- dikelola. Dengan mekanisme kendali: dikontrol secara elektrik, kimia dan mekanis;
- tidak terkendali. Mereka tidak memiliki mekanisme gerbang dan selalu terbuka, ion mengalir terus-menerus, tetapi perlahan.
Potensi istirahat adalah perbedaan potensial listrik antara bagian luar dan lingkungan internal sel.
Mekanisme pembentukan potensial istirahat. Penyebab langsung dari potensial istirahat adalah konsentrasi anion dan kation yang tidak seimbang di dalam dan di luar sel. Pertama, susunan ion ini disebabkan oleh perbedaan permeabilitas. Kedua, secara signifikan lebih banyak ion kalium yang keluar dari sel daripada natrium.
Potensi tindakan- ini adalah eksitasi sel, fluktuasi cepat potensial membran akibat difusi ion masuk dan keluar sel.
Ketika suatu stimulus bekerja pada sel-sel jaringan yang tereksitasi, saluran natrium mula-mula diaktifkan dan dinonaktifkan dengan sangat cepat, kemudian saluran kalium diaktifkan dan dinonaktifkan dengan beberapa penundaan.
Akibatnya, ion-ion dengan cepat berdifusi masuk atau keluar sel sepanjang gradien elektrokimia. Ini adalah kegembiraan. Berdasarkan perubahan besar dan tanda muatan sel, dibedakan tiga fase:
- Fase 1 - depolarisasi. Mengurangi muatan sel menjadi nol. Natrium bergerak menuju sel sesuai dengan konsentrasi dan gradien listrik. Kondisi gerak: gerbang saluran natrium terbuka;
- fase ke-2 - inversi. Membalikkan tanda muatan. Inversi melibatkan dua bagian: naik dan turun.
Bagian menaik. Natrium terus berpindah ke dalam sel sesuai dengan gradien konsentrasi, tetapi melawan gradien listrik (itu mengganggu).
Bagian menurun. Kalium mulai meninggalkan sel sesuai dengan konsentrasi dan gradien listrik. Gerbang saluran kalium terbuka;
- Fase ke-3 - repolarisasi. Kalium terus keluar sel sesuai dengan gradien konsentrasi, namun berlawanan dengan gradien listrik.
Kriteria rangsangan
Dengan berkembangnya potensial aksi, terjadi perubahan eksitabilitas jaringan. Perubahan ini terjadi secara bertahap. Keadaan polarisasi awal membran biasanya mencerminkan potensial membran istirahat, yang sesuai dengan keadaan awal rangsangan dan, oleh karena itu, keadaan awal sel yang dapat dirangsang. Ini adalah tingkat rangsangan yang normal. Periode pra-lonjakan adalah periode awal mula potensial aksi. Rangsangan jaringan sedikit meningkat. Fase eksitabilitas ini adalah peninggian primer (primary supernormal exitability). Selama perkembangan prespike, potensial membran mendekati tingkat kritis depolarisasi, dan untuk mencapai tingkat ini, kekuatan stimulus mungkin kurang dari ambang batas.
Selama periode perkembangan lonjakan (potensial puncak), terjadi aliran ion natrium seperti longsoran ke dalam sel, akibatnya membran terisi ulang, dan kehilangan kemampuan untuk merespons rangsangan di atas dengan eksitasi. -kekuatan ambang batas. Fase eksitabilitas ini disebut refraktori absolut, yaitu. ineksitabilitas absolut, yang berlangsung hingga akhir pengisian ulang membran. Refraktori membran absolut terjadi karena saluran natrium terbuka penuh dan kemudian dinonaktifkan.
Setelah fase pengisian ulang berakhir, rangsangannya secara bertahap dikembalikan ke tingkat semula - ini adalah fase refraktori relatif, yaitu. ketidakberdayaan relatif. Ini berlanjut sampai muatan membran dikembalikan ke nilai yang sesuai dengan tingkat kritis depolarisasi. Karena selama periode ini potensi membran istirahat belum pulih, rangsangan jaringan menurun, dan eksitasi baru hanya dapat timbul di bawah pengaruh stimulus superthreshold. Penurunan rangsangan pada fase refraktori relatif dikaitkan dengan inaktivasi parsial saluran natrium dan aktivasi saluran kalium.
Periode berikutnya sesuai peningkatan tingkat rangsangan: fase peninggian sekunder atau rangsangan supernormal sekunder. Karena potensial membran pada fase ini lebih dekat ke tingkat kritis depolarisasi, dibandingkan dengan keadaan istirahat polarisasi awal, ambang stimulasi menurun, yaitu. rangsangan sel meningkat. Selama fase ini, eksitasi baru dapat muncul dari aksi rangsangan dengan kekuatan di bawah ambang batas. Saluran natrium tidak sepenuhnya dinonaktifkan selama fase ini. Potensial membran meningkat—terjadi keadaan hiperpolarisasi membran. Menjauh dari tingkat kritis depolarisasi, ambang rangsangan sedikit meningkat, dan eksitasi baru hanya dapat muncul di bawah pengaruh rangsangan dengan nilai melebihi ambang batas.
Mekanisme terjadinya potensial membran istirahat
Setiap sel dalam keadaan istirahat ditandai dengan adanya beda potensial transmembran (potensial istirahat). Biasanya, perbedaan muatan antara permukaan dalam dan luar membran adalah -80 hingga -100 mV dan dapat diukur menggunakan mikroelektroda eksternal dan intraseluler (Gbr. 1).
Beda potensial antara sisi luar dan dalam membran sel pada keadaan istirahat disebut potensial membran (potensial istirahat).
Penciptaan potensi istirahat dipastikan melalui dua proses utama - distribusi yang tidak merata ion anorganik antara ruang intra dan ekstraseluler dan permeabilitas yang tidak sama membran sel. Analisis komposisi kimia cairan ekstra dan intraseluler menunjukkan distribusi ion yang sangat tidak merata (Tabel 1).
Saat istirahat, terdapat banyak anion asam organik dan ion K+ di dalam sel, yang konsentrasinya 30 kali lebih tinggi daripada di luar; Sebaliknya, terdapat ion Na+ 10 kali lebih banyak di luar sel dibandingkan di dalam; CI- juga lebih besar di bagian luar.
Saat istirahat, membran sel saraf paling permeabel terhadap K+, kurang permeabel terhadap CI- dan sangat sedikit permeabel terhadap Na+.Permeabilitas membran serabut saraf terhadap Na+ saat istirahat 100 kali lebih kecil dibandingkan K+. Bagi banyak anion asam organik, membran dalam keadaan diam benar-benar kedap air.
Beras. 1. Mengukur potensial istirahat serat otot (A) menggunakan mikroelektroda intraseluler: M - mikroelektroda; I - elektroda acuh tak acuh. Sinar pada layar osiloskop (B) menunjukkan bahwa sebelum membran ditembus oleh mikroelektroda, beda potensial antara M dan I sama dengan nol. Pada saat tusukan (ditunjukkan oleh panah), terdeteksi perbedaan potensial, yang menunjukkan bahwa sisi dalam membran bermuatan negatif relatif terhadap permukaan luarnya (menurut B.I. Khodorov)
Meja. Konsentrasi ion intra dan ekstraseluler dalam sel otot hewan berdarah panas, mmol/l (menurut J. Dudel)
|
Konsentrasi intraseluler |
Konsentrasi ekstraseluler |
|
|
A- (anion senyawa organik) |
Karena adanya gradien konsentrasi, K+ mencapai permukaan luar sel, membawa muatan positifnya. Anion dengan berat molekul tinggi tidak dapat mengikuti K+ karena membrannya kedap terhadapnya. Ion Na+ juga tidak dapat menggantikan ion kalium yang hilang, karena permeabilitas membran terhadap ion tersebut jauh lebih kecil. CI- sepanjang gradien konsentrasi hanya dapat bergerak di dalam sel, sehingga meningkatkan muatan negatif pada permukaan bagian dalam membran. Akibat pergerakan ion ini, polarisasi membran terjadi ketika permukaan luarnya bermuatan positif dan permukaan bagian dalam bermuatan negatif.
Medan listrik yang tercipta pada membran secara aktif mengganggu distribusi ion antara isi internal dan eksternal sel. Ketika muatan positif pada permukaan luar sel meningkat, ion K+ yang bermuatan positif semakin sulit berpindah dari dalam ke luar. Sepertinya sedang bergerak menanjak. Semakin besar muatan positif pada permukaan luar maka semakin sedikit ion K+ yang dapat mencapai permukaan sel. Pada potensial tertentu pada membran, jumlah ion K+ yang melintasi membran pada kedua arah adalah sama, yaitu Gradien konsentrasi kalium diseimbangkan oleh potensi yang ada melintasi membran. Potensial dimana fluks difusi ion-ion menjadi sama dengan fluks ion-ion sejenis yang masuk ke dalamnya arah sebaliknya, disebut potensial kesetimbangan untuk ion tertentu. Untuk ion K+, potensial kesetimbangannya adalah -90 mV. Pada serabut saraf bermielin, nilai potensial keseimbangan ion CI- mendekati nilai potensial membran istirahat (-70 mV). Oleh karena itu, meskipun konsentrasi ion CI- di luar serat lebih besar daripada di dalamnya, arus satu arah tidak diamati sesuai dengan gradien konsentrasi. Dalam hal ini, perbedaan konsentrasi diseimbangkan dengan potensial yang ada pada membran.
