Размената на супстанции кои влегуваат во клетката или се ослободуваат од неа надвор, како и размената на различни сигнали со микро- и макросредината, се случува преку надворешната мембрана на клетката. Како што е познато, клеточната мембрана е липиден двослој во кој се вградени различни протеински молекули кои делуваат како специјализирани рецептори, јонски канали, уреди кои активно транспортираат или отстрануваат разни хемикалии, меѓуклеточни контакти итн. Во здравите еукариотски клетки, фосфолипидите се дистрибуираат во мембраната асиметрично: надворешната површина се состои од сфингомиелин и фосфатидилхолин, внатрешната површина - од фосфатидилсерин и фосфатидилетаноламин. Одржувањето на таква асиметрија бара трошење на енергија. Затоа, во случај на оштетување на клетките, инфекција или енергетски глад, надворешната површина на мембраната се збогатува со фосфолипиди кои се невообичаени за неа, што станува сигнал за другите клетки и ензими за оштетување на клетките со соодветна реакција на ова. Најважна улога игра растворливата форма на фосфолипаза А2, која ја разградува арахидонската киселина и создава лизоформи од горенаведените фосфолипиди. Арахидонската киселина е ограничувачка врска за создавање на воспалителни медијатори како што се еикосаноидите, а заштитните молекули - пентраксини (C-реактивен протеин (CRP), прекурсори на амилоидни протеини) - се прикачени на лизоформите во мембраната, проследено со активирање на комплементот систем по класичната патека и уништување на клетките.
Структурата на мембраната помага да се зачуваат карактеристиките на внатрешната средина на клетката, нејзините разлики од надворешната средина. Ова е обезбедено со селективната пропустливост на клеточната мембрана и постоењето на активни транспортни механизми во неа. Нивното нарушување како резултат на директно оштетување, на пример, од тетродотоксин, уабаин, тетраетиламониум или во случај на недоволно снабдување со енергија на соодветните „пумпи“ доведува до нарушување на електролитниот состав на клетката, промени во неговиот метаболизам, нарушување на специфични функции - контракција, спроведување на импулси на побудување итн. Нарушувањето на клеточните јонски канали (калциум, натриум, калиум и хлорид) кај луѓето, исто така, може генетски да се определи со мутација на гените одговорни за структурата на овие канали. Таканаречените каналопатии предизвикуваат наследни болести на нервниот, мускулниот и дигестивниот систем. Прекумерното влегување на вода во клетката може да доведе до нејзино кинење - цитолиза - поради перфорација на мембраната кога се активира комплементот или напад од цитотоксични лимфоцити и природни клетки убијци.
Клеточната мембрана има многу рецептори вградени во неа - структури кои, кога се комбинираат со соодветните специфични сигнални молекули (лиганди), пренесуваат сигнал внатре во клетката. Ова се случува преку различни регулаторни каскади кои се состојат од ензимски активни молекули кои секвенцијално се активираат и на крајот придонесуваат за имплементација на различни клеточни програми, како што се растот и пролиферацијата, диференцијацијата, подвижноста, стареењето и клеточната смрт. Регулаторните каскади се доста бројни, но нивниот број сè уште не е целосно утврден. Системот на рецептори и регулаторни каскади поврзани со нив, исто така, постои внатре во клетката; тие создаваат специфична регулаторна мрежа со точки на концентрација, дистрибуција и избор на понатамошната патека на сигналот во зависност од функционалната состојба на клетката, фазата на нејзиниот развој и истовременото дејство на сигналите од другите рецептори. Резултатот од ова може да биде инхибиција или зајакнување на сигналот, насочувајќи го по различна регулаторна патека. И рецепторскиот апарат и патиштата за трансдукција на сигналот низ регулаторните каскади, на пример до јадрото, може да се нарушат како резултат на генетски дефект што се јавува како вроден дефект на ниво на организмот или поради соматска мутација во специфичен тип на клетка. Овие механизми можат да бидат оштетени од инфективни агенси, токсини, а исто така да се променат во текот на процесот на стареење. Последната фаза на ова може да биде нарушување на функциите на клетката, процесите на нејзино размножување и диференцијација.
На површината на клетките има и молекули кои играат важна улога во процесите на меѓуклеточната интеракција. Тие може да вклучуваат протеини за адхезија на клетките, антигени за хистокомпатибилност, ткивни специфични, диференцирачки антигени итн. Промените во составот на овие молекули предизвикуваат нарушување на меѓуклеточните интеракции и може да предизвикаат активирање на соодветни механизми за елиминација на таквите клетки, бидејќи тие претставуваат одредена опасност за интегритетот на телото како резервоар на инфекција, особено вирусна, или како потенцијални иницијатори за раст на туморот.
Повреда на снабдувањето со енергија на ќелијата
Изворот на енергија во клетката е храната, по чие разградување енергијата се ослободува во финални материи. Главното место за производство на енергија се митохондриите, во кои супстанциите се оксидираат со помош на ензими на респираторниот синџир. Оксидацијата е главниот снабдувач на енергија, бидејќи како резултат на гликолизата, не повеќе од 5% од енергијата се ослободува од иста количина на оксидациони супстрати (гликоза), во споредба со оксидацијата. Околу 60% од енергијата ослободена за време на оксидацијата се акумулира со оксидативна фосфорилација во високо-енергетски фосфати (ATP, креатин фосфат), а остатокот се троши како топлина. Последователно, клетката ги користи високо-енергетските фосфати за процеси како што се работата на пумпата, синтезата, поделбата, движењето, секрецијата итн. Постојат три механизми, чие оштетување може да предизвика нарушување во снабдувањето со енергија на клетката: првиот е механизам на синтеза на енергетскиот метаболизам ензими, вториот е механизмот на оксидативна фосфорилација, третиот е механизмот на употреба на енергија.
Нарушувањето на транспортот на електрони во митохондријалниот респираторен синџир или раздвојувањето на оксидацијата и фосфорилацијата на ADP со губење на протонскиот потенцијал, движечката сила за генерирање на АТП, доведува до слабеење на оксидативната фосфорилација на таков начин што најголемиот дел од енергијата се троши како топлина и се намалува бројот на високоенергетски соединенија. Одвојувањето на оксидацијата и фосфорилацијата под влијание на адреналин се користи од клетките на хомеотермните организми за да се зголеми производството на топлина додека се одржува константна телесна температура за време на ладењето или се зголемува за време на треска. Значајни промени во митохондријалната структура и енергетскиот метаболизам се забележани кај тиреотоксикозата. Овие промени се првично реверзибилни, но по одреден момент тие стануваат неповратни: митохондриите се фрагментираат, се распаѓаат или отекуваат, губат кристи, се претвораат во вакуоли и на крајот акумулираат супстанции како што се хијалин, феритин, калциум, липофусцин. Кај пациенти со скорбут, митохондриите се спојуваат за да формираат хондриосфери, веројатно поради оштетување на мембраната од соединенија на пероксид. Значително оштетување на митохондриите се јавува под влијание на јонизирачко зрачење при трансформацијата на нормална клетка во малигна.
Митохондриите се моќно складиште на јони на калциум, каде што неговата концентрација е неколку реда на големина поголема од онаа во цитоплазмата. Кога митохондриите се оштетени, калциумот влегува во цитоплазмата, предизвикувајќи активирање на протеинази со оштетување на интрацелуларните структури и нарушување на функциите на соодветната клетка, на пример, контрактури на калциум или дури и „калциумска смрт“ во невроните. Како резултат на нарушување на функционалниот капацитет на митохондриите, нагло се зголемува формирањето на соединенија на пероксид од слободни радикали, кои имаат многу висока реактивност и затоа ги оштетуваат важните компоненти на клетката - нуклеинските киселини, протеините и липидите. Оваа појава се забележува при таканаречениот оксидативен стрес и може да има негативни последици за постоењето на клетката. Така, оштетувањето на надворешната мембрана на митохондриите е придружено со ослободување во цитоплазмата на супстанции содржани во меѓумембранскиот простор, првенствено цитохром Ц и некои други биолошки активни супстанции, кои предизвикуваат верижни реакции кои предизвикуваат програмирана клеточна смрт - апоптоза. Со оштетување на митохондријалната ДНК, реакциите на слободните радикали ги искривуваат генетските информации неопходни за формирање на одредени ензими на респираторниот синџир, кои се произведуваат специјално во митохондриите. Ова доведува до уште поголемо нарушување на оксидативните процеси. Генерално, сопствениот генетски апарат на митохондриите, во споредба со генетскиот апарат на јадрото, е помалку заштитен од штетните влијанија кои можат да ги променат генетските информации кодирани во него. Како резултат на тоа, дисфункција на митохондриите се јавува во текот на животот, на пример, за време на процесот на стареење, за време на малигната трансформација на клетката, како и на позадината на наследни митохондријални заболувања поврзани со мутација на митохондријалната ДНК во јајцето. Во моментов, опишани се над 50 митохондријални мутации кои предизвикуваат наследни дегенеративни заболувања на нервниот и мускулниот систем. Тие се пренесуваат на детето исклучиво од мајката, бидејќи митохондриите на спермата не се дел од зиготот и, соодветно, новиот организам.
Повреда на зачувување и пренос на генетски информации
Јадрото на клетката содржи најголем дел од генетските информации и на тој начин го обезбедува неговото нормално функционирање. Преку селективна генска експресија, тој ја координира клеточната активност за време на интерфазата, складира генетски информации и повторно создава и пренесува генетски материјал за време на клеточната делба. Во јадрото се случува репликација на ДНК и транскрипција на РНК. Разни патогени фактори, како што се ултравиолетово и јонизирачко зрачење, оксидација на слободните радикали, хемикалии, вируси, може да ја оштетат ДНК. Се проценува дека на секоја клетка на топлокрвно животно му треба 1 ден. губи повеќе од 10.000 бази. Тука треба да додадеме прекршувања при копирање при поделба. Доколку овие оштетувања опстојуваат, клетката не би можела да преживее. Заштитата лежи во постоењето на моќни системи за поправка, како што се ултравиолетовите ендонуклеаза, поправка на репликација и рекомбинација поправка системи, кои го заменуваат оштетување на ДНК. Генетските дефекти во системите за поправка предизвикуваат развој на болести предизвикани од зголемена чувствителност на фактори кои ја оштетуваат ДНК. Ова е xeroderma pigmentosum, како и некои синдроми на забрзано стареење, придружени со зголемена тенденција за развој на малигни тумори.
Системот за регулирање на процесите на репликација на ДНК, транскрипција на гласник РНК (мРНК) и преведување на генетските информации од нуклеинските киселини во структурата на протеините е доста сложен и на повеќе нивоа. Покрај регулаторните каскади кои го активираат дејството на факторите на транскрипција со вкупен број од над 3000, кои активираат одредени гени, постои и регулаторен систем на повеќе нивоа со посредство на мали молекули на РНК (интерферирачка РНК; РНКи). Човечкиот геном, кој се состои од приближно 3 милијарди пурински и пиримидински бази, содржи само 2% од структурните гени одговорни за синтезата на протеините. Останатите обезбедуваат синтеза на регулаторни РНК, кои, истовремено со факторите на транскрипција, ја активираат или блокираат работата на структурните гени на ниво на ДНК во хромозомите или влијаат на процесите на транслација на гласник РНК (mRNA) за време на формирањето на полипептидна молекула во цитоплазмата. Повреда на генетските информации може да се случи и на ниво на структурни гени и на регулаторниот дел на ДНК со соодветни манифестации во форма на разни наследни болести.
Неодамна, големо внимание привлекуваат промените во генетскиот материјал кои настануваат при индивидуалниот развој на организмот и се поврзани со инхибиција или активирање на одредени делови од ДНК и хромозоми поради нивната метилација, ацетилација и фосфорилација. Овие промени опстојуваат долго време, понекогаш во текот на целиот животен век на организмот од ембриогенеза до старост и се нарекуваат епигеномска наследност.
Пролиферацијата на клетките со изменети генетски информации е спречена и со системи (фактори) кои го контролираат митотичниот циклус. Тие комуницираат со протеинските кинази зависни од циклин и нивните каталитички подединици - циклини - и ја блокираат клетката да мине низ целиот митотички циклус, запирајќи ја поделбата на границата помеѓу пресинтетичката и синтетичката фаза (G1/S блок) додека не заврши поправката на ДНК. и ако тоа е невозможно, тие иницираат програмирани клетки на смртта. Овие фактори го вклучуваат генот p53, чија мутација предизвикува губење на контролата врз пролиферацијата на трансформираните клетки; тоа е забележано кај речиси 50% од ракот кај луѓето. Втората контролна точка на митотичниот циклус е на границата G2/M. Овде, правилната дистрибуција на хромозомскиот материјал помеѓу ќерките клетки во митоза или мејоза се контролира со помош на збир на механизми кои го контролираат клеточното вретено, центарот и центромерите (кинетохори). Неефикасноста на овие механизми доведува до нарушување на дистрибуцијата на хромозомите или нивните делови, што се манифестира со отсуство на кој било хромозом во една од ќерките клетки (анеуплоидија), присуство на дополнителен хромозом (полиплоидија), одвојување на дел од хромозом (бришење) и негово пренесување на друг хромозом (транслокација). Ваквите процеси се многу често забележани за време на пролиферацијата на малигно дегенерирани и трансформирани клетки. Ако тоа се случи за време на мејозата со герминативните клетки, тоа доведува или до смрт на фетусот во рана фаза на ембрионалниот развој или до раѓање на организам со хромозомско заболување.
Неконтролираната клеточна пролиферација за време на растот на туморот се јавува како резултат на мутации во гените кои ја контролираат клеточната пролиферација и се нарекуваат онкогени. Меѓу повеќе од 70 моментално познати онкогени, повеќето припаѓаат на компонентите на регулацијата на клеточниот раст, некои се претставени со фактори на транскрипција кои ја регулираат генската активност, како и фактори кои ја инхибираат клеточната делба и раст. Друг фактор што го ограничува прекумерното проширување (ширење) на клетките кои се размножуваат е скратувањето на краевите на хромозомите - теломерите, кои не се во можност целосно да се реплицираат како резултат на чисто стерична интеракција, затоа, по секоја клеточна поделба, теломерите се скратуваат за одреден дел од основите. Така, клетките на возрасен организам што се размножуваат по одреден број поделби (обично од 20 до 100 во зависност од видот на организмот и неговата возраст) ја исцрпуваат должината на теломерите и понатамошната репликација на хромозомот престанува. Овој феномен не се јавува во епителот на спермата, ентероцитите и ембрионските клетки поради присуството на ензимот теломераза, кој ја враќа должината на теломерите по секоја поделба. Во повеќето клетки на возрасни организми, теломеразата е блокирана, но, за жал, таа се активира во клетките на туморот.
Врската помеѓу јадрото и цитоплазмата и транспортот на супстанции во двете насоки се врши преку порите во нуклеарната мембрана со учество на специјални транспортни системи кои трошат енергија. На овој начин енергијата и пластичните материи, сигналните молекули (фактори на транскрипција) се транспортираат до јадрото. Обратен проток ги носи во цитоплазмата молекулите на mRNA и трансферната РНК (tRNA), рибозоми неопходни за синтеза на протеини во клетката. Истиот пат на транспорт на супстанции е исто така својствен за вирусите, особено како што е ХИВ. Тие го пренесуваат својот генетски материјал во јадрото на клетката домаќин со негово понатамошно инкорпорирање во геномот на домаќинот и трансфер на новоформираната вирусна РНК во цитоплазмата за понатамошна синтеза на протеини на нови вирусни честички.
Повреда на процесите на синтеза
Процесите на синтеза на протеини се случуваат во цистерни на ендоплазматскиот ретикулум, тесно поврзани со порите во нуклеарната мембрана, преку кои рибозомите, tRNA и mRNA влегуваат во ендоплазматскиот ретикулум. Овде се врши синтеза на полипептидни синџири, кои последователно ја добиваат својата конечна форма во агрануларниот ендоплазматичен ретикулум и ламеларниот комплекс (комплекс Голги), каде што подлежат на пост-транслациска модификација и се комбинираат со јаглени хидрати и липидни молекули. Новоформираните протеински молекули не остануваат на местото на синтеза, туку преку сложен регулиран процес т.н. протеинкинеза, активно се пренесуваат во тој изолиран дел од клетката каде што ќе ја извршуваат замислената функција. Во овој случај, многу важен чекор е структурирањето на пренесената молекула во соодветна просторна конфигурација способна да ја врши својата вродена функција. Ова структуирање се случува со помош на специјални ензими или на матрица од специјализирани протеински молекули - шаперони, кои и помагаат на протеинската молекула, новоформирана или променета поради надворешно влијание, да ја стекне правилната тродимензионална структура. Во случај на негативен ефект врз клетката, кога постои можност за нарушување на структурата на протеинските молекули (на пример, со зголемување на телесната температура, инфективен процес, интоксикација), концентрацијата на чаперони во клетката се зголемува остро. Затоа, таквите молекули се нарекуваат и стрес протеини, или протеини од топлински шок. Повреда на структурата на протеинската молекула доведува до формирање на хемиски инертни конгломерати, кои се депонираат во клетката или надвор од неа за време на амилоидоза, Алцхајмерова болест итн. Понекогаш претходно структурирана слична молекула може да послужи како матрица, а во оваа случај, ако примарното структурирање се случи погрешно, сите наредни молекули исто така ќе бидат неисправни. Оваа ситуација се јавува кај таканаречените прионски болести (скрапи кај овци, бесни крави, куру, болест Кројцфелд-Јакоб кај луѓето), кога дефект на еден од мембранските протеини на нервната клетка предизвикува последователна акумулација на инертни маси во клетката и нарушување на неговите витални функции.
Нарушување на процесите на синтеза во клетката може да се случи во нејзините различни фази: транскрипција на РНК во јадрото, транслација на полипептиди во рибозомите, пост-транслациска модификација, хиперметилација и гликозилација на беж-молекулата, транспорт и дистрибуција на протеини во клетката и нивно отстранување кон надворешноста. Во овој случај, може да се забележи зголемување или намалување на бројот на рибозоми, распаѓање на полирибозомите, проширување на цистерните на зрнестиот ендоплазматичен ретикулум, губење на рибозоми и формирање на везикули и вакуоли. Така, кога ќе се отруе со бледа гребенка, ензимот РНК полимераза е оштетен, што ја нарушува транскрипцијата. Токсинот на дифтерија, со инактивирање на факторот на издолжување, ги нарушува процесите на превод, предизвикувајќи оштетување на миокардот. Причината за нарушување на синтезата на некои специфични протеински молекули може да бидат инфективни агенси. На пример, вирусите на херпес ја инхибираат синтезата и изразувањето на антигенските молекули на MHC, што им овозможува делумно да ја избегнат имунолошката контрола; бацилите на чума - синтеза на медијатори на акутно воспаление. Појавата на необични протеини може да го запре нивното понатамошно разградување и да доведе до акумулација на инертен, па дури и токсичен материјал. Ова може, до одреден степен, да се олесни со нарушување на процесите на распаѓање.
Нарушување на процесите на распаѓање
Истовремено со синтезата на протеинот во клетката, континуирано се случува негово разградување. Во нормални услови, ова има важно регулаторно и формативно значење, на пример, за време на активирање на неактивни форми на ензими, протеински хормони и протеини од митотичниот циклус. Нормалниот раст и развој на клетките бара фино контролирана рамнотежа помеѓу синтезата и деградацијата на протеините и органелите. Меѓутоа, во процесот на синтеза на протеини, поради грешки во работата на апаратот за синтеза, абнормално структуирање на протеинската молекула и нејзино оштетување од хемиски и бактериски агенси, постојано се формираат прилично голем број неисправни молекули. Според некои проценки, нивниот удел е околу една третина од сите синтетизирани протеини.
Клетките на цицачите имаат неколку главни Начини за уништување на протеини:преку лизозомални протеази (пентид хидролази), калциум-зависни протеинази (ендопептидази) и протеазомски систем. Покрај тоа, постојат и специјализирани протеинази, како што се каспазите. Главната органела во која се јавува деградација на супстанциите во еукариотските клетки е лизозомот, кој содржи бројни хидролитички ензими. Поради процесите на ендоцитоза и разни видови на автофагија во лизозомите и фаголизозомите, се уништуваат и неисправните протеински молекули и цели органели: оштетени митохондрии, делови од плазматската мембрана, некои екстрацелуларни протеини и содржината на секреторните гранули.
Важен механизам за разградување на протеините е протеазомот, мултикаталитичка протеиназа структура со сложена структура локализирана во цитозолот, јадрото, ендоплазматскиот ретикулум и на клеточната мембрана. Овој ензимски систем е одговорен за разградување на оштетените протеини, како и за здрави протеини кои мора да се отстранат за нормална функција на клетките. Во овој случај, протеините што треба да се уништат се прелиминарно комбинирани со специфичен полипептид, убиквитин. Сепак, не-убиквитинираните протеини, исто така, може делумно да се уништат во протеазомите. Разградувањето на протеинските молекули во протеазомите во кратки полипептиди (обработка), проследено со нивно претставување заедно со молекулите на MHC тип I, е важна врска во имунолошката контрола на антигенската хомеостаза во телото. Кога функцијата на протеазом е ослабена, се оштетуваат и се акумулираат непотребни протеини, што го придружува стареењето на клетките. Повреда на разградувањето на протеините зависни од циклин доведува до нарушување на клеточната делба, деградација на секреторните протеини - до развој на цистофиброза. Спротивно на тоа, зголемувањето на функцијата на протеазом го придружува осиромашувањето на телото (СИДА, рак).
Со генетски детерминирани нарушувања на разградувањето на протеините, организмот не е одржлив и умира во раните фази на ембриогенезата. Доколку се наруши разградувањето на мастите или јаглехидратите, се јавуваат болести на складирање (тезауризмоза). Во овој случај, во внатрешноста на клетката се акумулира прекумерна количина на одредени супстанции или производи од нивното нецелосно разградување - липиди, полисахариди, што значително ја оштетува функцијата на клетката. Ова најчесто се забележува кај епителните клетки на црниот дроб (хепатоцити), невроните, фибробластите и макрофагоцитите.
Стекнати нарушувања на процесите на распаѓање на супстанции може да настанат како резултат на патолошки процеси (на пример, протеини, масти, јаглени хидрати и пигментна дегенерација) и се придружени со формирање на необични супстанции. Нарушувањата во системот на лизозомална протеолиза доведуваат до намалена адаптација за време на постот или зголемен стрес и до појава на одредени ендокрини дисфункции - намалени нивоа на инсулин, тироглобулин, цитокини и нивните рецептори. Нарушената деградација на протеините ја забавува стапката на зараснување на раните, предизвикува развој на атеросклероза и влијае на имунолошкиот одговор. Со хипоксија, промени во интрацелуларната pH вредност, повреда на зрачење, која се карактеризира со зголемена пероксидација на мембранските липиди, како и под влијание на лизозомотропни супстанции - бактериски ендотоксини, метаболити на токсични габи (спорофусарин), кристали на силициум оксид - стабилност на мембраната на лизозомот промени, активираните лизозомални ензими се ослободуваат во цитоплазмата, што предизвикува уништување на клеточните структури и нејзина смрт.
