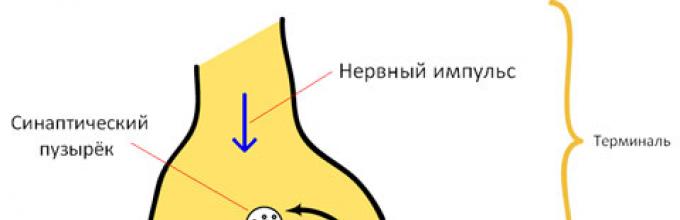
Mišićne i žljezdane ćelije se prenose kroz posebnu strukturnu formaciju - sinapsu.
Synapse- struktura koja osigurava provođenje signala od jednog do drugog. Termin je uveo engleski fiziolog C. Sherington 1897. godine.
Struktura sinapse
Sinapse se sastoje od tri glavna elementa: presinaptičke membrane, postsinaptičke membrane i sinaptičke pukotine (slika 1).
Rice. 1. Struktura sinapse: 1 - mikrotubule; 2 - mitohondrije; 3 - sinaptičke vezikule sa transmiterom; 4 - presinaptička membrana; 5 - postsinaptička membrana; 6 - receptori; 7 - sinaptički rascjep
Neki elementi sinapsi mogu imati druga imena. Na primjer, sinaptički plak je sinapsa između, krajnja ploča je postsinaptička membrana, motorni plak je presinaptički završetak aksona na mišićnom vlaknu.

presinaptička membrana pokriva prošireni nervni završetak, koji je neurosekretorni aparat. Presinaptički dio sadrži vezikule i mitohondrije koji osiguravaju sintezu medijatora. Medijatori se talože u granulama (mehurićima).
Postsinaptička membrana - zadebljani dio ćelijske membrane sa kojim je presinaptička membrana u kontaktu. Ima jonske kanale i sposoban je da generiše akcione potencijale. Osim toga, postoje posebne proteinske strukture- receptori koji percipiraju djelovanje medijatora.
Sinaptički rascjep je prostor između presinaptičke i postsinaptičke membrane, ispunjen tekućinom sličnom po sastavu.

Rice. Struktura sinapse i procesi koji se odvijaju tokom prenosa sinaptičkog signala
Vrste sinapsi
Sinapse se klasificiraju prema lokaciji, prirodi djelovanja i načinu prijenosa signala.
Po lokaciji Razlikuju neuromuskularne sinapse, neuroglandularne i neuroneuronske; potonji se, pak, dijele na akso-aksonalne, akso-dendritske, akso-somatske, dendro-somatske, dendro-dendrotske.
Po prirodi radnje Sinapse na perceptivnoj strukturi mogu biti ekscitatorne ili inhibitorne.
Metodom prenosa signala Sinapse se dijele na električne, kemijske i mješovite.
Tabela 1. Klasifikacija i tipovi sinapsi

