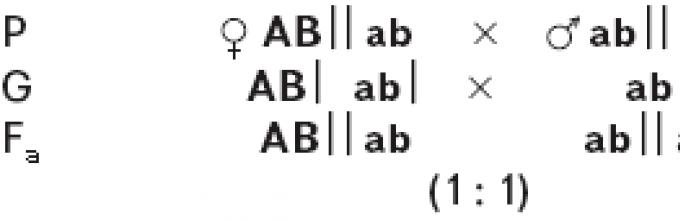
32. Peculiarities of inheritance of a trait with incomplete and complete coupling
LINKED INHERITANCE OF CHARACTERISTICS
We became acquainted with dihybrid crossing and realized that the independent combination of traits is explained by the fact that the splitting of one pair of allelic genes that determine the corresponding traits occurs independently of the other pair. However, this is observed only in the case when the genes of different pairs are located in different pairs of chromosomes and when the hybrid germ cells are formed in meiosis, the paternal and maternal chromosomes are independently combined. But the number of chromosomes is very limited compared to the number of characters, each of which develops under the control of a specific gene. Thus, about 7000 genes are known in Drosophila with four pairs of chromosomes. It is assumed that a person has at least 50 thousand genes with 23 pairs of chromosomes, etc. It follows that hundreds of alleles must be localized in each pair of chromosomes. Naturally; that there is linkage between genes that are located on the same chromosome and when germ cells are formed, they must be transmitted together.
Linked inheritance was discovered in 1906 by English geneticists W. Batson and R. Punnett while studying the inheritance of traits in sweet peas, but they were unable to reveal the reasons for this phenomenon. The nature of linked inheritance was clarified in 1910 by scientists T. Morgan and his collaborators K. Bridges and A. Sturtevant. As an object of study, they chose the fruit fly Drosophila, which turned out to be very convenient for genetic experiments. The cells of the Drosophila body contain 4 pairs of chromosomes. It is distinguished by very high fertility - one pair produces more than a hundred offspring. It has a high development speed - within 12-15 days after fertilization, a larva, pupa and an adult develop from the egg, which is almost immediately capable of producing offspring. More than twenty generations can be studied over the course of a year. Flies are gray in color, with red eyes. are small in size (about 3 mm), easily diluted in biological test tubes; You can use magnifying glasses to study their signs. When looking at hundreds of thousands of individuals, Morgan discovered many different mutations: there were flies with black and yellow bodies, with white and other colored eyes, with altered shape and position of wings, etc. Sometimes there were individuals with several mutations at once, for example black body, rudimentary wings, cinnabar eyes.
Studying the inheritance of different pairs of characters during dihybrid and polyhybrid crossings, Morgan and his colleagues discovered a large number of examples of their linked (joint) inheritance. All studied characters were divided into four linkage groups in accordance with the number and size of chromosomes in Drosophila. On this basis, Morgan concluded that the genes that determine these traits are located on chromosomes. Genes located on the same chromosome constitute a linkage group.
Gene linkage is the joint inheritance of genes located on the same chromosome. The number of linkage groups corresponds to the haploid number of chromosomes. For example, Drosophila has 4 linkage groups, humans have 23, cattle have 30, pigs have 19, etc.
The idea of the location of genes in chromosomes was expressed by Setton back in 1902. He discovered parallelism in the behavior of chromosomes in meiosis and the inheritance of traits in one of the grasshopper species. Further research conducted by Morgan showed that the linkage of genes located on the same chromosome can be complete or incomplete.
The difference in the behavior of linked and independently inherited genes is most clearly revealed when conducting an analytical cross. With independent inheritance of two pairs of characters in a hybrid Fi (АаББ) 4 types of gametes are formed with equal probability: AB,Ab, aB, ab. When crossed with a complete recessive (aabb) the number of varieties of gametes in a hybrid determines the number of types of descendants and the same probability of their appearance, since the gametes of a recessive individual (ab) cannot change the manifestations of dominant and recessive genes of the hybrid gametes. As a result, the ratio of offspring phenotypes will be 1:1:1:1. If both pairs of allelic genes are located in one pair of chromosomes, then during the formation of germ cells the genes of these alleles will not be able to combine freely. In this case, linked inheritance is observed.