§ 2. Главни компоненти на еукариотска клетка
Еукариотските клетки (сл. 8 и 9) се организирани многу посложени од прокариотските. Тие се многу разновидни по големина (од неколку микрометри до неколку сантиметри), во форма и по структурни карактеристики (сл. 10).
Ориз. 8. Структура на еукариотска клетка. Генерализирана шема
Ориз. 9. Структура на клетките според електронска микроскопија
Ориз. 10. Разни еукариотски клетки: 1 – епителни; 2 – крв (д – еритроитис, / – леукитис); 3 – 'рскавица; 4 – коски; 5 – мазни мускули; 6 – сврзно ткиво; 7 – нервни клетки; 8 – напречно-пругастите мускулни влакна
Сепак, општата организација и присуството на основните компоненти се исти во сите еукариотски клетки (сл. 11).
Ориз. 11. Еукариотска клетка (дијаграм)
Плазмалема (надворешна клеточна мембрана).Основата на плазматската мембрана, како и другите мембрани во клетките (на пример, митохондриите, пластидите итн.), е слој од липиди кој има два реда молекули (сл. 12). Бидејќи молекулите на липидите се поларни (еден пол е хидрофилен, т.е. привлечен од вода, а другиот е хидрофобен, т.е. одбиен од вода), тие се распоредени по одреден редослед. Хидрофилните краеви на молекулите на еден слој се насочени кон водната средина - во цитоплазмата на клетката, а другиот слој - нанадвор од клетката - кон меѓуклеточната супстанција (кај повеќеклеточните организми) или водната средина (кај едноклеточните организми ).
Ориз. 12. Структура на клеточната мембрана според моделот на флуиден мозаик. Протеините и гликопротеините се потопуваат во двоен слој на липидни молекули, со нивните хидрофилни краеви (кругови) свртени нанадвор, а нивните хидрофобни краеви (брановидни линии) свртени длабоко во мембраната.
Протеинските молекули се мозаично вградени во бимолекуларен слој од липиди. На надворешната страна на животинската клетка, полисахаридните молекули се прикачени на липидите и протеинските молекули на плазмалемата, формирајќи гликолипиди и гликопротеини.
Овој агрегат формира слој гликокаликс.Поврзан со него функцијата на рецепторотплазма мембрани (види подолу); може да акумулира и разни материи што ги користи клетката. Покрај тоа, гликокаликсот ја подобрува механичката стабилност на плазмалмата.
Во клетките на растенијата и габите има и клеточен ѕид кој игра потпорна и заштитна улога. Кај растенијата се состои од целулоза, а кај габите од хитин.
Надворешната клеточна мембрана врши голем број функции, вклучувајќи:
♦ механички(поддршка, градење форма);
♦ бариера-транспорт(селективна пропустливост на различни материи: влегување во клетката на потребните материи и отстранување на непотребните и штетните);
♦ рецептор(одредување на различни хемикалии кои се наоѓаат во непосредна близина на клетката; перцепција на сигнали во форма на хормони; препознавање на „туѓи“ протеин од клетките на имунолошкиот систем итн.).
Размената на супстанции помеѓу клетката и околината се врши на различни начини - пасивни и активни.
Молекулите на водата и разните јони пасивно (поради дифузија, осмоза), без клетката да троши енергија, влегуваат низ посебни пори - тоа се пасивен транспорт.Макромолекулите како што се протеините, полисахаридите, дури и цели клетки, се снабдуваат од фагоцитозаИ пиноцитозасо потрошувачка на енергија - активен транспорт.
Со фагоцитоза, цели клетки или големи честички се проголтани (на пример, размислете за исхраната во амебите или фагоцитозата со заштитни крвни зрнца на бактериите). За време на пиноцитозата, се апсорбираат мали честички или капки од течна супстанција. Заедничко за двата процеса е тоа што апсорбираните супстанции се опкружени со надворешна мембрана која инвагинира за да формира вакуола, која потоа се движи длабоко во цитоплазмата на клетката.
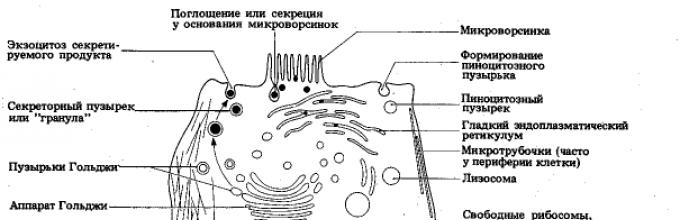
Егзоцитозата е процес (што е исто така активен транспорт) спротивен во насока на фагоцитозата и пиноцитозата (сл. 13). Со негова помош, несварената храна останува во протозои или може да се отстранат биолошки активните супстанции формирани во секреторната клетка.
Цитоплазма.Цитоплазмата е содржината на клетката, ограничена од плазмалемата, со исклучок на јадрото. Содржи земјата супстанција (хијалоплазма), органелиИ подмножества.
Хијалоплазма- вискозна течност која може да биде во состојба на било кое Зола(течност), или гел(како желе).
Доколку е потребно, цитоплазмата е способна за реверзибилна транзиција од една во друга состојба. На пример, за време на движењето на амебоидите (запомнете го делот „Протозои“ од курсот за зоологија), за време на формирањето на псевдопод, се случуваат брзи транзиции на цитоплазмата од гел во сол и обратно. Ова се должи на присуството во цитоплазмата на голем број на филаментозни протеински молекули актина.Кога ќе се поврзат едни со други за да формираат тродимензионална мрежа, цитоплазмата е во состојба на гел, а кога мрежата се распаѓа, таа е во солена состојба.
Хијалоплазмата содржи различни материи - ензими, протеини, јаглени хидрати, масти и други, органски и минерални. Овде се случуваат различни хемиски процеси - разградување на супстанции, нивна синтеза и модификации (промени).
Органоиди.Тоа се постојани компоненти на клетка со одредена структура и функции, сместени во нејзината цитоплазма. Во продолжение ќе зборуваме за органоиди за општа намена,својствени за сите типови на клетки на сите еукариоти. Тие се поврзани со обезбедување на виталните функции на второто. Органоиди за специјална наменасе наоѓа само во клетки од одреден (високо специјализиран) тип - на пример, миофибрили во мускулните клетки.
Органелите со општа намена имаат иста структура без разлика на кои клетки и на кои организми припаѓаат. Но, меѓу нив има и групи со мембрана (ендоплазматичен ретикулум, апарат Голџи, митохондрии, пластиди, лизозоми, вакуоли)како и немембрански ( рибозоми, клеточен центар)структура.
Ендоплазматичен ретикулум (ЕР).ЕПС се состои од мембрани и е сложен разгранет систем од тубули и цистерни што продира низ целата цитоплазма на клетката (сл. 14). Постојат два типа на EPS - грубоИ мазна.Рибозомите се прикачени на грубите мембрани (од цитоплазматската страна), додека не се присутни на мазните мембрани.
Ориз. 14. Ендоплазматичен ретикулум
Ендоплазматскиот ретикулум врши голем број важни функции во еукариотската клетка:
♦ разграничувајќи(поделба на внатрешниот волумен на клетката на различни реакциони простори);
♦ учество во синтезата на органски материи(рибозомите се наоѓаат на мембраните на грубиот ЕР, а ензимските комплекси се наоѓаат на мазните, обезбедувајќи синтеза на липиди, јаглени хидрати итн.);
♦ учество во формирањето на елементи на апаратот Голџи, лизозоми;
♦ транспорт на супстанции.
Голџи апарат.Апаратот Голџи (AG) е систем тенкови(рамни вакуоли) и меурчиња(везикули), лоцирани во непосредна близина на клеточното јадро, кои се формираат поради EPS како резултат на одвојувањето на неговите мали фрагменти (сл. 15). Кога овие фрагменти се спојуваат, се појавуваат нови цистерни на апаратот Голџи, додека од EPS се транспортираат различни супстанции, кои се вклучени во составувањето на сложените органски соединенија (протеини + јаглени хидрати, протеини + липиди, итн.), кои се отстрануваат со помош на АГ надвор од клетката. Овие биолошки активни супстанции или се отстранети од клетката (со користење на секреторни вакуоли со егзоцитоза) или се дел од лизозомите (види подолу) формирани од АГ.
Ориз. 15. Голџи апарат:
Апаратот Голџи ги извршува следниве функции:
♦ синтезабиолошки активни супстанции произведени од клетката;
♦ лачење (отстранување од клетката) на различни материи(хормони, ензими, супстанции од кои е изграден клеточниот ѕид и сл.);
♦ учество во формирањето на лизозоми.
Митохондриите.Сите видови на еукариотски клетки имаат митохондрии (сл. 16). Тие личат или на тркалезни тела или на прачки, поретко - на нишки. Нивните големини се движат од 1 до 7 микрони. Бројот на митохондриите во клетката се движи од неколку стотици до десетици илјади (во големите протозои).
Ориз. 16. Митохондрии. Погоре - митохондриите (а) во уринарните канали, видливи во светлосен микроскоп. Подолу е тродимензионален модел на митохондријална организација: 1 – cristae; 2 – надворешна мембрана; 3 – внатрешна мембрана; 4 – матрица
Митохондрионот е формиран од две мембрани - надворешенИ внатрешен,меѓу кои се наоѓа меѓумембрански простор.Внатрешната мембрана формира многу инвагинации - cristae, кои се или плочи или цевки. Оваа организација обезбедува огромна површина на внатрешната мембрана. Содржи ензими кои обезбедуваат конверзија на енергијата содржана во органските материи (јаглехидрати, липиди) во АТП енергија, неопходна за животот на клетката. Затоа, функцијата на митохондриите е да учествуваат во енергијаклеточни процеси. Затоа голем број митохондрии се својствени, на пример, во мускулните клетки кои вршат многу работа.
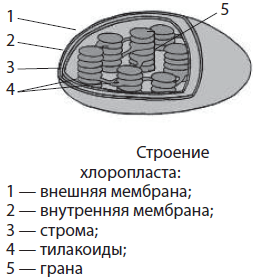
Пластиди.Во растителните клетки се наоѓаат специјални органели - пластиди, кои често имаат вретеновидна или заоблена форма, понекогаш посложена. Постојат три типа на пластиди - хлоропласти (сл. 17), хромопласти и леукопласти.
Хлоропластисе разликуваат по зелена боја, што се должи на пигментот - хлорофил,обезбедување на процесот фотосинтеза,т.е. синтеза на органски материи од вода (H 2 O) и јаглерод диоксид (CO 2) користејќи ја енергијата на сончевата светлина. Хлоропластите се наоѓаат главно во лисните клетки (кај вишите растенија). Тие се формирани од две мембрани лоцирани паралелно една на друга, околу содржината на хлоропластите - строма.Внатрешната мембрана формира бројни сплескани кеси - тилакоиди,кои се наредени (како куп монети) - зрна -и лежи во стромата. Токму тилакоидите содржат хлорофил.
Хромопластија одредува жолтата, портокаловата и црвената боја на многу цвеќиња и плодови, во чии ќелии се присутни во големи количини. Главните пигменти во нивниот состав се каротини.Функционалната цел на хромопластите е да привлекуваат животни со боја, обезбедувајќи опрашување на цвеќињата и растурање на семињата.
Ориз. 17. Пластиди: а – хлоропласти во клетките на листот Елодеа, видливи во светлосен микроскоп; б – дијаграм на внатрешната структура на хлоропластот со грана, кои се купишта рамни вреќички лоцирани нормално на површината на хлоропластот; в – подетален дијаграм кој прикажува анастомозирачки цевки кои ги поврзуваат одделните комори на вентилаторот
Леукопластисе безбојни пластиди содржани во клетките на подземните делови на растенијата (на пример, во клубени од компир), семиња и јадрото на стеблата. Во леукопластите, скробот главно се формира од гликоза и се акумулира во органите за складирање на растенијата.
Пластидите од еден тип може да се трансформираат во друг. На пример, кога лисјата ја менуваат бојата на есен, хлоропластите се претвораат во хромопласти.
Лизозоми.Овие органели изгледаат како везикули опкружени со мембрана со дијаметар до 2 микрони. Тие содржат неколку десетици ензими кои разградуваат протеини, нуклеински киселини, полисахариди и липиди. Функцијата на лизозомите е да учествуваат во процесите на интрацелуларно разградување на сложени органски соединенија (на пример, хранливи материи или супстанции од „потрошени“ клеточни компоненти). Лизозомите се спојуваат со фагоцитни (или пиноцитни) вакуоли за да формираат дигестивна вакуола.
Формирањето на лизозоми се јавува поради пукање од цистерните на апаратот Голџи.
Рибозоми.Рибозомите (сл. 18) се присутни во клетките и на еукариотите и на прокариотите, бидејќи тие вршат важна функција во биосинтеза на протеини(види поглавје 5). Секоја клетка содржи десетици, стотици илјади (до неколку милиони) од овие мали кружни органели.
Ориз. 18. Шема на структура на рибозом кој седи на мембраната на ендоплазматскиот ретикулум: 1 – мала подединица; 2 – tRNA; 3 – аминоацил-тРНК; 4 – амино киселина; 5 – голема подединица; 6 – мембрана на ендоплазматичен ретикулум; 7 – синтетизиран полипептиден синџир
Рибозомот се состои од две нееднакви подединици (делови). Тие се формираат одделно и се комбинираат, „опфаќајќи“ гласник РНК, за време на синтезата на протеинската молекула. Рибозомите содржат различни протеини и рибозомални РНК.
Клеточни подмножества.Така се нарекуваат непостојаните компоненти во клетката, присутни во главната супстанција на цитоплазмата во форма на зрна, гранули или капки. Вклучувањата може или не може да бидат опкружени со мембрана.
Функционално, постојат три типа на подмножества: резервни хранливи материи(скроб, гликоген, масти, протеини), секреторни подмножества(супстанции карактеристични за клетките на жлездата, произведени од нив - хормони на ендокрините жлезди итн.) и вклучување на посебна намена(во високо специјализирани клетки, на пример хемоглобин во црвените крвни зрнца).
§ 3. Организација на клеточното јадро. Хромозоми
Јадрото на клетката (види Сл. 8 и 9) е од најголема важност во животот на клетката, бидејќи служи како складиште на наследни информации содржани во хромозомите (види подолу).
Јадрото е ограничено со нуклеарна обвивка што ја одвојува неговата содржина (кариоплазма)од цитоплазмата. Школка се состои од две мембрани одделени со празнина. И двете се проникнати со бројни пори, благодарение на што е можна размена на материи помеѓу јадрото и цитоплазмата. Во клеточното јадро на повеќето еукариоти има од 1 до 7 јадра.Процесите на синтеза на РНК и tRNA се поврзани со нив.
Главни компоненти на кернелот - хромозоми,формирана од молекула на ДНК и разни протеини. Во светлосен микроскоп тие се јасно видливи само во периодот на клеточната делба (митоза, мејоза).Во клетка што не се дели, хромозомите изгледаат како долги тенки нишки распоредени низ целиот волумен на јадрото.
За време на клеточната делба, хромозомските нишки формираат густи спирали, како резултат на што тие стануваат видливи (со користење на конвенционален микроскоп) во форма на шипки, „шноли“. Целиот волумен на генетски информации се дистрибуира помеѓу хромозомите на јадрото. Во процесот на нивното проучување, беа идентификувани следниве модели:
♦ во јадрата на соматските клетки (т.е., телесни клетки, несексуални) сите индивидуи од ист вид содржат ист број на хромозоми што го сочинуваат збир на хромозоми(сл. 19);
Ориз. 19. Хромозоми на различни видови растенија и животни, прикажани на иста скала: 1,2 – амеба; 3,4 – дијатоми; 5–8, 18,19 – зелени алги; 9 – мушички агарик; 10 – липа; 11–12 – Дрософила; 13 – лосос; 14 – skerda (фамилија Asteraceae); 15 – растение од семејството на ароиди; 16 – пеперутка corydalis; 17 – инсект од семејството на скакулци; 20 – бубачка со воден шетач; 21 – цветна бубачка; 22 – амфибиска амбистома; 23 – алое (семејство крин)
♦ секој вид се карактеризира со свој сет на хромозоми според нивниот број (на пример, едно лице има 46 хромозоми, мувата Drosophila има 8, тркалезниот црв има 4, ракот има 196, коњот има 66 и пченката има 104);
♦ хромозомите во јадрата на соматските клетки може да се групираат во парови наречени хомологни хромозомиврз основа на нивната сличност (по структура и функција);
♦ во јадрата на герминативните клетки (гамети), секој пар на хомологни хромозоми содржи само еден, т.е. вкупниот сет на хромозоми е половина од соматските клетки;
♦ се нарекува единствен сет на хромозоми во герминативните клетки хаплоидени се означува со буквата n, а во соматски - диплоидна(2n).
Од горенаведеното, јасно е дека секој пар на хомологни хромозоми се формира со спојување на татковските и мајчините хромозоми за време на оплодувањето, т.е. фузија на полови клетки (гамети). Спротивно на тоа, за време на формирањето на герминативните клетки, од секој пар хомологни хромозоми само еден влегува во гаметата.
Хромозоми различнихомологните парови се разликуваат по големина и форма (сл. 20 и 21).
Ориз. 20. Структура и типови на хромозоми: а – изглед 1 – центромер; 2 – кратко рамо; 3 – долго рамо); внатрешна структура на истиот хромозом (1 – центромер; – молекули на ДНК); в – типови на хромозоми (1 – еднокраки; различно вооружени; 3 – еднакви краци: X – крак, V – центромер)
Ориз. 21. Хромозомот се состои од ДНК и протеини. Молекулата на ДНК се реплицира. Два идентични ДНК двојни спирали остануваат поврзани во регионот на центромерот. Овие копии стануваат посебни хромозоми подоцна за време на клеточната делба
Во телото на хромозомите постојат примарната констрикција (наречена центромер),на кој се прицврстени конците вретена на фисија.Го дели хромозомот на два рамоХромозомите можат да бидат со еднакво оружје, нееднакво вооружени или еднокраки.
Поглавје 5. Метаболизам
§ 1. Метаболизмот како единство на асимилација и дисимилација
Сите клетки и живи организми се отворени системи, односно се во состојба на постојана размена на енергии и материи со околината. Постојат отворени системи во неживата природа, но нивното постоење е квалитативно различно од живите организми. Размислете за овој пример: запаленото парче од мајчин сулфур е во состојба на размена со околината. Кога гори, O 2 се апсорбира, а SO 2 и енергија (во форма на топлина) се ослободуваат. Меѓутоа, во овој случај, парче сулфур како физичко тело се уништува и ја губи својата примарна структура.
За живите организми размената со околината се покажува како услов за зачувување и одржување на нивната структурна организација преку самообновување на сите супстанции и компоненти од кои се состојат.
Метаболизам (метаболизам) е збир на процеси кои се случуваат во живите организми (потрошувачка, трансформација, акумулација и ослободување на супстанции и енергија) кои ја обезбедуваат нивната витална активност, развој, раст, репродукција. Во текот на метаболичкиот процес, молекулите кои ги сочинуваат клетките се разградуваат и синтетизираат; обновување на клеточните структури и меѓуклеточната супстанција.
Метаболизмот се заснова на меѓусебно поврзани процеси асимилација(анаболизам) и дисимилација(катаболизам). За време на асимилација (пластична размена), сложените супстанции се синтетизираат од едноставни. Благодарение на тоа се создаваат сите органски материи во клетката неопходни за изградба на нејзините структурни компоненти, ензимски системи итн. Асимилацијата секогаш се одвива со трошење на енергија.
За време на дисимилацијата (енергетскиот метаболизам), сложените органски материи се разложуваат на поедноставни или неоргански. Во овој случај се ослободува енергија, која клетката ја троши за извршување на различни процеси кои ја обезбедуваат нејзината витална активност (синтеза и транспорт на супстанции, механичка работа и сл.).
Сите живи организми можат да се поделат во две групи: автотрофиИ хетеротрофи,кои се разликуваат по изворите на енергија и потребните материи за обезбедување на нивните витални функции.
Автотрофи- организми кои синтетизираат органски соединенија од неоргански материи користејќи ја енергијата на сончевата светлина (како фототрофи– растенија, цијанобактерии) или енергија добиена од оксидација на минерални (неоргански) супстанции (како на пр. хемотрофи– сулфурни бактерии, железни бактерии итн.). Следствено, тие се способни самостојно да ги создадат супстанциите потребни за нивната животна активност.
§ 2. Дисимилација кај анаеробните и аеробните организми
Организмите може да се поделат во две групи и според природата на дисимилацијата - аеробиИ анаероби.Аероби (од грчки. демон– воздух) бараат бесплатен кислород за живот. Кај анаеробите (грчки. ак– негативна честичка) не е потребно. Во нив, дисимилацијата се врши со ферментација - без кислород, ензимско разградување на органската материја со формирање на поедноставни органски материи и ослободување на енергија. На пример:
♦ ферментација на млечна киселина:
C 6 H 12 O 6 + 2H 3 PO 4 + 2ADP → 2PH + 2ATP + 2H 2 O;
♦ алкохолна ферментација:
C 6 H 12 O 6 + 2PH + 2ADP → 2C 2 H 5 OH + 2CO 2 + 2ATP + 2H 2 O.
Супстанциите што се формираат за време на ферментацијата се органски и затоа сè уште содржат многу енергија.
Ориз. 22. Врски помеѓу асимилација и дисимилација кај автотрофни и хетеротрофни организми
Кај аеробните организми, за време на процесот на дишење во митохондриите, се случува целосно распаѓање на органските материи (со користење на O 2) до енергетски сиромашните крајни производи CO 2 и H 2 O и се ослободува значително поголема количина на енергија:
C 6 H 12 0 6 (гликоза) + 0 2 > 6C0 2 + 6H 2 0 + енергија (поради што се синтетизираат 38 ATP молекули).
Да го разгледаме во форма на генерализирани шеми метаболизмот на автотрофните и хетеротрофните аеробни организми (сл. 22).
Асимилација.Нејзините најважни процеси се фотосинтезаИ биосинтеза на протеини.
Ве покануваме да се запознаете со материјалите и.
: целулозна мембрана, мембрана, цитоплазма со органели, јадро, вакуоли со клеточен сок.Присуството на пластиди е главната карактеристика на растителната клетка.
Функции на клеточната мембрана- го одредува обликот на клетката, штити од фактори на животната средина.
Плазма мембрана- тенок филм, кој се состои од интерактивни молекули на липиди и протеини, ја ограничува внатрешната содржина од надворешната средина, обезбедува транспорт на вода, минерали и органски материи во клетката со осмоза и активен транспорт, а исто така ги отстранува отпадните производи.
Цитоплазма- внатрешната полутечна средина на клетката, во која се наоѓаат јадрото и органелите, обезбедува врски меѓу нив и учествува во основните животни процеси.
Ендоплазматичен ретикулум- мрежа на разгранети канали во цитоплазмата. Тој е вклучен во синтезата на протеини, липиди и јаглени хидрати и во транспортот на супстанции. Рибозомите се тела лоцирани на ER или во цитоплазмата, кои се состојат од РНК и протеини и се вклучени во синтезата на протеините. EPS и рибозомите се единствен апарат за синтеза и транспорт на протеини.
Митохондриите- органели ограничени од цитоплазмата со две мембрани. Во нив се оксидираат органски материи и се синтетизираат молекулите на АТП со учество на ензими. Зголемување на површината на внатрешната мембрана на која се наоѓаат ензимите поради криста. АТП е органска супстанција богата со енергија.
Пластиди(хлоропласти, леукопласти, хромопласти), нивната содржина во клетката е главната карактеристика на растителниот организам. Хлоропластите се пластиди кои го содржат зелениот пигмент хлорофил, кој ја апсорбира светлосната енергија и ја користи за синтеза на органски материи од јаглерод диоксид и вода. Хлоропластите се одделени од цитоплазмата со две мембрани, бројни израстоци - грана на внатрешната мембрана, во која се наоѓаат молекули и ензими на хлорофилот.
Комплекс Голџи- систем на шуплини ограничени од цитоплазмата со мембрана. Акумулација на протеини, масти и јаглехидрати во нив. Спроведување на синтеза на масти и јаглени хидрати на мембраните.
Лизозоми- тела ограничени од цитоплазмата со една мембрана. Ензимите што ги содржат го забрзуваат разградувањето на сложените молекули на едноставни: протеини во аминокиселини, сложени јаглехидрати во едноставни, липиди во глицерол и масни киселини, а исто така ги уништуваат мртвите делови од клетката и цели клетки.
Вакуоли- шуплини во цитоплазмата исполнети со клеточен сок, место на акумулација на резервни хранливи материи и штетни материи; ја регулираат содржината на вода во ќелијата.
Јадро- главниот дел од ќелијата, покриен однадвор со нуклеарна обвивка со две мембрани, прободена со пори. Супстанциите влегуваат во јадрото и се отстрануваат од него преку порите. Хромозомите се носители на наследни информации за карактеристиките на организмот, главните структури на јадрото, од кои секоја се состои од една молекула на ДНК комбинирана со протеини. Јадрото е место на синтеза на ДНК, mRNA и rRNA.