Klasifikacija sinapsi i mehanizam prijenosa ekscitacije
Sinapse se klasifikuju na sledeći način:
- po lokaciji - periferni i centralni;
- po prirodi njihovog djelovanja - uzbudljivo i inhibitorno;
- po načinu prijenosa signala - hemijski, električni, mješoviti;
- prema medijatoru preko kojeg se prenosi - holinergički, adrenergički, serotonergički itd.
Uzbuđenje se prenosi kroz posrednici(posrednici).
Posrednici- molekule hemijske supstance, koji osiguravaju prijenos ekscitacije u sinapsama. Drugim riječima, kemijske tvari uključene u prijenos ekscitacije ili inhibicije s jedne ekscitabilne ćelije na drugu.
Svojstva medijatora
- Sintetizirano u neuronu
- Akumuliraju se na kraju ćelije
- Oslobađa se kada se ion Ca2+ pojavi u presinaptičkom terminalu
- Imaju specifičan učinak na postsinaptičku membranu
By hemijska struktura Medijatori se mogu podijeliti na amine (noradrenalin, dopamin, serotonin), aminokiseline (glicin, gama-aminobutirna kiselina) i polipeptide (endorfini, enkefalini). Acetilholin je uglavnom poznat kao ekscitatorni neurotransmiter i nalazi se u različitim dijelovima centralnog nervnog sistema. Predajnik se nalazi u vezikulama presinaptičkog zadebljanja (sinaptički plak). Medijator se sintetizira u neuronskim stanicama i može se ponovo sintetizirati iz metabolita njegovog cijepanja u sinaptičkom pukotinu.
Kada su terminali aksona pobuđeni, membrana sinaptičkog plaka se depolarizira, uzrokujući da ioni kalcija teku iz ekstracelularnog okruženja u nervni završetak kroz kalcijumske kanale. Joni kalcija stimuliraju kretanje sinaptičkih vezikula do presinaptičke membrane, njihovo spajanje s njom i naknadno oslobađanje transmitera u sinaptički rascjep. Nakon prodiranja u jaz, transmiter difundira do postsinaptičke membrane koja sadrži receptore na svojoj površini. Interakcija transmitera sa receptorima izaziva otvaranje natrijumskih kanala, što doprinosi depolarizaciji postsinaptičke membrane i pojavi ekscitatornog postsinaptičkog potencijala. U neuromuskularnoj sinapsi ovaj potencijal se naziva potencijal krajnje ploče. Lokalne struje nastaju između depolarizirane postsinaptičke membrane i susjednih polariziranih dijelova iste membrane, koje depolariziraju membranu do kritičnog nivoa, nakon čega slijedi stvaranje akcionog potencijala. Akcijski potencijal se širi preko svih membrana, na primjer, mišićnog vlakna i uzrokuje njegovu kontrakciju.
Odašiljač koji se oslobađa u sinaptičku pukotinu vezuje se za receptore postsinaptičke membrane i cijepa ga odgovarajući enzim. Dakle, holinesteraza uništava neurotransmiter acetilkolin. Nakon toga, određena količina produkata razgradnje medijatora ulazi u sinaptički plak, gdje se iz njih ponovo sintetizira acetilkolin.
Tijelo sadrži ne samo ekscitatorne, već i inhibitorne sinapse. Po mehanizmu prijenosa ekscitacije slični su ekscitatornim sinapsama. U inhibitornim sinapsama, transmiter (na primjer, gama-aminobutirna kiselina) se vezuje za receptore na postsinaptičkoj membrani i potiče otvaranje u njoj. U tom slučaju se aktivira prodor ovih jona u ćeliju i razvija se hiperpolarizacija postsinaptičke membrane, što uzrokuje pojavu inhibitornog postsinaptičkog potencijala.
Sada je otkriveno da se jedan medijator može vezati za nekoliko različitih receptora i izazvati različite reakcije.
Hemijske sinapse
Fiziološka svojstva hemijskih sinapsi
Sinapse sa hemijskim prenosom ekscitacije imaju određena svojstva:
- ekscitacija se provodi u jednom smjeru, budući da se transmiter oslobađa samo iz sinaptičkog plaka i stupa u interakciju s receptorima na postsinaptičkoj membrani;
- širenje ekscitacije kroz sinapse događa se sporije nego duž nervnog vlakna (sinaptičko kašnjenje);
- prijenos ekscitacije vrši se pomoću specifičnih medijatora;
- mijenja se ritam ekscitacije u sinapsama;
- sinapse se mogu umoriti;
- sinapse su vrlo osjetljive na razne kemikalije i hipoksiju.
Jednosmjerni prijenos signala. Signal se prenosi samo sa presinaptičke membrane na postsinaptičku membranu. Ovo proizilazi iz strukturnih karakteristika i svojstava sinaptičkih struktura.
Sporo prenos signala. Uzrokovano sinaptičkim kašnjenjem u prijenosu signala iz jedne ćelije u drugu. Kašnjenje je uzrokovano vremenom potrebnim za procese oslobađanja transmitera, njegove difuzije na postsinaptičku membranu, vezivanja za receptore postsinaptičke membrane, depolarizacije i konverzije postsinaptičkog potencijala u AP (akcioni potencijal). Trajanje sinaptičkog kašnjenja kreće se od 0,5 do 2 ms.
Sposobnost sumiranja efekata signala koji pristižu u sinapsu. Ovo zbrajanje se pojavljuje ako sljedeći signal stigne u sinapsu kratko vrijeme (1-10 ms) nakon prethodnog. U takvim slučajevima, EPSP amplituda se povećava i viša AP frekvencija se može generirati na postsinaptičkom neuronu.
Transformacija ritma uzbuđenja. Učestalost nervnih impulsa koji pristižu na presinaptičku membranu obično ne odgovara učestalosti AP koje generiše postsinaptički neuron. Izuzetak su sinapse koje prenose ekscitaciju s nervnog vlakna na skeletni mišić.
Mala labilnost i visok zamor sinapsi. Sinapse mogu provesti 50-100 nervnih impulsa u sekundi. To je 5-10 puta manje od maksimalne AP frekvencije koju nervna vlakna mogu reproducirati kada su električno stimulirana. Ako se nervna vlakna smatraju praktički neumornima, onda se na sinapsama vrlo brzo razvija umor. To se događa zbog iscrpljivanja rezervi transmitera, energetskih resursa, razvoja trajne depolarizacije postsinaptičke membrane itd.
Visoka osjetljivost sinapsi na djelovanje biološki aktivnih supstanci, lijekova i otrova. Na primjer, otrov strihnin blokira funkciju inhibitornih sinapsi u centralnom nervnom sistemu vezivanjem za receptore osjetljive na posrednik glicin. Tetanusni toksin blokira inhibitorne sinapse, ometajući oslobađanje transmitera iz presinaptičkog terminala. U oba slučaja razvijaju se po život opasne pojave. Gore su razmotreni primjeri djelovanja biološki aktivnih supstanci i otrova na prijenos signala u neuromuskularnim sinapsama.
Olakšavajuća i depresivna svojstva sinoptičke transmisije. Olakšavanje sinaptičke transmisije nastaje kada nervni impulsi stignu u sinapsu nakon kratkog vremena (10-50 ms) jedan za drugim, tj. dovoljno često. Štaviše, tokom određenog vremenskog perioda, svaki naredni PD koji stiže na presinaptičku membranu uzrokuje povećanje sadržaja transmitera u sinaptičkom pukotinu, povećanje amplitude EPSP-a i povećanje efikasnosti sinaptičke transmisije.
Jedan od mehanizama olakšavanja je nakupljanje Ca 2 jona u presinaptičkom terminalu. Potrebno je nekoliko desetina milisekundi da kalcijumska pumpa ukloni dio kalcija koji je ušao u sinaptički terminal po dolasku AP-a. Ako u to vrijeme dođe novi akcioni potencijal, tada novi dio kalcija ulazi u terminal i njegov učinak na oslobađanje neurotransmitera dodaje se preostaloj količini kalcija koju kalcijeva pumpa nije uspjela ukloniti iz neuroplazme terminal.
Postoje i drugi mehanizmi za razvoj reljefa. Ovaj fenomen se naziva i u klasičnim udžbenicima fiziologije post-tetanična potenciranje. Olakšavanje sinaptičkog prijenosa važno je u funkcioniranju mehanizama pamćenja, za formiranje uslovljeni refleksi i obuku. Olakšavanje prijenosa signala je u osnovi razvoja sinaptičke plastičnosti i poboljšanja njihovih funkcija uz čestu aktivaciju.
Depresija (inhibicija) prijenosa signala u sinapsama nastaje kada vrlo česti (za neuromuskularnu sinapsu više od 100 Hz) nervni impulsi stignu do presinaptičke membrane. U mehanizmima razvoja fenomena depresije dolazi do iscrpljivanja rezervi transmitera u presinaptičkom terminalu, smanjenja osjetljivosti receptora postsinaptičke membrane na transmiter, te razvoja uporne depolarizacije postsinaptičke membrane, što otežava generiranje AP na membrani postsinaptičke ćelije, su važne.
Električne sinapse
Pored sinapsi sa hemijskim prenosom ekscitacije, telo ima sinapse sa električnim prenosom. Ove sinapse imaju vrlo uski sinaptički rascjep i reducirani električni otpor između dve membrane. Zbog prisutnosti poprečnih kanala između membrana i malog otpora, električni impuls lako prolazi kroz membrane. Električne sinapse su obično karakteristične za ćelije istog tipa.
Kao rezultat izlaganja stimulusu, presinaptički akcijski potencijal pobuđuje postsinaptičku membranu, gdje se javlja propagirajući akcijski potencijal.
Odlikuje ih veća brzina ekscitacije u odnosu na hemijske sinapse i niska osetljivost na dejstvo hemikalija.
Električne sinapse imaju jednosmjerni i dvosmjerni prijenos ekscitacije.
Električne inhibitorne sinapse se također nalaze u tijelu. Inhibicijski učinak nastaje djelovanjem struje koja uzrokuje hiperpolarizaciju postsinaptičke membrane.
U mješovitim sinapsama, ekscitacija se može prenijeti pomoću električnih impulsa i medijatora.
Moscow PsychologicalDruštveni institut (MSSI)
Sažetak o anatomiji centralnog nervnog sistema na temu:
SYNAPSES(struktura, struktura, funkcije).
student 1. godine Psihološkog fakulteta,
grupa 21/1-01 Logachev A.Yu.
Učitelj:
Kholodova Marina Vladimirovna.
godine 2001.
Plan rada:
1.Prolog.
2. Fiziologija neurona i njegova struktura.
3. Struktura i funkcije sinapse.
4. Hemijska sinapsa.
5. Izolacija posrednika.
6. Hemijski posrednici i njihove vrste.
7.Epilog.
8. Spisak referenci.
PROLOG:
Naše tijelo je jedan veliki satni mehanizam. Sastoji se od ogromnog broja sićušnih čestica koje se nalaze u u strogom redu a svaki od njih obavlja određene funkcije i ima svoje jedinstvena svojstva. Ovaj mehanizam - tijelo, sastoji se od ćelija, koje povezuju svoja tkiva i sisteme: sve to kao cjelina predstavlja jedan lanac, supersistem tijela. Najveća raznolikost ćelijskih elemenata ne bi mogla funkcionirati kao jedinstvena cjelina da u tijelu ne postoji sofisticirani regulatorni mehanizam. Nervni sistem igra posebnu ulogu u regulaciji. Sav naporan rad nervni sistem- regulisanje rada unutrašnje organe, kontrola pokreta, bilo da su jednostavni i nesvjesni pokreti (na primjer, disanje) ili složeni pokreti ruku osobe - sve se to, u suštini, temelji na interakciji stanica jedna s drugom. Sve se to u suštini zasniva na prijenosu signala iz jedne ćelije u drugu. Štaviše, svaka ćelija radi svoj posao, a ponekad ima i nekoliko funkcija. Raznolikost funkcija obezbjeđuju dva faktora: način na koji su ćelije međusobno povezane i način na koji su te veze raspoređene.
FIZIOLOGIJA NEURONA I NJEGOVA STRUKTURA:
Najjednostavnija reakcija nervnog sistema na spoljašnji stimulans - to je refleks. Prije svega, razmotrimo strukturu i fiziologiju strukturne elementarne jedinice nervnog tkivaživotinje i ljudi - neuron. Funkcionalna i osnovna svojstva neurona određena su njegovom sposobnošću da se uzbuđuje i samouzbudi. Prenos ekscitacije vrši se duž procesa neurona - aksona i dendrita.
Aksoni su duži i širi procesi. Imaju niz specifičnih svojstava: izolovanu provodljivost ekscitacija i bilateralno provođenje.
Nervne ćelije su sposobne ne samo da percipiraju i obrađuju vanjsku stimulaciju, već i spontano proizvode impulse koji nisu uzrokovani vanjskom stimulacijom (samopobuda). Kao odgovor na stimulaciju, neuron odgovara impuls aktivnosti- akcijski potencijal čija se frekvencija generiranja kreće od 50-60 impulsa u sekundi (za motorne neurone) do 600-800 impulsa u sekundi (za interneurone mozga). Akson se završava mnogim tankim granama tzv terminali. Od terminala impuls prelazi na druge ćelije, direktno na njihova tijela ili, češće, na njihove dendritske procese. Broj terminala u aksonu može doseći i do hiljadu, koji završavaju u različitim ćelijama. S druge strane, tipični neuron kralježnjaka ima između 1.000 i 10.000 terminala iz drugih stanica.
Dendriti - kraći i brojniji procesineurona. Oni opažaju ekscitaciju od susjednih neurona i provode je do tijela ćelije. Postoje kašaste i nepulpirane nervne ćelije i vlakna.
Vlakna pulpe su dio osjetljivih imotoričkih nerava skeletnih mišića i osjetilnih organaPrekrivene su lipidnom mijelinskom ovojnicom. Vlakna pulpe su "brzodjelujuća": u takvim vlaknima promjera 1-3,5 mikromilimetara, ekscitacija se širi brzinom od 3-18 m/s. To se objašnjava činjenicom da se provođenje impulsa duž mijeliniziranog živca odvija grčevito. U ovom slučaju, akcioni potencijal "skače" kroz područje živca prekrivenog mijelinom i na Ranvierovom čvoru (izloženo područje živca), prelazi na omotač aksijalnog cilindra živca. vlakna. Mijelinska ovojnica je dobar izolator i sprječava prijenos ekscitacije na spoj paralelnih nervnih vlakana.
Nemišićna vlakna čine većinu simpatičkih nerava. Nemaju mijelinsku ovojnicu i odvojene su jedna od druge neuroglijalnim ćelijama.
U vlaknima bez pulpe ćelije djeluju kao izolatori. neuroglia(nervno potporno tkivo). Schwannove ćelije - jedna od vrsta glijalnih ćelija. Pored unutrašnjih neurona koji percipiraju i transformišu impulse koji dolaze od drugih neurona, postoje neuroni koji percipiraju uticaje direktno od okruženje- Ovo receptori, kao i neuroni koji direktno utiču na izvršne organe - efektori, na primjer, na mišićima ili žlijezdama. Ako neuron djeluje na mišić, naziva se motorni neuron ili motorni neuron. Među neuroreceptorima postoji 5 vrsta ćelija, u zavisnosti od vrste patogena:
- fotoreceptori, koji se pobuđuju pod uticajem svetlosti i obezbeđuju funkcionisanje organa vida,
- mehanoreceptori, oni receptori koji reaguju na mehaničke uticaje. Nalaze se u organima sluha i ravnoteže. Dodirne ćelije su takođe mehanoreceptori. Neki mehanoreceptori se nalaze u mišićima i mjere stepen njihovog istezanja.
- hemoreceptori - selektivno reagiraju na prisutnost ili promjenu koncentracije raznih hemikalija, na njima se zasniva rad organa mirisa i ukusa,
- termoreceptori, reaguju na promjene temperature ili njenog nivoa - receptori za hladnoću i toplinu,
- elektroreceptori reaguju na trenutne impulse, a prisutni su kod nekih riba, vodozemaca i sisara, na primjer, platipusa.
Na osnovu gore navedenog, želio bih napomenuti da je dugo vremena među biolozima koji su proučavali nervni sistem postojalo mišljenje da nervne ćelije formiraju dugačke složene mreže koje se kontinuirano pretvaraju jedna u drugu.
Međutim, 1875. godine, italijanski naučnik, profesor histologije na Univerzitetu u Paviji, došao je do novi način bojenje ćelija - srebrenje. Kada jedna od hiljada obližnjih ćelija posrebri, samo je ona obojena - jedina, ali potpuno, sa svim svojim procesima. Golgijeva metoda uvelike pomogao proučavanju strukture nervnih ćelija. Njegova upotreba je pokazala da je, unatoč činjenici da su stanice u mozgu smještene izuzetno blizu jedna drugoj, a njihovi procesi zbunjeni, svaka stanica još uvijek jasno odvojena. To jest, mozak se, kao i druga tkiva, sastoji od pojedinačnih ćelija koje nisu ujedinjene u zajedničku mrežu. Ovaj zaključak je izveo španski histolog S. Ramon y Cahalem, koji je time proširio ćelijsku teoriju na nervni sistem. Odbacivanje koncepta povezane mreže značilo je to u nervnom sistemu puls prelazi od ćelije do ćelije ne direktnim električnim kontaktom, već kroz jaz
Kada se koristio u biologiji? elektronski mikroskop, koji je izmišljen 1931 M. Knollem I E. Ruska, ove ideje o prisustvu jaza dobile su direktnu potvrdu.
STRUKTURA I FUNKCIJA SINAPSE:
Svaki višećelijski organizam, svakom tkivu koje se sastoji od ćelija potrebni su mehanizmi koji osiguravaju međućelijske interakcije. Pogledajmo kako se izvode interneuronskiinterakcije. Informacije putuju duž nervnih ćelija u obliku akcioni potencijali. Prijenos ekscitacije sa terminala aksona na inervirani organ ili drugu živčanu ćeliju odvija se kroz međustanične strukturne formacije - sinapse(iz grčkog "Sinapsis"- veza, veza). Koncept sinapse uveo je engleski fiziolog C. Sherington 1897. godine, da označi funkcionalni kontakt između neurona. Treba napomenuti da je još 60-ih godina prošlog vijeka NJIH. Sechenov naglasio da je bez međućelijske komunikacije nemoguće objasniti metode nastanka čak i najelementarnijeg nervnog procesa. Što je nervni sistem složeniji, i veći broj komponente nervnih moždanih elemenata, značaj sinaptičkih kontakata postaje važniji.
Različiti sinaptički kontakti se međusobno razlikuju. Međutim, uz svu raznolikost sinapsi, postoje određene opšta svojstva njihove strukture i funkcije. Stoga prvo opisujemo opšte principe njihovog funkcionisanja.
Sinapsa - je složena struktura formacija koja se sastoji od presinaptičke membrane (najčešće je to krajnja grana aksona), postsinaptičke membrane (najčešće je to dio tjelesne membrane ili dendrita drugog neurona), kao i sinaptičke pukotine.
Mehanizam prijenosa kroz sinapse dugo je ostao nejasan, iako je bilo očito da se prijenos signala u sinaptičkom području oštro razlikuje od procesa provođenja akcionog potencijala duž aksona. Međutim, početkom 20. stoljeća formulirana je hipoteza da se sinaptički prijenos događa ili električni ili hemijski. Električna teorija sinaptičke transmisije u centralnom nervnom sistemu bila je priznata do ranih 50-ih godina, ali je značajno izgubila tlo nakon što je hemijska sinapsa demonstrirana u brojnim slučajevima. periferne sinapse. Na primjer, A.V. Kibjakov, provođenje eksperimenta na nervnom gangliju, kao i korištenje mikroelektrodne tehnologije za unutarćelijsko snimanje sinaptičkih potencijala
neuroni centralnog nervnog sistema omogućili su nam da izvučemo zaključak o hemijskoj prirodi prenosa u interneuronskim sinapsama kičmena moždina.
Studije mikroelektroda posljednjih godina pokazala je da na određenim interneuronskim sinapsama postoji mehanizam električnog prijenosa. Sada je postalo očigledno da postoje sinapse i sa mehanizmom hemijskog prenosa i sa električnim. Štoviše, u nekim sinaptičkim strukturama i električni i hemijski mehanizmi prenosi su tzv mješovite sinapse.
Područje kontakta između dva neurona naziva se sinapse.
Unutrašnja struktura aksodendritske sinapse.A) Električne sinapse. Električne sinapse su rijetke u nervnom sistemu sisara. Nastaju od spojeva (neksusa) između dendrita ili soma susjednih neurona, koji su povezani citoplazmatskim kanalima promjera 1,5 nm. Proces prijenosa signala odvija se bez sinaptičkog kašnjenja i bez učešća medijatora.
Kroz električne sinapse, elektrotonični potencijali se mogu širiti s jednog neurona na drugi. Zbog bliskog sinaptičkog kontakta, modulacija prijenosa signala je nemoguća. Zadatak ovih sinapsi je da istovremeno pobuđuju neurone koji obavljaju istu funkciju. Primjer su neuroni respiratornog centra duguljaste moždine, koji sinhrono generiraju impulse tijekom udisanja. Osim toga, primjer su neuronski krugovi koji kontroliraju sakadama, u kojima se tačka fiksacije pogleda pomiče s jednog objekta pažnje na drugi.
b) Hemijske sinapse. Većina sinapsi u nervnom sistemu su hemijske. Funkcioniranje takvih sinapsi ovisi o oslobađanju transmitera. Klasična hemijska sinapsa je predstavljena presinaptičkom membranom, sinaptičkim rascjepom i postsinaptičkom membranom. Presinaptička membrana je dio štapićastog produžetka nervnog završetka ćelije koji prenosi signal, a postsinaptička membrana je dio ćelije koji prima signal.
Transmiter se oslobađa iz produžetka klavate egzocitozom, prolazi kroz sinaptički rascjep i vezuje se za receptore na postsinaptičkoj membrani. Ispod postsinaptičke membrane nalazi se subsinaptička aktivna zona, u kojoj nakon aktivacije receptora postsinaptičke membrane nastaju različiti biohemijski procesi.
Nastavak u obliku štapa sadrži sinaptičke vezikule koje sadrže medijatore, kao i veliki broj mitohondrija i cisterni glatkog endoplazmatskog retikuluma. Korištenje tradicionalnih tehnika fiksacije u proučavanju stanica omogućuje razlikovanje presinaptičkih brtvila na presinaptičkoj membrani, ograničavajući aktivne zone sinapse, na koje se sinaptičke vezikule usmjeravaju uz pomoć mikrotubula.
 Aksodendritska sinapsa.
Aksodendritska sinapsa. Presjek uzorka kičmene moždine: sinapsa između terminalnog dijela dendrita i, vjerovatno, motornog neurona.
Za ekscitatorne sinapse karakteristično je prisustvo okruglih sinaptičkih vezikula i postsinaptička zbijenost.
Dendrit je izrezan u poprečnom smjeru, o čemu svjedoči prisustvo mnogih mikrotubula.
Osim toga, vidljivi su i neki neurofilamenti. Mjesto sinapse je okruženo protoplazmatskim astrocitom.
 Procesi koji se javljaju u dvije vrste nervnih završetaka.
Procesi koji se javljaju u dvije vrste nervnih završetaka. (A) Sinaptički prijenos male molekule(npr. glutamat).
(1) Transportne vezikule koje sadrže membranske proteine sinaptičkih vezikula usmjeravaju se duž mikrotubula do plazma membrane zadebljanja u obliku štapa.
Istovremeno, molekuli enzima i glutamata se prenose sporim transportom.
(2) Proteini membrane vezikula izlaze iz plazma membrane i formiraju sinaptičke vezikule.
(3) Glutamat se puni u sinaptičke vezikule; dolazi do akumulacije medijatora.
(4) Vezikule koje sadrže glutamat približavaju se presinaptičkoj membrani.
(5) Kao rezultat depolarizacije dolazi do egzocitoze medijatora iz djelomično uništenih vezikula.
(6) Oslobođeni transmiter se difuzno širi u predelu sinaptičke pukotine i aktivira specifične receptore na postsinaptičkoj membrani.
(7) Membrane sinaptičkih vezikula se transportuju nazad u ćeliju endocitozom.
(8) Djelomično ponovno preuzimanje glutamata u ćeliju se dešava za ponovnu upotrebu.
(B) Prenos neuropeptida (npr. supstance P) koji se odvija istovremeno sa sinaptičkim prenosom (npr. glutamat).
Zajednički prijenos ovih supstanci odvija se u centralnim nervnim završecima unipolarnih neurona, koji osiguravaju osjetljivost na bol.
(1) Vezikule i peptidni prekursori (propeptidi) sintetizirani u Golgijevom kompleksu (u perikarionskoj regiji) se brzim transportom transportuju u produžetak u obliku batine.
(2) Kada uđu u područje zadebljanja u obliku štapa, proces formiranja molekule peptida je završen, a vezikule se transportuju do plazma membrane.
(3) Depolarizacija membrane i prijenos sadržaja vezikula u međućelijski prostor egzocitozom.
(4) Istovremeno se oslobađa glutamat.
1. Aktivacija receptora. Molekuli transmitera prolaze kroz sinaptički rascjep i aktiviraju receptorske proteine smještene u parovima na postsinaptičkoj membrani. Aktivacija receptora pokreće ionske procese koji dovode do depolarizacije postsinaptičke membrane (ekscitatorno postsinaptičko djelovanje) ili hiperpolarizacije postsinaptičke membrane (inhibitorno postsinaptičko djelovanje). Promjena elektrotoničnosti se prenosi na somu u obliku elektrotonskog potencijala koji se raspada kako se širi, zbog čega se mijenja potencijal mirovanja u početnom segmentu aksona.
Jonski procesi su detaljno opisani u posebnom članku na web stranici. Kada ekscitatorni postsinaptički potencijali prevladavaju, početni segment aksona se depolarizira do graničnog nivoa i stvara akcioni potencijal.
Najčešći ekscitatorni neurotransmiter centralnog nervnog sistema je glutamat, a inhibitorni gama-aminobutirna kiselina (GABA). U perifernom nervnom sistemu acetilholin služi kao transmiter za motorne neurone prugasto-prugastih mišića, a glutamat za senzorne neurone.
Redoslijed procesa koji se odvijaju u glutamatergijskim sinapsama prikazan je na donjoj slici. Kada se glutamat prenosi zajedno sa drugim peptidima, oslobađanje peptida se dešava preko ekstrasinaptičkih puteva.
Većina senzornih neurona, osim glutamata, luči i druge peptide (jedan ili više), koji se oslobađaju u različitim dijelovima neurona; međutim, glavna funkcija ovih peptida je da moduliraju (povećavaju ili smanjuju) efikasnost sinaptičke transmisije glutamata.
Osim toga, neurotransmisija se može dogoditi putem difuznog ekstrasinaptičkog prijenosa signala, karakterističnog za monoaminergičke neurone (neurone koji koriste biogene amine da posreduju u neurotransmisiji). Postoje dvije vrste monoaminergičkih neurona. U nekim neuronima, kateholamini (noradrenalin ili dopamin) se sintetiziraju iz aminokiseline tirozin, a u drugima se serotonin sintetizira iz aminokiseline triptofana. Na primjer, dopamin se oslobađa i u sinaptičkom području i iz aksonalnih varikoziteta, u kojima se također događa sinteza ovog neurotransmitera.
Dopamin prodire u međućelijsku tečnost centralnog nervnog sistema i, pre razgradnje, može da aktivira specifične receptore na udaljenosti do 100 mikrona. Monoaminergički neuroni su prisutni u mnogim strukturama centralnog nervnog sistema; poremećaj prijenosa impulsa od strane ovih neurona dovodi do raznih bolesti, uključujući Parkinsonovu bolest, šizofreniju i veliku depresiju.
Dušikov oksid (gasoviti molekul) je također uključen u difuznu neurotransmisiju u glutamatergičnom neuronskom sistemu. Prekomjerna količina dušikovog oksida ima citotoksično djelovanje, posebno u onim područjima gdje je poremećena opskrba krvlju zbog arterijske tromboze. Glutamat je također potencijalno citotoksični neurotransmiter.
Za razliku od difuzne neurotransmisije, tradicionalni sinaptički prijenos signala naziva se "provodnikom" zbog svoje relativne stabilnosti.
V) Sažetak. Multipolarni neuroni CNS-a sastoje se od some, dendrita i aksona; akson formira kolateralne i terminalne grane. Soma sadrži glatki i grubi endoplazmatski retikulum, Golgi komplekse, neurofilamente i mikrotubule. Mikrotubule prožimaju ceo neuron, učestvuju u procesu anterogradnog transporta sinaptičkih vezikula, mitohondrija i supstanci za izgradnju membrane, a obezbeđuju i retrogradni transport „markerskih“ molekula i uništenih organela.
Postoje tri tipa hemijskih interneuronskih interakcija: sinaptičke (npr. glutamatergične), ekstrasinaptičke (peptidergijske) i difuzne (npr. monoaminergičke, serotonergičke).
Hemijske sinapse se prema njihovoj anatomskoj strukturi dijele na aksodendritske, aksosomatske, aksoaksonalne i dendro-dendritske. Sinapsu predstavljaju pre- i postsinaptičke membrane, sinaptički rascjep i subsinaptička aktivna zona.
Električne sinapse obezbeđuju istovremenu aktivaciju čitavih grupa, formirajući električne veze između njih zahvaljujući kontaktima nalik prazninama (neksusima).
Aksoni glutamatergičnih (1) i dopaminergičkih (2) neurona formiraju čvrste sinaptičke kontakte sa procesom zvezdastog neurona (3) striatuma.
Dopamin se oslobađa ne samo iz presinaptičke regije, već i iz varikoznog zadebljanja aksona, odakle difundira u međućelijski prostor i aktivira dopaminske receptore dendritičnog stabla i zidova kapilarnih pericita.
 Dezinhibicija.
Dezinhibicija. (A) Ekscitatorni neuron 1 aktivira inhibitorni neuron 2, koji zauzvrat inhibira neuron 3.
(B) Pojava drugog inhibitornog neurona (2b) ima suprotan efekat na neuron 3, pošto je neuron 2b inhibiran.
Spontano aktivni neuron 3 generira signale u odsustvu inhibicijskih utjecaja.
2. Lijekovi - "ključevi" i "brave". Receptor se može uporediti sa bravom, a posrednik sa ključem koji mu odgovara. Ako je proces oslobađanja medijatora poremećen s godinama ili kao posljedica bilo koje bolesti, lijek može igrati ulogu “rezervnog ključa” koji obavlja funkciju sličnu medijatoru. Ovaj lijek se naziva agonist. Istovremeno, u slučaju prekomjerne proizvodnje, posrednika može "presresti" blokator receptora - "lažni ključ", koji će kontaktirati receptor "brave", ali neće uzrokovati njegovu aktivaciju.
3. Kočenje i dezinhibicija. Funkcionisanje spontano aktivnih neurona inhibirano je uticajem inhibitornih neurona (obično GABAergičnih). Aktivnost inhibitornih neurona, zauzvrat, može biti inhibirana drugim inhibitornim neuronima koji djeluju na njih, što rezultira dezinhibicijom ciljne stanice. Proces dezinhibicije je važna karakteristika neuronske aktivnosti u bazalnim ganglijama.
4. Rijetke vrste hemijskih sinapsi. Postoje dvije vrste aksoaksonalnih sinapsi. U oba slučaja, zadebljanje u obliku batine formira inhibitorni neuron. Sinapse prvog tipa formiraju se u području početnog segmenta aksona i prenose snažan inhibitorni efekat inhibitornog neurona. Sinapse drugog tipa formiraju se između zadebljanja u obliku štapa inhibitornog neurona i zadebljanja u obliku štapa ekscitatornih neurona, što dovodi do inhibicije oslobađanja transmitera. Ovaj proces se naziva presinaptička inhibicija. U tom smislu, tradicionalna sinapsa pruža postsinaptičku inhibiciju.
Dendro-dendritske (D-D) sinapse se formiraju između dendritskih bodlji dendrita susjednih bodljikavih neurona. Njihov zadatak nije da generišu nervni impuls, već da promene električni ton ciljne ćelije. U uzastopnim D-D sinapsama, sinaptičke vezikule se nalaze u samo jednoj dendritskoj kralježnici, au recipročnim D-D sinapsama u obje. Ekscitatorne D-D sinapse prikazane su na donjoj slici. Inhibitorne D-D sinapse su široko zastupljene u preklopnim jezgrima talamusa.
Osim toga, postoji nekoliko somato-dendritskih i somato-somatskih sinapsi.
 Aksoaksonalne sinapse moždane kore.
Aksoaksonalne sinapse moždane kore. Strelice pokazuju smjer impulsa.
 (1) presinaptička i (2) postsinaptička inhibicija spinalnog neurona koji putuje do mozga.
(1) presinaptička i (2) postsinaptička inhibicija spinalnog neurona koji putuje do mozga. Strelice pokazuju smjer provođenja impulsa (moguća je inhibicija prekidačkog neurona pod utjecajem inhibitornih utjecaja).
 Ekscitatorne dendro-dendritske sinapse. Prikazani su dendriti tri neurona.
Ekscitatorne dendro-dendritske sinapse. Prikazani su dendriti tri neurona. Recipročna sinapsa (desno). Strelice pokazuju smjer širenja elektrotoničnih valova.
Edukativni video - struktura sinapse
Sinapsa je određena zona kontakta između procesa nervnih ćelija i drugih ne-ekscitabilnih i ekscitabilnih ćelija koje osiguravaju prijenos informacijskog signala. Sinapsa je morfološki formirana kontaktnim membranama 2 ćelije. Membrana povezana s procesom naziva se presinaptička membrana stanice u koju se signal prima; njeno drugo ime je postsinaptička. Zajedno sa postsinaptičkom membranom, sinapsa može biti interneuronska, neuromuskularna i neurosekretorna. Riječ sinapsa uveo je 1897. Charles Sherington (engleski fiziolog).
Šta je sinapsa?
Sinapsa je posebna struktura koja osigurava prijenos nervnog impulsa sa nervnog vlakna na drugo nervno vlakno ili nervnu ćeliju, a kako bi na nervno vlakno delovala receptorna ćelija (područje kontakta između nervnih ćelija i još jedno nervno vlakno), potrebne su dvije nervne ćelije.
Sinapsa je mali dio na kraju neurona. Uz njegovu pomoć, informacije se prenose s prvog neurona na drugi. Sinapsa se nalazi u tri područja nervnih ćelija. Takođe, sinapse se nalaze na mestu gde nervna ćelija ulazi u vezu sa različitim žlezdama ili mišićima tela.
Od čega se sastoji sinapsa?
Struktura sinapse ima jednostavan dijagram. Sastoji se od 3 dijela, od kojih svaki obavlja određene funkcije tokom prijenosa informacija. Stoga se ova struktura sinapse može nazvati pogodnom za prijenos.Na proces direktno utiču dvije glavne ćelije: primajuća i odašiljajuća. Na kraju aksona prenosive ćelije nalazi se presinaptički završetak (početni dio sinapse). Može uticati na pokretanje neurotransmitera u ćeliji (ova riječ ima nekoliko značenja: posrednici, posrednici ili neurotransmiteri) - definira se po kojem se ostvaruje prijenos električnog signala između 2 neurona.

