Lizozomi. Mitohondrije. Plastidi
1. Koja je struktura i funkcije ATP?
2. Koje vrste plastida poznajete?
Kada različiti nutrijenti uđu u ćeliju putem fagocitoze ili pinocitoze, moraju se probaviti. Gde vjeverice moraju se razgraditi na pojedinačne aminokiseline, polisaharide - na molekule glukoze ili fruktoze, lipida- na glicerol i masne kiseline. Da bi intracelularna probava bila moguća, fagocitni ili pinocitni vezikuli moraju se stopiti sa lizozomom (slika 25). Lizozom je mali mjehur, prečnika samo 0,5-1,0 mikrona, koji sadrži veliki skup enzima koji mogu uništiti hranjive tvari. Jedan lizozom može sadržavati 30-50 različitih enzima.

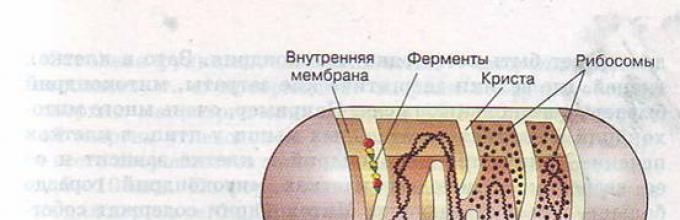
Mitohondrije su trajne membranske organele okruglog ili štapićastog (često granastog) oblika. Debljina - 0,5 mikrona, dužina - 5-7 mikrona. Broj mitohondrija u većini životinjskih ćelija je 150-1500; u ženskim jajnim ćelijama - do nekoliko stotina hiljada, u spermi - jedna spiralna mitohondrija uvijena oko aksijalnog dela flageluma.
Glavne funkcije mitohondrija:
1) igraju ulogu energetskih stanica ćelija;
2) skladištiti nasljedni materijal u obliku mitohondrijske DNK.
Nus funkcije - sudjelovanje u sintezi steroidnih hormona, nekih aminokiselina (na primjer, glutamina).
Struktura mitohondrija
Mitohondrije imaju dvije membrane: vanjsku (glatku) i unutrašnju (tvoreći izrasline - lisnate (kriste) i cjevaste (tubule)).
U mitohondrijama je unutarnji sadržaj matriks - koloidna tvar u kojoj su pomoću elektronskog mikroskopa otkrivena zrna promjera 20-30 nm (akumuliraju ione kalcija i magnezija, rezerve hranjivih tvari, na primjer, glikogen).
U matrici se nalazi aparat za biosintezu proteina organele: 2-6 kopija kružne DNK bez histonskih proteina, ribozomi, set t-RNA, enzimi za reduplikaciju, transkripciju i translaciju nasljednih informacija.
Mitohondrije se množe ligacijom; mitohondrije karakteriše relativna autonomija unutar ćelije.
Lizozomi su vezikule prečnika 200-400 µm. (obično). Imaju jednomembransku ljusku, koja je ponekad izvana prekrivena vlaknastim proteinskim slojem. Glavna funkcija je unutarćelijska probava različitih kemijskih spojeva i ćelijskih struktura.
Postoje primarni (neaktivni) i sekundarni lizozomi (u njima se odvija proces probave). Sekundarni lizozomi nastaju od primarnih. Dijele se na heterolizozome i autolizozome.
U heterolizosomima (ili fagolizosomima) proces varenja materijala koji ulazi u ćeliju izvana odvija se aktivnim transportom (pinocitoza i fagocitoza).
U autolizosomima (ili citolizosomima), njihove vlastite ćelijske strukture koje su završile svoj život su uništene.
Sekundarni lizosomi koji su prestali probaviti materijal nazivaju se rezidualna tijela. Ne sadrže hidrolaze i sadrže neprobavljeni materijal.
Kada je narušen integritet membrane lizosoma ili kada je ćelija bolesna, hidrolaze ulaze u ćeliju iz lizosoma i vrše njenu samoprobavu (autolizu). Isti proces je u osnovi procesa prirodne smrti svih ćelija (apoptoza).
Mikrotijela
Mikrotijela čine zajedničku grupu organela. To su vezikule promjera 100-150 nm, ograničene jednom membranom. Sadrže fino zrnastu matricu i često proteinske inkluzije.
Stalne ćelijske strukture, ćelijski organi koji obezbeđuju obavljanje specifičnih funkcija tokom života ćelije - skladištenje i prenos genetskih informacija, prenos supstanci, sinteza i transformacija supstanci i energije, deoba, kretanje itd.
Na organoide (organele) ćelija eukarioti vezati:
- hromozomi;
- stanične membrane;
- mitohondrije;
- Golgijev kompleks;
- endoplazmatski retikulum;
- ribosomi;
- mikrotubule;
- mikrofilamenti;
- lizozomi.
Životinjske stanice također sadrže centriole i mikrofibrile, a biljne ćelije sadrže plastide koji su im jedinstveni.
Ponekad se jezgro kao cjelina klasificira kao organele eukariotskih stanica.
Prokarioti nemaju većinu organela, imaju samo ćelijsku membranu i ribozome, koji se razlikuju od citoplazmatskih ribozoma eukariotskih ćelija.
Specijalizovane eukariotske ćelije mogu imati složene strukture zasnovane na univerzalnim organelama, kao što su mikrotubule i centriole - glavne komponente bičaka i cilija. Mikrofibrile su u osnovi tono- i neurofibrila. Posebne strukture jednoćelijskih organizama, kao što su flagele i cilije (izgrađene na isti način kao u višećelijskim ćelijama), obavljaju funkciju organa kretanja.
U modernoj literaturi češće se koriste termini „ organoidi " i " organele " se koriste kao sinonimi.
Strukture zajedničke životinjskim i biljnim ćelijama
| Shematska ilustracija | Struktura | Funkcije |
|
| Plazma membrana (plazmalema, ćelijska membrana) | Dva sloja lipida (dvosloj) između dva sloja proteina | Selektivno propusna barijera koja reguliše razmjenu između ćelije i okoline |
|
| Core | Najveća organela, zatvorena u školjku od dvije membrane, prožeta nuklearne pore. Sadrži hromatin- u ovom obliku, odmotani hromozomi su u interfazi. Također sadrži strukturu tzv nucleolus | Hromozomi sadrže DNK - supstancu naslijeđa.DNK se sastoji od gena koji regulišu sve vrste ćelijske aktivnosti. Nuklearna podjela je u osnovi reprodukcije stanica, a samim tim i procesa reprodukcije. Ribosomi se formiraju u nukleolu |
|
| Endoplazmatski retikulum (ER) | Sistem spljoštenih membranskih vreća - tenkovi- u obliku cijevi i ploča. Sa vanjskom membranom nuklearnog omotača čini jednu jedinicu | Ako je površina ER prekrivena ribosomima, onda se zove grubo.Protein sintetizovan na ribosomima transportuje se kroz rezervoare takvog ER. Glatko ER(bez ribozoma) služi kao mjesto za sintezu lipida i steroida |
|
| Ribosomi | Vrlo male organele koje se sastoje od dvije podčestice - velike i male. Sadrže proteine i RNK u približno jednakim omjerima. Ribosomi, koji se nalaze u mitohondrijima (kao i u hloroplastima u biljkama), još su manji | Mjesto sinteze proteina, gdje se različiti molekuli u interakciji drže u ispravnom položaju. Ribosomi su povezani sa ER ili leže slobodni u citoplazmi. Mogu se formirati mnogi ribozomi polizom (poliribozom), u kojem su nanizani na jednu lancu glasničke RNK |
|
| Mitohondrije | Mitohondrij je okružen ljuskom od dvije membrane, unutrašnja membrana formira nabore ( cristas). Sadrži matriks koji sadrži mali broj ribozoma, jednu kružnu DNK molekulu i granule fosfata | Tokom aerobnog disanja, oksidativna fosforilacija i prijenos elektrona se dešavaju u kristama, a enzimi uključeni u Krebsov ciklus i oksidaciju masnih kiselina djeluju u matriksu. |
|
| Golgijev aparat | Gomila spljoštenih membranskih vrećica - tenkovi. Na jednom kraju se kontinuirano formiraju hrpe vrećica, a na drugom su povezane u obliku mjehurića. Stopovi mogu postojati kao diskretni diktiosomi, kao u biljnim stanicama, ili formirati prostornu mrežu, kao u mnogim životinjskim stanicama | Mnogi ćelijski materijali, kao što su enzimi iz ER, prolaze kroz modifikaciju u cisternama i transportuju se u vezikulama. Golgijev aparat je uključen u proces sekrecije i u njemu se formiraju lizozomi |
|
| Lizozomi | Jednostavna sferna membranska vreća (jedna membrana) ispunjena probavnim (hidrolitičkim) enzimima. Sadržaj se čini homogenim | Obavlja mnoge funkcije, uvijek povezane s dezintegracijom bilo koje strukture ili molekula |
|
| Mikrotijela | Organela nije sasvim pravilnog sfernog oblika, okružena je jednom membranom. Sadržaj ima granularnu strukturu, ali ponekad u njemu postoji kristaloid ili skup niti | Sva mikrotijela sadrže katalazu, enzim koji katalizuje razgradnju vodikovog peroksida. Svi su povezani s oksidativnim reakcijama |
|
| Ćelijski zid, srednja lamina, plazmodesma |
|||