Присуство на надворешна мембрана, цитоплазма со органели и јадро со хромозоми.
Надворешна или плазма мембрана- ја разграничува содржината на клетката од околината (други клетки, меѓуклеточна супстанција), се состои од липидни и протеински молекули, обезбедува комуникација помеѓу клетките, транспорт на супстанции во клетката (пиноцитоза, фагоцитоза) и надвор од клетката.
Цитоплазма- внатрешната полутечна средина на клетката, која обезбедува комуникација помеѓу јадрото и органелите лоцирани во неа. Главните животни процеси се одвиваат во цитоплазмата.
Клеточни органели:
1) ендоплазматичен ретикулум (ER)- систем на разгранети тубули, учествува во синтезата на протеини, липиди и јаглени хидрати, во транспортот на супстанции во клетката;
2) рибозоми- телата кои содржат rRNA се наоѓаат на ER и во цитоплазмата и учествуваат во синтезата на протеините. EPS и рибозомите се единствен апарат за синтеза и транспорт на протеини;
3) митохондриите- „електрични станици“ на клетката, ограничени од цитоплазмата со две мембрани. Внатрешниот формира криста (набори), зголемувајќи ја неговата површина. Ензимите на кристаите ја забрзуваат оксидацијата на органските материи и синтезата на молекулите на АТП богати со енергија;
4) Комплекс Голџи- група на шуплини ограничени со мембрана од цитоплазмата, исполнета со протеини, масти и јаглехидрати, кои или се користат во виталните процеси или се отстрануваат од клетката. Мембраните на комплексот вршат синтеза на масти и јаглени хидрати;
5) лизозоми- телата исполнети со ензими го забрзуваат разградувањето на протеините во амино киселини, липидите во глицерол и масни киселини, полисахаридите во моносахариди. Во лизозомите, мртвите делови од клетката, цели клетки, се уништуваат.
Клеточни подмножества- акумулации на резервни хранливи материи: протеини, масти и јаглени хидрати.
Јадро- најважниот дел од клетката. Тој е покриен со двомембранска обвивка со пори, преку која некои супстанции продираат во јадрото, а други влегуваат во цитоплазмата. Хромозомите се главните структури на јадрото, носители на наследни информации за карактеристиките на организмот. Се пренесува при поделбата на матичната клетка на ќерките, а со герминативните клетки на ќерките организми. Јадрото е место на синтеза на ДНК, mRNA и rRNA.
Вежба:
Објасни зошто органелите се нарекуваат специјализирани клеточни структури?
Одговор:органелите се нарекуваат специјализирани клеточни структури, бидејќи тие вршат строго дефинирани функции, наследни информации се складираат во јадрото, АТП се синтетизира во митохондриите, фотосинтезата се јавува во хлоропластите итн.
Ако имате прашања во врска со цитологијата, можете да контактирате
Теорија за задача 5 од Единствен државен испит по биологија
Клеточна структура. Односот помеѓу структурата и функциите на деловите и органелите на клетката е основата на нејзиниот интегритет
Клеточна структура
Структура на прокариотски и еукариотски клетки
Главните структурни компоненти на клетките се плазма мембраната, цитоплазмата и наследниот апарат. Во зависност од карактеристиките на организацијата, се разликуваат два главни типа на клетки: прокариотски и еукариотски. Главната разлика помеѓу прокариотските клетки и еукариотските клетки е организацијата на нивниот наследен апарат: кај прокариотите се наоѓа директно во цитоплазмата (оваа област на цитоплазмата се нарекува нуклеоид) и не е одвоена од него со мембрански структури, додека кај еукариотите поголемиот дел од ДНК е концентрирана во јадрото, опкружена со двојна мембрана. Покрај тоа, генетските информации на прокариотските клетки, лоцирани во нуклеоидот, се запишуваат во кружна молекула на ДНК, додека кај еукариотите молекулите на ДНК се отворени.
За разлика од еукариотите, цитоплазмата на прокариотските клетки содржи и мал број органели, додека еукариотските клетки се карактеризираат со значителна разновидност на овие структури.
Структура и функции на биолошките мембрани
Структурата на биомембраната.Мембраните што ги врзуваат клетките и мембранските органели на еукариотските клетки имаат заеднички хемиски состав и структура. Тие вклучуваат липиди, протеини и јаглени хидрати. Мембранските липиди главно се претставени со фосфолипиди и холестерол. Повеќето мембрански протеини се сложени протеини, како што се гликопротеините. Јаглехидратите не се појавуваат независно во мембраната, тие се поврзани со протеини и липиди. Дебелината на мембраните е 7-10 nm.
Според моментално општо прифатениот течен мозаичен модел на структурата на мембраната, липидите формираат двоен слој, или липиден двослој, во која хидрофилните „глави“ на липидните молекули се свртени кон надвор, а хидрофобните „опашки“ се скриени во мембраната. Овие „опашки“, поради нивната хидрофобност, обезбедуваат одвојување на водените фази на внатрешната средина на клетката и нејзината околина. Протеините се поврзани со липидите преку различни видови на интеракции. Некои протеини се наоѓаат на површината на мембраната. Таквите протеини се нарекуваат периферни, или површни. Другите протеини се делумно или целосно потопени во мембраната - тоа се интегрален,или потопени протеини. Мембранските протеини вршат структурни, транспортни, каталитички, рецепторни и други функции.
Мембраните не се како кристали; нивните компоненти се постојано во движење, како резултат на што се појавуваат празнини помеѓу липидните молекули - пори преку кои различни супстанции можат да влезат или да излезат од клетката.
Биолошките мембрани се разликуваат по нивната локација во клетката, хемискиот состав и функциите. Главните типови на мембрани се плазма и внатрешни. Плазма мембранасодржи околу 45% липиди (вклучувајќи гликолипиди), 50% протеини и 5% јаглени хидрати. Над површината на мембраната излегуваат синџири на јаглени хидрати, кои се дел од сложени протеини-гликопротеини и сложени липиди-гликолипиди. Гликопротеините на плазмалемата се исклучително специфични. На пример, тие се користат за меѓусебно препознавање на клетките, вклучително и спермата и јајце клетката.
На површината на животинските клетки, синџирите на јаглени хидрати формираат тенок површински слој - гликокаликс.Тој е откриен во скоро сите животински клетки, но неговиот степен на експресивност варира (10-50 μm). Гликокаликсот обезбедува директна комуникација помеѓу клетката и надворешната средина, каде што се случува екстрацелуларното варење; Рецепторите се наоѓаат во гликокаликсот. Покрај плазмалемата, клетките на бактериите, растенијата и габите се опкружени и со клеточни мембрани.
Внатрешни мембраниеукариотските клетки ограничуваат различни делови од клетката, формирајќи чудни „оддели“ - прегради, кој промовира одвојување на различни метаболички и енергетски процеси. Тие може да се разликуваат во хемискиот состав и функции, но нивниот генерален структурен план останува ист.
Функции на мембраната:
- Ограничување.Идејата е дека тие го одвојуваат внатрешниот простор на клетката од надворешната средина. Мембраната е полупропустлива, односно низ неа слободно можат да поминат само оние материи кои и се потребни на клетката, а постојат механизми за транспортирање на потребните материи.
- Рецептор.Тоа е првенствено поврзано со перцепцијата на еколошките сигнали и преносот на овие информации во ќелијата. Специјални рецепторни протеини се одговорни за оваа функција. Мембранските протеини се одговорни и за клеточно препознавање според принципот „пријател или непријател“, како и за формирање на меѓуклеточни врски, од кои најпроучени се синапсите на нервните клетки.
- Каталитички.На мембраните се наоѓаат бројни ензимски комплекси, како резултат на што на нив се случуваат интензивни синтетички процеси.
- Трансформирање на енергија.Поврзано со формирање на енергија, нејзино складирање во форма на АТП и потрошувачка.
- Одделување.Мембраните, исто така, го ограничуваат просторот внатре во клетката, со што се одвојуваат почетните материјали на реакцијата и ензимите кои можат да ги спроведат соодветните реакции.
- Формирање на меѓуклеточни контакти.И покрај фактот дека дебелината на мембраната е толку мала што не може да се разликува со голо око, таа, од една страна, служи како прилично сигурна бариера за јоните и молекулите, особено оние растворливи во вода, а од друга страна , обезбедува нивен транспорт во и надвор од ќелијата.
- Транспорт.