Sinaptički rascjep je srednji dio sinapse - ovo je jaz između 2 nervne ćelije koje međusobno djeluju. Kroz ovaj razmak dolazi električni impuls iz ćelije za prijenos. Završnim dijelom sinapse smatra se receptivni dio ćelije, koji je postsinaptički završetak (fragment ćelije u kontaktu sa različitim osjetljivim receptorima u svojoj strukturi).
Posrednici sinapse
Posrednik (od latinskog Media - prenosilac, posrednik ili sredina). Takvi sinaptički posrednici su veoma važni u procesu transmisije
Morfološka razlika između inhibitornih i ekscitatornih sinapsi je u tome što one nemaju mehanizam za oslobađanje transmitera. Odašiljačem u inhibitornoj sinapsi, motornom neuronu i drugim inhibitornim sinapsama smatra se aminokiselina glicin. Ali inhibitorna ili ekscitatorna priroda sinapse nije određena njihovim medijatorima, već svojstvom postsinaptičke membrane. Na primjer, acetilholin djeluje stimulativno na završetke neuromuskularnih sinapsa (vagusni nervi u miokardu).
Acetilholin služi kao ekscitatorni transmiter u kolinergičkim sinapsama (presinaptičku membranu u njoj igra završetak kičmene moždine motornog neurona), u sinapsi na Renshawovim stanicama, u presinaptičkom terminalu znojnih žlijezda, meduli nadbubrežne žlijezde, u intestinalnoj sinapsi i u ganglijama simpatičkog nervnog sistema. Acetilkolinesteraza i acetilholin su također pronađeni u frakcijama različitih dijelova mozga, ponekad u velike količine, ali osim kolinergičke sinapse na Renshawovim stanicama, još uvijek nisu uspjeli identificirati druge kolinergičke sinapse. Prema naučnicima, vrlo je vjerovatna medijatorska ekscitatorna funkcija acetilholina u centralnom nervnom sistemu.