| ćelijski zid | Kruti ćelijski zid koji okružuje ćeliju sastoji se od celuloznih mikrofibrila ugrađenih u matriks koji sadrži druge složene polisaharide, naime hemiceluloze i pektine. U nekim ćelijama, ćelijski zidovi prolaze kroz sekundarno zadebljanje | Pruža mehaničku podršku i zaštitu. Zahvaljujući njemu nastaje turgorski pritisak, što pojačava funkciju potpore. Sprečava rupturu osmotskih ćelija. Kretanje vode i mineralnih soli odvija se duž ćelijskog zida. Različite modifikacije, kao što je impregnacija ligninom, pružaju specijalizirane funkcije |
|
| srednja ploča | Tanak sloj pektinskih supstanci (pektati kalcija i magnezija) | Drži ćelije zajedno |
|
| plazmodesma | Tanak citoplazmatski filament koji povezuje citoplazmu dvije susjedne ćelije kroz tanku poru u ćelijskom zidu. Pora je obložena plazma membranom.Dezmotubul prolazi kroz poru, često povezan na oba kraja sa ER | Ujedinite protoplaste susjednih ćelija u jedan kontinuirani sistem - simplast, kroz koje se supstance transportuju između ovih ćelija |
|
| Hloroplast | Veliki plastid koji sadrži hlorofil u kojem se odvija fotosinteza. Hloroplast je okružen dvostrukom membranom i ispunjen želatinozom stroma. Stroma sadrži sistem membrana sastavljenih u hrpe, ili zrna. U njega se takođe može deponovati skrob. Osim toga, stroma sadrži ribozome, kružni DNK molekul i kapljice ulja | U ovoj organeli se dešava fotosinteza, odnosno sinteza šećera i drugih supstanci iz CO 2 i vode zahvaljujući svetlosnoj energiji koju hvata hlorofil. Svetlosna energija se pretvara u hemijsku energiju |
|
| Velika centralna vakuola | Vrećica formirana od jedne membrane tzv tonoplast. Vakuola sadrži ćelijski sok – koncentriranu otopinu različitih supstanci, kao što su mineralne soli, šećeri, pigmenti, organske kiseline i enzimi. U zrelim ćelijama vakuole su obično velike | Ovdje se pohranjuju razne tvari, uključujući i krajnje produkte metabolizma. Osmotska svojstva ćelije jako zavise od sadržaja vakuole. Ponekad vakuola funkcionira kao lizozom |
|
Komparativne karakteristike RNK i DNK
| Znakovi | RNA | DNK |
| Lokacija u kavezu | Nukleus, ribozomi, citoplazma, mitohondrije, hloroplasti | Nukleus, mitohondrije, hloroplasti |
| Lokacija u jezgru | Nucleolus | hromozomi |
| Struktura makromolekula | Jednostruki polinukleotidni lanac | Dvostruki nerazgranati linearni polimer, umotan u desnu spiralu |
| Monomeri | Ribonukleotidi | Deoksiribonukleotidi |
| Nukleotidni sastav | Azotne baze (purin - adenin, gvanin, pirimidin - uracil, citozin); riboza (ugljikohidrat): ostatak fosforne kiseline | Azotne baze (purin - adenin, gvanin, pirimidin - timin, citozin); deoksiriboza (ugljikohidrat): ostatak fosforne kiseline |
| Vrste nukleotida | alenil (A), guanil (G), uridil (U), citidil (C) | alenil (A), gvanil (G), timidil (T), citidil (C) |
| Svojstva | Nesposoban za samoudvostručavanje. Labilna | Sposoban za samoumnožavanje po principu komplementarnosti (reduplikacija): A-T, T-A, G-C, C-G Stabilan |
| Funkcije | Informativni (mRNA) - prenosi kod nasljedne informacije o primarnoj strukturi proteinskog molekula; ribosomalna (rRNA) - dio ribozoma; transport (tRNA) - prenosi aminokiseline do ribozoma; mitohondrijalna i plastidna RNK - dio su ribozoma ovih organela | Hemijska osnova hromozomskog genetskog materijala (gena); Sinteza DNK, sinteza RNK, informacije o strukturi proteina |
Mitohondrije su mikroskopske organele vezane za membranu koje opskrbljuju ćeliju energijom. Stoga se nazivaju energetskim stanicama (baterijama) ćelija.
Mitohondrije su odsutne u ćelijama jednostavnih organizama, bakterija i entameba, koje žive bez upotrebe kiseonika. Neke zelene alge tripanosomi sadrže jedan veliki mitohondrij, a ćelije srčanog mišića i mozga imaju od 100 do 1000 ovih organela.
Strukturne karakteristike
Mitohondrije su dvomembranske organele; imaju vanjsku i unutarnju membranu, međumembranski prostor između njih i matriks.
Vanjska membrana. Glatka je, nema nabora i odvaja unutrašnji sadržaj od citoplazme. Njegova širina je 7 nm i sadrži lipide i proteine. Važnu ulogu igra porin, protein koji formira kanale u vanjskoj membrani. Oni obezbjeđuju jonsku i molekularnu izmjenu.
Intermembranski prostor. Veličina intermembranskog prostora je oko 20 nm. Supstanca koja ga ispunjava po sastavu je slična citoplazmi, s izuzetkom velikih molekula koji ovdje mogu prodrijeti samo aktivnim transportom.

Unutrašnja membrana. Građen je uglavnom od proteina, samo trećina se izdvaja za lipidne supstance. Veliki broj proteina su transportni proteini, jer unutrašnja membrana nema slobodno prohodne pore. Formira mnoge izrasline - kriste, koje izgledaju kao spljošteni grebeni. Oksidacija organskih jedinjenja do CO 2 u mitohondrijima se dešava na membranama krista. Ovaj proces ovisi o kisiku i odvija se pod djelovanjem ATP sintetaze. Oslobođena energija se pohranjuje u obliku ATP molekula i koristi se po potrebi.
Matrix– unutrašnje okruženje mitohondrija ima granularnu, homogenu strukturu. U elektronskom mikroskopu možete vidjeti granule i filamente u kuglicama koje slobodno leže između krista. Matrica sadrži poluautonomni sistem sinteze proteina - DNK, sve vrste RNK i ribozomi se nalaze ovdje. Ipak, većina proteina se opskrbljuje iz jezgre, zbog čega se mitohondrije nazivaju poluautonomne organele.
Lokacija i podjela ćelije
Hondriom je grupa mitohondrija koja je koncentrisana u jednoj ćeliji. Različito se nalaze u citoplazmi, što zavisi od specijalizacije ćelija. Položaj u citoplazmi zavisi i od okolnih organela i inkluzija. U biljnim ćelijama oni zauzimaju periferiju, jer su mitohondrije gurnute prema membrani centralnom vakuolom. U bubrežnim epitelnim stanicama membrana formira izbočine između kojih se nalaze mitohondriji.
U matičnim ćelijama, gde se energija podjednako koristi za sve organele, mitohondrije su raspoređene haotično. U specijalizovanim ćelijama oni su uglavnom koncentrisani u područjima najveće potrošnje energije. Na primjer, u poprečnoprugastim mišićima nalaze se u blizini miofibrila. Kod spermatozoida spiralno prekrivaju os flageluma, jer je potrebno mnogo energije da se ona pokrene i pokrene spermatozoid. Protozoji koji se kreću pomoću cilija također sadrže veliki broj mitohondrija u svojoj osnovi.
Division. Mitohondrije su sposobne za samostalnu reprodukciju, imaju vlastiti genom. Organele su podijeljene konstrikcijama ili septama. Formiranje novih mitohondrija u različitim stanicama razlikuje se po učestalosti; na primjer, u tkivu jetre one se zamjenjuju svakih 10 dana.
Funkcije u ćeliji
- Glavna funkcija mitohondrija je formiranje ATP molekula.
- Taloženje jona kalcijuma.
- Učešće u razmjeni vode.
- Sinteza prekursora steroidnih hormona.
Molekularna biologija je nauka koja proučava ulogu mitohondrija u metabolizmu. Oni također pretvaraju piruvat u acetil-koenzim A i beta-oksidaciju masnih kiselina.
| Tabela: struktura i funkcije mitohondrija (ukratko) | ||
|---|---|---|
| Strukturni elementi | Struktura | Funkcije |
| Vanjska membrana | Glatka ljuska, napravljena od lipida i proteina | Odvaja unutrašnji sadržaj od citoplazme |
| Intermembranski prostor | Postoje joni vodonika, proteini, mikromolekule | Stvara protonski gradijent |
| Unutrašnja membrana | Formira izbočine - kriste, sadrži sisteme za transport proteina | Prijenos makromolekula, održavanje gradijenta protona |
| Matrix | Lokacija enzima Krebsovog ciklusa, DNK, RNK, ribozoma | Aerobna oksidacija sa oslobađanjem energije, pretvaranje piruvata u acetil koenzim A. |
| Ribosomi | Kombinovane dve podjedinice | Sinteza proteina |
Sličnosti između mitohondrija i hloroplasta
Zajednička svojstva mitohondrija i hloroplasta prvenstveno su posljedica prisutnosti dvostruke membrane.