Мембрански транспорт.Поради фактот што клетките, како елементарни биолошки системи, се отворени системи, за да се обезбеди метаболизам и енергија, одржување на хомеостазата, растот, раздразливоста и други процеси, потребен е пренос на супстанции преку мембраната - мембрански транспорт. Во моментов, транспортот на супстанции преку клеточната мембрана е поделен на активна, пасивна, ендо- и егзоцитоза.
Пасивен транспорт- Станува збор за вид транспорт кој се случува без потрошувачка на енергија од повисоки кон помали концентрации. Мали неполарни молекули растворливи во липиди (O 2, CO 2) лесно продираат во клетката преку едноставна дифузија. Оние кои се нерастворливи во липиди, вклучувајќи ги и наелектризираните мали честички, се собираат од протеините-носители или минуваат низ посебни канали (гликоза, амино киселини, K +, PO 4 3-). Овој вид на пасивен транспорт се нарекува олеснета дифузија. Водата влегува во клетката преку порите во липидната фаза, како и преку специјални канали обложени со протеини. Транспортот на вода низ мембрана се нарекува со осмоза.

Осмозата е исклучително важна во животот на клетката, бидејќи ако се стави во раствор со поголема концентрација на соли отколку во клеточниот раствор, тогаш водата ќе почне да ја напушта клетката и волуменот на живата содржина ќе почне да се намалува. Во животинските клетки, клетката како целина се намалува, а во растителните клетки цитоплазмата заостанува зад клеточниот ѕид, што се нарекува. плазмолиза. Кога клетката се става во раствор помалку концентриран од цитоплазмата, транспортот на вода се случува во спротивна насока - во клетката. Сепак, постојат ограничувања за растегливоста на цитоплазматската мембрана, а животинската клетка на крајот пука, додека растителната клетка не дозволува тоа да се случи поради силниот клеточен ѕид. Феноменот на пополнување на целиот внатрешен простор на ќелијата со клеточна содржина се нарекува деплазмолиза. Треба да се земе предвид интрацелуларната концентрација на соли при подготовка на лекови, особено за интравенска администрација, бидејќи тоа може да доведе до оштетување на крвните клетки (за ова се користи солен раствор со концентрација од 0,9% натриум хлорид). Ова не е помалку важно кога се одгледуваат клетки и ткива, како и животински и растителни органи.