FULL CLUTCH
T. Morgan crossed black long-winged females with gray males with rudimentary wings. In Drosophila, gray body color dominates over black, and long wings dominate over rudimentary wings. Let us denote the gray body color gene IN, allelic gene for black body color b; Longwing™ gene V, allelic to it is the gene for rudimentary wings v. Both pairs of these genes are located on the same second pair of chromosomes. For both pairs of Traits, the parental forms were homozygous: female for the recessive trait of black body (bb) and the dominant trait long-winged™ (V.V.), male by dominant gray coloration. (BB) and the recessive trait of rudimentary wings (w>. The gametes of the parents, during reduction division, receive from the maternal form a chromosome with genes b And V, from the father's - with genes IN and v. All offspring of the first generation (Fi) had a gray body and long wings (Fig. 13) and were heterozygous for both pairs of traits (bV/ Bv). Then males were selected from Fi and crossed with females homozygous for both recessive genes, black rudiments (bv/ bv), i.e., an analytical crossing was carried out, as a result of which, with an independent combination of characters, the offspring of four phenotypes should have been obtained in equal proportions: gray long-winged, gray with rudimentary wings, black long-winged, black with rudimentary wings, but the descendants of only two phenotypes were obtained , similar to the original parental forms: black long-winged and gray short-winged. In this case, complete concatenation of characteristics is observed. This is due to the fact that in a heterozygous male, both the gene for black coloration and the gene for long wings are located on the same chromosome from a homologous pair, and the gene for gray coloration and the gene for rudimentary wings are located on the other.
During spermiogenesis during meiosis, homologous chromosomes separate into different germ cells. Only two varieties of gametes are produced: one with a chromosome that carries genes Kommersant And V, the other with the chromosome on which the genes are located IN and v. When these gametes are combined with gametes of an individual with recessive traits, only two types of offspring are formed. In complete linkage, genes located on the same chromosome are always transmitted together. Complete linkage has so far been established only in fruit flies and female silkworms.
INCOMPLETE CLUTCH
In the next experiment, as in the previous one, Morgan crossed black long-winged females with gray rudimentary-winged males. In the first generation, all the offspring were gray long-winged. Then he again made an analytical cross, but from the first generation he selected not a male, but a female and crossed her with a black male with rudimentary wings (Fig. 14). In this case, the offspring appeared not of two types, as with complete linkage, but of four: gray with rudimentary wings, black long-winged, gray long-winged and black with rudimentary wings, but not in equal proportions, as with an independent combination of characters, but with a significant predominance phenotypes similar to the parental forms. 41.5% of the flies were gray with rudimentary wings, like one original parent, and 41.5% were black long-winged, like the other original parent. Only 17% of the offspring were born with a new combination of characteristics: 8.5% were black with rudimentary wings and 8.5% were gray long-winged. Thus, 83% of the descendants had a combination of characteristics like the original parental forms, but individuals also appeared with a new combination of characteristics. Therefore, the clutch is incomplete./ The question arose: why did individuals with a new combination of parental characteristics appear? To explain this phenomenon, Morgan used and developed the theory chiasmotypes Belgian cytologist Janssens. In 1909, Janssens observed that during spermiogenesis in a salamander in the prophase of meiosis, homologous chromosomes conjugate, and then, at the beginning of divergence, form figures in chromosomes exchange sections. If linked genes lie on the same chromosome and recombination of these genes occurs in heterozygotes during the formation of gametes, it means that homologous chromosomes exchanged their parts during meiosis. The exchange of homologous chromosomes with their parts is called decussation or cross-singover(the English word crossingover means the formation of a cross). Individuals with new combinations of characters resulting from crossing over are called crossovers.
Let's return to Figure 14. In female Fi, heterozygous for both pairs of traits, genes are located on one of the homologous chromosomes Kommersant And V, in the other - genes allelic to them IN and v. In the prophase of reduction division, when two homologous chromosomes are united into one bivalent, each chromosome is doubled and consists of two chromatids. There will be 4 chromatids in total. Between two chromatids of homologous chromosomes, an exchange of their parts occurs. As a result, the gene Kommersant, located in the chromatid of one homologous chromosome, can connect with the v gene located in the chromatid of another homologous chromosome, and as a result of one event, a second chromatid is formed, where the genes are connected IN And V. Subsequently, the chromatids will separate and crossover gametes with chromosomes with a new combination of genes will be formed. (bv And B.V.).