Znakovi sličnosti uključuju i sposobnost neovisne sinteze proteina. Ove organele imaju svoju DNK, RNK i ribozome.
I mitohondrije i hloroplasti se mogu podijeliti sužavanjem.
Ujedinjuje ih i sposobnost proizvodnje energije; mitohondrije su specijalizovanije za ovu funkciju, ali hloroplasti takođe proizvode molekule ATP-a tokom fotosintetskih procesa. Dakle, biljne stanice imaju manje mitohondrija od životinjskih, jer hloroplasti za njih djelomično obavljaju funkcije.
Hajde da ukratko opišemo sličnosti i razlike:
- One su dvomembranske organele;
- unutrašnja membrana formira izbočine: kriste su karakteristične za mitohondrije, a tilakoidi su karakteristični za hloroplaste;
- imaju svoj genom;
- sposoban da sintetiše proteine i energiju.
Ove organele se razlikuju po svojim funkcijama: mitohondrije su namijenjene za sintezu energije, ovdje se javlja ćelijsko disanje, kloroplasti su potrebni biljnim stanicama za fotosintezu.
Sadržaj članka
ĆELIJA, elementarne jedinice živih bića. Ćelija je odvojena od drugih ćelija ili od vanjskog okruženja posebnom membranom i ima jezgro ili njegov ekvivalent, u kojem je koncentriran najveći dio kemijskih informacija koje kontroliraju nasljeđe. Citologija proučava strukturu ćelija, a fiziologija se bavi njihovim funkcionisanjem. Nauka koja proučava tkivo koje se sastoji od ćelija naziva se histologija.
Postoje jednoćelijski organizmi čije se cijelo tijelo sastoji od jedne ćelije. Ova grupa uključuje bakterije i protiste (protozoe i jednoćelijske alge). Ponekad se nazivaju i bezćelijski, ali se češće koristi termin jednoćelijski. Prave višećelijske životinje (Metazoa) i biljke (Metaphyta) sadrže mnogo ćelija.
Velika većina tkiva sastoji se od ćelija, ali postoje izuzeci. Tijelo sluzavih plijesni (miksomiceta), na primjer, sastoji se od homogene tvari koja nije podijeljena na ćelije s brojnim jezgrama. Neka životinjska tkiva, posebno srčani mišić, organizirana su na sličan način. Vegetativno tijelo (talus) gljiva formirano je od mikroskopskih niti - hifa, često segmentiranih; svaka takva nit se može smatrati ekvivalentom ćelije, iako netipičnog oblika.
Neke strukture tijela koje ne sudjeluju u metabolizmu, posebno školjke, biseri ili mineralna osnova kostiju, ne formiraju ćelije, već proizvodi njihovog lučenja. Drugi, kao što su drvo, kora, rogovi, kosa i vanjski sloj kože, nisu sekretornog porijekla, već se formiraju od mrtvih ćelija.
Mali organizmi, kao što su rotiferi, sastoje se od svega nekoliko stotina ćelija. Poređenja radi: u ljudskom tijelu ima cca. 10 14 ćelija, svake sekunde 3 miliona crvenih krvnih zrnaca umire i zamenjuju se novim, a to je samo jedan desetmilioni deo ukupnog broja telesnih ćelija.
Obično se veličine biljnih i životinjskih ćelija kreću od 5 do 20 mikrona u prečniku. Tipična bakterijska ćelija je mnogo manja – cca. 2 mikrona, a najmanji poznati je 0,2 mikrona.
Neke slobodno živeće ćelije, kao što su protozoe kao što je foraminifera, mogu doseći nekoliko centimetara; uvijek imaju mnogo jezgara. Ćelije tankih biljnih vlakana dostižu dužinu od jednog metra, a procesi nervnih ćelija kod velikih životinja dosežu i nekoliko metara. Sa takvom dužinom, volumen ovih ćelija je mali, ali je površina vrlo velika.
Najveće ćelije su neoplođena ptičja jaja napunjena žumanjkom. Najveće jaje (a samim tim i najveća ćelija) pripadalo je izumrloj ogromnoj ptici - apyornisu ( Aepyornis). Pretpostavlja se da je žumanca bilo teško cca. 3,5 kg. Najveće jaje među živim vrstama pripada noju čije je žumance teško cca. 0,5 kg.
U pravilu, stanice velikih životinja i biljaka su tek nešto veće od stanica malih organizama. Slon je veći od miša ne zato što su njegove ćelije veće, već uglavnom zato što samih ćelija ima mnogo više. Postoje grupe životinja, poput rotifera i nematoda, kod kojih broj ćelija u tijelu ostaje konstantan. Dakle, iako velike vrste nematoda imaju veći broj ćelija od malih, glavna razlika u veličini je u ovom slučaju posljedica većih veličina ćelija.
Unutar datog tipa ćelije, njihove veličine obično zavise od ploidnosti, tj. na broj skupova hromozoma prisutnih u jezgru. Tetraploidne ćelije (sa četiri seta hromozoma) su duplo veće po zapremini od diploidne ćelije (sa dva seta hromozoma). Ploidnost biljke može se povećati unošenjem biljnog lijeka kolhicina u nju. Budući da biljke izložene ovom dejstvu imaju veće ćelije, one su same veće. Međutim, ovaj fenomen se može uočiti samo kod poliploida novijeg porijekla. U evolucijski drevnim poliploidnim biljkama, veličine ćelija su podložne „obrnutoj regulaciji“ prema normalnim vrijednostima unatoč povećanju broja kromosoma.
STRUKTURA ĆELIJE
Nekada se ćelija smatrala manje ili više homogenom kapljicom organske materije, koja se nazivala protoplazma ili živa supstanca. Ovaj termin je zastario nakon što je otkriveno da se ćelija sastoji od mnogih jasno različitih struktura koje se nazivaju ćelijske organele („mali organi“).
Hemijski sastav.
Tipično, 70-80% ćelijske mase čini voda, u kojoj su otopljene različite soli i niskomolekularna organska jedinjenja. Najkarakterističnije komponente ćelije su proteini i nukleinske kiseline. Neki proteini su strukturne komponente ćelije, drugi su enzimi, tj. katalizatori koji određuju brzinu i smjer kemijskih reakcija koje se odvijaju u stanicama. Nukleinske kiseline služe kao nosioci nasljedne informacije, koja se ostvaruje u procesu intracelularne sinteze proteina.
Često ćelije sadrže određenu količinu skladišnih supstanci koje služe kao rezerva hrane. Biljne ćelije prvenstveno skladište škrob, polimerni oblik ugljikohidrata. Drugi ugljikohidratni polimer, glikogen, pohranjuje se u jetri i mišićnim stanicama. Često pohranjene namirnice također uključuju masnoće, iako neke masti imaju drugu funkciju, odnosno služe kao bitne strukturne komponente. Proteini u ćelijama (sa izuzetkom ćelija sjemena) obično se ne skladište.
Nije moguće opisati tipičan sastav ćelije, prvenstveno zato što postoje velike razlike u količini uskladištene hrane i vode. Ćelije jetre sadrže, na primjer, 70% vode, 17% proteina, 5% masti, 2% ugljikohidrata i 0,1% nukleinskih kiselina; preostalih 6% dolazi od soli i organskih spojeva male molekularne težine, posebno aminokiselina. Biljne ćelije obično sadrže manje proteina, znatno više ugljikohidrata i nešto više vode; izuzetak su ćelije koje su u stanju mirovanja. Ćelija pšeničnog zrna, koja je izvor hranljivih materija za embrion, sadrži cca. 12% proteina (uglavnom uskladištenih proteina), 2% masti i 72% ugljenih hidrata. Količina vode dostiže normalan nivo (70–80%) tek na početku klijanja zrna.
Glavni dijelovi ćelije.
Neke ćelije, uglavnom biljne i bakterijske, imaju vanjski ćelijski zid. U višim biljkama sastoji se od celuloze. Zid okružuje samu ćeliju, štiteći je od mehaničkog stresa. Ćelije, posebno bakterijske, također mogu lučiti sluzave tvari, formirajući tako oko sebe kapsulu koja, kao i ćelijski zid, ima zaštitnu funkciju.
Upravo s uništavanjem staničnih zidova povezana je smrt mnogih bakterija pod utjecajem penicilina. Činjenica je da je unutar bakterijske stanice koncentracija soli i niskomolekularnih spojeva vrlo visoka, pa stoga, u nedostatku armaturnog zida, dotok vode u ćeliju uzrokovan osmotskim tlakom može dovesti do njenog pucanja. Penicilin, koji sprečava stvaranje njegovog zida tokom rasta ćelije, dovodi do rupture ćelije (lize).