Ion Na+ sepanjang gradien konsentrasi harus masuk ke dalam sel (potensial kesetimbangannya adalah +60 mV), dan keberadaan muatan negatif di dalam sel tidak boleh mengganggu aliran ini. Dalam hal ini, Na+ yang masuk akan menetralkan muatan negatif di dalam sel. Namun hal ini sebenarnya tidak terjadi karena membran dalam keadaan diam mempunyai permeabilitas yang buruk terhadap Na+.
Mekanisme terpenting yang mempertahankan konsentrasi ion Na+ intraseluler yang rendah dan konsentrasi ion K+ yang tinggi adalah pompa natrium-kalium (transpor aktif). Diketahui bahwa di dalam membran sel terdapat sistem pembawa yang masing-masing diikat oleh ion Na+ sanggurdi yang terletak di dalam sel dan membawanya keluar. Dari luar, pembawa mengikat dua ion K+ yang terletak di luar sel, yang ditransfer ke sitoplasma. Pasokan energi untuk pengoperasian sistem transporter disediakan oleh ATP. Pengoperasian pompa menggunakan sistem seperti itu memberikan hasil sebagai berikut:
- konsentrasi ion K+ yang tinggi dipertahankan di dalam sel, yang menjamin nilai potensial istirahat yang konstan. Karena fakta bahwa selama satu siklus pertukaran ion, satu ion positif lebih banyak dikeluarkan dari sel daripada yang dimasukkan, transpor aktif berperan dalam menciptakan potensial istirahat. Dalam hal ini, mereka berbicara tentang pompa elektrogenik, karena pompa itu sendiri menghasilkan pompa yang kecil, tapi D.C. muatan positif dari sel, dan karena itu memberikan kontribusi langsung pada pembentukan potensial negatif di dalamnya. Namun besarnya kontribusi pompa elektrogenik terhadap arti umum potensi istirahat biasanya kecil dan berjumlah beberapa milivolt;
- konsentrasi ion Na + yang rendah dipertahankan di dalam sel, yang, di satu sisi, memastikan berfungsinya mekanisme pembangkitan potensial aksi, dan di sisi lain, memastikan pelestarian osmolaritas normal dan volume sel;
- mempertahankan gradien konsentrasi Na+ yang stabil, pompa natrium-kalium mendorong pengangkutan K+, Na+ asam amino dan gula melintasi membran sel.
Dengan demikian, terjadinya beda potensial transmembran (potensial istirahat) disebabkan oleh tingginya konduktivitas membran sel dalam keadaan diam untuk ion K+, CI-, asimetri ionik konsentrasi ion K+ dan ion CI-, kerja dari sistem transpor aktif (Na + / K + -ATPase), yang menciptakan dan mempertahankan asimetri ionik.
Potensi aksi serabut saraf, impuls saraf
Potensi aksi - Ini adalah fluktuasi jangka pendek dalam perbedaan potensial membran sel yang tereksitasi, disertai dengan perubahan tanda muatannya.
Potensi aksi adalah tanda spesifik utama dari eksitasi. Pendaftarannya menunjukkan bahwa sel atau strukturnya merespons dampak dengan eksitasi. Namun sebagaimana telah disebutkan, PD pada beberapa sel dapat terjadi secara spontan (spontan). Sel-sel tersebut terdapat pada alat pacu jantung, dinding pembuluh darah, sistem saraf. AP digunakan sebagai pembawa informasi, mentransmisikannya dalam bentuk sinyal listrik (electrical signaling) sepanjang serabut saraf aferen dan eferen, sistem konduksi jantung, dan juga untuk memulai kontraksi sel otot.
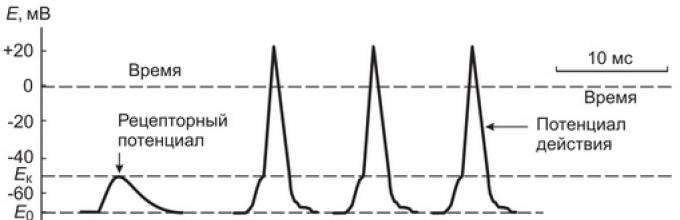
Mari kita perhatikan alasan dan mekanisme pembentukan AP di serabut saraf aferen yang membentuk reseptor sensorik primer. Penyebab langsung terjadinya (generasi) AP di dalamnya adalah potensi reseptor.
Jika kita mengukur beda potensial pada membran nodus Ranvier yang paling dekat dengan ujung saraf, maka dalam interval antara tumbukan pada kapsul sel darah Pacinian, ia tetap tidak berubah (70 mV), dan selama pemaparan ia mengalami depolarisasi hampir bersamaan dengan depolarisasi. membran reseptor ujung saraf.
Dengan peningkatan gaya tekanan pada badan Pacinian, menyebabkan peningkatan potensial reseptor hingga 10 mV, osilasi cepat potensial membran biasanya dicatat di simpul Ranvier terdekat, disertai dengan pengisian ulang membran - potensial aksi. (AP), atau impuls saraf (Gbr. 2). Jika kekuatan tekanan pada tubuh semakin meningkat, maka amplitudo potensial reseptor meningkat dan sejumlah potensial aksi dengan frekuensi tertentu dihasilkan di ujung saraf.

Beras. 2. Gambaran skematis mekanisme pengubahan potensial reseptor menjadi potensial aksi (impuls saraf) dan perambatan impuls sepanjang serabut saraf
Inti dari mekanisme pembangkitan AP adalah potensial reseptor menyebabkan munculnya arus sirkular lokal antara membran reseptor yang terdepolarisasi pada bagian ujung saraf yang tidak bermielin dan membran nodus pertama Ranvier. Arus ini, yang dibawa oleh Na+, K+, CI- dan ion mineral lainnya, “mengalir” tidak hanya sepanjang, tetapi juga melintasi membran serabut saraf di area nodus Ranvier. Di membran nodus Ranvier, berbeda dengan membran reseptor ujung saraf itu sendiri, terdapat saluran natrium dan kalium yang bergantung pada tegangan ion dengan kepadatan tinggi.
Ketika nilai depolarisasi sekitar 10 mV tercapai pada membran intersepsi Ranvier, saluran natrium yang bergantung pada tegangan cepat terbuka dan melalui saluran tersebut aliran ion Na+ mengalir ke aksoplasma sepanjang gradien elektrokimia. Hal ini menyebabkan depolarisasi cepat dan pengisian ulang membran di nodus Ranvier. Namun, bersamaan dengan terbukanya saluran natrium berpintu tegangan cepat di membran nodus Ranvier, saluran kalium berpintu tegangan lambat terbuka dan ion K+ mulai meninggalkan axoillasma, keluarannya tertinggal dibandingkan masuknya ion Na+. Jadi, ion Na+ yang memasuki aksoplasma dengan kecepatan tinggi dengan cepat mendepolarisasi dan mengisi ulang membran dalam waktu singkat (0,3-0,5 ms), dan ion K+ yang keluar mengembalikan distribusi muatan awal pada membran (repolarisasi membran). Akibatnya, ketika terjadi benturan mekanis pada sel darah Pacinian dengan kekuatan yang sama dengan atau melebihi ambang batas, osilasi potensial jangka pendek diamati pada membran simpul Ranvier terdekat dalam bentuk depolarisasi dan repolarisasi membran yang cepat. , yaitu PD (impuls saraf) dihasilkan.
Karena penyebab langsung timbulnya AP adalah potensial reseptor, dalam hal ini disebut juga potensial generator. Jumlah impuls saraf dengan amplitudo dan durasi yang sama yang dihasilkan per satuan waktu sebanding dengan amplitudo potensial reseptor, dan oleh karena itu dengan kekuatan tekanan pada reseptor. Proses mengubah informasi tentang kekuatan pengaruh yang terkandung dalam amplitudo potensial reseptor menjadi sejumlah impuls saraf diskrit disebut pengkodean informasi diskrit.
Secara detail mekanisme ion dan dinamika waktu proses pembangkitan AP dipelajari dalam kondisi eksperimental di bawah pengaruh buatan pada serabut saraf sengatan listrik dengan kekuatan dan durasi yang berbeda-beda.