The other two chromatids from a pair of homologous chromosomes do not participate in the crossover and retain the maternal ones in the original combination (bV) and paternal (Bv) genes. The formation of new crossover gametes ensured the appearance of fruit flies with a new combination of characters: black with rudimentary wings and gray long-winged ones. However, most of the offspring will be similar to the original parents (black long-winged and gray short-winged). Morgan concludes that the number of appearances of new forms depends on the frequency of crossover, which is determined by the following formula:
If, for example, the total number of descendants is 900, and the new crossover forms are 180, then the crossover frequency will be 20 %. Morgan found that frequency of crossover between a specific pair of genes- a relatively constant value, but different for different pairs of genes. Based on this, it was concluded that The frequency of crossover can be used to judge the distances between genes. The unit of measurement of the cross is taken to be equal to 1%. Sometimes she is called Morganida. The amount of crossover depends on the distance between the genes being studied. The more distant the genes are from each other, the more often crossover occurs; the closer they are located, the less likely it is to cross. It has been established that the number of crossover individuals to the total number of descendants never exceeds 50%, since at very large distances between genes, double crossing over occurs more often and some crossover individuals remain unaccounted for. They can be taken into account when studying not two pairs of linked traits, but three or four. In this case, taking into account double and triple crossovers, it is possible to more accurately judge the distances and frequency of crossover between genes.
Block of information.
Linkage of genes.
At the beginning of the 20th century (1902-1907), the American scientist W. Setton and the German embryologist T. Boveri discovered parallelism in the inheritance of characteristics and the behavior of the chromosomes of the cell nucleus during the process of gametogenesis and fertilization. This confirmed the localization of hereditary information in chromosomes. It has been established that the number of genes significantly exceeds the number of chromosomes. So, a person has 46 chromosomes, and there are from 70,000 to 100,000 genes. Consequently, a large number of genes are localized on each chromosome. Genes on the same chromosome are inherited together (linked). An experimental study of this phenomenon was carried out by the American geneticist T. Morgan and his collaborators: A. Sturtevant, A. Bridges and G. Möller in 1910-1916. These studies confirmed the chromosomal localization of genes and formed the basis of the chromosomal theory of heredity.
Basic provisions of the chromosomal theory of Heredity.
1. Each gene occupies a specific place on the chromosome - a locus.
2. Genes on a chromosome are arranged linearly in a certain sequence.
3. Different chromosomes contain different numbers of genes. The set of genes on each non-homologous chromosome is unique.
4. Genes of one chromosome form a “linkage group and are inherited together, i.e. linked.
5. The number of linkage groups is equal to the number of chromosomes in the haploid set (there are four in Drosophila, 10 in corn, 20 in mice, 23 in humans).
6. An exchange of allelic genes, i.e. crossing over, can occur between homologous chromosomes.
7. The frequency of crossing over is directly proportional to the distance between genes in the linkage group.
8. A special unit, the morganide (M), is taken as the unit of distance between genes in a linkage group. 1M=1% crossing over.
There are complete and incomplete linkage of genes.
Full grip.
In experiments on Drosophila it was found that the development
traits that are inherited linked and controlled by genes of one
chromosomes. Genes for body color (b - gray and B - black) and wing length
(v - normal and V - short, vestigial wings) are localized in one pair of homologous chromosomes.
Crossing gray flies with normal wings and gray flies with vestigial wings produces gray hybrids with normal wings in the first generation.
When carrying out the analyzing crossing, males were selected from p, since it is known that male Drosophila have achiasmatic spermatogenesis (i.e., crossing over does not occur and the completeness of gene linkage is not disturbed in any way). As a result of such crossing, individuals of two phenotypes similar to the original parental forms, and in equal quantities: 1:1 phenotypic splitting.
Considered together, the results of both crossings convince us that the development of the analyzed traits is controlled by different genes, and linked inheritance is explained by the localization of genes on the same chromosome. The completeness of adhesion in this case is not disturbed by anything. This gene linkage is complete.
To study incomplete linkage from P, females (genotype B||b) were selected; crossing over occurs in C females during gametogenesis. Therefore, a diheterozygous individual forms additional, i.e. crossover varieties of gametes. The probability of their formation is determined by the probability of crossing over, i.e. depends on the distance between genes in the linkage group.
Not recombinant individuals; y Recombinant individuals; at
they are observed to have the same formation of their genotypes
combinations, like the original ones, involved crossover
parental forms of the gamete.