Ćelijski zidovi i kapsule ne učestvuju u metabolizmu i često se mogu odvojiti bez ubijanja ćelije. Stoga se mogu smatrati vanjskim pomoćnim dijelovima ćelije. Životinjskim ćelijama obično nedostaju ćelijski zidovi i kapsule.
Sama ćelija se sastoji od tri glavna dijela. Ispod ćelijskog zida, ako postoji, nalazi se ćelijska membrana. Membrana okružuje heterogeni materijal koji se naziva citoplazma. Okruglo ili ovalno jezgro je uronjeno u citoplazmu. U nastavku ćemo detaljnije pogledati strukturu i funkcije ovih dijelova ćelije.
STANIČNE MEMBRANE
Ćelijska membrana je veoma važan dio ćelije. On drži sve ćelijske komponente zajedno i razgraničava unutrašnje i spoljašnje okruženje. Osim toga, modificirani nabori ćelijske membrane formiraju mnoge ćelijske organele.
Stanična membrana je dvostruki sloj molekula (bimolekularni sloj ili dvosloj). To su uglavnom molekule fosfolipida i drugih srodnih supstanci. Molekuli lipida imaju dvostruku prirodu, koja se očituje u tome kako se ponašaju u odnosu na vodu. Glave molekula su hidrofilne, tj. imaju afinitet prema vodi, a njihovi ugljikovodični repovi su hidrofobni. Stoga, kada se pomiješaju s vodom, lipidi stvaraju film na svojoj površini sličan uljnom filmu; Štaviše, svi njihovi molekuli su orijentisani na isti način: glave molekula su u vodi, a ugljikovodični repovi iznad njene površine.
U ćelijskoj membrani postoje dva takva sloja, i u svakom od njih su glave molekula okrenute prema van, a repovi okrenuti unutar membrane, jedan prema drugom, tako da ne dolaze u dodir s vodom. Debljina takve membrane je cca. 7 nm. Pored glavnih lipidnih komponenti, sadrži velike proteinske molekule koje su u stanju da „lebde” u lipidnom dvosloju i raspoređene su tako da je jedna strana okrenuta ka unutrašnjosti ćelije, a druga u kontaktu sa spoljašnjim okruženjem. Neki proteini se nalaze samo na vanjskoj ili samo na unutarnjoj površini membrane ili su samo djelomično uronjeni u lipidni dvosloj.
Glavna funkcija stanične membrane je regulacija transporta tvari u ćeliju i iz nje. Budući da je membrana fizički donekle slična ulju, tvari koje su topljive u ulju ili organskim otapalima, kao što je eter, lako prolaze kroz nju. Isto vrijedi i za plinove kao što su kisik i ugljični dioksid. U isto vrijeme, membrana je praktički nepropusna za većinu tvari topljivih u vodi, posebno šećera i soli. Zahvaljujući ovim svojstvima, u stanju je da održava hemijsko okruženje unutar ćelije koje se razlikuje od spoljašnjeg. Na primjer, u krvi je visoka koncentracija jona natrijuma, a niska iona kalija, dok su u unutarćelijskoj tekućini ti joni prisutni u suprotnom omjeru. Slična situacija je tipična za mnoga druga hemijska jedinjenja.
Očigledno je da stanica, međutim, ne može biti potpuno izolirana od okoline, jer mora primiti tvari potrebne za metabolizam i osloboditi se svojih konačnih proizvoda. Osim toga, lipidni dvosloj nije potpuno nepropustan čak ni za tvari topive u vodi, te za one tzv. koje prodiru u njega. Proteini koji "formiraju kanale" stvaraju pore ili kanale koji se mogu otvarati i zatvarati (u zavisnosti od promjena u konformaciji proteina) i, u otvorenom stanju, provode određene ione (Na +, K +, Ca 2+) duž gradijenta koncentracije . Posljedično, razlika u koncentracijama unutar i izvan ćelije ne može se održati samo zbog niske permeabilnosti membrane. U stvari, sadrži proteine koji obavljaju funkciju molekularne "pumpe": prenose određene tvari u ćeliju i van nje, radeći protiv gradijenta koncentracije. Kao rezultat toga, kada je koncentracija, na primjer, aminokiselina u ćeliji visoka i niska izvana, aminokiseline ipak mogu teći iz vanjskog okruženja u onu unutarnju. Ovaj prijenos se naziva aktivni transport i koristi energiju dobivenu metabolizmom. Membranske pumpe su vrlo specifične: svaka od njih može transportirati ili samo jone određenog metala, ili aminokiselinu, ili šećer. Membranski jonski kanali su takođe specifični.
Takva selektivna permeabilnost je fiziološki veoma važna, a njeno odsustvo je prvi dokaz ćelijske smrti. To je lako ilustrirati na primjeru cvekle. Ako se živi korijen repe potopi u hladnu vodu, zadržava svoj pigment; ako se cvekla prokuha, ćelije umiru, postaju lako propusne i gube pigment, što vodu postaje crveno.
Ćelija može "progutati" velike molekule kao što su proteini. Pod uticajem određenih proteina, ako su prisutni u tečnosti koja okružuje ćeliju, dolazi do invaginacije u ćelijskoj membrani, koja se zatim zatvara, formirajući vezikulu – malu vakuolu koja sadrži vodu i proteinske molekule; Nakon toga, membrana oko vakuole puca, a sadržaj ulazi u ćeliju. Ovaj proces se naziva pinocitoza (bukvalno „ispijanje ćelije“) ili endocitoza.
Veće čestice, poput čestica hrane, mogu se apsorbirati na sličan način tokom tzv. fagocitoza. Obično je vakuola formirana tokom fagocitoze veća, a hrana se probavlja lizozomskim enzimima unutar vakuole prije nego što okolna membrana pukne. Ova vrsta ishrane tipična je za protozoe, kao što su amebe, koje jedu bakterije. Međutim, sposobnost fagocitoze je karakteristična i za crijevne stanice nižih životinja i za fagocite, jednu od vrsta bijelih krvnih stanica (leukocita) kralježnjaka. U potonjem slučaju smisao ovog procesa nije u ishrani samih fagocita, već u njihovom uništavanju bakterija, virusa i drugog stranog materijala štetnog za tijelo.
Funkcije vakuola mogu biti različite. Na primjer, protozoe koje žive u slatkoj vodi doživljavaju stalan osmotski priliv vode, jer je koncentracija soli unutar ćelije mnogo veća nego izvan nje. Oni su u stanju da luče vodu u posebnu ekskretornu (kontraktilnu) vakuolu, koja povremeno istiskuje svoj sadržaj.
Biljne ćelije često imaju jednu veliku centralnu vakuolu koja zauzima gotovo cijelu ćeliju; citoplazma formira samo vrlo tanak sloj između ćelijskog zida i vakuole. Jedna od funkcija takve vakuole je akumulacija vode, omogućavajući ćeliji da se brzo poveća u veličini. Ova sposobnost je posebno neophodna u periodu kada biljna tkiva rastu i formiraju vlaknaste strukture.
U tkivima, na mjestima gdje su ćelije čvrsto povezane, njihove membrane sadrže brojne pore formirane od proteina koji prodiru kroz membranu - tzv. konektori. Pore susjednih ćelija nalaze se jedna naspram druge, tako da niskomolekularne supstance mogu prelaziti od ćelije do ćelije - ovaj hemijski komunikacioni sistem koordinira njihovu vitalnu aktivnost. Jedan primjer takve koordinacije je manje-više sinhrona podjela susjednih stanica uočena u mnogim tkivima.
CITOPLAZMA
Citoplazma sadrži unutrašnje membrane koje su slične vanjskoj membrani i formiraju organele različitih tipova. Ove membrane se mogu smatrati naborima vanjske membrane; ponekad su unutrašnje membrane integralne sa vanjskom, ali često se unutrašnji nabor razvezuje i kontakt sa vanjskom membranom je prekinut. Međutim, čak i ako se kontakt održava, unutrašnja i vanjska membrana nisu uvijek kemijski identične. Konkretno, sastav membranskih proteina se razlikuje u različitim ćelijskim organelama.
Endoplazmatski retikulum.
Mreža unutrašnjih membrana koja se sastoji od tubula i vezikula proteže se od površine ćelije do jezgra. Ova mreža se naziva endoplazmatski retikulum. Često se primjećuje da se tubuli otvaraju na površini ćelije, te stoga endoplazmatski retikulum igra ulogu mikrocirkulacijskog aparata preko kojeg vanjska sredina može direktno komunicirati sa cjelokupnim sadržajem ćelije. Ova interakcija je pronađena u nekim ćelijama, posebno mišićnim ćelijama, ali još nije jasno da li je univerzalna. U svakom slučaju, transport većeg broja supstanci kroz ove tubule iz jednog dela ćelije u drugi se zapravo dešava.