Sifat potensial aksi serabut saraf (impuls saraf)
Selaput serabut saraf pada titik lokalisasi elektroda perangsang bereaksi terhadap pengaruh arus yang sangat lemah yang belum mencapai nilai ambang batas. Respon ini disebut lokal, dan osilasi beda potensial pada membran disebut potensial lokal.
Respon lokal pada membran sel yang tereksitasi dapat mendahului terjadinya potensial aksi atau terjadi sebagai proses yang berdiri sendiri. Ini mewakili fluktuasi jangka pendek (depolarisasi dan repolarisasi) dari potensial istirahat, tidak disertai dengan pengisian ulang membran. Depolarisasi membran selama pengembangan potensial lokal disebabkan oleh masuknya ion Na+ ke dalam aksoplasma, dan repolarisasi disebabkan oleh tertundanya keluarnya ion K+ dari aksoplasma.
Jika membran terkena arus listrik dengan kekuatan yang meningkat, maka pada nilai ini, yang disebut ambang batas, depolarisasi membran dapat mencapai tingkat kritis - Ec, di mana saluran natrium yang bergantung pada tegangan cepat terbuka. Akibatnya, terjadi peningkatan aliran ion Na+ ke dalam sel seperti longsoran salju. Proses depolarisasi yang terinduksi menjadi semakin cepat, dan potensi lokal berkembang menjadi potensial aksi.
Telah disebutkan hal itu fitur karakteristik PD adalah inversi (perubahan) jangka pendek dari tanda muatan pada membran. Di luar, ia menjadi bermuatan negatif dalam waktu singkat (0,3-2 ms), dan di dalam bermuatan positif. Besarnya inversi bisa mencapai 30 mV, dan besarnya seluruh potensial aksi adalah 60-130 mV (Gbr. 3).
Meja. Karakteristik komparatif potensi lokal dan potensi aksi
|
Ciri |
Potensi lokal |
Potensi tindakan |
|
Daya konduksi |
Menyebar secara lokal, 1-2 mm dengan redaman (penurunan) |
Menyebar tanpa redaman dalam jarak jauh di sepanjang serabut saraf |
|
Hukum "kekuatan" |
Mengirimkan |
Tidak patuh |
|
Hukum semua atau tidak sama sekali |
Tidak patuh |
Mengirimkan |
|
Fenomena penjumlahan |
Ringkasnya, meningkat dengan stimulasi subthreshold yang sering dan berulang |
Tidak bertambah |
|
Nilai amplitudo |
||
|
Sifat dpt dirangsang |
Meningkat |
Menurun sampai pada titik tidak dapat dirangsang sepenuhnya (refractoriness) |
|
Besaran stimulus |
Bawah sadar |
Ambang batas dan ambang batas super |
Potensi aksi, tergantung pada sifat perubahan muatan pada permukaan bagian dalam membran, dibagi menjadi fase depolarisasi, repolarisasi dan hiperpolarisasi membran. Depolarisasi sebut saja seluruh bagian menaik dari PD, yang didalamnya diidentifikasi wilayah-wilayah yang sesuai dengan potensi lokal (dari tingkat E 0 sebelum Ek), depolarisasi cepat (dari level Ek ke tingkat 0 mV), inversi tanda muatan (dari 0 mV ke nilai puncak atau awal repolarisasi). Repolarisasi disebut bagian AP yang menurun, yang mencerminkan proses pemulihan polarisasi asli membran. Pada awalnya, repolarisasi terjadi dengan cepat, tetapi semakin mendekati level tersebut E 0, kecepatannya bisa melambat dan bagian ini disebut melacak hal-hal negatif(atau menelusuri potensi negatif). Di beberapa sel, setelah repolarisasi, terjadi hiperpolarisasi (peningkatan polarisasi membran). Mereka memanggilnya menelusuri potensi positif.
Bagian awal AP yang mengalir cepat dengan amplitudo tinggi disebut juga puncak, atau paku. Ini mencakup fase depolarisasi dan repolarisasi cepat.
Dalam mekanisme perkembangan PD, peran paling penting adalah saluran ion yang bergantung pada tegangan dan peningkatan permeabilitas membran sel terhadap ion Na+ dan K+ yang tidak simultan. Jadi, ketika arus listrik bekerja pada sel, hal ini menyebabkan depolarisasi membran dan, ketika muatan membran menurun hingga tingkat kritis (Ec), saluran natrium dengan gerbang tegangan terbuka. Seperti yang telah disebutkan, saluran ini dibentuk oleh molekul protein yang tertanam di dalam membran, di dalamnya terdapat pori dan dua mekanisme gerbang. Salah satu mekanisme gerbang, aktivasi, memastikan (dengan partisipasi segmen 4) pembukaan (aktivasi) saluran selama depolarisasi membran, dan yang kedua (dengan partisipasi loop intraseluler antara domain ke-3 dan ke-4) memastikan inaktivasinya , yang berkembang ketika membran diisi ulang (Gbr. 4). Karena kedua mekanisme ini dengan cepat mengubah posisi gerbang saluran, saluran natrium dengan gerbang tegangan adalah saluran ion cepat. Keadaan ini sangat penting bagi generasi PD di jaringan yang bersemangat dan untuk konduksinya sepanjang membran serabut saraf dan otot.

Beras. 3. Potensial aksi, fasa dan arus ioniknya (a, o). Deskripsi dalam teks

Beras. 4. Posisi gerbang dan keadaan aktivitas saluran natrium dan kalium dengan gerbang tegangan pada berbagai tingkat polarisasi membran
Agar saluran natrium berpintu tegangan memungkinkan ion Na+ masuk ke dalam sel, hanya gerbang aktivasi yang harus dibuka, karena gerbang inaktivasi terbuka dalam kondisi istirahat. Inilah yang terjadi ketika depolarisasi membran mencapai suatu tingkat Ek(Gbr. 3, 4).
Pembukaan gerbang aktivasi saluran natrium menyebabkan masuknya natrium seperti longsoran ke dalam sel, didorong oleh gaya gradien elektrokimia. Karena ion Na+ membawa muatan positif, ion-ion tersebut menetralkan kelebihan muatan negatif pada permukaan bagian dalam membran, mengurangi beda potensial melintasi membran dan mendepolarisasinya. Segera, ion Na+ memberikan kelebihan muatan positif ke permukaan bagian dalam membran, yang disertai dengan inversi (perubahan) tanda muatan dari negatif menjadi positif.
Namun, saluran natrium tetap terbuka hanya sekitar 0,5 ms dan setelah jangka waktu ini sejak permulaan
AP menutup gerbang inaktivasi, saluran natrium menjadi tidak aktif dan kedap terhadap ion Na+, sehingga masuknya ion Na+ ke dalam sel sangat terbatas.
Dari saat depolarisasi membran hingga levelnya Ek aktivasi saluran kalium dan pembukaan gerbangnya untuk ion K+ juga diamati. Ion K+, di bawah pengaruh gaya gradien konsentrasi, meninggalkan sel, menghilangkan muatan positif darinya. Namun, mekanisme gerbang saluran kalium berfungsi lambat dan laju keluarnya muatan positif yang mengandung ion K+ dari sel ke luar sel tertinggal dibandingkan masuknya ion Na+. Aliran ion K+, menghilangkan kelebihan muatan positif dari sel, menyebabkan pemulihan distribusi muatan awal pada membran atau repolarisasinya, dan di sisi dalam, sesaat setelah pengisian ulang, muatan negatif dipulihkan.
Terjadinya AP pada membran tereksitasi dan selanjutnya pemulihan potensial istirahat awal pada membran dimungkinkan karena dinamika masuk dan keluarnya muatan positif ion Na+ dan K+ ke dalam sel dan keluar sel berbeda. Pintu masuk ion Na+ mendahului keluarnya ion K+. Jika proses-proses ini berada dalam kesetimbangan, maka beda potensial melintasi membran tidak akan berubah. Perkembangan kemampuan untuk merangsang dan menghasilkan AP oleh sel otot dan saraf yang tereksitasi disebabkan oleh pembentukan dua jenis saluran ion dengan kecepatan berbeda di membrannya - natrium cepat dan kalium lambat.
Untuk menghasilkan satu AP, diperlukan sejumlah energi yang relatif kecil untuk masuk ke dalam sel. jumlah besar Ion Na+ yang tidak mengganggu distribusinya ke luar dan ke dalam sel. Jika AP yang dihasilkan dalam jumlah besar, distribusi ion di kedua sisi membran sel dapat terganggu. Namun, di kondisi normal hal ini dapat dicegah dengan pengoperasian pompa Na+, K+.
Dalam kondisi alami, di neuron sistem saraf pusat, potensial aksi terutama muncul di daerah bukit akson, di neuron aferen - di simpul Ranvier dari ujung saraf yang paling dekat dengan reseptor sensorik, yaitu. di bagian membran yang terdapat saluran natrium dengan gerbang tegangan selektif cepat dan saluran kalium lambat. Pada jenis sel lain (misalnya alat pacu jantung, miosit halus), tidak hanya saluran natrium dan kalium, tetapi juga saluran kalsium berperan dalam terjadinya AP.