In this example, the phenotypic cleavage in the offspring was as follows: gray flies with long wings - 41.5%; black with short wings - 41.5%; gray shortwings - 8.5%; black longwings - 8.5%. Thus, the probability of the appearance of recombinant individuals in the offspring is 17%. Therefore, the distance between genes B and V in the linkage group is 17 morganiids.
The predominance of gray long-winged and black short-winged flies in the offspring indicates that genes B and V; b and v are indeed linked. On the other hand, the appearance of recombinant individuals indicates that in a certain number of cases there is a break in the linkage between genes B and V and genes B and V. This is the result of crossing over.
An example of complete linkage of genes in humans is the inheritance of the Rh factor. It is caused by three pairs C, D, K, closely linked to each other, therefore inheritance of Rhesus affiliation occurs according to the type of monohybrid crossing. Another example of the close linkage of genes in humans is the inheritance of cataracts and polydactyly. The genes for hemophilia and color blindness are localized on the X chromosome at a distance of 9.8 morganids (M), i.e. undergo crossing over and are therefore inherited as incompletely linked. Autosomal genes for the Rh factor and the shape of erythrocytes, located at a distance of 3 M from each other, are also an example of incomplete linkage.
In 1909, F. Janssens, while studying meiosis in amphibians, discovered chiasmata (crossovers) of chromosomes, which are cytological evidence of crossing over. Since that time, many attempts have been made to explain the mechanism of this phenomenon. There are several theories of crossing over. The most common are two hypotheses.
Lesson planning 10th grade
Subject: “Complete and incomplete linkage of genes. Genetic maps of chromosomes."
Objective of the lesson: familiarize students with the process of complete and incomplete gene linkage; and form an idea of the genetic maps of chromosomes.
Educational tasks:
Reveal the essence of the phenomenon of linked inheritance of genes.
To develop knowledge about the main provisions of the T. Morgan law.
Introduce the principle of compiling genetic maps
Develop students' logical thinking.
Equipment, visual aids : tables on general biology illustrating the linked inheritance of genes and traits, presentation for the lesson, tasks to reinforce new material.
Lesson type: A lesson in learning new material.
Methods: explanatory - illustrative.
Lesson progress:
I Organizational moment
Checking the personnel of students and the visual readiness of the class for the lesson.
II Testing students' knowledge
Frontal survey:
1. Name three laws of G. Mendel?
2. What rules did G. Mendel adhere to when conducting his experiments?
3. Formulate the law of gamete purity. Who is responsible for the discovery of this law?
4. Can traits always be clearly divided into dominant and recessive?
5. What name did this phenomenon get?
6. Is it always possible to determine from the phenotype which genes a given individual contains? Give an example.
7. Is it possible to establish the genotype of individuals that do not differ in phenotype? What method is used for this?
8. What features characterize dihybrid crossing?
Well done! You have completed this stage of work
III Learning new material:
Linked inheritance of genes
G. Mendel traced the inheritance of seven pairs of traits in peas. Many researchers, repeating Mendel's experiments, confirmed the laws he discovered. It was recognized that these laws were of a universal nature. However, in 1906, English geneticists W. Batson and R. Pennett, crossing sweet pea plants and analyzing the inheritance of pollen shape and flower color, discovered that these characters do not give independent distribution in the offspring. Descendants always repeated the characteristics of their parent forms. Gradually, evidence of exceptions to Mendel's third law accumulated. It became clear that not all genes are characterized by independent distribution in the offspring and free combination.
Any organism has a variety of morphological, physiological, biochemical and other characteristics and properties, and each characteristic or property is controlled by one or more genes localized in chromosomes.
However, if the number of genes in an organism is huge and can amount to tens of thousands, then the number of chromosomes is relatively small and, as a rule, is measured in several tens. Therefore, hundreds and thousands of allelic genes forming linkage groups are localized in each pair of chromosomes.
A complete correspondence has been established between the number of linkage groups and the number of chromosome pairs. For example, corn has a set of chromosomes 2n = 20 and 10 linkage groups, and Drosophila has 2n = 8 and 4 linkage groups, that is, the number of linkage groups is equal to the haploid set of chromosomes.
Thomas Morgan Law
Genes localized on the same chromosome are transmitted together, and their mode of inheritance differs from the inheritance of genes localized in different pairs of homologous chromosomes.
So, for example, with an independent distribution of chromosomes, a dihybridAaBb forms four types of gametes (AB , aB , Ab , ab ), and under the condition of complete linkage, the same dihybrid will give only two types of gametes (AB Andab ), since these genes are located on the same chromosome.