Katelhomini (dopamin, norepinefrin i epinefrin) se smatraju adrenergičnim medijatorima. Adrenalin i norepinefrin se sintetiziraju na kraju simpatičkog živca, u moždanoj ćeliji nadbubrežne žlijezde, kičmene moždine i mozga. Aminokiseline (tirozin i L-fenilalanin) se smatraju polaznim materijalom, a adrenalin je konačni proizvod sinteze. Međusupstanca, koja uključuje norepinefrin i dopamin, također funkcionira kao posrednici u sinapsi stvorenoj na završecima simpatičkih živaca. Ova funkcija može biti inhibitorna (sekretorne žlijezde crijeva, nekoliko sfinktera i glatki mišići bronha i crijeva) ili ekscitacijska (glatki mišići određenih sfinktera i krvnih žila, u sinapsi miokarda - norepinefrin, u potkožnim jezgrama mozga - dopamin).
Kada medijatori sinapse završe svoju funkciju, presinaptički živčani završetak apsorbira kateholamin i aktivira se transmembranski transport. Tokom apsorpcije predajnika, sinapse su zaštićene od preranog iscrpljivanja zaliha tokom dugog i ritmičnog rada.
Synapse: glavni tipovi i funkcije
Langley je 1892. sugerirao da sinaptički prijenos u autonomnom gangliju sisara ne elektricne prirode, ali hemijski. Deset godina kasnije, Elliott je otkrio da se adrenalin proizvodi iz nadbubrežnih žlijezda istim djelovanjem kao i stimulacija simpatičkih živaca.