Mekanisme persepsi dan transformasi sinyal menjadi potensial aksi pada reseptor sensorik sekunder berbeda dengan mekanisme yang dibahas pada reseptor sensorik primer. Dalam reseptor ini, persepsi sinyal dilakukan oleh sel neurosensori khusus (fotoreseptor, penciuman) atau sensoroepithelial (pengecapan, pendengaran, vestibular). Masing-masing sel sensitif ini memiliki mekanisme khusus untuk menangkap sinyal. Namun, di semua sel, energi sinyal yang dirasakan (stimulus) diubah menjadi osilasi beda potensial membran plasma, yaitu. menjadi potensi reseptor.
Dengan demikian, poin kunci dalam mekanisme sel-sel sensorik mengubah sinyal yang dirasakan menjadi potensi reseptor adalah perubahan permeabilitas saluran ion sebagai respons terhadap stimulus. Pembukaan saluran ion Na +, Ca 2+, K + - selama persepsi dan transformasi sinyal dicapai dalam sel-sel ini dengan partisipasi protein G, pembawa pesan intraseluler kedua, pengikatan ligan, dan fosforilasi saluran ion. Biasanya, potensi reseptor yang timbul di sel sensorik menyebabkan pelepasan celah sinaptik neurotransmitter yang memastikan transmisi sinyal ke membran postsinaptik dari ujung saraf aferen dan menghasilkan impuls saraf pada membrannya. Proses-proses ini dijelaskan secara rinci dalam bab tentang sistem sensorik.
Potensial aksi dapat dicirikan oleh amplitudo dan durasi, yang untuk serabut saraf yang sama tetap sama selama aksi merambat sepanjang serabut tersebut. Oleh karena itu, potensial aksi disebut potensial diskrit.
Ada hubungan tertentu antara sifat dampak pada reseptor sensorik dan jumlah AP yang timbul di serabut saraf aferen sebagai respons terhadap dampak tersebut. Hal ini terletak pada kenyataan bahwa dengan kekuatan atau durasi paparan yang besar, a jumlah yang lebih besar impuls saraf, mis. ketika efeknya meningkat, impuls dengan frekuensi yang lebih tinggi akan dikirim dari reseptor ke sistem saraf. Proses mengubah informasi tentang sifat efek menjadi frekuensi dan parameter lain dari impuls saraf yang ditransmisikan ke sistem saraf pusat disebut pengkodean informasi diskrit.
Untuk penjelasan asal mula potensi istirahat disarankan berbagai teori. Asal usul pemahaman modern tentang masalah ini terletak pada karya V. Yu.Chagovets, yang pada tahun 1896, sebagai mahasiswa kedokteran, mengungkapkan gagasan tentang sifat ionik dari proses bioelektrik dan mencoba menerapkan teori Arrhenius tentang disosiasi elektrolitik untuk menjelaskan asal usul potensial ini. Selanjutnya, pada tahun 1902, Y. Bernstein mengembangkan teori membran-ion, yang dimodifikasi dan dibuktikan secara eksperimental oleh A. Hodgkin dan A. Huxley (1952) dan sekarang diterima secara luas. Menurut teori ini, potensi bioelektrik disebabkan oleh perbedaan konsentrasi ion K, Na, dan Cl di dalam dan di luar sel serta perbedaan permeabilitas membran permukaan terhadap ion tersebut.
Protoplasma sel saraf dan otot mengandung ion kalium 30-50 kali lebih banyak, ion natrium 8-10 kali lebih sedikit, dan ion klorin 50 kali lebih sedikit dibandingkan cairan ekstraseluler.
Hambatan dalam pemerataan cepat perbedaan konsentrasi ini adalah membran plasma tertipis (sekitar 100 Å) yang menutupi sel-sel hidup.
|
Gagasan tentang struktur membran ini didasarkan pada data yang diperoleh dengan mikroskop elektron, mikroskop optik, difraksi sinar-X dan analisis kimia. Diasumsikan bahwa membran terdiri dari lapisan ganda molekul fosfolipid, ditutupi di bagian dalam dengan lapisan molekul protein, dan di bagian luar dengan lapisan molekul karbohidrat kompleks - mukopolisakarida. Struktur membran tiga lapis ditunjukkan secara skematis pada beras. 116. Beras. 116. Skema struktur molekul membran. Lapisan lipid bimolekuler Z ditampilkan (lingkaran menunjukkan gugus polar fosfolipid) dan dua lapisan tunggal non-lipid: lapisan luar adalah mukopolisakarida - X, lapisan dalam adalah protein - Y (menurut Robertson). |
Membran sel mengandung tubulus yang sangat tipis - “pori-pori” dengan diameter beberapa angstrom. Melalui tubulus ini, molekul air dan zat lain, serta ion dengan diameter yang sesuai dengan ukuran pori-pori, masuk dan keluar sel.
Pada elemen struktural ion-ion pribadi terfiksasi di dalam membran, yang memberikan muatan tertentu pada dinding pori-porinya dan dengan demikian mempersulit atau memudahkan ion untuk melewatinya. Dengan demikian, diasumsikan bahwa adanya gugus fosfat dan karboksil yang terdisosiasi dalam membran merupakan alasan mengapa membran serabut saraf kurang permeabel terhadap anion dibandingkan kation.
Permeabilitas membran untuk kation yang berbeda juga tidak sama, dan secara alami berubah pada keadaan fungsional jaringan yang berbeda. Saat istirahat, membran serabut saraf kira-kira 20-100 kali lebih permeabel terhadap ion K dibandingkan ion Na, dan ketika tereksitasi, permeabilitas natrium mulai secara signifikan melebihi permeabilitas kalium membran.
Untuk memahami mekanisme munculnya potensial membran istirahat dari sudut pandang teori Bernstein-Hodgkin, mari kita perhatikan model eksperimen. Paruh pertama kapal ( beras. 117), dipisahkan oleh membran semipermeabel buatan, yang pori-porinya dengan bebas memungkinkan ion K bermuatan positif melewatinya dan tidak membiarkan ion SO"4 bermuatan negatif melewatinya, diisi dengan larutan pekat K2SO4, dan bagian kiri setengahnya juga diisi dengan larutan K2SO4, tetapi konsentrasinya lebih rendah.
 |
Karena adanya gradien konsentrasi, ion K akan mulai berdifusi melalui membran terutama dari bagian kanan wadah (yang konsentrasinya C1) ke kiri (dengan konsentrasi C2). Oleh karena itu, anion SO"4 yang bermuatan negatif, yang membrannya kedap air, akan terkonsentrasi di bagian kanan bejana dekat permukaan membran. Dengan muatan negatifnya, mereka akan menahan ion K secara elektrostatis pada permukaan membran sebelah kiri. Akibatnya, membran terpolarisasi: timbul perbedaan potensial antara kedua permukaannya. Beras. 117. Munculnya beda potensial pada membran buatan yang memisahkan larutan K2SO4 dengan konsentrasi berbeda (C1 dan C2). Membran secara selektif permeabel terhadap kation K (lingkaran kecil) dan tidak memungkinkan anion SO4 melewatinya (lingkaran besar). 1 dan 2 - elektroda direndam dalam larutan; 3 - alat pengukur listrik. |
Jika sekarang kita menurunkan elektroda ke bagian kanan dan kiri bejana, alat pengukur listrik akan mendeteksi adanya beda potensial, sedangkan larutan dengan konsentrasi ion K2SO4 yang lebih rendah, di mana sebagian besar terjadi difusi ion K bermuatan positif, memperoleh muatan positif relatif terhadap larutan dengan konsentrasi K2SO4 yang lebih tinggi.
Beda potensial (E) pada kasus yang dipertimbangkan dapat dihitung dengan menggunakan rumus Nernst:

Ada banyak alasan untuk percaya bahwa hubungan serupa terjadi pada serabut saraf yang hidup, karena konsentrasi ion K dalam protoplasma lebih dari 30 kali lebih tinggi daripada konsentrasi ion-ion ini dalam larutan eksternal, dan anion organik (protein, dll.) dari protoplasma melalui membran praktis tidak menembus.
Dalam keadaan istirahat fisiologis, difusi ion K bermuatan positif dari protoplasma ke cairan luar memberikan muatan positif pada permukaan luar membran, dan permukaan bagian dalam bermuatan negatif.
Argumen penting yang mendukung kebenaran gagasan ini adalah kenyataan bahwa beda potensial yang dihitung menggunakan rumus Nernst antara sisi luar dan dalam membran serat otot (sekitar 90 mV) ternyata mendekati yang diukur dalam percobaan menggunakan mikroelektroda intraseluler.