The development of the problem of linked inheritance of genes belongs to the school of T. Morgan (1866–1945). If Mendel conducted his experiments on peas, then for Morgan the main object was the fruit fly Drosophila. The fly produces numerous offspring every two weeks at a temperature of 25 °C. The male and female are clearly distinguishable - the male has a smaller and darker abdomen. In addition, they differ in numerous characteristics and can reproduce in test tubes on a cheap nutrient medium.
Studying patterns of inheritancegenes localized on the same chromosome , Morgan concluded that theyare inherited linked . This is T. Morgan's law.
Crossing over - process of exchange of plotsduring in prophase I . In addition to meiotic, it has also been described . The chromosome is divided into these sections at certain points, the same for one species, which can be the definition of the species at the genetic level; the location of these points is determined by a single gene.
Since crossing over introduces disturbances into the picture , it was successfully used to map “linkage groups” (chromosomes). The mapping feasibility was based on the assumption that the more often crossover occurs between two , the further these genes are located from each other in the linkage group and the more often deviations from linked inheritance will be observed.
Full and partial clutch
To determine the type of inheritance of two pairs of genes (linked or independent), it is necessary to conduct an analytical crossing and, based on its results, draw a conclusion about the nature of gene inheritance. Let's consider three possible options for the results of an analyzing cross.
1) Independent inheritance .
If, as a result of analytical crossing, four classes of phenotypes are formed among hybrids, it means that the genes are inherited independently.
2) Complete gene linkage .

With complete gene linkageA
AndIN
according to the results of analyzing crossing, they find -
There are two phenotypic classes of hybrids that completely copy their parents.
3) Incomplete gene linkage .

In case of incomplete linkage of genesA AndIN When analyzing crosses, four phenotypes appear, two of which have a new combination of genes:Ab ‖ ab ; aB ‖ ab . The appearance of such forms indicates that the dihybrid with gametesAB │ andab │ forms crossover gametesAb │ andaB │. The appearance of such gametes is possible only as a result of the exchange of sections of homologous chromosomes, that is, in the process of crossing over. The number of crossover gametes is significantly smaller than non-crossover gametes.
The crossover frequency is proportional to the distance between genes. The closer the genes are located on a chromosome, the closer the linkage between them and the less often they separate during crossover. And vice versa, the farther the genes are from each other, the weaker the linkage between them and the more often they cross over. Consequently, the distance between genes on chromosomes can be judged by the frequency of crossover.
Genetic maps
Genetic mapping usually means determining the position of a gene in relation to other genes.
Let's consider the procedure for compiling genetic maps.
1. Establishment of the linkage group (that is, determination of the chromosome in which the gene is localized). To do this, it is necessary to have at least one marker gene in each linkage group.
2. Finding the location of the gene under study in the chromosome. To do this, the mutant form is crossed with a normal one and the result of crossing over is taken into account.
3. Determination of the distance between linked genes, which makes it possible to draw up genetic maps of chromosomes, which indicate the order of genes in the chromosomes and their relative distances from each other. The higher the crossing over frequency, the greater the distance between the genes. If it is established that between linked genesA AndIN crossing over frequency is 10%, and between genesIN AndWITH – 20%, then it is obvious that the distanceSun 2 times more thanAB . The distance between genes is expressed in units corresponding to 1% crossing over. These units are called morganids.
Thus, based on data on the frequency of crossing over, genetic maps are compiled.
IV Consolidation of knowledge
Solving a genetic problem
A female Drosophila heterozygous for the recessive genes for dark body color and miniature wings was crossed with a male who had a dark body and miniature wings. From this crossing the following was obtained:
– 244 flies with dark bodies and miniature wings;
– 20 flies with gray body color and miniature wings;
– 15 flies with dark body color and normal wings;
– 216 flies with gray body color and normal wings.
Based on the given data, determine whether these two pairs of genes are linked or not. How are genes linked?
Given :
A
– gray body
A
– dark body
IN
– normal wings
b
– miniature wings
The nature of gene inheritanceA AndIN - ?
Solution
The results of splitting among hybrids (two phenotypic classes are dominant and repeat phenotypically and genotypically the parental forms, and the other two classes of phenotypes are represented by a small number of individuals) indicate incomplete linkage of genes§41, pp.161-164. Answer the questions p.165.