Активен транспортпродолжува со трошење на енергијата на АТП од помала концентрација на супстанција до повисока. Се изведува со помош на специјални протеини на пумпата. Протеините пумпаат K + , Na + , Ca 2 + и други јони низ мембраната, што го промовира транспортот на есенцијалните органски материи, како и појавата на нервни импулси итн.
Ендоцитоза- ова е активен процес на апсорпција на супстанции од клетката, во кој мембраната формира инвагинации, а потоа формира мембрански везикули - фагозоми, кои ги содржат апсорбираните предмети. Тогаш примарниот лизозом се спојува со фагозомот и се формира секундарен лизозом, или фаголизозом, или дигестивна вакуола. Содржината на везикулата се вари со ензими на лизозом, а производите на распаѓање се апсорбираат и асимилираат од клетката. Несварените остатоци се отстрануваат од клетката со егзоцитоза. Постојат два главни типа на ендоцитоза: фагоцитоза и пиноцитоза.
Фагоцитозае процес на фаќање од страна на површината на клетката и апсорпција на цврсти честички од страна на клетката, и пиноцитоза- течности. Фагоцитозата се јавува главно во животинските клетки (едноклеточни животни, човечки леукоцити), ја обезбедува нивната исхрана и често го штити телото. Со пиноцитоза, протеините, комплексите антиген-антитела се апсорбираат за време на имунолошките реакции итн. Меѓутоа, многу вируси влегуваат во клетката и со пиноцитоза или фагоцитоза. Во растителните и габичните клетки, фагоцитозата е практично невозможна, бидејќи тие се опкружени со издржливи клеточни мембрани.

Егзоцитоза- процес обратен на ендоцитоза. На тој начин, од дигестивните вакуоли се ослободуваат несварените остатоци од храна, а се отстрануваат и супстанциите неопходни за животот на клетката и телото во целина. На пример, преносот на нервните импулси се случува поради ослободување на хемиски гласници од невронот што го испраќа импулсот - медијатори, а во растителните клетки вака се излачуваат помошните јаглехидрати на клеточната мембрана.
Клеточни ѕидови на растителни клетки, габи и бактерии.Надвор од мембраната, клетката може да лачи силна рамка - клеточната мембрана,или клеточен ѕид.
Кај растенијата, основата на клеточниот ѕид е целулоза, спакуван во снопови од 50-100 молекули. Просторите меѓу нив се полни со вода и други јаглехидрати. Ѕидот на растителните клетки е проникнат со тубули - плазмодезмата, низ кој минуваат мембраните на ендоплазматскиот ретикулум. Плазмодесмата врши транспорт на супстанции помеѓу клетките. Меѓутоа, транспортот на супстанции, како што е водата, може да се случи и по самите клеточни ѕидови. Со текот на времето, во клеточниот ѕид на растенијата се акумулираат различни супстанции, вклучувајќи танини или супстанции слични на маснотии, што доведува до лигнификација или суберизација на самиот клеточен ѕид, поместување на водата и смрт на клеточната содржина. Помеѓу клеточните ѕидови на соседните растителни ќелии има разделувачи слични на желе - средни плочи кои ги држат заедно и го цементираат телото на растението како целина. Тие се уништуваат само за време на процесот на зреење на овошјето и кога лисјата паѓаат.

Се формираат клеточни ѕидови на габични клетки хитин- јаглехидрат кој содржи азот. Тие се прилично силни и се надворешниот скелет на клетката, но сепак, како и кај растенијата, ја спречуваат фагоцитозата.
Кај бактериите, клеточниот ѕид содржи јаглехидрати со пептидни фрагменти - муреин, сепак, неговата содржина значително варира кај различни групи бактерии. Други полисахариди, исто така, може да се излачат на врвот на клеточниот ѕид, формирајќи мукозна капсула која ги штити бактериите од надворешни влијанија.
Мембраната го одредува обликот на клетката, служи како механичка потпора, врши заштитна функција, ги обезбедува осмотските својства на клетката, го ограничува истегнувањето на живата содржина и го спречува пукањето на клетката, кое се зголемува поради навлегувањето на вода. . Покрај тоа, водата и супстанциите растворени во неа го надминуваат клеточниот ѕид пред да влезат во цитоплазмата или, обратно, кога ја напуштаат, додека водата се транспортира низ клеточните ѕидови побрзо отколку низ цитоплазмата.
Цитоплазма
Цитоплазма- Ова е внатрешната содржина на ќелијата. Во него се потопени сите клеточни органели, јадрото и разни отпадни продукти.
Цитоплазмата ги поврзува сите делови на клетката еден со друг, а во неа се случуваат бројни метаболички реакции. Цитоплазмата е одвоена од околината и поделена на одделенија со мембрани, односно клетките имаат мембранска структура. Може да биде во две состојби - сол и гел. Сол- ова е полутечна, желеста состојба на цитоплазмата, во која најинтензивно се одвиваат виталните процеси, и гел- погуста, желатинозна состојба која го попречува појавувањето на хемиски реакции и транспортот на материи.
Течниот дел од цитоплазмата без органели се нарекува хијалоплазма. Хијалоплазмата или цитозолот е колоиден раствор во кој има еден вид суспензија на прилично големи честички, на пример протеини, опкружени со диполи на молекули на вода. Врнежите на оваа суспензија не се случуваат поради фактот што тие имаат ист полнеж и се одбиваат едни со други.
Органоиди
Органоиди- Тоа се постојани компоненти на клетката кои вршат специфични функции.
Во зависност од нивните структурни карактеристики, тие се делат на мембрански и немембрански. Мембранаорганелите, пак, се класифицирани како едномембрански (ендоплазматичен ретикулум, комплекс Голџи и лизозоми) или двомембрани (митохондрии, пластиди и јадро). Не-мембранаОрганелите се рибозоми, микротубули, микрофиламенти и клеточниот центар. Од наведените органели, само рибозомите се својствени за прокариотите.
Структура и функции на јадрото. Јадро- голема двомембранска органела која лежи во центарот на клетката или на нејзината периферија. Димензиите на јадрото може да се движат од 3-35 микрони. Обликот на јадрото е најчесто сферичен или елипсоиден, но има и јадра во форма на прачка, фузиформни, гравовидни, лобуси, па дури и сегментирани. Некои истражувачи веруваат дека обликот на јадрото одговара на обликот на самата клетка.
Повеќето клетки имаат едно јадро, но, на пример, во клетките на црниот дроб и срцето може да има две од нив, а во голем број неврони - до 15. Скелетните мускулни влакна обично содржат многу јадра, но тие не се клетки во целосна смисла на зборот, бидејќи тие се формираат како резултат на спојување на неколку клетки.
Јадрото е опкружено нуклеарна обвивка, а неговиот внатрешен простор е исполнет нуклеарен сок, или нуклеоплазма (кариоплазма), во кој се потопуваат хроматинИ јадро. Јадрото врши такви важни функции како складирање и пренесување на наследни информации, како и контролирање на животот на клетката.

Улогата на јадрото во преносот на наследни информации беше убедливо докажана во експериментите со зелената алга Acetabularia. Во една џиновска клетка, достигнувајќи должина од 5 см, се разликуваат капа, стебленце и ризоид. Покрај тоа, содржи само едно јадро лоцирано во ризоидот. Во 1930-тите, И. Хемерлинг го пресадил јадрото на еден вид ацетабуларија со зелена боја во ризоидот на друг вид, со кафена боја, од кој јадрото било отстрането. По некое време, на растението со пресаденото јадро му пораснало ново капаче, како алгите донори на јадрото. Во исто време, капачето или стебленцето, одвоено од ризоидот и кое не содржи јадро, умрело по некое време.

Нуклеарна обвивкаформирана од две мембрани - надворешна и внатрешна, меѓу кои има простор. Меѓумембранскиот простор комуницира со шуплината на грубиот ендоплазматичен ретикулум, а надворешната мембрана на јадрото може да носи рибозоми. Нуклеарната обвивка е проникната со бројни пори обложени со специјални протеини. Транспортот на супстанции се случува низ порите: потребните протеини (вклучувајќи ензими), јони, нуклеотиди и други супстанции влегуваат во јадрото, а молекулите на РНК, потрошените протеини и подединици на рибозоми го напуштаат. Така, функциите на нуклеарната обвивка се одвојување на содржината на јадрото од цитоплазмата, како и регулирање на метаболизмот помеѓу јадрото и цитоплазмата.
Нуклеоплазманаречена содржина на јадрото, во кое се потопени хроматинот и јадрото. Тоа е колоиден раствор, хемиски потсетува на цитоплазмата. Ензимите на нуклеоплазмата ја катализираат размената на амино киселини, нуклеотиди, протеини итн. Нуклеоплазмата е поврзана со хијалоплазмата преку нуклеарните пори. Функциите на нуклеоплазмата, како и хијалоплазмата, се да обезбеди меѓусебно поврзување на сите структурни компоненти на јадрото и да спроведе голем број ензимски реакции.
Хроматиннаречена збирка од тенки нишки и гранули потопени во нуклеоплазмата. Може да се открие само со боење, бидејќи индексите на рефракција на хроматин и нуклеоплазма се приближно исти. Филаментозната компонента на хроматин се нарекува еухроматини зрнести - хетерохроматин. Еухроматинот е слабо набиен, бидејќи од него се читаат наследни информации, додека повеќе спирализираниот хетерохроматин е генетски неактивен.
Хроматинот е структурна модификација на хромозомите во јадрото што не се дели. Така, хромозомите се постојано присутни во јадрото, само нивната состојба се менува во зависност од функцијата што јадрото ја извршува во моментот.
Составот на хроматинот главно вклучува нуклеопротеински протеини (деоксирибонуклеопротеини и рибонуклеопротеини), како и ензими, од кои најважните се поврзани со синтезата на нуклеинските киселини и некои други супстанции.
Функциите на хроматинот се состојат, прво, во синтеза на нуклеински киселини специфични за даден организам, кои ја насочуваат синтезата на специфични протеини, и второ, во преносот на наследните својства од матичната клетка до клетките ќерки, за која цел хроматинските нишки се пакуваат во хромозоми за време на процесот на делење.
Нуклеолус- сферично тело, јасно видливо под микроскоп, со дијаметар од 1-3 микрони. Се формира на делови од хроматин во кои се кодирани информации за структурата на rRNA и рибозомалните протеини. Често има само едно јадро во јадрото, но во оние клетки каде што се случуваат интензивни витални процеси, може да има две или повеќе јадра. Функциите на јадрата се синтеза на rRNA и склопување на рибозомални подединици со комбинирање на rRNA со протеини кои доаѓаат од цитоплазмата.
Митохондриите- двомембрански органели со тркалезна, овална или прачка форма, иако се среќаваат и спирални (кај сперматозоидите). Дијаметарот на митохондриите е до 1 µm, а должината е до 7 µm. Просторот во митохондриите е исполнет со матрица. Матрица- Ова е главната супстанца на митохондриите. Во него се потопени кружна молекула на ДНК и рибозоми. Надворешната мембрана на митохондриите е мазна и непропустлива за многу супстанции. Внатрешната мембрана има проекции - cristas, зголемување на површината на мембраните за да се појават хемиски реакции. На површината на мембраната има бројни протеински комплекси кои го сочинуваат таканаречениот респираторен синџир, како и ензими на АТП синтетаза во облик на печурка. Аеробната фаза на дишење се јавува во митохондриите, при што се синтетизира АТП.