Ditemukan juga bahwa peningkatan konsentrasi ion K di lingkungan luar sel, dan oleh karena itu penurunan perbedaan konsentrasi ion-ion ini di kedua sisi membran, menyebabkan penurunan potensial istirahat, dan dalam rentang konsentrasi tertentu, pergeseran ini secara kuantitatif bertepatan dengan yang dihitung menggunakan rumus Nernst.
Namun, bukti langsung yang paling penting tentang kebenaran gagasan ini diperoleh oleh A. Hodgkin dan rekan-rekannya (1962) dalam eksperimen penggantian protoplasma pada serabut saraf raksasa moluska cumi-cumi dengan larutan garam. Protoplasma diperas dengan hati-hati dari serat yang berdiameter sekitar 1 ml, dan cangkang yang runtuh diisi dengan larutan garam buatan.
Dalam kasus ketika konsentrasi ion kalium dalam larutan ini mendekati intraseluler, terjadi perbedaan potensial antara sisi dalam dan luar membran, kira-kira sama dengan potensial istirahat serat normal (50-80 mV). Penurunan konsentrasi ion K dalam larutan internal menyebabkan penurunan alami atau bahkan distorsi potensial istirahat.
Eksperimen tersebut menunjukkan bahwa gradien konsentrasi ion K memang merupakan faktor utama yang menentukan nilai potensial istirahat serabut saraf.
Selain ion K, ion Na yang berdifusi ke dalam protoplasma dari cairan ekstraseluler yang konsentrasinya tinggi juga berperan dalam munculnya potensial istirahat. Difusi ini sangat terhambat oleh rendahnya permeabilitas natrium pada membran saat istirahat. Namun, saat berdifusi melalui membran ke dalam protoplasma, ion Na mentransfer muatan positifnya ke sini, yang agak mengurangi nilai potensial istirahat yang diciptakan oleh difusi ion K dari sel. Hal ini menjelaskan fakta bahwa potensi istirahat sebagian besar sel dan serabut saraf bukanlah 90 mV, seperti yang diharapkan jika potensi ini diciptakan hanya oleh ion K, tetapi 60-70 mV.
Dengan demikian, nilai potensial istirahat serabut saraf dan sel ditentukan oleh perbandingan jumlah ion K bermuatan positif yang berdifusi per satuan waktu dari sel ke luar, dan ion Na bermuatan positif yang berdifusi melalui membran dalam arah yang berlawanan. Semakin tinggi rasio ini, semakin besar potensi istirahatnya, dan sebaliknya.
bidang_teks
bidang_teks
panah_ke atas
Potensi membran istirahat (MPP) atau potensi istirahat (PP) adalah beda potensial sel istirahat antara sisi dalam dan sisi luar membran. Bagian dalam membran sel bermuatan negatif dibandingkan bagian luarnya. Dengan mengambil potensial solusi eksternal sebagai nol, maka MPP ditulis dengan tanda minus. Besarnya MPP tergantung pada jenis jaringan dan bervariasi dari -9 hingga -100 mV. Oleh karena itu, membran sel dalam keadaan istirahat terpolarisasi. Penurunan nilai MPP disebut depolarisasi, meningkatkan - hiperpolarisasi, mengembalikan nilai aslinya MPP- ulangpolarisasi membran.
Ketentuan dasar teori asal usul membran MPP intinya sebagai berikut. Dalam keadaan istirahat, membran sel sangat permeabel terhadap ion K+ (pada beberapa sel dan SG), kurang permeabel terhadap Na+ dan praktis kedap terhadap protein intraseluler dan ion organik lainnya. Ion K+ berdifusi keluar sel sepanjang gradien konsentrasi, dan anion yang tidak berpenetrasi tetap berada di sitoplasma, memberikan tampilan perbedaan potensial melintasi membran.
Beda potensial yang dihasilkan menghalangi keluarnya K+ dari sel dan pada nilai tertentu terjadi keseimbangan antara keluarnya K+ sepanjang gradien konsentrasi dan masuknya kation-kation tersebut sepanjang gradien listrik yang dihasilkan. Potensi membran di mana keseimbangan ini tercapai disebut potensi keseimbangankirmizi Nilainya dapat dihitung dari persamaan Nernst:
Di mana Ek- potensi keseimbangan untuk KE + ; R- konstanta gas; T- suhu absolut; F - nomor Faraday; P- valensi K + (+1), [Kn + ] - [K + penginapan ] - konsentrasi eksternal dan internal K + -
Jika Anda pergi dari logaritma natural menjadi desimal dan substitusikan nilai numerik konstanta ke dalam persamaan, maka persamaan tersebut akan berbentuk: 
Pada neuron tulang belakang (Tabel 1.1) E k = -90 mV. Nilai MPP yang diukur menggunakan mikroelektroda jauh lebih rendah - 70 mV.
Tabel 1.1. Konsentrasi beberapa ion di dalam dan di luar neuron motorik tulang belakang mamalia
| Dan dia |
Konsentrasi |
(mmol/l H2O) |
Potensi berat (mv) |
|
di dalam sel |
di luar kandang |
||
| Tidak+ | 15,0 | 150,0 | |
| K+ | 150,0 | 5,5 | |
| sl - | 125,0 | ||
|
Potensial membran istirahat = -70 mV |
|||
Jika potensial membran sel bersifat kalium, maka sesuai dengan persamaan Nernst, nilainya akan menurun secara linier dengan penurunan gradien konsentrasi ion-ion tersebut, misalnya dengan peningkatan konsentrasi K+ di ekstraseluler. cairan. Namun ketergantungan linier Besarnya RMP (potensial membran istirahat) pada gradien konsentrasi K+ hanya ada bila konsentrasi K+ dalam cairan ekstraseluler berada di atas 20 mM. Pada konsentrasi K+ yang lebih rendah di luar sel, kurva ketergantungan E m pada logaritma rasio konsentrasi kalium di luar dan di dalam sel berbeda dengan teori. Penyimpangan yang terjadi dalam ketergantungan eksperimental nilai MPP dan gradien konsentrasi K + yang dihitung secara teoritis menggunakan persamaan Nernst dapat dijelaskan dengan asumsi bahwa MPP sel yang dapat dirangsang ditentukan tidak hanya oleh kalium, tetapi juga oleh potensial kesetimbangan natrium dan klorin. . Dengan argumen serupa dengan argumen sebelumnya, kita dapat menulis:
Nilai potensi keseimbangan natrium dan klorin untuk neuron tulang belakang (Tabel 1.1) masing-masing sama dengan +60 dan -70 mV. Nilai E Cl sama dengan nilai MPP. Hal ini menunjukkan distribusi pasif ion klorin melintasi membran sesuai dengan gradien kimia dan listrik. Untuk ion natrium, gradien kimia dan listrik diarahkan ke dalam sel.
Kontribusi masing-masing potensial kesetimbangan terhadap nilai MPP ditentukan oleh perbandingan antara permeabilitas membran sel untuk masing-masing ion tersebut. Potensi membran dihitung menggunakan persamaan Goldmann:

E m- potensi membran; R- konstanta gas; T- suhu absolut; F- Nomor Faraday; RK, P Na Dan RKl- konstanta permeabilitas membran untuk K + Na + dan Cl; [KE+ n ], [ K + ay, [ Tidak+ n [ Tidak + ay], [Cl - n] dan [Cl - ext] - konsentrasi K+, Na+ dan Cl di luar (n) dan di dalam (dalam) sel.
Dengan mensubstitusi konsentrasi ion dan nilai MPP yang diperoleh dalam studi eksperimental ke dalam persamaan ini, dapat ditunjukkan bahwa untuk akson cumi-cumi raksasa harus terdapat rasio konstanta permeabilitas P ke: P Na: P C1 = I: 0,04: 0,45. Jelas, karena membran permeabel terhadap ion natrium (P N a =/ 0) dan potensi kesetimbangan ion-ion ini memiliki tanda plus, maka masuknya ion-ion tersebut ke dalam sel sepanjang gradien kimia dan listrik akan mengurangi keelektronegatifan sitoplasma, yaitu. meningkatkan RMP (potensial membran istirahat).
Ketika konsentrasi ion kalium dalam larutan luar meningkat di atas 15 mM, MPP meningkat dan rasio konstanta permeabilitas berubah menuju kelebihan Pk yang lebih signifikan terhadap P Na dan P C1. P k : P Na : P C1 = 1 : 0,025 : 0,4. Dalam kondisi seperti itu, MPP ditentukan hampir secara eksklusif oleh gradien ion kalium, sehingga ketergantungan eksperimental dan teoritis nilai MPP pada logaritma rasio konsentrasi kalium di luar dan di dalam sel mulai bertepatan.