Sitna tijela nazvana ribosomi pokrivaju površinu endoplazmatskog retikuluma, posebno blizu jezgra. Prečnik ribosoma cca. 15 nm, sastoje se od pola proteina, pola ribonukleinskih kiselina. Njihova glavna funkcija je sinteza proteina; glasnička RNK i aminokiseline povezane s prijenosnom RNK su vezane za njihovu površinu. Područja retikuluma prekrivena ribozomima nazivaju se hrapavi endoplazmatski retikulum, a oni bez njih se nazivaju glatkim. Osim ribozoma, različiti enzimi se adsorbiraju na endoplazmatskom retikulumu ili na neki drugi način vezani za njega, uključujući enzimske sisteme koji obezbjeđuju korištenje kisika za stvaranje sterola i za neutralizaciju određenih otrova. U nepovoljnim uslovima endoplazmatski retikulum brzo degeneriše, pa njegovo stanje služi kao osetljiv pokazatelj zdravlja ćelija.
Golgijev aparat.
Golgijev aparat (Golgijev kompleks) je specijalizirani dio endoplazmatskog retikuluma koji se sastoji od naslaganih ravnih membranskih vrećica. Učestvuje u sekreciji proteina od strane ćelije (u njoj dolazi do pakovanja izlučenih proteina u granule) i stoga je posebno razvijen u ćelijama koje obavljaju sekretornu funkciju. Važne funkcije Golgijevog aparata također uključuju vezivanje grupa ugljikohidrata za proteine i korištenje ovih proteina za izgradnju stanične membrane i membrane lizosoma. U nekim algama, celulozna vlakna se sintetiziraju u Golgijevom aparatu.
Lizozomi
- To su mali mehurići okruženi jednom membranom. Pupaju iz Golgijevog aparata i možda iz endoplazmatskog retikuluma. Lizozomi sadrže razne enzime koji razgrađuju velike molekule, posebno proteine. Zbog svog destruktivnog djelovanja, ovi enzimi su takoreći "zaključani" u lizozomima i oslobađaju se samo kada je to potrebno. Dakle, tokom intracelularne probave, enzimi se oslobađaju iz lizosoma u probavne vakuole. Lizozomi su takođe neophodni za uništavanje ćelija; na primjer, tijekom transformacije punoglavca u odraslu žabu, oslobađanje lizosomalnih enzima osigurava uništavanje repnih stanica. U ovom slučaju, to je normalno i korisno za tijelo, ali ponekad je takvo uništavanje stanica patološko. Na primjer, kada se azbestna prašina udiše, ona može prodrijeti u ćelije pluća, a zatim pucaju lizozomi, uništavanje stanica i razvija se plućna bolest.
Mitohondrije i hloroplasti.
Mitohondrije su relativno velike strukture nalik vrećama s prilično složenom strukturom. Sastoje se od matriksa okruženog unutrašnjom membranom, međumembranskog prostora i vanjske membrane. Unutrašnja membrana je presavijena u nabore zvane kriste. Klasteri proteina nalaze se na kristama. Mnogi od njih su enzimi koji kataliziraju oksidaciju proizvoda razgradnje ugljikohidrata; drugi kataliziraju reakcije sinteze i oksidacije masti. Pomoćni enzimi uključeni u ove procese rastvaraju se u mitohondrijskom matriksu.
Oksidacija organskih supstanci se javlja u mitohondrijima, zajedno sa sintezom adenozin trifosfata (ATP). Razgradnja ATP-a u adenozin difosfat (ADP) je praćena oslobađanjem energije koja se troši na različite vitalne procese, na primjer, na sintezu proteina i nukleinskih kiselina, transport tvari u i iz stanice, prijenos nervnih impulsa ili kontrakcije mišića. Mitohondrije su stoga energetske stanice koje prerađuju “gorivo” – masti i ugljikohidrate – u oblik energije koji može koristiti stanica, a time i tijelo u cjelini.
Biljne ćelije takođe sadrže mitohondrije, ali glavni izvor energije za njihove ćelije je svetlost. Ove stanice koriste svjetlosnu energiju za proizvodnju ATP-a i sintezu ugljikohidrata iz ugljičnog dioksida i vode. Hlorofil, pigment koji akumulira svjetlosnu energiju, nalazi se u hloroplastima. Kloroplasti, poput mitohondrija, imaju unutrašnju i vanjsku membranu. Iz izraslina unutrašnje membrane tokom razvoja hloroplasta nastaju takozvani hloroplasti. tilakoidne membrane; potonji formiraju spljoštene vreće, skupljene u hrpe poput stupca novčića; ove hrpe, zvane grana, sadrže hlorofil. Osim hlorofila, hloroplasti sadrže i sve ostale komponente neophodne za fotosintezu.
Neki specijalizovani hloroplasti ne provode fotosintezu, ali imaju druge funkcije, poput skladištenja škroba ili pigmenata.
Relativna autonomija.
U nekim aspektima, mitohondrije i hloroplasti se ponašaju kao autonomni organizmi. Na primjer, baš kao i same ćelije, koje nastaju samo iz stanica, mitohondrije i hloroplasti nastaju samo od već postojećih mitohondrija i hloroplasta. To je pokazano u eksperimentima na biljnim stanicama, u kojima je stvaranje hloroplasta potisnuto antibiotikom streptomicinom, i na stanicama kvasca, gdje je stvaranje mitohondrija potisnuto drugim lijekovima. Nakon ovakvih efekata, ćelije nikada nisu obnovile nedostajuće organele. Razlog je taj što mitohondrije i hloroplasti sadrže određenu količinu vlastitog genetskog materijala (DNK) koji kodira dio njihove strukture. Ako se ova DNK izgubi, što se dešava kada je formiranje organela potisnuto, struktura se ne može ponovo stvoriti. Obje vrste organela imaju svoj vlastiti sistem za sintezu proteina (ribozomi i prijenosne RNK), koji se donekle razlikuje od glavnog sistema ćelije za sintezu proteina; poznato je, na primjer, da se sistem organela koji sintetiše proteine može potisnuti uz pomoć antibiotika, dok oni nemaju nikakvog efekta na glavni sistem.
Organela DNK je odgovorna za većinu ekstrahromozomskog ili citoplazmatskog nasljeđivanja. Ekstrahromozomsko naslijeđe ne pokorava se Mendelovim zakonima, jer kada se stanica podijeli, DNK organela se prenosi na ćelije kćeri na drugačiji način od hromozoma. Proučavanje mutacija koje se javljaju u DNK organela i hromozomskoj DNK pokazalo je da je DNK organela odgovorna samo za mali dio strukture organela; većina njihovih proteina je kodirana u genima smještenim na hromozomima.
Djelomična genetska autonomija organela koji se razmatraju i karakteristike njihovih sistema za sintezu proteina poslužili su kao osnova za pretpostavku da mitohondrije i hloroplasti potječu od simbiotskih bakterija koje su se naselile u stanicama prije 1-2 milijarde godina. Moderan primjer takve simbioze su male fotosintetske alge koje žive u ćelijama nekih koralja i mekušaca. Alge obezbeđuju kiseonik svojim domaćinima i od njih primaju hranljive materije.
Fibrilarne strukture.
Citoplazma ćelije je viskozna tekućina, tako da bi površinska napetost uzrokovala da se očekuje da će stanica biti sferna osim ako ćelije nisu čvrsto zbijene. Međutim, to se obično ne primjećuje. Mnoge protozoe imaju guste integumente ili membrane koje daju ćeliji specifičan, nesferičan oblik. Ipak, čak i bez membrane, stanice mogu zadržati nesferični oblik zbog činjenice da je citoplazma strukturirana uz pomoć brojnih, prilično krutih, paralelnih vlakana. Potonje su formirane šupljim mikrotubulama, koje se sastoje od proteinskih jedinica organiziranih u spiralu.
Neke protozoe formiraju pseudopodije - duge, tanke citoplazmatske projekcije kojima hvataju hranu. Pseudopodije zadržavaju svoj oblik zbog krutosti mikrotubula. Ako se hidrostatički pritisak poveća na približno 100 atmosfera, mikrotubule se raspadaju i ćelija poprima oblik kapi. Kada se pritisak vrati u normalu, mikrotubule se ponovo sastavljaju i ćelija formira pseudopodije. Mnoge druge ćelije na sličan način reaguju na promene pritiska, što potvrđuje učešće mikrotubula u održavanju oblika ćelije. Sastavljanje i dezintegracija mikrotubula, neophodnih da bi ćelija brzo promenila oblik, dešava se čak i u odsustvu promena pritiska.