Пластиди- големи двомембрански органели, карактеристични само за растителните клетки. Внатрешниот простор на пластидите е исполнет строма, или матрица. Стромата содржи повеќе или помалку развиен систем на мембрански везикули - тилакоиди, кои се собираат во купови - зрна, како и сопствената кружна молекула на ДНК и рибозоми. Постојат четири главни типа на пластиди: хлоропласти, хромопласти, леукопласти и пропластиди.
Хлоропласти- ова се зелени пластиди со дијаметар од 3-10 микрони, јасно видливи под микроскоп. Ги има само во зелените делови на растенијата - лисјата, младите стебла, цветовите и плодовите. Хлоропластите се генерално овални или елипсоидни по форма, но исто така можат да бидат во облик на чаша, во форма на спирала, па дури и со лобуси. Бројот на хлоропласти во една клетка е во просек од 10 до 100 парчиња. Меѓутоа, на пример, кај некои алги може да биде една, да има значителни димензии и сложена форма - тогаш се нарекува хроматофор. Во други случаи, бројот на хлоропласти може да достигне неколку стотици, додека нивните големини се мали. Бојата на хлоропластите се должи на главниот пигмент на фотосинтезата - хлорофил, иако содржат и дополнителни пигменти - каротеноиди. Каротеноидите стануваат забележливи само на есен, кога хлорофилот во стареењето на листовите се распаѓа. Главната функција на хлоропластите е фотосинтезата. Светлосните реакции на фотосинтезата се случуваат на тилакоидните мембрани, на кои се прикачени молекулите на хлорофилот, а темните реакции се случуваат во стромата, каде што се содржани бројни ензими.
Хромопласти- Станува збор за жолти, портокалови и црвени пластиди кои содржат каротеноидни пигменти. Обликот на хромопластите исто така може значително да варира: тие можат да бидат цевчести, топчести, кристални итн. Хромопластите им даваат боја на цветовите и плодовите на растенијата, привлекувајќи опрашувачи и дистрибутери на семиња и плодови.
Леукопласти- Станува збор за бели или безбојни пластиди, претежно кружни или овални во форма. Вообичаени се во нефотосинтетичките делови на растенијата, на пример во кората на лисјата, клубени од компири итн. Тие складираат хранливи материи, најчесто скроб, но кај некои растенија тоа може да бидат протеини или масло.
Пластидите се формираат во растителните клетки од пропластиди, кои се веќе присутни во клетките на образовното ткиво и се мали двомембрански тела. Во раните фази на развој, различни видови пластиди се способни да се трансформираат едни во други: кога се изложени на светлина, леукопластите на клубенот од компир и хромопластите на коренот на морковот стануваат зелени.
Пластидите и митохондриите се нарекуваат полуавтономни органели на клетката, бидејќи тие имаат свои ДНК молекули и рибозоми, вршат синтеза на протеини и се делат независно од клеточната делба. Овие карактеристики се објаснуваат со нивното потекло од едноклеточни прокариотски организми. Сепак, „независноста“ на митохондриите и пластидите е ограничена, бидејќи нивната ДНК содржи премалку гени за слободно постоење, додека остатокот од информациите се кодирани во хромозомите на јадрото, што му овозможува да ги контролира овие органели.
Ендоплазматичен ретикулум (ЕР), или ендоплазматичен ретикулум (ER), е едномембранска органела, која е мрежа од мембрански шуплини и тубули кои зафаќаат до 30% од содржината на цитоплазмата. Дијаметарот на EPS тубулите е околу 25-30 nm. Постојат два вида EPS - груб и мазен. Груб XPSноси рибозоми, каде што се јавува синтеза на протеини. Нежните XPSнема рибозоми. Неговата функција е синтеза на липиди и јаглени хидрати, како и транспорт, складирање и неутрализација на токсични материи. Особено е развиен во оние клетки каде што се случуваат интензивни метаболички процеси, на пример во клетките на црниот дроб - хепатоцитите - и скелетните мускулни влакна. Супстанциите синтетизирани во ЕР се транспортираат до апаратот Голџи. Склопувањето на клеточните мембрани се случува и во ЕР, но нивното формирање е завршено во апаратот Голџи.
Голџи апарат,или Комплекс Голџи, е едномембранска органела формирана од систем на рамни цистерни, тубули и везикули одвоени од нив. Структурната единица на апаратот Голџи е диктозом- куп тенкови, на едниот пол од кои доаѓаат супстанции од ЕПС, а од спротивниот пол, откако претрпеа одредени трансформации, тие се пакуваат во везикули и се испраќаат во други делови од клетката. Дијаметарот на резервоарите е околу 2 микрони, а дијаметарот на малите меурчиња е околу 20-30 микрони. Главните функции на комплексот Голџи се синтеза на одредени супстанции и модификација (промена) на протеини, липиди и јаглени хидрати кои доаѓаат од ER, конечно формирање на мембрани, како и транспорт на супстанции низ клетката, обновување на нејзините структури. и формирање на лизозоми. Апаратот Голџи го доби своето име во чест на италијанскиот научник Камило Голџи, кој прв ја откри оваа органела (1898).

Лизозоми- мали едномембрански органели со дијаметар до 1 μm, кои содржат хидролитички ензими вклучени во интрацелуларното варење. Мембраните на лизозомите се слабо пропустливи за овие ензими, така што лизозомите ги извршуваат своите функции многу прецизно и насочено. Така, тие земаат активно учество во процесот на фагоцитоза, формирајќи дигестивни вакуоли, а во случај на гладување или оштетување на одредени делови од клетката, ги варат без да влијаат на другите. Неодамна е откриена улогата на лизозомите во процесите на клеточна смрт.
Вакуолае празнина во цитоплазмата на растителните и животинските клетки, ограничена со мембрана и исполнета со течност. Дигестивните и контрактилните вакуоли се наоѓаат во клетките на протозоите. Првите учествуваат во процесот на фагоцитоза, бидејќи ги разградуваат хранливите материи. Вторите обезбедуваат одржување на рамнотежа вода-сол поради осморегулација. Кај повеќеклеточните животни главно се наоѓаат дигестивни вакуоли.
Во растителните клетки, вакуоли се секогаш присутни; тие се опкружени со посебна мембрана и исполнети со клеточен сок. Мембраната што ја опкружува вакуолата е слична по хемиски состав, структура и функции на плазма мембраната. Клеточен соке воден раствор од различни неоргански и органски материи, вклучувајќи минерални соли, органски киселини, јаглени хидрати, протеини, гликозиди, алкалоиди итн. Вакуолата може да заземе до 90% од волуменот на клетката и да го турка јадрото до периферијата. Овој дел од клетката врши складишни, екскреторни, осмотски, заштитни, лизозомални и други функции, бидејќи акумулира хранливи материи и отпадни материи, обезбедува снабдување со вода и го одржува обликот и волуменот на клетката, а содржи и ензими за разградување на многу клеточни компоненти. Покрај тоа, биолошки активните супстанции на вакуолите можат да спречат многу животни да ги јадат овие растенија. Кај голем број растенија, поради отекување на вакуоли, растот на клетките се јавува со издолжување.
Вакуоли се присутни и во клетките на некои габи и бактерии, но кај габите тие вршат само функција на осморегулација, додека кај цијанобактериите одржуваат пловност и учествуваат во процесот на асимилација на азотот од воздухот.
Рибозоми- мали немембрански органели со дијаметар од 15-20 микрони, составени од две подединици - големи и мали. Еукариотските рибозомални подединици се собираат во јадрото, а потоа се транспортираат во цитоплазмата. Рибозомите кај прокариотите, митохондриите и пластидите се помали по големина од рибозомите кај еукариотите. Рибозомалните подединици вклучуваат rRNA и протеини.

Бројот на рибозоми во клетката може да достигне неколку десетици милиони: во цитоплазмата, митохондриите и пластидите тие се во слободна состојба, а на грубиот ER - во врзана состојба. Тие учествуваат во синтезата на протеините, особено го спроведуваат процесот на транслација - биосинтеза на полипептиден синџир на молекула на mRNA. Слободните рибозоми ги синтетизираат протеините на хијалоплазмата, митохондриите, пластидите и нивните сопствени рибозомални протеини, додека рибозомите прикачени на грубиот ER вршат превод на протеини за отстранување од клетките, склопување на мембраните и формирање на лизозоми и вакуоли.
Рибозомите може да се најдат поединечно во хијалоплазмата или собрани во групи за време на истовремена синтеза на неколку полипептидни синџири на една mRNA. Таквите групи на рибозоми се нарекуваат полирибозоми, или полисоми.

Микротубули- Тоа се цилиндрични шупливи немембрански органели кои продираат низ целата цитоплазма на клетката. Нивниот дијаметар е околу 25 nm, дебелината на ѕидот е 6-8 nm. Тие се формирани од бројни протеински молекули тубулин,кои прво формираат 13 нишки кои личат на монистра и потоа се собираат во микротубула. Микротубулите формираат цитоплазматичен ретикулум, кој и дава облик и волумен на клетката, ја поврзува плазматската мембрана со другите делови на клетката, обезбедува транспорт на супстанции низ клетката, учествува во движењето на клетката и интрацелуларните компоненти, како и во поделбата на генетскиот материјал. Тие се дел од клеточниот центар и органели на движење - флагели и цилии.
Микрофиламенти,или микронишки, исто така се немембрански органели, но тие имаат филаментозна форма и се формираат не од тубулин, туку актин. Тие учествуваат во процесите на мембрански транспорт, меѓуклеточно препознавање, поделба на клеточната цитоплазма и во нејзиното движење. Во мускулните клетки, интеракцијата на микрофиламентите на актин со филаментите на миозин посредува во контракцијата.
Микротубулите и микрофиламентите го формираат внатрешниот скелет на клетката - цитоскелет. Тоа е комплексна мрежа на влакна кои обезбедуваат механичка поддршка за плазма мембраната, го одредуваат обликот на клетката, локацијата на клеточните органели и нивното движење при клеточната делба.

Центар за клетки- немембранска органела сместена во животински клетки во близина на јадрото; го нема во растителните клетки. Неговата должина е околу 0,2-0,3 микрони, а дијаметарот е 0,1-0,15 микрони. Центарот на клетките е формиран од двајца центриоли, лежејќи во меѓусебно нормални рамнини, и зрачна сфераод микротубули. Секоја центриола е формирана од девет групи микротубули, собрани во групи од три, т.е. тројки. Клеточниот центар учествува во процесите на склопување на микротубули, поделба на наследниот материјал на клетката, како и во формирањето на флагели и цилии.

Органели на движење. ФлагелиИ цилииТие се клеточни израстоци покриени со плазмалема. Основата на овие органели е составена од девет пара микротубули лоцирани долж периферијата и две слободни микротубули во центарот. Микротубулите се меѓусебно поврзани со различни протеини, обезбедувајќи нивно координирано отстапување од оската - осцилација. Осцилациите се зависни од енергија, односно енергијата на високоенергетските АТП обврзници се троши на овој процес. Реставрацијата на изгубените флагели и цилии е функција базалните тела, или кинетозомителоцирани во нивната база.
Должината на цилиите е околу 10-15 nm, а должината на флагелите е 20-50 μm. Поради строго насочените движења на флагелите и цилиите, не се јавува само движење на едноклеточни животни, сперматозоиди и сл., туку и чистење на респираторниот тракт и движење на јајце клетката низ јајцеводите, бидејќи сите овие делови на човечкото тело се обложени со цилијарен епител.

Вклучувања
Вклучувања- Тоа се непостојани компоненти на клетката кои се формираат и исчезнуваат во текот на нејзиниот живот. Тие вклучуваат и резервни супстанции, на пример, зрна скроб или протеини во растителните клетки, гранули на гликоген во клетките на животните и габите, волутинот во бактериите, капките маснотии во сите видови клетки и отпадните производи, особено остатоците од храна несварен како резултат на фагоцитоза, формирајќи таканаречени резидуални тела.
Односот помеѓу структурата и функциите на деловите и органелите на клетката е основата на нејзиниот интегритет
Секој од деловите на клетката, од една страна, е посебна структура со специфична структура и функции, а од друга, компонента на покомплексен систем наречен клетка. Повеќето од наследни информации за еукариотската клетка се концентрирани во јадрото, но самото јадро не е во состојба да ја обезбеди неговата имплементација, бидејќи за тоа е потребна барем цитоплазмата, која делува како главна супстанција, и рибозомите, на кои се јавува оваа синтеза. . Повеќето рибозоми се наоѓаат на зрнестиот ендоплазматичен ретикулум, од каде што протеините најчесто се транспортираат до комплексот Голџи, а потоа, по модификацијата, до оние делови од клетката за кои се наменети или се излачуваат. Мембранското пакување на протеини и јаглени хидрати може да се вметне во мембраните на органелите и цитоплазматската мембрана, обезбедувајќи нивно постојано обновување. Лизозомите и вакуолите, кои вршат важни функции, исто така се одвојуваат од комплексот Голџи. На пример, без лизозоми, клетките брзо би се претвориле во еден вид депонија за отпадни молекули и структури.
Појавата на сите овие процеси бара енергија произведена од митохондриите, а кај растенијата од хлоропластите. И иако овие органели се релативно автономни, бидејќи имаат свои молекули на ДНК, некои од нивните протеини сè уште се кодирани од нуклеарниот геном и се синтетизираат во цитоплазмата.
Така, клетката е нераскинливо единство на нејзините составни компоненти, од кои секоја врши своја уникатна функција.
Метаболизмот и енергетската конверзија се својства на живите организми. Енергија и пластичен метаболизам, нивната врска. Фази на енергетскиот метаболизам. Ферментација и дишење. Фотосинтеза, неговото значење, космичка улога. Фази на фотосинтеза. Светлосни и темни реакции на фотосинтезата, нивната врска. Хемосинтеза. Улогата на хемосинтетичките бактерии на Земјата
Метаболизам и конверзија на енергија - својства на живите организми
Една клетка може да се спореди со минијатурна хемиска фабрика во која се случуваат стотици и илјадници хемиски реакции.
Метаболизам- збир на хемиски трансформации насочени кон зачувување и саморепродукција на биолошки системи.
Вклучува внесување на супстанции во телото за време на исхрана и дишење, интрацелуларен метаболизам или метаболизмот, како и изолација на финалните метаболички производи.
Метаболизмот е нераскинливо поврзан со процесите на претворање на еден вид енергија во друг. На пример, во процесот на фотосинтеза, светлосната енергија се складира во форма на енергија на хемиски врски на сложени органски молекули, а во процесот на дишење се ослободува и троши за синтеза на нови молекули, механичка и осмотска работа, дисипани во форма на топлина итн.
Појавата на хемиски реакции кај живите организми е обезбедена благодарение на биолошките катализатори од протеинска природа - ензими, или ензими. Како и другите катализатори, ензимите ја забрзуваат појавата на хемиски реакции во клетката за десетици и стотици илјади пати, а понекогаш дури и ги овозможуваат, но не ја менуваат природата или својствата на финалниот производ(ите) од реакцијата и прават не се менуваат себеси. Ензимите можат да бидат и едноставни и сложени протеини, кои, покрај протеинскиот дел, вклучуваат и непротеински дел - кофактор (коензим). Примери за ензими се плунковната амилаза, која ги разградува полисахаридите при продолжено џвакање и пепсинот, кој обезбедува варење на протеините во желудникот.
Ензимите се разликуваат од непротеинските катализатори по нивната висока специфичност на дејство, значително зголемување на брзината на реакцијата со нивна помош, како и способноста да се регулира дејството со менување на условите на реакцијата или интеракцијата на различни супстанции со нив. Покрај тоа, условите под кои се случува ензимската катализа значително се разликуваат од оние под кои се случува неензимска катализа: оптималната температура за функционирање на ензимите во човечкото тело е $37°C$, притисокот треба да биде блиску до атмосферскиот, а $pH$ на околината може значително да се двоуми. Така, амилазата бара алкална средина, а пепсин бара кисела средина.
Механизмот на дејство на ензимите е да ја намали енергијата на активирање на супстанциите (супстратите) кои влегуваат во реакција поради формирање на средно ензимско-супстрат комплекси.