Jadi, adanya perbedaan potensial stasioner antara sitoplasma dan lingkungan luar dalam sel istirahat disebabkan oleh adanya gradien konsentrasi untuk K+, Na+ dan Cl serta perbedaan permeabilitas membran terhadap ion-ion tersebut. Peran utama dalam pembentukan MPP dimainkan oleh difusi ion kalium dari sel ke dalam larutan eksternal. Bersamaan dengan itu, MPP juga ditentukan oleh potensial kesetimbangan natrium dan klorin, dan kontribusi masing-masing ditentukan oleh hubungan antara permeabilitas membran plasma sel terhadap ion-ion tersebut.
Semua faktor yang disebutkan di atas merupakan apa yang disebut komponen ionik RMP (potensial membran istirahat). Karena potensial kesetimbangan kalium dan natrium tidak sama dengan MPP. sel harus menyerap Na+ dan kehilangan K+. Keteguhan konsentrasi ion-ion ini di dalam sel dipertahankan karena kerja Na + K + -ATPase.
Namun, peran pompa ion ini tidak terbatas pada mempertahankan gradien natrium dan kalium. Diketahui bahwa pompa natrium bersifat elektrogenik dan ketika berfungsi, aliran bersih muatan positif muncul dari sel ke dalam cairan ekstraseluler, menyebabkan peningkatan elektronegativitas sitoplasma terhadap lingkungan. Elektrogenisitas pompa natrium terungkap dalam percobaan pada neuron moluska raksasa. Injeksi elektroforesis ion Na+ ke dalam tubuh satu neuron menyebabkan hiperpolarisasi membran, di mana MPP secara signifikan lebih rendah daripada potensial kesetimbangan kalium. Hiperpolarisasi ini dilemahkan dengan menurunkan suhu larutan di mana sel berada dan ditekan oleh ouabain inhibitor Na + , K + -ATPase spesifik.
Dari penjelasan di atas dapat disimpulkan bahwa MPP dapat dibagi menjadi dua komponen - "ionik" Dan "metabolik". Komponen pertama bergantung pada gradien konsentrasi ion dan permeabilitas membrannya. Yang kedua, “metabolik”, disebabkan oleh transpor aktif natrium dan kalium dan memiliki efek ganda MPP. Di satu sisi, pompa natrium mempertahankan gradien konsentrasi antara sitoplasma dan lingkungan luar. Di sisi lain, karena bersifat elektrogenik, pompa natrium mempunyai efek langsung pada MPP. Kontribusinya terhadap nilai MPP tergantung pada kepadatan arus “pemompaan” (arus per satuan luas permukaan membran sel) dan resistansi membran.
Potensi aksi membran
bidang_teks
bidang_teks
panah_ke atas
Jika iritasi diterapkan pada saraf atau otot di atas ambang eksitasi, maka MPP saraf atau otot akan cepat menurun dan dalam waktu singkat (milidetik) membran akan terisi ulang: bagian dalamnya akan menjadi bermuatan positif relatif terhadap yang bagian luar. Ini perubahan MPP jangka pendek yang terjadi selama eksitasi sel, yang pada layar osiloskop berbentuk puncak tunggal, disebut potensial aksi membran (MPD).
IVD pada jaringan saraf dan otot terjadi bila terjadi penurunan nilai mutlak MPP (depolarisasi membran) sampai nilai kritis tertentu disebut ambang generasi MTD. Pada serabut saraf cumi-cumi raksasa, IVD adalah 60 mV. Ketika membran didepolarisasi hingga -45 mV (ambang batas pembangkitan IVD), terjadi IVD (Gbr. 1.15).
Beras. 1.15 Potensi aksi serabut saraf (A) dan perubahan konduksi membran ion natrium dan kalium (B).Pada saat terjadinya IVD pada akson cumi, resistansi membran berkurang 25 kali lipat, dari 1000 menjadi 40 Ohm.cm 2, sedangkan kapasitansi listrik tidak berubah. Penurunan resistensi membran ini disebabkan oleh peningkatan permeabilitas ionik membran saat eksitasi.
Pada amplitudonya (100-120 mV), MAP (Membrane Action Potensial) lebih tinggi 20-50 mV dibandingkan MPP (Resting Membrane Potensial). Dengan kata lain, sisi dalam membran untuk waktu yang singkat menjadi bermuatan positif dibandingkan dengan sisi luarnya - "melampaui" atau pembalikan biaya.
Dari persamaan Goldman dapat disimpulkan bahwa hanya peningkatan permeabilitas membran terhadap ion natrium yang dapat menyebabkan perubahan potensial membran. Nilai E k selalu lebih kecil dari nilai MPP, sehingga peningkatan permeabilitas membran untuk K+ akan meningkatkan nilai absolut MPP. Potensi kesetimbangan natrium memiliki tanda plus, sehingga peningkatan tajam permeabilitas membran terhadap kation ini menyebabkan pengisian ulang membran.
Selama IVD, permeabilitas membran terhadap ion natrium meningkat. Perhitungan menunjukkan bahwa jika dalam keadaan diam perbandingan konstanta permeabilitas membran untuk K + , Na + dan SG adalah 1: 0,04: 0,45, maka pada MTD - P ke: P Na: P = 1: 20: 0,45 . Akibatnya, dalam keadaan eksitasi, membran serabut saraf tidak hanya kehilangan permeabilitas ion selektifnya, tetapi sebaliknya, dari permeabel selektif terhadap ion kalium saat istirahat, menjadi permeabel selektif terhadap ion natrium. Peningkatan permeabilitas natrium pada membran dikaitkan dengan terbukanya saluran natrium berpintu tegangan.
Mekanisme yang menjamin terbuka dan tertutupnya saluran ion disebut gerbang kanal. Merupakan kebiasaan untuk membedakannya pengaktifan(m) dan inaktivasi(h) gerbang. Saluran ion dapat berada dalam tiga keadaan utama: tertutup (gerbang m tertutup; gerbang h terbuka), terbuka (gerbang m dan h terbuka) dan tidak aktif (gerbang m terbuka, gerbang h tertutup) (Gambar 1.16) .
Beras. 1.16 Diagram posisi gerbang aktivasi (m) dan inaktivasi (h) saluran natrium, sesuai dengan keadaan tertutup (istirahat, A), terbuka (aktivasi, B) dan tidak aktif (C).
Depolarisasi membran, yang disebabkan oleh stimulus yang mengiritasi, misalnya arus listrik, membuka gerbang m saluran natrium (transisi dari keadaan A ke B) dan memastikan munculnya aliran muatan positif ke dalam - ion natrium. Hal ini menyebabkan depolarisasi membran lebih lanjut, yang pada gilirannya meningkatkan jumlah saluran natrium terbuka dan, oleh karena itu, meningkatkan permeabilitas natrium membran. Terjadi depolarisasi “regeneratif” pada membran, akibatnya potensial sisi dalam membran cenderung mencapai potensial kesetimbangan natrium.
Penyebab terhentinya pertumbuhan IVD (Potensial aksi membran) dan repolarisasi membran sel adalah:
A) Peningkatan depolarisasi membran, mis. ketika E m -» E Na, mengakibatkan penurunan gradien elektrokimia ion natrium, sama dengan E m -> E Na. Dengan kata lain, kekuatan “mendorong” natrium ke dalam sel berkurang;
B) Depolarisasi membran menimbulkan proses inaktivasi saluran natrium (penutupan gerbang-h; keadaan saluran B), yang menghambat pertumbuhan permeabilitas natrium membran dan menyebabkan penurunannya;
V) Depolarisasi membran meningkatkan permeabilitasnya terhadap ion kalium. Arus kalium yang keluar cenderung menggeser potensial membran menuju potensial kesetimbangan kalium.
Mengurangi potensi elektrokimia ion natrium dan menonaktifkan saluran natrium mengurangi besarnya arus natrium yang masuk. Pada titik waktu tertentu, besarnya arus natrium yang masuk dibandingkan dengan peningkatan arus keluar - pertumbuhan IVD terhenti. Ketika total arus keluar melebihi arus masuk, repolarisasi membran dimulai, yang juga bersifat regeneratif. Permulaan repolarisasi menyebabkan penutupan gerbang aktivasi (m), yang mengurangi permeabilitas natrium membran, mempercepat repolarisasi, dan yang terakhir meningkatkan jumlah saluran tertutup, dll.
Fase repolarisasi IVD di beberapa sel (misalnya, di kardiomiosit dan beberapa sel otot polos) dapat melambat, membentuk dataran AP disebabkan oleh perubahan kompleks waktu arus masuk dan keluar melalui membran. Akibat IVD, hiperpolarisasi dan/atau depolarisasi membran dapat terjadi. Inilah yang disebut menelusuri potensi. Hiperpolarisasi jejak memiliki sifat ganda: ionik Dan metaboliksaya menempa. Yang pertama disebabkan oleh fakta bahwa permeabilitas kalium dalam serabut saraf membran tetap meningkat selama beberapa waktu (puluhan bahkan ratusan milidetik) setelah pembentukan IVD dan menggeser potensial membran menuju potensial keseimbangan kalium. Jejak hiperpolarisasi setelah stimulasi ritmik sel dikaitkan terutama dengan aktivasi pompa natrium elektrogenik, karena akumulasi ion natrium di dalam sel.