Nakon toga, sugerirano je da se adrenalin može lučiti od strane neurona i, kada je uzbuđen, oslobađa ga nervni završetak. Ali 1921. godine, Levy je napravio eksperiment u kojem je ustanovio hemijsku prirodu transmisije u autonomnoj sinapsi između srca i vagusnih nerava. Napunio je sudove fiziološkim rastvorom i stimulisao vagusni nerv, uzrokujući usporavanje rada srca. Kada je tečnost prenesena iz srca sa inhibicijom pejsinga u srce bez pejsinga, ono je kucalo sporije. Jasno je da je stimulacija vagusnog živca izazvala oslobađanje inhibitorne tvari u otopinu. Acetilholin je u potpunosti ponovio učinak ove tvari. Godine 1930. Feldberg i njegov saradnik konačno su utvrdili ulogu acetilholina u sinaptičkom prijenosu u ganglijima.
Hemijska sinapsa
Hemijska sinapsa se bitno razlikuje u prijenosu iritacije uz pomoć transmitera iz presinapse u postsinapsu. Zbog toga nastaju razlike u morfologiji hemijske sinapse. Hemijska sinapsa je češća u vertebralnom CNS-u. Sada je poznato da je neuron sposoban osloboditi i sintetizirati par transmitera (koegzistirajući prijenosnici). Neuroni takođe imaju plastičnost neurotransmitera - sposobnost da menjaju glavni transmiter tokom razvoja.