Енергија и пластичен метаболизам, нивната врска
Метаболизмот се состои од два процеси кои се случуваат истовремено во клетката: пластика и енергетски метаболизам.
Пластичен метаболизам (анаболизам, асимилација)е збир на синтезни реакции кои вклучуваат трошење на АТП енергија. Во процесот на пластичен метаболизам се синтетизираат органски материи неопходни за клетката. Примери за пластична размена на реакции се фотосинтезата, биосинтезата на протеините и репликацијата на ДНК (самодуплирање).
Енергетски метаболизам (катаболизам, дисимилација)е збир на реакции кои ги разградуваат сложените материи на поедноставни. Како резултат на енергетскиот метаболизам, енергијата се ослободува и складира во форма на АТП. Најважните процеси на енергетскиот метаболизам се дишењето и ферментацијата.
Пластиката и енергетската размена се нераскинливо поврзани, бидејќи во процесот на пластична размена се синтетизираат органски материи и за тоа е потребна АТП енергија, а во процесот на размена на енергија органските материи се разградуваат и се ослободува енергија, која потоа ќе се троши на процесите на синтеза. .
Организмите добиваат енергија за време на процесот на исхрана, а ја ослободуваат и ја претвораат во достапна форма главно за време на процесот на дишење. Според начинот на исхрана, сите организми се делат на автотрофи и хетеротрофи. Автотрофиспособни самостојно да синтетизираат органски материи од неоргански и хетеротрофикористете исклучиво подготвени органски материи.
Фази на енергетскиот метаболизам
И покрај сложеноста на реакциите на енергетскиот метаболизам, конвенционално е поделен на три фази: подготвителна, анаеробна (без кислород) и аеробна (кислород).
На подготвителна фазамолекули на полисахариди, липиди, протеини, нуклеински киселини се распаѓаат на поедноставни, на пример, гликоза, глицерол и масни киселини, амино киселини, нуклеотиди итн. Оваа фаза може да се појави директно во клетките или во цревата, од каде што надолу супстанции се испорачуваат преку крвотокот.
Анаеробна фазаенергетскиот метаболизам е придружен со понатамошно распаѓање на мономери на органски соединенија во уште поедноставни меѓупроизводи, на пример, пирувична киселина или пируват. Не бара присуство на кислород, а за многу организми кои живеат во калта на мочуриштата или во човечките црева, тоа е единствениот начин да се добие енергија. Анаеробната фаза на енергетскиот метаболизам се јавува во цитоплазмата.
Различни супстанции можат да претрпат расцепување без кислород, но доста често супстратот на реакциите е гликозата. Процесот на негово разделување без кислород се нарекува гликолиза. За време на гликолизата, молекулата на гликоза губи четири атоми на водород, т.е. се оксидира и се формираат две молекули пирувична киселина, две молекули АТП и две молекули на редуцираниот водороден носач $NADH + H^(+)$:
$C_6H_(12)O_6 + 2H_3PO_4 + 2ADP + 2NAD → 2C_3H_4O_3 + 2ATP + 2NADH + H^(+) + 2H_2O$.
Формирањето на АТП од АДП настанува поради директен трансфер на фосфатниот анјон од предфосфорилираниот шеќер и се нарекува. фосфорилација на подлогата.
Аеробна фазаразмена на енергија може да се случи само во присуство на кислород, додека меѓусоединенијата формирани за време на расцепувањето без кислород се оксидираат до крајните производи (јаглерод диоксид и вода) и најголемиот дел од енергијата складирана во хемиските врски на органските соединенија се ослободува. Се претвора во енергија на високо-енергетски врски од 36 ATP молекули. Оваа фаза е исто така наречена ткиво дишење. Во отсуство на кислород, меѓусоединенијата се претвораат во други органски материи, процес наречен ферментација.
Здив
Механизмот на клеточно дишење е шематски прикажан на Сл.

Аеробното дишење се јавува во митохондриите, при што пирувична киселина прво губи еден јаглероден атом, што е придружено со синтеза на еден редукциски еквивалент од $NADH + H^(+) $ и молекула на ацетил коензим А (ацетил-CoA):
$C_3H_4O_3 + NAD + H~CoA → CH_3CO~CoA + NADH + H^(+) + CO_2$.
Ацетил-CoA во митохондријалната матрица е вклучен во синџир на хемиски реакции, чиј тотал се нарекува Кребсовиот циклус (циклус на трикарбоксилна киселина, циклус на лимонска киселина). За време на овие трансформации, се формираат две ATP молекули, ацетил-CoA целосно се оксидира до јаглерод диоксид, а неговите водородни јони и електрони се додаваат на водородните носители $NADH + H^(+)$ и $FADH_2$. Носачите транспортираат водородни протони и електрони до внатрешните мембрани на митохондриите, формирајќи кристаи. Со помош на носечки протеини, водородните протони се пумпаат во меѓумембранскиот простор, а електроните се пренесуваат преку таканаречениот респираторен синџир на ензими лоцирани на внатрешната мембрана на митохондриите и се испуштаат на атоми на кислород:
$O_2+2e^(-)→O_2^-$.
Треба да се напомене дека некои протеини од респираторниот синџир содржат железо и сулфур.
Од меѓумембранскиот простор, водородните протони се транспортираат назад во митохондријалната матрица со помош на специјални ензими - ATP синтази, а енергијата што се ослободува во овој случај се троши на синтеза на 34 ATP молекули од секоја молекула на гликоза. Овој процес се нарекува оксидативна фосфорилација. Во митохондријалната матрица, водородните протони реагираат со кислородните радикали за да формираат вода:
$4H^(+)+O_2^-→2H_2O$.
Збирот на реакции на кислородно дишење може да се изрази на следниов начин:
$2C_3H_4O_3 + 6O_2 + 36H_3PO_4 + 36ADP → 6CO_2 + 38H_2O + 36ATP.$
Целокупната равенка за дишење изгледа вака:
$C_6H_(12)O_6 + 6O_2 + 38H_3PO_4 + 38ADP → 6CO_2 + 40H_2O + 38ATP.$
Ферментација
Во отсуство на кислород или негов недостаток, се јавува ферментација. Ферментацијата е еволутивно порана метода за добивање енергија отколку дишењето, но енергетски е помалку корисна бидејќи ферментацијата произведува органски материи кои се уште се богати со енергија. Постојат неколку главни типови на ферментација: млечна киселина, алкохолна, оцетна киселина итн. Така, во скелетните мускули во отсуство на кислород за време на ферментацијата, пирувична киселина се редуцира на млечна киселина, додека претходно формираните редукциони еквиваленти се консумираат и само остануваат две ATP молекули:
$2C_3H_4O_3 + 2NADH + H^(+) → 2C_3H_6O_3 + 2NAD$.
За време на ферментацијата со помош на квасец, пирувична киселина во присуство на кислород се претвора во етил алкохол и јаглерод моноксид (IV):
$C_3H_4O_3 + NADH + H^(+) → C_2H_5OH + CO_2 + NAD^(+)$.
При ферментација со помош на микроорганизми, од пирувична киселина може да се формираат и оцетна, маслена, мравја киселина и др.
АТП, добиен како резултат на енергетскиот метаболизам, се троши во клетката за различни видови на работа: хемиска, осмотска, електрична, механичка и регулаторна. Хемиската работа вклучува биосинтеза на протеини, липиди, јаглени хидрати, нуклеински киселини и други витални соединенија. Осмотската работа ги вклучува процесите на апсорпција од клетката и отстранување од неа на супстанции кои се во екстрацелуларниот простор во концентрации поголеми отколку во самата клетка. Електричната работа е тесно поврзана со осмотската работа, бидејќи како резултат на движењето на наелектризираните честички низ мембраните се формира мембрански полнеж и се стекнуваат својствата на ексцитабилност и спроводливост. Механичката работа вклучува движење на супстанции и структури внатре во клетката, како и на клетката како целина. Регулаторната работа ги вклучува сите процеси насочени кон координирање на процесите во ќелијата.
Фотосинтеза, неговото значење, космичка улога
Фотосинтезае процес на претворање на светлосната енергија во енергија на хемиски врски на органски соединенија со учество на хлорофил.
Како резултат на фотосинтезата, годишно се произведуваат околу 150 милијарди тони органска материја и приближно 200 милијарди тони кислород. Овој процес го обезбедува циклусот на јаглерод во биосферата, спречувајќи акумулација на јаглерод диоксид и со тоа спречување на ефектот на стаклена градина и прегревање на Земјата. Органските супстанции формирани како резултат на фотосинтезата не се целосно консумирани од други организми; значителен дел од нив во текот на милиони години формирале депозити на минерали (тврд и кафеав јаглен, нафта). Неодамна, маслото од репка („биодизел“) и алкохолот добиен од растителни остатоци исто така почнаа да се користат како гориво. Озонот се формира од кислород под влијание на електрични празнења, што формира озонски екран кој го штити целиот живот на Земјата од деструктивните ефекти на ултравиолетовите зраци.
Нашиот сонародник, извонредниот физиолог на растенијата К.
Фази на фотосинтеза. Светлосни и темни реакции на фотосинтезата, нивната врска
Во 1905 година, англискиот растителен физиолог Ф. Блекман открил дека стапката на фотосинтеза не може да се зголемува бесконечно; некој фактор ја ограничува. Врз основа на ова, тој претпостави дека постојат две фази на фотосинтезата: светлинаИ темно. При низок интензитет на светлина, брзината на светлосните реакции се зголемува пропорционално со зголемувањето на интензитетот на светлината, а покрај тоа, овие реакции не зависат од температурата, бидејќи не бараат ензими да се појават. На тилакоидните мембрани се јавуваат светлосни реакции.
Брзината на темните реакции, напротив, се зголемува со зголемување на температурата, меѓутоа, по достигнувањето на прагот на температурата од $30°C $, ова зголемување престанува, што укажува на ензимската природа на овие трансформации што се случуваат во стромата. Треба да се напомене дека светлината има одредено влијание и врз темните реакции, и покрај тоа што тие се нарекуваат темни реакции.