Alasan depolarisasi yang berkembang setelah pembentukan MAP (Potensi Aksi Membran) adalah akumulasi ion kalium pada permukaan luar membran. Yang terakhir, sebagai berikut dari persamaan Goldman, menyebabkan peningkatan RMP (potensial membran istirahat).
Inaktivasi saluran natrium dikaitkan dengan sifat penting dari serabut saraf yang disebutsifat tahan api .
Selama absokecapi periode refraktori serabut saraf benar-benar kehilangan kemampuan untuk dirangsang oleh rangsangan dengan kekuatan apa pun.
Relatif sifat tahan api, mengikuti yang absolut, ditandai dengan ambang batas yang lebih tinggi untuk terjadinya MTD (Potensi aksi membran).
Gagasan tentang proses membran yang terjadi selama eksitasi serabut saraf menjadi dasar pemahaman dan fenomena tersebut akomodasi. Dasar akomodasi jaringan pada tingkat peningkatan arus iritasi yang rendah adalah peningkatan ambang eksitasi, yang melampaui depolarisasi membran yang lambat. Peningkatan ambang eksitasi hampir seluruhnya ditentukan oleh inaktivasi saluran natrium. Peran peningkatan permeabilitas kalium membran dalam perkembangan akomodasi menyebabkan penurunan resistensi membran. Karena penurunan resistensi, laju depolarisasi membran menjadi lebih lambat. Laju akomodasi semakin tinggi, semakin besar jumlah saluran natrium pada potensial istirahat yang berada dalam keadaan tidak aktif, semakin tinggi laju perkembangan inaktivasi dan semakin tinggi permeabilitas kalium membran.
Melakukan eksitasi
bidang_teks
bidang_teks
panah_ke atas
Konduksi eksitasi sepanjang serabut saraf dilakukan karena arus lokal antara bagian membran yang tereksitasi dan istirahat. Urutan kejadian dalam kasus ini disajikan sebagai berikut.
Ketika rangsangan titik diterapkan pada serabut saraf di bagian membran yang sesuai, timbul potensial aksi. Sisi dalam membran pada suatu titik tertentu ternyata bermuatan positif relatif terhadap sisi dalam membran yang diam. Arus muncul antara titik-titik serat yang mempunyai potensial berbeda. (arus lokal), diarahkan dari tereksitasi (tanda (+) di bagian dalam membran) ke tidak tereksitasi (tanda (-) di bagian dalam membran) ke bagian serat. Arus ini mempunyai efek depolarisasi pada membran serat di area istirahat, dan ketika tingkat depolarisasi membran kritis di area ini tercapai, terjadi MAP (Potensi Aksi Membran). Proses ini secara berurutan menyebar ke seluruh area serabut saraf.
Pada beberapa sel (neuron, otot polos), IVD tidak bersifat natrium, tetapi disebabkan oleh masuknya ion Ca 2+ melalui saluran kalsium yang bergantung pada tegangan. Pada kardiomiosit, pembentukan IVD dikaitkan dengan aliran natrium dan natrium-kalsium yang masuk.
Topik ini akan membahas dua kation - natrium (Na) dan kalium (K). Berbicara tentang anion, mari kita perhatikan bahwa sejumlah anion terletak di sisi luar dan dalam membran sel.
Bentuk sel bergantung pada jaringan mana sel tersebut berada. Dengan caranya sendiri membentuk sel dapat berupa:
· berbentuk silinder dan kubik (sel kulit);
· berbentuk cakram (eritrosit);
· bulat (ovula);
· fusiform (otot polos);
· stellata dan piramidal (sel saraf);
· tidak mempunyai bentuk permanen - amoeboid (leukosit).
Sel itu mempunyai nomor properti: ia mencari makan, tumbuh, berkembang biak, memulihkan diri, beradaptasi dengan lingkungannya, bertukar energi dan zat dengannya lingkungan, menjalankan fungsi bawaannya (tergantung pada jaringan tempat sel tersebut berada). Selain itu, sel memiliki sifat dpt dirangsang.
Sifat dpt dirangsang – Ini adalah kemampuan sel, sebagai respons terhadap rangsangan, untuk berpindah dari keadaan istirahat ke keadaan aktif.
Iritasi dapat berasal dari lingkungan luar atau berasal dari dalam sel. Rangsangan yang menimbulkan eksitasi dapat berupa : rangsangan listrik, kimia, mekanik, suhu dan rangsangan lainnya.
Sebuah sel dapat berada dalam dua keadaan utama: diam dan tereksitasi. Istirahat dan eksitasi sel disebut - potensial membran istirahat dan potensial aksi membran.
Ketika sel tidak mengalami iritasi apapun, maka sel berada dalam keadaan istirahat. Istirahat sel disebut juga potensial membran istirahat (RMP).
Saat diam, permukaan bagian dalam membrannya bermuatan negatif, dan permukaan luarnya bermuatan positif. Hal ini dijelaskan oleh fakta bahwa di dalam sel terdapat banyak anion dan sedikit kation, sedangkan di belakang sel, sebaliknya, kation mendominasi.
Karena sel berisi muatan listrik, maka listrik yang dihasilkannya dapat diukur. Potensial membran istirahat adalah: - 70 mV (minus 70, karena terdapat muatan negatif di dalam sel). Nilai ini bersyarat, karena setiap sel dapat memiliki nilai potensial istirahatnya sendiri.
Saat istirahat, pori-pori membran terbuka terhadap ion kalium dan tertutup terhadap ion natrium. Artinya ion kalium dapat dengan mudah keluar masuk sel. Ion natrium tidak dapat masuk ke dalam sel karena pori-pori membran tertutup. Tetapi sejumlah kecil ion natrium menembus ke dalam sel karena mereka tertarik oleh sejumlah besar anion yang terletak di permukaan bagian dalam membran (muatan yang berlawanan tarik-menarik). Pergerakan ion ini adalah pasif , karena tidak memerlukan energi.
Agar sel berfungsi normal, nilai MPP-nya harus tetap pada tingkat yang konstan. Namun pergerakan ion natrium dan kalium melalui membran menyebabkan fluktuasi nilai ini, yang dapat menyebabkan penurunan atau peningkatan nilai: - 70 mV.
Agar nilai MPP relatif konstan disebut pompa natrium-kalium . Fungsinya adalah mengeluarkan ion natrium dari sel, dan memompa ion kalium ke dalam sel. Ini adalah rasio tertentu ion natrium dan kalium di dalam sel dan di belakang sel yang menciptakan nilai MPP yang diperlukan. Pengoperasian pompa adalah mekanisme aktif , karena memerlukan energi.
Sumber energi dalam sel adalah ATP. ATP menyediakan energi hanya ketika dipecah menjadi asam yang lebih sederhana - ADP, dengan partisipasi wajib enzim ATPase dalam reaksi:
ATP + enzim ATPase ADP + energi
Semua sel hidup memiliki kemampuan, di bawah pengaruh rangsangan, untuk berpindah dari keadaan istirahat fisiologis ke keadaan aktif atau eksitasi.
Perangsangan adalah kompleks perubahan listrik, kimia, dan fungsional aktif pada jaringan yang dapat dirangsang (saraf, otot, atau kelenjar), yang dengannya jaringan merespons pengaruh eksternal. Peran penting dalam eksitasi dimainkan oleh proses kelistrikan yang memastikan konduksi eksitasi sepanjang serabut saraf dan membawa jaringan ke keadaan aktif (bekerja).
Potensi membran
Sel hidup memiliki sifat penting: permukaan bagian dalam sel selalu bermuatan negatif dibandingkan dengan sisi luarnya. Antara permukaan luar sel, bermuatan elektropositif terhadap protoplasma, dan sisi dalam membran sel, terdapat beda potensial yang berkisar antara 60-70 mV. Menurut P.G.Kostyuk (2001), sel saraf perbedaan ini berkisar antara 30 hingga 70 mV. Beda potensial antara sisi luar dan dalam membran sel disebut potensi membran, atau potensi istirahat(Gbr. 2.1).