Mikrotubule također formiraju fibrilarne strukture koje služe kao organi kretanja stanica. Neke ćelije imaju izbočine nalik na bičeve koje se nazivaju flagele, ili cilije - njihovo udaranje osigurava kretanje ćelije u vodi. Ako je ćelija nepomična, ove strukture guraju vodu, čestice hrane i druge čestice prema ili od ćelije. Flagele su relativno velike, i obično ćelija ima samo jednu, a ponekad i nekoliko flagela. Cilije su mnogo manje i pokrivaju cijelu površinu ćelije. Iako su ove strukture karakteristične uglavnom za najjednostavnije, one mogu biti prisutne i u visoko organiziranim oblicima. U ljudskom tijelu svi respiratorni putevi su obloženi cilijama. Male čestice koje uđu u njih obično se hvata sluz na površini ćelije, a cilije ih istiskuju zajedno sa sluzom napolje, štiteći tako pluća. Muške reproduktivne stanice većine životinja i nekih nižih biljaka kreću se uz pomoć bičaka.
Postoje i druge vrste ćelijskog kretanja. Jedan od njih je ameboidno kretanje. Amebe, kao i neke ćelije višećelijskih organizama, „teče“ od mesta do mesta, tj. pomjerati zbog struje sadržaja ćelije. Stalna struja materije postoji i unutar biljnih ćelija, ali ona ne podrazumeva kretanje ćelije kao celine. Najproučavaniji tip ćelijskog kretanja je kontrakcija mišićnih ćelija; odvija se klizećim fibrilima (proteinskim nitima) jedna u odnosu na drugu, što dovodi do skraćivanja ćelije.
CORE
Jezgro je okruženo dvostrukom membranom. Vrlo uzak (oko 40 nm) prostor između dvije membrane naziva se perinuklearni. Nuklearne membrane prelaze u membrane endoplazmatskog retikuluma, a perinuklearni prostor se otvara u retikularni prostor. Obično nuklearna membrana ima vrlo uske pore. Očigledno, veliki molekuli se transportuju kroz njih, kao što je glasnička RNK, koja se sintetizira na DNK, a zatim ulazi u citoplazmu.
Najveći dio genetskog materijala nalazi se u hromozomima ćelijskog jezgra. Hromozomi se sastoje od dugih lanaca dvolančane DNK, za koju su vezani osnovni (tj. alkalni) proteini. Ponekad hromozomi imaju nekoliko identičnih DNK lanaca koji leže jedan pored drugog - takvi se hromozomi nazivaju politeni (višelančani). Broj hromozoma varira među vrstama. Diploidne ćelije ljudskog tela sadrže 46 hromozoma, odnosno 23 para.
U ćeliji koja se ne dijeli, hromozomi su pričvršćeni na jednoj ili više tačaka za nuklearnu membranu. U svom normalnom neumotanom stanju, hromozomi su toliko tanki da se ne vide pod svetlosnim mikroskopom. Na određenim lokusima (presjecima) jednog ili više hromozoma formira se gusto tijelo, koje je prisutno u jezgrima većine stanica - tzv. nucleolus. U nukleolima dolazi do sinteze i akumulacije RNK koja se koristi za izgradnju ribozoma, kao i nekih drugih vrsta RNK.
ĆELIJA DIVISION
Iako sve ćelije nastaju deobom prethodne ćelije, ne nastavljaju sve da se dele. Na primjer, nervne ćelije u mozgu, kada se jednom formiraju, ne dijele se. Njihov broj se postepeno smanjuje; Oštećeno moždano tkivo nije u stanju da se oporavi regeneracijom. Ako ćelije nastave da se dijele, tada ih karakterizira ćelijski ciklus koji se sastoji od dvije glavne faze: interfaze i mitoze.
Sama međufaza se sastoji od tri faze: G 1, S i G 2. Ispod je njihovo trajanje, tipično za biljne i životinjske ćelije.
G 1 (4–8 sati). Ova faza počinje odmah nakon rođenja ćelije. Tokom G 1 faze, ćelija, sa izuzetkom hromozoma (koji se ne menjaju), povećava svoju masu. Ako se ćelija dalje ne dijeli, ostaje u ovoj fazi.
S (6–9 sati). Ćelijska masa nastavlja da raste i dolazi do udvostručavanja (duplikacije) hromozomske DNK. Međutim, hromozomi ostaju jednostruke strukture, iako su udvostručeni u masi, budući da su dvije kopije svakog hromozoma (hromatide) i dalje povezane jedna s drugom cijelom svojom dužinom.
G2. Masa ćelije nastavlja da raste sve dok ne bude približno dvostruko veća od svoje prvobitne mase, a zatim dolazi do mitoze.
Nakon što se hromozomi udvostruče, svaka od ćelija kćeri treba da dobije pun set hromozoma. Jednostavnom diobom stanica to se ne može postići - ovaj rezultat se postiže kroz proces koji se zove mitoza. Ne ulazeći u detalje, početak ovog procesa treba smatrati poravnanjem hromozoma u ekvatorijalnoj ravni ćelije. Zatim se svaki hromozom dijeli uzdužno na dvije hromatide, koje počinju divergirati u suprotnim smjerovima, postajući nezavisni hromozomi. Kao rezultat, kompletan set hromozoma nalazi se na oba kraja ćelije. Ćelija se tada dijeli na dva dijela, a svaka kćerka stanica prima puni set hromozoma.
Slijedi opis mitoze u tipičnoj životinjskoj ćeliji. Obično se dijeli u četiri faze.
I. Profaza. Posebna ćelijska struktura - centriol - udvostručuje se (ponekad se ovo udvostručenje događa u S-periodu interfaze), a dva centriola počinju da se razilaze na suprotne polove jezgra. Nuklearna membrana je uništena; u isto vrijeme, posebni proteini se kombinuju (agregiraju), formirajući mikrotubule u obliku niti. Centriole, koje se sada nalaze na suprotnim polovima ćelije, imaju organizacioni efekat na mikrotubule, koje se kao rezultat radijalno poređaju, formirajući strukturu koja izgledom podseća na cvet astere („zvezde“). Ostale niti mikrotubula protežu se od jednog centriola do drugog, formirajući tzv. fisiono vreteno. U ovom trenutku, hromozomi su u spiralnom stanju, nalik na oprugu. Oni su jasno vidljivi u svjetlosnom mikroskopu, posebno nakon bojenja. U profazi, hromozomi su podeljeni, ali hromatide i dalje ostaju vezane u parovima u zoni centromere - hromozomske organele slične po funkciji centriolu. Centromeri takođe imaju organizacioni efekat na filamente vretena, koji se sada protežu od centriola do centromere i od nje do drugog centriola.
II. Metafaza. Kromosomi, do ovog trenutka raspoređeni nasumično, počinju se kretati, kao da su povučeni nitima vretena pričvršćenim za njihove centromere, i postepeno se nižu u istoj ravni u određenom položaju i na jednakoj udaljenosti od oba pola. Centromere koje leže u istoj ravni zajedno sa hromozomima čine tzv. ekvatorijalna ploča. Centromeri koji povezuju parove hromatida se dijele, nakon čega se sestrinski hromozomi potpuno odvajaju.
III. Anafaza. Kromosomi svakog para kreću se u suprotnim smjerovima prema polovima, kao da ih vuku niti vretena. U ovom slučaju, niti se formiraju između centromera uparenih hromozoma.
IV. Telofaza. Čim se kromosomi približe suprotnim polovima, sama stanica počinje se dijeliti duž ravnine u kojoj se nalazila ekvatorijalna ploča. Kao rezultat, formiraju se dvije ćelije. Niti vretena se uništavaju, hromozomi se odmotaju i postaju nevidljivi, a oko njih se formira nuklearna membrana. Ćelije se vraćaju u G 1 fazu interfaze. Cijeli proces mitoze traje oko sat vremena.
Detalji mitoze se donekle razlikuju među različitim tipovima ćelija. Tipična biljna ćelija formira vreteno, ali nema centriole. Kod gljiva se mitoza javlja unutar jezgre, bez prethodnog raspada nuklearne membrane.
Sama podjela ćelije, nazvana citokineza, nema strogu vezu sa mitozom. Ponekad se javlja jedna ili više mitoza bez diobe stanica; Kao rezultat, formiraju se višejezgrene ćelije, koje se često nalaze u algama. Ako se jezgro ukloni iz jajeta morskog ježa mikromanipulacijom, vreteno nastavlja da se formira, a jaje nastavlja da se deli. Ovo pokazuje da prisustvo hromozoma nije neophodan uslov za deobu ćelije.
Reprodukcija mitozom naziva se aseksualna reprodukcija, vegetativna reprodukcija ili kloniranje. Njegov najvažniji aspekt je genetski: kod takve reprodukcije nema divergencije nasljednih faktora u potomstvu. Rezultirajuće ćelije kćeri su genetski potpuno iste kao i matična ćelija. Mitoza je jedini način samoreprodukcije kod vrsta koje nemaju spolnu reprodukciju, kao što su mnogi jednoćelijski organizmi. Međutim, čak i kod vrsta sa seksualnom reprodukcijom, tjelesne ćelije se dijele mitozom i dolaze iz jedne ćelije, oplođenog jajeta, i stoga su sve genetski identične. Više biljke mogu se razmnožavati aseksualno (koristeći mitozu) sadnicama i viticama (poznati primjer su jagode).