Task No. 1. A diheterozygous pea plant with smooth seeds and tendrils was crossed with a plant with wrinkled seeds without tendrils. It is known that both dominant genes (smooth seeds and the presence of tendrils) are localized on the same chromosome; crossing over does not occur. Make a diagram for solving the problem. Determine the genotypes of the parents, phenotypes and genotypes of the offspring, the ratio of individuals with different genotypes and phenotypes. What law is manifested in this case?
A - smooth seeds, a - wrinkled seeds
B - presence of antennae, b - without antennae
AB/ab
ab/ab
AB/ab
ab/ab
smooth
seeds,
mustache
wrinkled
seeds,
without mustache
50%
50%
If crossing over does not occur, then the diheterozygous parent produces only two types of gametes (full linkage).
Genes located on the same chromosome are clutch group. Gene linkage- This is the joint inheritance of genes located on the same chromosome. The number of linkage groups corresponds to the haploid number of chromosomes. The linkage of genes located on the same chromosome may be complete or incomplete. Full grip: Morgan crossed black long-winged females with gray males with rudimentary wings. In Drosophila, gray body color dominates over black, and long wings dominate over rudimentary wings. Gray body - A, black body a; long-winged - B, rudimentary wings - c. During spermiogenesis during meiosis, homologous chromosomes separate into different germ cells. 1) AA//AB*av//av=4AB//av; 2) AB//av*AB//av=AB//AB, AB//av, ab//AB, ab//av. If the genes are found in autosomes, then with complete linkage in F1 there will be uniformity in phenotype, and in F2 - 3:1, no matter how many characteristics the parents differ in, because one pair of chromosomes is studied.
The phenomenon of incomplete linkage in the inheritance of traits
As a result of crossing, the descendants had a combination of characteristics, like the original parental forms, but individuals also appeared with a new combination of characteristics - clutch incomplete. B – gray, c – black, V – normal, v – rudimentary. Bv||Bv*bV||bV=Bv||bV; females from the first generation were crossed with analyzer males: BV//bV*bv//bv=Bv//bv,bV//bv – not crossover. Bv//bV*bv//bv=2bv//bv, 2BV//bv – crossover. The exchange of homologous chromosomes with their parts is called crossover or crossing over. Individuals with new combinations of characters resulting from crossing over are called crossovers. The number of appearances of new forms depends on the crossover frequency, which is determined by the following formula: Crossover frequency = (Number of crossover forms) 100/ Total number of descendants. The unit of measurement of the cross is taken to be equal to 1%. She is called morganida. The amount of crossover depends on the distance between the genes being studied. The more distant the genes are from each other, the more often crossover occurs; the closer they are located, the less likely it is to cross.
Chromosome maps. An example of their construction
Chromosome map- plan of the location of genes on a chromosome. Genes are located on chromosomes in a linear sequence at certain distances from each other. The phenomenon of inhibition of crossing over in one area by crossing over in another is called interference. The smaller the distance separating the three genes, the greater the interference. Taking into account the linear arrangement of genes in a chromosome, taking the crossing over frequency as a unit of distance, Morgan compiled the first map of the location of genes in one of the Drosophila chromosomes: сh___13.6___ y___28.2___b. When constructing maps, they do not indicate the distance between genes, but the distance to each gene from the zero point of the beginning of the chromosome. The dominant allele is indicated by a capital letter, and the recessive allele by a lowercase letter. After constructing genetic maps, the question arose of whether the location of genes on the chromosome, constructed on the basis of the crossing-over frequency, corresponds to the true location. Each chromosome along its length has specific patterns of disks, which makes it possible to distinguish its different sections from each other. The material for testing was chromosomes in which, as a result of mutation, various chromosomal rearrangements arose: individual disks were missing, or they were inverted, or doubled. The physical distances between genes on the genetic map do not quite correspond to the established cytological distances. However, this does not reduce the value of genetic maps of chromosomes for predicting the likelihood of individuals with new combinations of traits. Based on an analysis of the results of numerous experiments with Drosophila, T. Morgan formulated chromosome theory heredity, the essence of which is the following: 1) genes are located in chromosomes, located linearly in them at a certain distance from each other; 2) genes located on the same chromosome belong to the same linkage group. The number of linkage groups corresponds to the haploid number of chromosomes; H) traits whose genes are located on the same chromosome are inherited linked; 4) in the offspring of heterozygous parents, new combinations of genes located in the same pair of chromosomes can arise as a result of crossing over during the process of meiosis. The frequency of crossing over depends on the distance between genes; 5) based on the linear arrangement of genes in a chromosome and the frequency of crossing over as an indicator of the distance between genes, chromosome maps can be constructed.