Potensial membran istirahat terdapat pada membran selama sel masih hidup dan menghilang ketika sel mati. L. Galvani menunjukkan kembali pada tahun 1794 bahwa jika Anda merusak saraf atau otot dengan melakukan persilangan dan dengan menempelkan elektroda-elektroda yang dihubungkan dengan galvanometer pada bagian yang rusak dan pada tempat yang rusak, maka galvanometer akan menunjukkan arus yang selalu mengalir dari bagian jaringan yang tidak rusak ke tempat sayatan. Dia menyebut aliran ini sebagai arus diam. Dalam esensi fisiologisnya, arus istirahat dan potensial membran istirahat adalah satu dan sama. Beda potensial yang diukur dalam percobaan ini adalah 30-50 mV, karena ketika jaringan rusak, sebagian arus dialihkan ke ruang antar sel dan cairan di sekitar struktur yang diteliti. Beda potensial dapat dihitung dengan menggunakan rumus Nernst:
dimana R adalah konstanta gas, T adalah suhu absolut, F adalah bilangan Faraday, [K] int. dan [K] adv. - konsentrasi kalium di dalam dan di luar sel.

Beras. 2.1.
Penyebab potensial istirahat umum terjadi pada semua sel. Antara protoplasma sel dan lingkungan ekstraseluler terdapat distribusi ion yang tidak merata (asimetri ion). Komposisi darah manusia ditinjau dari keseimbangan garam menyerupai komposisi air laut. Lingkungan ekstraseluler di sistem saraf pusat juga banyak mengandung natrium klorida. Komposisi ionik sitoplasma sel lebih buruk. Di dalam sel terdapat ion Na+ 8-10 kali lebih sedikit dan ion C 50 kali lebih sedikit!". Kation utama sitoplasma adalah K+. Konsentrasinya di dalam sel 30 kali lebih tinggi daripada di lingkungan ekstraseluler, dan kira-kira sama dengan konsentrasi Na ekstraseluler. Ion lawan K + dalam sitoplasma adalah anion organik, khususnya anion aspartat, histamin dan asam amino lainnya. Asimetri seperti itu merupakan pelanggaran keseimbangan termodinamika. Untuk memulihkannya, kalium ion harus meninggalkan sel secara bertahap, dan ion natrium harus meninggalkan sel. Namun, hal ini tidak terjadi.
Kendala pertama dalam menyamakan perbedaan konsentrasi ion adalah membran plasma sel. Terdiri dari dua lapis molekul fosfolipid, bagian dalam ditutupi dengan lapisan molekul protein, dan bagian luar dengan lapisan karbohidrat (mukopolisakarida). Beberapa protein seluler tertanam langsung di lapisan ganda lipid. Ini adalah protein internal.
Protein membran semua sel dibagi menjadi lima kelas: pompa, saluran, reseptor, enzim Dan protein struktural. Pompa berfungsi untuk memindahkan ion dan molekul melawan gradien konsentrasi menggunakan energi metabolisme. Saluran protein, atau pori-pori, memberikan permeabilitas selektif (difusi) melalui membran ion dan molekul yang sesuai dengan ukurannya. Protein reseptor memiliki spesifisitas tinggi, mereka mengenali dan mengikat, menempel pada membran, banyak jenis molekul yang diperlukan untuk kehidupan sel pada waktu tertentu. Enzim mempercepat aliran reaksi kimia pada permukaan membran. Protein struktural memastikan koneksi sel ke organ dan pemeliharaan struktur subseluler.
Semua protein ini spesifik, tetapi tidak sepenuhnya spesifik. Dalam kondisi tertentu, protein tertentu secara bersamaan dapat menjadi pompa, enzim, dan reseptor. Melalui saluran membran, molekul air, serta ion yang sesuai dengan ukuran pori-pori, masuk dan keluar sel. Permeabilitas membran untuk kation yang berbeda tidak sama dan berubah pada keadaan fungsional jaringan yang berbeda. Saat istirahat, membran 25 kali lebih permeabel terhadap ion kalium dibandingkan ion natrium, dan ketika tereksitasi, permeabilitas natrium kira-kira 20 kali lebih tinggi daripada kalium. Saat istirahat, konsentrasi kalium yang sama di sitoplasma dan natrium di lingkungan ekstraseluler harus menghasilkan jumlah muatan positif yang sama di kedua sisi membran. Tetapi karena permeabilitas ion kalium 25 kali lebih tinggi, kalium, meninggalkan sel, membuat permukaannya semakin bermuatan positif dibandingkan dengan sisi dalam membran, di dekatnya terdapat molekul aspartik, histamin, dan lain-lain yang bermuatan negatif, terlalu besar. untuk pori-pori membran, semakin banyak asam amino yang terakumulasi yang “melepaskan” kalium ke luar sel, tetapi “mencegah” keluarnya jauh karena muatan negatifnya. Muatan negatif terakumulasi di bagian dalam membran, dan muatan positif di bagian luar. Perbedaan potensial muncul. Arus difusi ion natrium ke dalam protoplasma dari cairan ekstraseluler menjaga perbedaan ini pada tingkat 60-70 mV, mencegah peningkatannya. Arus difusi ion natrium dalam keadaan diam 25 kali lebih lemah dibandingkan arus berlawanan ion kalium. Ion natrium, yang menembus ke dalam sel, mengurangi potensi istirahat, membiarkannya tetap pada tingkat tertentu. Dengan demikian, nilai potensial istirahat sel otot dan saraf, serta serabut saraf, ditentukan oleh perbandingan jumlah ion kalium bermuatan positif yang berdifusi per satuan waktu dari sel ke luar, dan ion natrium bermuatan positif yang berdifusi melalui sel. membran dalam arah yang berlawanan. Semakin tinggi rasio ini, semakin besar potensi istirahatnya, dan sebaliknya.
Kendala kedua yang menjaga beda potensial pada tingkat tertentu adalah pompa natrium-kalium (Gbr. 2.2). Ini disebut natrium-kalium atau ionik, karena secara aktif menghilangkan (memompa) ion natrium yang menembus ke dalamnya dari protoplasma dan memasukkan (memompa) ion kalium ke dalamnya. Sumber energi untuk pengoperasian pompa ion adalah pemecahan ATP (adenosin trifosfat), yang terjadi di bawah pengaruh enzim adenosin trifosfatase, terlokalisasi di membran sel dan diaktifkan oleh ion yang sama, yaitu kalium dan natrium (natrium). -ATPase yang bergantung pada kalium).

Beras. 2.2.
Ini adalah protein berukuran besar, melebihi ketebalan membran sel. Molekul protein ini, menembus membran, mengikat sebagian besar natrium dan ATP di bagian dalam, dan kalium serta berbagai inhibitor seperti glikosida di bagian luar. Dalam hal ini, terjadi arus membran. Berkat arus ini, arah transpor ion yang tepat dapat dipastikan. Transfer ion terjadi dalam tiga tahap. Pertama, ion bergabung dengan molekul pembawa untuk membentuk kompleks pengangkut ion. Kompleks ini kemudian melewati membran atau mentransfer muatan melewatinya. Akhirnya, ion dilepaskan dari pembawa di sisi berlawanan dari membran. Pada saat yang sama, proses serupa terjadi, mengangkut ion ke arah yang berlawanan. Jika pompa mentransfer satu ion natrium ke satu ion kalium, maka pompa hanya mempertahankan gradien konsentrasi di kedua sisi membran, tetapi tidak berkontribusi pada penciptaan potensial membran. Untuk memberikan kontribusi ini, pompa ion harus mengangkut natrium dan kalium dengan perbandingan 3:2, yaitu untuk setiap 2 ion kalium yang masuk ke dalam sel, pompa harus mengeluarkan 3 ion natrium dari sel. Bekerja pada beban maksimum, setiap pompa mampu memompa sekitar 130 ion kalium dan 200 ion natrium per detik melalui membran. Ini kecepatan tertinggi. Dalam kondisi nyata, pengoperasian masing-masing pompa disesuaikan dengan kebutuhan sel. Kebanyakan neuron memiliki antara 100 dan 200 pompa ion per mikron persegi permukaan membran. Akibatnya, membran sel saraf mana pun mengandung 1 juta pompa ion yang mampu menggerakkan hingga 200 juta ion natrium per detik.
Dengan demikian, potensial membran (potensial istirahat) tercipta sebagai hasil mekanisme pasif dan aktif. Derajat partisipasi mekanisme tertentu pada sel yang berbeda tidak sama, yang berarti bahwa potensial membran pada struktur yang berbeda mungkin berbeda. Aktivitas pompa mungkin bergantung pada diameter serabut saraf: semakin tipis serat, semakin tinggi rasio ukuran permukaan terhadap volume sitoplasma; oleh karena itu, aktivitas pompa diperlukan untuk mempertahankan perbedaan konsentrasi ion. di permukaan dan di dalam serat harus lebih besar. Dengan kata lain, potensial membran mungkin bergantung pada strukturnya jaringan saraf, dan karena itu pada tujuan fungsionalnya. Polarisasi listrik pada membran adalah syarat utama rangsangan sel. Ini adalah kesiapannya yang terus-menerus untuk bertindak. Ini stoknya energi potensial sel yang dapat dia gunakan seandainya sistem saraf memerlukan respons segera.