Neuromuskularni spoj
Ova sinapsa prenosi ekscitaciju, ali tu vezu mogu uništiti različiti faktori. Transmisija se završava blokadom oslobađanja acetilholina u sinaptičku pukotinu, kao i viškom njegovog sadržaja u području postsinaptičkih membrana. Mnogi otrovi i lijekovi utječu na unos, izlaz, koji je povezan s holinergičkim receptorima postsinaptičke membrane, zatim mišićna sinapsa blokira prijenos ekscitacije. Tijelo umire prilikom gušenja i zaustavljanja kontrakcije respiratornih mišića.

Botulinus je mikrobni toksin u sinapsi; blokira prijenos ekscitacije uništavanjem proteina sintaksina u presinaptičkom terminalu, koji je kontroliran oslobađanjem acetilkolina u sinaptičku pukotinu. Nekoliko toksičnih ratnih agenasa, farmakološki lijekovi (neostigmin i prozerin), kao i insekticidi blokiraju provođenje ekscitacije u neuromuskularnoj sinapsi inaktivirajući acetilkolinesterazu, enzim koji uništava acetilkolin. Stoga se acetilholin akumulira u području postsinaptičke membrane, smanjuje se osjetljivost na medijator, a blok receptora se oslobađa iz postsinaptičke membrane i uranja u citosol. Acetilholin će biti neefikasan i sinapsa će biti blokirana.
Nervna sinapsa: karakteristike i komponente
Sinapsa je veza između kontaktne tačke između dvije ćelije. Štaviše, svaki od njih je zatvoren u vlastitu elektrogenu membranu. Nervna sinapsa se sastoji od tri glavne komponente: postsinaptičke membrane, sinaptičke pukotine i presinaptičke membrane. Postsinaptička membrana je nervni završetak koji prolazi do mišića i spušta se u mišićno tkivo. U presinaptičkoj regiji nalaze se vezikule - to su zatvorene šupljine koje sadrže transmiter. Uvek su u pokretu.

Približavajući se membrani nervnih završetaka, vezikule se spajaju s njom, a transmiter ulazi u sinaptičku pukotinu. Jedna vezikula sadrži kvant medijatora i mitohondrije (potrebni su za sintezu medijatora - glavnog izvora energije), zatim se iz holina sintetizira acetilkolin i pod utjecajem enzima acetilkolin transferaze prerađuje u acetilCoA. .
Sinaptički rascjep među post- i presinaptičkim membranama
Veličina jaza je različita u različitim sinapsama. ispunjena međućelijskom tečnošću koja sadrži posrednik. Postsinaptička membrana pokriva mjesto kontakta između nervnog završetka i inervirane ćelije u mioneuralnoj sinapsi. U određenim sinapsama postsinaptička membrana se nabora i površina kontakta se povećava.
Dodatne supstance koje čine postsinaptičku membranu
U zoni postsinaptičke membrane prisutne su sljedeće tvari:
Receptor (holinergički receptor u mioneuralnoj sinapsi).
Lipoprotein (veoma sličan acetilkolinu). Ovaj protein ima elektrofilni kraj i ionsku glavu. Glava ulazi u sinaptički rascjep i stupa u interakciju s kationskom glavom acetilholina. Zbog ove interakcije dolazi do promjene postsinaptičke membrane, zatim dolazi do depolarizacije i otvaraju se potencijalno vođeni Na kanali. Depolarizacija membrane se ne smatra samopojačavajućim procesom;
Postepeno je, njegov potencijal na postsinaptičkoj membrani ovisi o broju medijatora, odnosno potencijal je karakteriziran svojstvom lokalne ekscitacije.
Kolinesteraza se smatra proteinom koji ima enzimsku funkciju. Po strukturi je sličan holinergičkom receptoru i ima slična svojstva kao acetilkolin. Kolinesteraza uništava acetilholin, prvo onaj koji je povezan sa holinergičkim receptorom. Pod dejstvom holinesteraze, holinergički receptor uklanja acetilkolin, što dovodi do repolarizacije postsinaptičke membrane. Acetilholin se razgrađuje na sirćetna kiselina i holin, neophodan za trofizam mišićnog tkiva.
Uz pomoć aktivnog transporta, kolin se uklanja u presinaptičku membranu, koristi se za sintezu novog transmitera. Pod uticajem medijatora menja se permeabilnost u postsinaptičkoj membrani, a pod uticajem holinesteraze osetljivost i permeabilnost se vraća na početnu vrednost. Hemoreceptori mogu stupiti u interakciju s novim medijatorima.
Predavanje 2. Fiziologija sinapsi: struktura, klasifikacija i mehanizmi djelovanja. Medijatori, neurohemijska osnova ponašanja.
Krajem 19. vijeka postojale su dvije paralelne teorije organizacije nervnog sistema (NS). Retikularna teorija vjeruje da je NS funkcionalni sincicij: neuroni su povezani procesima, sličnim kapilarama cirkulacijskog sistema. Prema Waldeyerova ćelijska teorija(1981) NS se sastoji od pojedinačnih neurona odvojenih membranama. Da bi se riješilo pitanje interakcije između pojedinačnih neurona, Sherington 1987. predložio je prisustvo posebne membranske formacije - sinapse. Koristeći elektronski mikroskop, prisustvo sinapsi je nedvosmisleno potvrđeno. Međutim, ćelijska teorija strukture NS-a postala je općeprihvaćena; ironično, Fershpan i Potter su 1959. godine otkrili sinapsu sa praznim spojevima (električna sinapsa) u NS-u rakova.
Synapse je membranska formacija od dvije (ili više) ćelija u kojoj se ekscitacija (informacija) prenosi s jedne ćelije na drugu.
Postoji sljedeća klasifikacija sinapsi:
1) po mehanizmu prijenosa pobude (i po strukturi):
Hemijski;
Električni (ephaps);
Miješano.
2) prema oslobođenom neurotransmiteru:
Adrenergički – neurotransmiter norepinefrin;
Holinergik – neurotransmiter acetilholin;
Dopaminergički – neurotransmiter dopamin;
Serotonergički – neurotransmiter serotonin;
GABAergic – neurotransmiter gama-aminobutirna kiselina (GABA)
3) uticajem:
uzbudljivo;
Kočnice.
4) po lokaciji:
Neuromuscular;
Neuro-neuralni:
a) akso-somatski;
b) akso-aksonalni;
c) akso-dendritski;
d) dendrosomatski.
Razmotrimo tri vrste sinapsi: hemijski, električni i mješoviti(kombinacija svojstava hemijskih i električnih sinapsi).
Bez obzira na vrstu, sinapse imaju zajedničke strukturne karakteristike: nervni proces na kraju formira produžetak ( sinaptički plak, SB); terminalna membrana SB razlikuje se od ostalih dijelova neuronske membrane i naziva se presinaptička membrana(PreSM); specijalizovana membrana druge ćelije je označena kao postsinaptička membrana (PostSM); nalazi između membrana sinapse sinaptički rascjep(SCH, sl. 1, 2).
Rice. 1. Šema strukture hemijske sinapse
Električne sinapse(ephapses, ES) se danas nalaze u NS ne samo rakova, već i mekušaca, člankonožaca i sisara. ES ima niz jedinstvenih svojstava. Imaju uski sinaptički rascjep (oko 2-4 nm), zbog čega se ekscitacija može prenijeti elektrokemijski (kao kroz nervno vlakno zbog EMF-a) velikom brzinom iu oba smjera: i od PreSM membrane do PostSM, i od PostSM do PreSM. Između ćelija postoje praznine (koneksoni ili koneksoni), formirani od dva proteina nazvana koneksini. Šest podjedinica svakog konneksina formira PreSM i PostSM kanale, kroz koje ćelije mogu razmjenjivati niskomolekularne tvari molekularne težine od 1000-2000 Daltona. Rad koneksona može se regulisati jonima Ca 2+ (slika 2).