Seksualno razmnožavanje organizama vrši se uz pomoć specijalizovanih ćelija, tzv. gamete - oocite (jajne ćelije) i spermatozoide (spermatozoide). Gamete se spajaju i formiraju jednu ćeliju - zigotu. Svaka gameta je haploidna, tj. ima jedan set hromozoma. Unutar skupa, svi hromozomi su različiti, ali svaki hromozom jajne ćelije odgovara jednom od hromozoma sperme. Zigota, dakle, već sadrži par hromozoma koji odgovaraju jedan drugom, a koji se nazivaju homologni. Homologni hromozomi su slični jer imaju iste gene ili njihove varijante (alele) koji određuju specifične karakteristike. Na primjer, jedan od uparenih hromozoma može imati gen koji kodira krvnu grupu A, a drugi može imati varijantu koja kodira krvnu grupu B. Kromozomi zigota koji potiču iz jajeta su majčinski, a oni koji potiču iz sperme su očinski.
Kao rezultat ponovljenih mitotičkih dioba, iz nastale zigote nastaju ili višećelijski organizam ili brojne slobodno živeće stanice, kao što se događa kod protozoa koje imaju spolnu reprodukciju i kod jednoćelijskih algi.
Tokom formiranja gameta, diploidni skup hromozoma prisutan u zigoti mora biti smanjen za polovinu. Ako se to ne bi dogodilo, onda bi u svakoj generaciji fuzija gameta dovela do udvostručavanja seta hromozoma. Redukcija na haploidni broj hromozoma nastaje kao rezultat redukcijske diobe - tzv. mejoza, koja je varijanta mitoze.
Cijepanje i rekombinacija.
Posebnost mejoze je u tome što se tokom diobe ćelije ekvatorijalna ploča formira od parova homolognih hromozoma, a ne od dupliranih pojedinačnih hromozoma, kao u mitozi. Upareni hromozomi, od kojih svaki ostaje pojedinačni, divergiraju na suprotne polove ćelije, ćelija se deli, i kao rezultat, ćelije kćeri dobijaju polovinu skupa hromozoma u poređenju sa zigotom.
Na primjer, pretpostavimo da se haploidni skup sastoji od dva hromozoma. U zigoti (i prema tome u svim ćelijama organizma koje proizvode gamete) prisutni su majčinski hromozomi A i B i očevi hromozomi A" i B". Tokom mejoze mogu se podijeliti na sljedeći način:
Najvažnija stvar u ovom primjeru je činjenica da kada hromozomi divergiraju, originalni majčinski i očinski set nije nužno formiran, ali je moguća rekombinacija gena, kao kod gameta AB" i A"B u gornjem dijagramu.
Sada pretpostavimo da par hromozoma AA" sadrži dva alela - a I b– gen koji određuje krvne grupe A i B. Slično, par hromozoma “BB” sadrži alele m I n još jedan gen koji određuje krvne grupe M i N. Razdvajanje ovih alela može se odvijati na sljedeći način:
Očigledno, rezultirajuće gamete mogu sadržavati bilo koju od sljedećih kombinacija alela dva gena: am, bn, bm ili an.
Ako ima više hromozoma, tada će se parovi alela odvojiti nezavisno po istom principu. To znači da iste zigote mogu proizvesti gamete s različitim kombinacijama genskih alela i dovesti do različitih genotipova u potomstvu.
Mejotička podjela.
Oba primjera ilustruju princip mejoze. U stvari, mejoza je mnogo složeniji proces, jer uključuje dvije uzastopne diobe. Glavna stvar u mejozi je da se hromozomi udvostručuju samo jednom, dok se stanica dijeli dva puta, uslijed čega se broj kromosoma smanjuje i diploidni skup se pretvara u haploidni.
Tokom profaze prve podjele, homologni hromozomi se konjugiraju, odnosno spajaju se u parove. Kao rezultat ovog vrlo preciznog procesa, svaki gen završava nasuprot svom homologu na drugom hromozomu. Tada se oba hromozoma udvostručuju, ali hromatide ostaju međusobno povezane zajedničkom centromerom.
U metafazi, četiri povezane hromatide se poredaju da formiraju ekvatorijalnu ploču, kao da su jedan duplicirani hromozom. Suprotno onome što se dešava u mitozi, centromere se ne dijele. Kao rezultat, svaka ćelija kćerka prima par hromatida koje su još povezane centromerom. Tokom druge podjele, hromozomi, koji su već pojedinačni, ponovo se poređaju, formirajući, kao u mitozi, ekvatorijalnu ploču, ali do njihovog udvostručenja ne dolazi tokom ove diobe. Centromere se tada dijele i svaka ćerka ćelija prima jednu hromatidu.
Citoplazmatska podjela.
Kao rezultat dvije mejotičke diobe diploidne ćelije nastaju četiri ćelije. Kada se formiraju muške reproduktivne ćelije, dobiju se četiri spermatozoida približno iste veličine. Kada se formiraju jaja, podjela citoplazme odvija se vrlo neravnomjerno: jedna ćelija ostaje velika, dok su druge tri toliko male da su gotovo u potpunosti zauzete jezgrom. Ove male ćelije, tzv. polarna tijela služe samo za smještaj viška kromosoma nastalih kao rezultat mejoze. Najveći dio citoplazme potrebne za zigotu ostaje u jednoj ćeliji - jajetu.
Konjugacija i ukrštanje.
Tokom konjugacije, hromatide homolognih hromozoma mogu se razbiti i zatim spojiti u novom redosledu, razmjenjujući sekcije na sljedeći način:

Ova razmjena dijelova homolognih hromozoma naziva se crossing. Kao što je gore prikazano, ukrštanje dovodi do pojave novih kombinacija alela povezanih gena. Dakle, ako su originalni hromozomi imali kombinacije AB I ab, tada će nakon prelaska sadržavati Ab I aB. Ovaj mehanizam za nastanak novih kombinacija gena dopunjuje efekat nezavisnog sortiranja hromozoma koji se javlja tokom mejoze. Razlika je u tome što križanje razdvaja gene na istom hromozomu, dok nezavisno sortiranje razdvaja samo gene na različitim hromozomima.
ALTERNATIVNE GENERACIJE
PRIMITIVNE ĆELIJE: PROKARIOTI
Sve navedeno odnosi se na stanice biljaka, životinja, protozoa i jednoćelijskih algi, koje se zajednički nazivaju eukarioti. Eukarioti su evoluirali iz jednostavnijeg oblika, prokariota, koji su sada predstavljeni bakterijama, uključujući arhebakterije i cijanobakterije (potonje koje su se ranije nazivale plavo-zelene alge). U poređenju sa eukariotskim ćelijama, prokariotske ćelije su manje i imaju manje ćelijskih organela. Imaju ćelijsku membranu, ali im nedostaje endoplazmatski retikulum, a ribosomi slobodno plutaju u citoplazmi. Mitohondrije su odsutne, ali su oksidativni enzimi obično vezani za ćelijsku membranu, koja tako postaje ekvivalent mitohondrijama. Prokariotima također nedostaju hloroplasti, a hlorofil, ako je prisutan, prisutan je u obliku vrlo malih granula.
Prokarioti nemaju jezgro zatvoreno membranom, iako se lokacija DNK može identificirati po njenoj optičkoj gustoći. Ekvivalent hromozoma je lanac DNK, obično kružnog oblika, sa mnogo manje vezanih proteina. DNK lanac je vezan za ćelijsku membranu u jednom trenutku. Kod prokariota nema mitoze. Zamijenjuje se sljedećim procesom: DNK se udvostručuje, nakon čega stanična membrana počinje rasti između susjednih tačaka vezivanja dvije kopije molekule DNK, koje kao rezultat postupno divergiraju. Ćelija se na kraju podijeli između tačaka vezivanja molekula DNK, formirajući dvije ćelije, svaka sa svojom kopijom DNK.
ĆELIJA DIFERENCIJACIJA
Višećelijske biljke i životinje evoluirale su od jednoćelijskih organizama čije su ćelije, nakon podjele, ostale zajedno kako bi formirale koloniju. U početku su sve ćelije bile identične, ali dalja evolucija dovela je do diferencijacije. Prije svega, diferenciraju se somatske stanice (tj. tjelesne ćelije) i zametne stanice. Dalja diferencijacija postala je složenija - nastajalo je sve više različitih tipova ćelija. Ontogeneza - individualni razvoj višećelijskog organizma - ponavlja u opštem smislu ovaj evolucijski proces (filogenezu).
Fiziološki, ćelije se djelimično diferenciraju pojačavanjem jedne ili druge osobine zajedničke svim stanicama. Na primjer, kontraktilna funkcija je poboljšana u mišićnim stanicama, što može biti rezultat poboljšanja mehanizma koji izvodi ameboidne ili druge vrste pokreta u manje specijaliziranim stanicama. Sličan primjer su ćelije korijena tankih stijenki sa svojim procesima, tzv. korijenske dlake, koje služe za upijanje soli i vode; u jednom ili drugom stepenu, ova funkcija je inherentna svim ćelijama. Ponekad se specijalizacija povezuje sa stjecanjem novih struktura i funkcija - primjer je razvoj lokomotornog organa (flageluma) u spermi.