Bisexuality, intersexuality, gyandromorphism, chimerism according to sex chromes. The role of hormones and environmental conditions in the development of sex characteristics
Any zygote has x-chromes and autosomes, i.e. has both female and male genes, i.e. genetically any organism bisexual(bisexual). Intersex– hermaphrodites – individuals with developed both female and male characteristics. 2 types: true - have female and male gonads due to an imbalance of genes; conditional - have glands of one sex, and the external sexual characteristics of the other sex due to an imbalance of hormones. Sometimes found in insects and animals gyandromorphs– one part of the body has female characteristics, and the other has male characteristics. Causes: The female zygote is divided into 2 blastomeres. One of them lost one x-lame. From this blastomere the male half of the body will develop. Chimerism half of the chromosomes xx/xy are found in multiparous animals, in bulls - when the same organism contains xx chromosomes, and the reproduction of xy chromosomes is impaired. With normal feeding, males grow, and if female sex hormones are added to the food, females (fish fry) grow. If the larva of a sea worm attaches itself to the bottom of the sea, it is a female; if it attaches to the proboscis of a female, it is a male.
Types of sex determination in animals. Primary and secondary sex ratio. The problem of gender regulation
Determination ensures the formation of an equal number of males and females, which is necessary for the normal self-reproduction of the species. Types: 1) epigamous - the sex of an individual is determined during ontogenesis and depends on the external environment. 2) progamous - sex is determined during gametogenesis in the parents of the individual. 3) syngamous - sex is determined at the moment of fusion of gametes. Primary and secondary sex ratio: sex ratio, the cat is determined at the moment of fusion of gametes, called primary, always 1:1. Any change in sex ratio, either before or after birth, is called secondary. Usually after birth it shifts in favor of the female sex, therefore, in many animal species and in humans, more males are born than females: rabbits - 57%, humans - 51%, birds - 59%. Gender regulation problem: is of great economic importance. For example: in dairy farming and egg-laying poultry farming, females are desirable, and where the main product is meat, males are better. The problem is to separate the sperm into x- and y-fractions. Methods: 1) electrophoresis - x - sperm have a negative charge - move towards the cathode, and y - sperm move towards the anode. 80% guarantee. 2) Sedimentation method - x - the sperm is more dense and settles down, and y - remains on top. 3) Using a set of acids to change the pH of the female reproductive tract to create conditions only for x - or only for y-. 4) Parthenogenesis: genogenesis - obtaining females - the oocyte is irradiated with X-rays. first order, thereby delaying the divergence of chromium-m, an egg with a diploid set of chromium-m is formed, and a female develops into a cat without fertilization. Androgenesis - obtaining males - the nucleus of the egg is killed with X-rays, then two sperm penetrate it, the nuclei merge, giving a diploid set, there will be a male. 5) Method of dividing sperm into fractions based on the amount of DNA in sperm. 6) The younger the parents, the more likely they are to be born male. 7) The more sperm in the female genital tract, the more likely the birth of a male. 8) The longer the sperm is stored – the female. 9) Poultry feeding: if you add Ca to the rooster’s feed, then it’s female, and if K, it’s male. 10) In any population the law of equilibrium applies, i.e. The sex ratio tends to be 1:1.
Linked inheritance of traits. An analysis of the simultaneous inheritance of several traits in Drosophila, carried out by T. Morgan, showed that the results of analytical crossing of Fi hybrids sometimes differ from those expected in the case of their independent inheritance. In the descendants of such crosses, instead of freely combining traits from different pairs, a tendency was observed to inherit predominantly parental combinations of traits. This inheritance of traits was called linked. Linked inheritance is explained by the location of the corresponding genes on the same chromosome. As part of the latter, they are transmitted from generation to generation of cells and organisms, preserving the combination of parental alleles.
The dependence of linked inheritance of traits on the localization of genes on one chromosome gives grounds to consider chromosomes as separate clutch groups.
When analyzing crossing males from F 1, only two types of offspring appeared, similar to the parents in the combination of variants of the analyzed characters (gray body color and normal wings or black body color and short wings) in a 1:1 ratio. This indicates that F 1 males produce only two types of gametes with equal probability, which include the original parental combinations of alleles of the genes that control the named traits (BV or bv).