Rice. 2. Dijagram električne sinapse
ES imaju veću specijalizaciju u poređenju sa hemijskim sinapsama i obezbeđuju veliku brzinu prenosa pobude. Međutim, čini se da je lišen mogućnosti suptilnije analize (regulacije) prenesenih informacija.
Hemijske sinapse dominiraju NS. Povijest njihovog proučavanja počinje radovima Claudea Bernarda, koji je 1850. objavio članak “Istraživanje o Curareu”. Evo šta je napisao: “Kurare je jak otrov koji pripremaju neki narodi (uglavnom kanibali) koji žive u šumama... Amazonije.” I dalje, „Curare je sličan zmijskom otrovu po tome što se može nekažnjeno uneti u probavni trakt ljudi ili životinja, dok injekcija pod kožu ili u bilo koji dio tijela brzo dovodi do smrti. ...nakon nekoliko trenutaka životinje leže kao da su umorne. Tada prestaje disanje i njihova osjetljivost i život nestaju, a da životinje ne kriju ili ne pokazuju znakove bola.” Iako C. Bernard nije došao na ideju kemijskog prijenosa nervnih impulsa, njegovi klasični eksperimenti s kurareom omogućili su da se ova ideja pojavi. Prošlo je više od pola veka kada je J. Langley ustanovio (1906) da je paralizirajući efekat kurarea povezan sa posebnim delom mišića, koji je nazvao receptivnom supstancom. Prvu sugestiju o prenošenju ekscitacije sa nerva na efektorski organ pomoću hemijske supstance dao je T. Eliot (1904).
Međutim, samo su radovi G. Dalea i O. Löwyja konačno odobrili hipotezu o hemijskoj sinapsi. Dale je 1914. ustanovio da se iritacija parasimpatičkog živca imitira acetilkolinom. Löwy je 1921. dokazao da se acetilholin oslobađa iz nervnog završetka vagusnog živca, a 1926. otkrio je acetilkolinesterazu, enzim koji uništava acetilholin.
Ekscitacija u hemijskoj sinapsi se prenosi pomoću posrednik. Ovaj proces uključuje nekoliko faza. Razmotrimo ove karakteristike na primeru acetilholinske sinapse, koja je rasprostranjena u centralnom nervnom sistemu, autonomnom i perifernom nervnom sistemu (slika 3).

Rice. 3. Šema funkcionisanja hemijske sinapse
1. Posrednik acetilholin (ACh) se sintetiše u sinaptičkom plaku iz acetil-CoA (acetil-koenzim A nastaje u mitohondrijima) i holina (sintetizira se u jetri) pomoću acetilkolin transferaze (slika 3, 1).
2. Pijuk je zapakovan sinaptičke vezikule ( Castillo, Katz; 1955). Količina medijatora u jednoj vezikuli je nekoliko hiljada molekula ( kvantni posrednik). Neki od vezikula se nalaze na PreSM-u i spremni su za oslobađanje medijatora (sl. 3, 2).
3. Posrednika pušta egzocitoza nakon pobuđivanja PreSM-a. Dolazna struja igra važnu ulogu u rupturi membrane i kvantnom oslobađanju odašiljača. Sa 2+ (sl. 3, 3).
4. Pušten izbor vezuje se za specifični receptorski protein PostSM (sl. 3, 4).
5. Kao rezultat interakcije između posrednika i receptora promjene jonske provodljivosti PostSM: kada se otvore Na+ kanali, depolarizacija; otvaranje K+ ili Cl - kanala dovodi do hiperpolarizacija(sl. 3, 5).
6 . Nakon depolarizacije, u postsinaptičkoj citoplazmi pokreću se biohemijski procesi (sl. 3, 6).
7. Receptor se oslobađa od medijatora: ACh se uništava acetilholinesterazom (AChE, sl. 3. 7).
Početak forme
Imajte na umu da medijator normalno stupa u interakciju sa specifičnim receptorom određene snage i trajanja. Zašto je curare otrov? Mjesto djelovanja kurarea je upravo ACh sinapsa. Curare se čvršće vezuje za acetilkolinski receptor i lišava ga interakcije s neurotransmiterom (ACh). Ekscitacija od somatskih živaca do skeletnih mišića, uključujući i od freničnog živca do glavnog respiratornog mišića (dijafragme) prenosi se uz pomoć ACh, pa kurare uzrokuje opuštanje mišića i prestanak disanja (što, zapravo, uzrokuje smrt).
Zabilježimo glavno Osobine prijenosa ekscitacije u kemijskoj sinapsi.
1. Ekscitacija se prenosi pomoću hemijskog posrednika - posrednika.
2. Ekscitacija se prenosi u jednom smjeru: od PreSm do PostSm.
3. Nastaje hemijska sinapsa privremeno odlaganje u provođenju ekscitacije, dakle sinapsa ima niska labilnost.
4. Hemijska sinapsa je vrlo osjetljiva na djelovanje ne samo medijatora, već i drugih biološki aktivnih supstanci, lijekova i otrova.
5. U hemijskoj sinapsi dolazi do transformacije ekscitacije: elektrohemijska priroda ekscitacije na PreSM nastavlja se u biohemijski proces egzocitoze sinaptičkih vezikula i vezivanja medijatora za određeni receptor. Nakon toga slijedi promjena ionske provodljivosti PostSM-a (također elektrohemijski proces), koji se nastavlja biokemijskim reakcijama u postsinaptičkoj citoplazmi.
U principu, takav višestepeni prijenos pobude trebao bi biti značajan biološki značaj. Imajte na umu da je u svakoj fazi moguće regulirati proces prijenosa pobude. Uprkos ograničenom broju medijatora (nešto više od desetak), u hemijskoj sinapsi postoje uslovi za široku raznolikost u odlučivanju o sudbini nervnog pobuđenja koji dolazi u sinapsu. Kombinacija karakteristika hemijskih sinapsi objašnjava individualnu biohemijsku raznolikost nervnih i mentalnih procesa.
Zaustavimo se sada na dva važna procesa koji se dešavaju u postsinaptičkom prostoru. Primijetili smo da se kao rezultat interakcije ACh sa receptorom na PostSM-u mogu razviti i depolarizacija i hiperpolarizacija. Šta određuje da li će medijator biti ekscitatorni ili inhibitorni? Rezultat interakcije između posrednika i receptora određena svojstvima receptorskog proteina(još jedno važno svojstvo hemijske sinapse je da je PostSM aktivan u odnosu na ekscitaciju koja dolazi do njega). U principu, hemijska sinapsa je dinamička formacija; promjenom receptora, stanica koja prima pobudu može utjecati na nju buduća sudbina. Ako su svojstva receptora takva da njegova interakcija sa transmiterom otvara Na + kanale, onda kada izolacijom jednog kvanta medijatora na PostSM-u razvija se lokalni potencijal(za neuromuskularni spoj naziva se minijaturni potencijal krajnje ploče - MEPP).
Kada se javlja PD? PostSM ekscitacija (ekscitatorni postsinaptički potencijal - EPSP) nastaje kao rezultat sumiranja lokalnih potencijala. Možete odabrati dvije vrste procesa sumiranja. At sekvencijalno oslobađanje nekoliko kvanta medijatora u istoj sinapsi(voda nosi kamen) nastaje privremeniA Ja sam sumiranje. Ako kvanti medijatori se oslobađaju istovremeno na različitim sinapsama(na membrani neurona ih može biti nekoliko hiljada). prostorna sumacija. Repolarizacija PostSM membrane se odvija sporo i nakon oslobađanja pojedinačnih kvanta medijatora, PostSM je neko vrijeme u stanju egzaltacije (tzv. sinaptička potencijacija, sl. 4). Možda na taj način dolazi do treninga sinapse (oslobađanje kvanta odašiljača u određenim sinapsama može „pripremiti“ membranu za odlučujuću interakciju sa transmiterom).
Kada se K+ ili Cl - kanali otvore na PostSM-u, pojavljuje se inhibitorni postsinaptički potencijal (IPSP, slika 4).