Diferencijacija na ćelijskom ili tkivnom nivou je detaljno proučavana. Znamo, na primjer, da se ponekad javlja autonomno, tj. jedna vrsta ćelija može se pretvoriti u drugu bez obzira na to koja vrsta ćelija su susjedne ćelije. Međutim, tzv embrionalna indukcija je fenomen u kojem jedna vrsta tkiva stimulira ćelije druge vrste da se diferenciraju u datom smjeru.
U opštem slučaju, diferencijacija je nepovratna, tj. visoko diferencirane ćelije ne mogu se transformisati u drugu vrstu ćelija. Međutim, to nije uvijek slučaj, posebno u biljnim stanicama.
Razlike u strukturi i funkciji u konačnici su određene tipom proteina koji se sintetizira u ćeliji. Budući da je sinteza proteina pod kontrolom gena, a skup gena je isti u svim stanicama tijela, diferencijacija mora ovisiti o aktivaciji ili inaktivaciji određenih gena u različitim tipovima stanica. Regulacija aktivnosti gena se dešava na nivou transkripcije, tj. formiranje glasničke RNK koristeći DNK kao šablon. Samo transkribovani geni proizvode proteine. Sintetizirani proteini mogu blokirati transkripciju, ali je ponekad i aktivirati. Osim toga, budući da su proteini proizvodi gena, neki geni mogu kontrolirati transkripciju drugih gena. Hormoni, posebno steroidi, također su uključeni u regulaciju transkripcije. Vrlo aktivni geni mogu se udvostručiti (udvostručiti) mnogo puta kako bi se proizvelo više glasničke RNK.
Razvoj malignih tumora često se smatra posebnim slučajem stanične diferencijacije. Međutim, pojava malignih ćelija je rezultat promena u strukturi DNK (mutacije), a ne procesa transkripcije i translacije u protein normalne DNK.
METODE ZA PROUČAVANJE ĆELIJA
Svetlosni mikroskop.
U proučavanju oblika i strukture ćelije, prvo oruđe je bio svetlosni mikroskop. Njegova moć razlučivanja ograničena je dimenzijama uporedivim sa talasnom dužinom svetlosti (0,4–0,7 μm za vidljivu svetlost). Međutim, mnogi elementi stanične strukture su mnogo manjih dimenzija.
Druga poteškoća je u tome što je većina ćelijskih komponenti providna i ima indeks prelamanja gotovo isti kao voda. Da bi se poboljšala vidljivost, često se koriste boje koje imaju različite afinitete za različite ćelijske komponente. Bojenje se također koristi za proučavanje ćelijske hemije. Na primjer, neke boje se prvenstveno vezuju za nukleinske kiseline i time otkrivaju njihovu lokalizaciju u ćeliji. Mali dio boja – koje se nazivaju intravitalne boje – može se koristiti za bojenje živih stanica, ali obično se ćelije moraju prvo fiksirati (koristeći supstancije za koagulaciju proteina) prije nego što se mogu bojati. Cm. HISTOLOGIJA.
Prije testiranja, ćelije ili komadi tkiva obično se ugrađuju u parafin ili plastiku, a zatim se mikrotomom režu na vrlo tanke dijelove. Ova metoda se široko koristi u kliničkim laboratorijama za identifikaciju tumorskih stanica. Osim konvencionalne svjetlosne mikroskopije, razvijene su i druge optičke metode za proučavanje ćelija: fluorescentna mikroskopija, fazno-kontrastna mikroskopija, spektroskopija i analiza difrakcije rendgenskih zraka.
Elektronski mikroskop.
Elektronski mikroskop ima rezoluciju od cca. 1–2 nm. Ovo je dovoljno za proučavanje velikih proteinskih molekula. Obično je potrebno obojiti i kontrastirati predmet metalnim solima ili metalima. Iz tog razloga, kao i zbog toga što se objekti ispituju u vakuumu, pomoću elektronskog mikroskopa mogu se proučavati samo ubijene ćelije.
Autoradiografija.
Ako se radioaktivni izotop koji apsorbiraju stanice tijekom metabolizma doda mediju, njegova unutarćelijska lokalizacija se tada može otkriti pomoću autoradiografije. Ovom metodom tanki dijelovi ćelija se postavljaju na film. Film potamni ispod onih mjesta gdje se nalaze radioaktivni izotopi.
Centrifugiranje.
Za biohemijsko proučavanje ćelijskih komponenti, ćelije se moraju uništiti - mehanički, hemijski ili ultrazvukom. Oslobođene komponente su suspendovane u tečnosti i mogu se izolovati i pročistiti centrifugiranjem (najčešće u gradijentu gustine). Obično takve pročišćene komponente zadržavaju visoku biohemijsku aktivnost.
Ćelijske kulture.
Neka tkiva se mogu podijeliti na pojedinačne ćelije tako da ćelije ostaju žive i često se mogu razmnožavati. Ova činjenica definitivno potvrđuje ideju o ćeliji kao životnoj jedinici. Sunđer, primitivni višećelijski organizam, može se razdvojiti na ćelije trljanjem kroz sito. Nakon nekog vremena, ove ćelije se ponovo povezuju i formiraju spužvu. Embrionalna tkiva životinja mogu se natjerati da se razdvoje pomoću enzima ili drugih sredstava koja slabe veze između stanica.
Američki embriolog R. Harrison (1879–1959) bio je prvi koji je pokazao da embrionalne, pa čak i neke zrele ćelije mogu rasti i razmnožavati se izvan tijela u prikladnom okruženju. Ovu tehniku, nazvanu kultivisanje ćelija, usavršio je francuski biolog A. Carrel (1873–1959). Biljne ćelije se takođe mogu uzgajati u kulturi, ali u poređenju sa životinjskim ćelijama one formiraju veće nakupine i čvršće su vezane jedna za drugu, tako da se tkiva formiraju kako kultura raste, a ne pojedinačne ćelije. U kulturi ćelija, cijela odrasla biljka, kao što je šargarepa, može se uzgajati iz jedne ćelije.
Mikrohirurgija.
Koristeći mikromanipulator, pojedinačni dijelovi ćelije mogu se ukloniti, dodati ili na neki način modificirati. Velika ćelija amebe može se podijeliti na tri glavne komponente - ćelijsku membranu, citoplazmu i jezgro, a zatim se te komponente mogu ponovo sastaviti i formirati živu ćeliju. Na taj način se mogu dobiti umjetne ćelije koje se sastoje od komponenti različitih vrsta ameba.
Ako uzmemo u obzir da je moguće umjetno sintetizirati neke ćelijske komponente, onda bi eksperimenti u sklapanju umjetnih stanica mogli biti prvi korak ka stvaranju novih oblika života u laboratoriju. Budući da se svaki organizam razvija iz jedne ćelije, metoda proizvodnje umjetnih stanica u principu omogućava izgradnju organizama date vrste, ako se istovremeno koriste komponente koje se neznatno razlikuju od onih koje se nalaze u postojećim stanicama. U stvarnosti, međutim, nije potrebna potpuna sinteza svih ćelijskih komponenti. Strukturu većine, ako ne i svih, komponenti ćelije određuju nukleinske kiseline. Dakle, problem stvaranja novih organizama svodi se na sintezu novih vrsta nukleinskih kiselina i njihovu zamjenu prirodnim nukleinskim kiselinama u određenim stanicama.
Fuzija ćelija.
Druga vrsta umjetnih stanica može se dobiti spajanjem stanica iste ili različitih vrsta. Da bi se postigla fuzija, ćelije su izložene virusnim enzimima; u ovom slučaju, vanjske površine dvije ćelije su zalijepljene zajedno, a membrana između njih je uništena i formira se ćelija u kojoj su dva seta hromozoma zatvorena u jednom jezgru. Moguće je spajanje ćelija različitih tipova ili u različitim fazama diobe. Ovom metodom bilo je moguće dobiti hibridne ćelije miša i piletine, čovjeka i miša te čovjeka i žabe. Takve ćelije su hibridne samo u početku, a nakon brojnih dioba stanica gube većinu hromozoma jednog ili drugog tipa. Konačni proizvod postaje, na primjer, u suštini mišja ćelija bez prisutnih ljudskih gena ili ih ima samo u tragovima. Posebno je zanimljiva fuzija normalnih i malignih ćelija. U nekim slučajevima hibridi postaju maligni, u drugima ne, tj. oba svojstva mogu se manifestovati i kao dominantna i kao recesivna. Ovaj rezultat nije neočekivan, jer malignitet može biti uzrokovan raznim faktorima i ima složen mehanizam.
književnost:
Ham A., Cormack D. Histologija, tom 1. M., 1982
Alberts B., Bray D., Lewis J., Raff M., Roberts K., Watson J. Molekularna biologija ćelije, tom 1. M., 1994