When analyzing the crossing of F 1 females, four types of descendants appeared with all possible combinations of characteristics. At the same time, offspring with parental combinations of traits were found in 83%. 17% of the offspring had new combinations of characters (gray body color and short wings or black body color and normal wings). It is clear that in these crosses there is also a tendency to linked inheritance of either dominant or recessive traits (83%). Partial disruption of linkage (17% of descendants) was explained by the process of crossing over - the exchange of corresponding sections of homologous chromosomes in prophase I of meiosis (see Fig. 3.72).
From the results of crossing it follows that Drosophila females form four types of gametes, most of which (83%) are non-crossover ((BV) and (bv)), 17% of the gametes they form appear as a result of crossing over and carry new combinations of alleles of the analyzed genes ((Bv ) and (bV)). The differences observed when males and females from F 1 are crossed with recessive homozygous partners are explained by the fact that, for reasons that are poorly understood, crossing over does not occur in Drosophila males. As a result, males who are diheterozygous for genes located on the same chromosome form two types of gametes. In females, crossing over occurs and leads to the formation of non-crossover and crossover gametes, two types of each. Therefore, four phenotypes appear in the offspring of an analyzing cross, two of which have new combinations of traits compared to the parents.
Rice. 6.14. Linked inheritance of traits
(body color and wing length in Drosophila):
I- crossing pure lines, II, III - analyzing crosses between males and females from F 1.
A study of the inheritance of other combinations of traits has shown that the percentage of crossover offspring for each pair of traits is always the same, but it varies for different pairs. This observation led to the conclusion that genes on chromosomes are arranged in a linear order. It was noted above that a chromosome is a linkage group of certain genes. Homologous chromosomes are identical linkage groups that differ from each other only alleles of individual genes. During conjugation, homologues are brought closer together by their allelic genes, and during crossing over they exchange corresponding regions. As a result, crossover chromosomes appear with a new set of alleles. The frequency with which exchange occurs at a site between two given genes depends on the distance between them (T. Morgan rule). The percentage of crossover gametes indirectly reflects the distance between genes. This distance is usually expressed in santshmorganids. One centimorganide is taken to be the distance between genes at which 1% of crossover offspring (crossover gametes) are formed.
As the distance between genes increases, the probability of crossing over in the area between them in gamete precursor cells increases. Since two of the four chromatids present in the bivalent are involved in the act of crossing over, even if there is an exchange between the genes of a given pair in all gamete precursor cells, the percentage of crossover germ cells cannot exceed 50. However, such a situation is only theoretically possible. In practice, as the distance between genes increases, the possibility of several crossovers occurring simultaneously in a given area increases (see Fig. 5.9). Since every second crossover leads to the restoration of the previous combination of alleles in the chromosome, with increasing distance the number of crossover gametes may not increase, but decrease. It follows from this that the percentage of crossover gametes is an indicator of the true distance between genes only when they are located sufficiently close, when the possibility of a second crossing over is excluded.
Disruption of linked inheritance of parental alleles as a result of crossing over allows us to talk about incomplete clutch unlike full clutch, observed, for example, in male Drosophila.
The use of analytical crossing in the experiments of T. Morgan showed that with its help it is possible to determine not only the composition of pairs of non-allelic genes, but also the nature of their joint inheritance. In the case of linked inheritance of traits, based on the results of analyzing crossing, the distance between genes in the chromosome can also be determined.

Rice. 6.15. Diagram explaining the low percentage of crossover gametes
(in relation to these two genes)
Plus denotes gamete precursor cells in which crossing over took place in the area between these two genes; crossover gametes are blackened
Genetic maps of chromosomes- this is a diagram of the relative position and relative distances between the genes of certain chromosomes located in the same linkage group.
For the first time, T. Morgan and his colleagues pointed out the possibility of constructing genetic maps of chromosomes. They experimentally showed that based on the phenomena of gene linkage and crossing over, it is possible to construct genetic maps of chromosomes. The ability to map is based on the constancy of the percentage of crossing over between certain genes. Genetic maps of chromosomes have been compiled for many types of organisms: insects (drosophila, mosquito, cockroach, etc.), fungi (yeast, aspergillus), bacteria and viruses.
Human genetic maps are used in medicine to diagnose a number of severe hereditary human diseases. Studies of the evolutionary process compare the genetic maps of different species of living organisms.