Rice. 4. Postsinaptički membranski potencijali
Naravno, ako se IPSP razvije, dalje širenje ekscitacije može se zaustaviti. Druga opcija za zaustavljanje procesa ekscitacije je presinaptička inhibicija. Ako se na membrani sinaptičkog plaka formira inhibitorna sinapsa, tada kao rezultat hiperpolarizacije PreSM-a može biti blokirana egzocitoza sinaptičkih vezikula.
Drugi važan proces je razvoj biohemijskih reakcija u postsinaptičkoj citoplazmi. Promjena jonske provodljivosti PostSM-a aktivira tzv sekundarni glasnici (posrednici): cAMP, cGMP, Ca 2+-zavisna protein kinaza, koje zauzvrat aktiviraju različite protein kinaze tako što ih fosforiliraju. Ove biohemijske reakcije mogu se „spustiti“ duboko u citoplazmu sve do jezgra neurona, regulišući procese sinteze proteina. Dakle, nervna ćelija može odgovoriti na dolazeću ekscitaciju ne samo odlučivanjem o svojoj daljnjoj sudbini (odgovarati EPSP ili IPSP, tj. provoditi ili ne nastaviti dalje), već i promijeniti broj receptora, ili sintetizirati receptorski protein s novim svojstva u odnosu na određenog na posrednika. Shodno tome, još jedno važno svojstvo hemijske sinapse: zahvaljujući biohemijskim procesima postsinaptičke citoplazme, ćelija se priprema (uči) za buduće interakcije.
U nervnom sistemu funkcionišu različite sinapse koje se razlikuju po medijatorima i receptorima. Ime sinapse određuje medijator, tačnije, naziv receptora za određeni posrednik. Stoga, razmotrimo klasifikaciju glavnih medijatora i receptora nervnog sistema (pogledajte i materijal koji je podijeljen na predavanju!!).
Već smo primijetili da je učinak interakcije između medijatora i receptora određen svojstvima receptora. Stoga poznati medijatori, sa izuzetkom g-aminomaslačne kiseline, mogu obavljati funkcije i ekscitatornih i inhibitornih medijatora. hemijska struktura Razlikuju se sljedeće grupe medijatora.
Acetilholin, široko rasprostranjen u centralnom nervnom sistemu, posrednik je u holinergičkim sinapsama autonomnog nervnog sistema, kao iu somatskim neuromuskularnim sinapsama (slika 5).

Rice. 5. Molekul acetilholina
Poznato dva tipa holinergičkih receptora: nikotin ( H-holinergički receptori) i muskarinici ( M-holinergički receptori). Ime su dale supstance koje u ovim sinapsama izazivaju efekat sličan acetilkolinu: N-holinomimetik je nikotin, A M-holinomimetik- toksin muharice Amanita muscaria ( muskarin). Blokator H-holinergičkih receptora (antiholinergik) je d-tubokurarin(glavna komponenta otrova kurare), i M-antiholinergik je toksin beladone Atropa belladonna – atropin. Zanimljivo je da su svojstva atropina odavno poznata i bilo je vremena kada su žene koristile atropin iz beladone da izazovu proširenje vidnih zjenica (da oči budu tamne i „lijepe“).
Sljedeća četiri glavna medijatora imaju sličnosti u hemijskoj strukturi, pa su klasifikovani kao monoamini. Ovo serotonin ili 5-hidroksitriptamin (5-HT), igra važnu ulogu u mehanizmima pojačanja (hormon radosti). Sintetizira se iz esencijalne aminokiseline za ljude - triptofana (slika 6).

Rice. 6. Molekul serotonina (5-hidroksitriptamin).
Tri druga medijatora sintetiziraju se iz esencijalne aminokiseline fenilalanina, pa su stoga ujedinjeni pod zajedničkim imenom kateholamini- Ovo dopamin (dopamin), norepinefrin (norepinefrin) i adrenalin (epinefrin, slika 7).

Rice. 7. Kateholamini
Među amino kiseline medijatori uključuju gama-aminobutirna kiselina(g-AMK ili GABA - poznat kao jedini inhibitorni neurotransmiter), glicin, glutaminska kiselina, asparaginska kiselina.
Medijatori uključuju brojne peptidi. Godine 1931. Euler je otkrio tvar u ekstraktima mozga i crijeva koja uzrokuje kontrakciju glatkih mišića crijeva i proširenje krvnih žila. Ovaj transmiter je izoliran u svom čistom obliku iz hipotalamusa i dobio je ime supstanca P(od engleskog praha - prah, sastoji se od 11 aminokiselina). Kasnije je ustanovljeno da supstanca P igra važnu ulogu u provođenju bolnih ekscitacija (naziv nije morao mijenjati, jer je bol na engleskom bol).
Delta peptid spavanja dobio je ime po svojoj sposobnosti da izaziva spore ritmove velike amplitude (delta ritmove) u elektroencefalogramu.
U mozgu se sintetizira niz proteinskih medijatora narkotičke (opijatske) prirode. Ovo su pentapeptidi Met-enkefalin I Leu-enkefalin, i endorfini. Ovo su najvažniji blokatori ekscitacije bola i posrednici pojačanja (radosti i zadovoljstva). Drugim riječima, naš mozak je velika fabrika endogeni droge. Glavna stvar je naučiti mozak da ih proizvodi. "Kako?" - pitate. Jednostavno je - endogeni opijati nastaju kada doživimo zadovoljstvo. Radite sve sa zadovoljstvom, natjerajte svoju endogenu fabriku da sintetiše opijate! Prirodno nam je data ova prilika od rođenja - velika većina neurona reaguje na pozitivno pojačanje.
Istraživanja posljednjih decenija omogućila su otkrivanje još jednog vrlo zanimljivog posrednika - dušikov oksid (NO). Pokazalo se da NO ne samo da igra važnu ulogu u regulaciji tonusa krvnih sudova (nitroglicerin za koji znate da je izvor NO i širi koronarne sudove), već se sintetiše i u neuronima centralnog nervnog sistema.
U principu, istorija medijatora još nije gotova, postoji niz supstanci koje su uključene u regulaciju nervnog uzbuđenja. Samo činjenica njihove sinteze u neuronima još nije precizno utvrđena, nisu pronađeni u sinaptičkim vezikulama, a specifični za njih receptori nisu pronađeni.