Лесната фаза на фотосинтезата се јавува на тилакоидните мембрани кои носат неколку видови протеински комплекси, од кои главни се фотосистеми I и II, како и АТП синтаза. Фотосистемите вклучуваат пигментни комплекси, кои покрај хлорофилот содржат и каротеноиди. Каротеноидите ја фаќаат светлината во областите на спектарот каде што не ја прави хлорофилот, а исто така го штитат хлорофилот од уништување со светлина со висок интензитет.
Покрај пигментните комплекси, фотосистемите вклучуваат и голем број протеини кои примаат електрони, кои последователно ги пренесуваат електроните од молекулите на хлорофилот едни на други. Редоследот на овие протеини се нарекува транспортен ланец на електрони на хлоропласти.
Посебен комплекс на протеини е поврзан и со фотосистемот II, кој обезбедува ослободување на кислород за време на фотосинтезата. Овој комплекс за ослободување на кислород содржи јони на манган и хлор.
ВО лесна фазасветлосни кванти или фотони, кои паѓаат на молекулите на хлорофилот лоцирани на тилакоидните мембрани, ги пренесуваат во возбудена состојба, која се карактеризира со поголема електронска енергија. Во овој случај, возбудените електрони од хлорофилот на фотосистемот I се пренесуваат преку синџир на посредници до водородниот носач NADP, кој прикачува водородни протони, секогаш присутни во воден раствор:
$NADP + 2e^(-) + 2H^(+) → NADPH + H^(+)$.
Намалените $NADPH + H^(+)$ последователно ќе се користат во темната фаза. Електроните од хлорофилот на фотосистемот II исто така се пренесуваат по синџирот на транспорт на електрони, но тие ги пополнуваат „електронските дупки“ на хлорофилот на фотосистемот I. Недостигот на електрони во хлорофилот на фотосистемот II се пополнува со одземање на молекули на вода, кои се јавува со учество на комплексот за ослободување на кислород веќе споменат погоре. Како резултат на распаѓањето на молекулите на водата, што се нарекува фотолиза, се формираат водородни протони и се ослободува молекуларен кислород, кој е нуспроизвод на фотосинтезата:
$H_2O → 2H^(+) + 2e^(-) + (1)/(2)O_2$.
Генетски информации во клетка. Гени, генетски код и неговите својства. Матрична природа на реакциите на биосинтезата. Биосинтеза на протеини и нуклеински киселини
Генетски информации во клетка
Репродукцијата на сопствениот вид е едно од основните својства на живите суштества. Благодарение на овој феномен, постои сличност не само помеѓу организмите, туку и помеѓу поединечните клетки, како и нивните органели (митохондрии и пластиди). Материјалната основа на оваа сличност е преносот на генетски информации шифрирани во ДНК нуклеотидната секвенца, што се врши преку процесите на репликација на ДНК (самодуплирање). Сите карактеристики и својства на клетките и организмите се реализираат благодарение на протеините, чија структура првенствено е одредена од низата на ДНК нуклеотиди. Затоа, биосинтезата на нуклеинските киселини и протеините има огромно значење во метаболичките процеси. Структурната единица на наследни информации е генот.
Гени, генетски код и неговите својства
Наследните информации во клетката не се монолитни, тие се поделени на посебни „зборови“ - гени.
гене елементарна единица на генетски информации.
Работата на програмата „Човечки геном“, која беше спроведена истовремено во неколку земји и беше завршена на почетокот на овој век, ни даде разбирање дека една личност има само околу 25-30 илјади гени, но информации од поголемиот дел од нашата ДНК никогаш не се чита, бидејќи содржи огромен број бесмислени делови, повторувања и гени кои кодираат особини кои го изгубиле значењето за луѓето (опашка, влакна на телото итн.). Дополнително, дешифрирани се голем број гени одговорни за развој на наследни болести, како и гени за целни лекови. Сепак, практичната примена на резултатите добиени при спроведувањето на оваа програма се одложува додека не се дешифрираат геномите на повеќе луѓе и не стане јасно како тие се разликуваат.
Се нарекуваат гени кои ја кодираат примарната структура на протеинот, рибозомалната или трансферната РНК структурнии гени кои обезбедуваат активирање или потиснување на читање информации од структурни гени - регулаторни. Сепак, дури и структурните гени содржат регулаторни региони.
Наследните информации на организмите се шифрирани во ДНК во форма на одредени комбинации на нуклеотиди и нивната низа - генетски код. Неговите својства се: тројност, специфичност, универзалност, вишок и непреклопување. Покрај тоа, во генетскиот код нема интерпункциски знаци.
Секоја аминокиселина е кодирана во ДНК со три нуклеотиди - тројка,на пример, метионинот е кодиран од тројката TAC, односно кодот е тројка. Од друга страна, секоја тројка шифрира само една аминокиселина, што е нејзината специфичност или недвосмисленост. Генетскиот код е универзален за сите живи организми, односно наследни информации за човечките протеини може да ги читаат бактериите и обратно. Ова укажува на единството на потеклото на органскиот свет. Сепак, 64 комбинации од три нуклеотиди одговараат на само 20 амино киселини, како резултат на што една амино киселина може да биде кодирана од 2-6 тројки, односно генетскиот код е излишен или дегенериран. Три тројки немаат соодветни амино киселини, тие се нарекуваат стоп кодони, бидејќи тие укажуваат на крајот на синтезата на полипептидниот синџир.
Редоследот на бази во ДНК тројки и аминокиселините што тие ги кодираат
*Стоп кодон, што укажува на крајот на синтезата на полипептидниот синџир.
Кратенки за имиња на амино киселини:
Ала - аланин
Арг - аргинин
Асн - аспарагин
Asp - аспарагинска киселина
Вал - валин
Неговиот - хистидин
Гли - глицин
Gln - глутамин
Глу - глутаминска киселина
Иле - изолеуцин
Леу - леуцин
Лиз - лизин
Мет - метионин
Про - пролин
Сер - серин
Тир - тирозин
Тре - треонин
Три - триптофан
Фен - фенилаланин
Cys - цистеин
Ако почнете да читате генетски информации не од првиот нуклеотид во тројката, туку од вториот, тогаш не само што ќе се помести рамката за читање, туку и протеинот синтетизиран на овој начин ќе биде сосема различен не само во нуклеотидната низа, туку и во структура и својства. Меѓу тројките нема интерпункциски знаци, така што нема пречки за поместување на рамката за читање, што отвора простор за појава и одржување на мутации.
Матрична природа на реакциите на биосинтезата
Бактериските клетки се способни да се удвојуваат на секои 20-30 минути, а еукариотските клетки - секој ден и уште почесто, што бара голема брзина и точност на репликацијата на ДНК. Покрај тоа, секоја клетка содржи стотици и илјадници копии на многу протеини, особено ензими, затоа, методот „парчено“ на нивно производство е неприфатлив за нивна репродукција. Попрогресивен метод е печат, кој ви овозможува да добиете бројни точни копии на производот и исто така да ја намалите неговата цена. За печат, потребна е матрица од која се прави отпечатокот.
Во клетките, принципот на синтеза на шаблоните е дека новите молекули на протеини и нуклеински киселини се синтетизираат во согласност со програмата вградена во структурата на веќе постоечките молекули на истите нуклеински киселини (ДНК или РНК).
Биосинтеза на протеини и нуклеински киселини
репликација на ДНК.ДНК е двоверижен биополимер, чии мономери се нуклеотиди. Доколку биосинтезата на ДНК би се случила на принципот на фотокопирање, тогаш неизбежно би се појавиле бројни искривувања и грешки во наследни информации, што на крајот би довело до смрт на нови организми. Затоа, процесот на удвојување на ДНК се случува поинаку, на полуконзервативен начин: молекулата на ДНК се одмотува, а на секоја од синџирите се синтетизира нов синџир според принципот на комплементарност. Процесот на само-репродукција на молекула на ДНК, обезбедувајќи точно копирање на наследни информации и нивно пренесување од генерација на генерација, се нарекува. репликација(од лат. репликација- повторување). Како резултат на репликацијата, се формираат две апсолутно точни копии на матичната молекула на ДНК, од кои секоја носи по една копија од матичната ДНК молекула.

Процесот на репликација е всушност исклучително сложен, бидејќи голем број на протеини се вклучени во него. Некои од нив ја одмотуваат двојната спирала на ДНК, други ги кршат водородните врски помеѓу нуклеотидите на комплементарните синџири, други (на пример, ензимот ДНК полимераза) избираат нови нуклеотиди врз основа на принципот на комплементарност итн. Две молекули на ДНК формирани како резултат на репликација се разминуваат на два за време на поделбата новоформираните ќерки клетки.
Грешките во процесот на репликација се случуваат исклучително ретко, но ако се појават, тие многу брзо се елиминираат и од ДНК полимеразите и од специјалните ензими за поправка, бидејќи секоја грешка во нуклеотидната низа може да доведе до неповратна промена во структурата и функциите на протеинот. и, во крајна линија, негативно влијае на одржливоста на нова клетка или дури и поединец.
Биосинтеза на протеини.Како што фигуративно рече извонредниот филозоф од 19 век Ф. Енгелс: „Животот е форма на постоење на протеински тела“. Структурата и својствата на протеинските молекули се одредуваат според нивната примарна структура, т.е. низата на амино киселини кодирани во ДНК. Не само постоењето на самиот полипептид, туку и функционирањето на клетката како целина зависи од точноста на репродукцијата на оваа информација, затоа процесот на синтеза на протеини е од големо значење. Се чини дека тоа е најкомплексниот процес на синтеза во клетката, бидејќи вклучува до триста различни ензими и други макромолекули. Покрај тоа, тече со голема брзина, што бара уште поголема прецизност.
Постојат две главни фази во биосинтезата на протеините: транскрипција и транслација.
Транскрипција(од лат. транскрипција- препишување) е биосинтеза на mRNA молекули на ДНК матрица.

Бидејќи молекулата на ДНК содржи два антипаралелни синџири, читањето информации од двата синџири би довело до формирање на сосема различни mRNA, па затоа нивната биосинтеза е можна само на еден од синџирите, кој се нарекува кодирање, или кодоген, за разлика од вториот, некодирачки или некодогени. Процесот на препишување е обезбеден со посебен ензим, РНК полимераза, кој ги избира нуклеотидите на РНК според принципот на комплементарност. Овој процес може да се случи и во јадрото и во органели кои имаат своја ДНК - митохондрии и пластиди.
Молекулите на mRNA синтетизирани за време на транскрипцијата се подложени на сложен процес на подготовка за транслација (митохондријалните и пластидните mRNA може да останат внатре во органелите, каде што се случува втората фаза од биосинтезата на протеините). За време на процесот на созревање на mRNA, првите три нуклеотиди (AUG) и опашка од аденил нуклеотиди се прикачени на него, чија должина одредува колку копии од протеинот може да се синтетизираат на дадена молекула. Само тогаш зрелите mRNA го напуштаат јадрото низ нуклеарните пори.
Паралелно, процесот на активирање на аминокиселините се случува во цитоплазмата, при што аминокиселината се приклучува на соодветната слободна tRNA. Овој процес се катализира со посебен ензим и бара АТП.
Емитување(од лат. емитување- трансфер) е биосинтеза на полипептиден синџир на mRNA матрица, при што генетските информации се преведуваат во амино киселинската секвенца на полипептидниот синџир.

Втората фаза на синтеза на протеини најчесто се јавува во цитоплазмата, на пример на грубиот ЕР. За нејзино појавување, неопходно е присуство на рибозоми, активирање на tRNA, при што тие ги закачуваат соодветните амино киселини, присуство на јони Mg2+, како и оптимални услови на животната средина (температура, pH, притисок и сл.).
За да започнете со емитување ( иницијација) мала рибозомална подединица е прикачена на молекула на mRNA подготвена за синтеза, а потоа, според принципот на комплементарност на првиот кодон (AUG), се избира tRNA што ја носи аминокиселината метионин. Само после ова се прикачува големата рибозомална подединица. Во склопениот рибозом има два кодони на мРНК, од кои првиот е веќе окупиран. Втората тРНК, која исто така носи амино киселина, се додава во кодонот што е во непосредна близина до него, по што се формира пептидна врска помеѓу остатоците од аминокиселините со помош на ензими. Рибозомот поместува еден кодон од mRNA; првата tRNA ослободена од амино киселина се враќа во цитоплазмата по следната амино киселина, а фрагмент од идниот полипептиден синџир виси, како да е, на преостанатата tRNA. Следната tRNA е прикачена на новиот кодон кој се наоѓа во рамките на рибозомот, процесот се повторува и чекор по чекор полипептидниот синџир се издолжува, т.е. издолжување.
Крај на синтезата на протеини ( раскинување) се јавува веднаш штом ќе се сретне специфична нуклеотидна секвенца во молекулата на mRNA која не кодира за амино киселина (стоп-кодон). По ова, рибозомот, mRNA и полипептидниот синџир се одвојуваат, а новосинтетизираниот протеин добива соодветна структура и се транспортира до делот од клетката каде што ќе ги извршува своите функции.
Преведувањето е многу енергетски интензивен процес, бидејќи енергијата на една ATP молекула се троши за прикачување на една амино киселина на tRNA, а уште неколку се користат за движење на рибозомот по молекулата на mRNA.
За да се забрза синтезата на одредени протеински молекули, неколку рибозоми може последователно да се прикачат на молекула на mRNA, кои формираат единствена структура - полисом.
Клетката е генетска единица на живо суштество. Хромозоми, нивната структура (облик и големина) и функции. Бројот на хромозоми и нивната константност на видот. Соматски и герминативни клетки. Животен циклус на клетките: интерфаза и митоза. Митозата е поделба на соматски клетки. Мејоза. Фази на митоза и мејоза. Развој на герминативните клетки кај растенијата и животните. Клеточната делба е основа за раст, развој и репродукција на организмите. Улогата на мејозата и митозата
Клетката е генетска единица на живо суштество.
И покрај фактот дека нуклеинските киселини се носители на генетски информации, имплементацијата на овие информации е невозможна надвор од клетката, што лесно се докажува со примерот на вирусите. Овие организми, кои често содржат само ДНК или РНК, не можат да се репродуцираат независно; за да го направат тоа, тие мора да го користат наследниот апарат на клетката. Тие дури и не можат да навлезат во клетка без помош на самата клетка, освен преку употреба на механизми за транспорт на мембраната или поради оштетување на клетките. Повеќето вируси се нестабилни; тие умираат по само неколку часа изложеност на отворено. Следствено, клетката е генетска единица на живо суштество, која има минимален сет на компоненти за зачувување, менување и спроведување на наследни информации, како и нејзино пренесување на потомците.
Повеќето генетски информации на еукариотската клетка се наоѓаат во јадрото. Особеноста на неговата организација е тоа што, за разлика од ДНК на прокариотска клетка, молекулите на ДНК на еукариотите не се затворени и формираат комплексни комплекси со протеини - хромозоми.
Хромозоми, нивната структура (облик и големина) и функции
Хромозом(од грчки хром- боја, боење и сома- тело) е структура на клеточното јадро, кое содржи гени и носи одредени наследни информации за карактеристиките и својствата на организмот.
Понекогаш кружните молекули на ДНК на прокариотите се нарекуваат и хромозоми. Хромозомите се способни за самодуплирање; тие имаат структурна и функционална индивидуалност и ја задржуваат со генерации. Секоја клетка ги носи сите наследни информации на телото, но само мал дел работи во неа.
Основата на хромозомот е двоверижна молекула на ДНК преполна со протеини. Кај еукариотите, хистонските и нехистонските протеини комуницираат со ДНК, додека кај прокариотите, хистонските протеини се отсутни.
Хромозомите најдобро се гледаат под светлосен микроскоп за време на клеточната делба, кога, како резултат на набивање, добиваат изглед на тела во облик на прачка, одделени со примарно стегање - центромер — на рамениците. На хромозомот исто така може да има секундарна констрикција, што во некои случаи го одвојува т.н сателит. Краевите на хромозомите се нарекуваат теломерите. Теломерите ги спречуваат краевите на хромозомите да се залепат заедно и обезбедуваат нивно прицврстување на нуклеарната мембрана во клетка што не се дели. На почетокот на поделбата, хромозомите се удвојуваат и се состојат од два ќерки хромозома - хроматид, прицврстена на центромерот.

Според нивната форма, хромозомите се делат на хромозоми со еднакви раце, нееднакви вооружени и шипки. Големините на хромозомите значително се разликуваат, но просечниот хромозом има димензии од 5 $×$ 1,4 микрони.
Во некои случаи, хромозомите, како резултат на бројни дупликации на ДНК, содржат стотици и илјадници хроматиди: таквите гигантски хромозоми се нарекуваат политен. Тие се наоѓаат во плунковните жлезди на ларвите на Drosophila, како и во дигестивните жлезди на кружните црви.
Бројот на хромозоми и нивната константност на видот. Соматски и герминативни клетки
Според клеточната теорија, клетката е единица на структура, витална активност и развој на организмот. Така, на клеточно ниво се обезбедуваат такви важни функции на живите суштества како раст, репродукција и развој на организмот. Клетките на повеќеклеточните организми можат да се поделат на соматски и репродуктивни клетки.
Соматски клетки- тоа се сите клетки на телото формирани како резултат на митотична поделба.
Студијата за хромозомите овозможи да се утврди дека соматските клетки на телото на секој биолошки вид се карактеризираат со постојан број на хромозоми. На пример, едно лице има 46. Множеството хромозоми на соматски клетки се нарекува диплоидна(2n), или двојно.

Полови клетки, или гамети, се специјализирани клетки кои се користат за сексуална репродукција.
Гаметите секогаш содржат половина колку хромозоми од соматските клетки (кај луѓето - 23), затоа збирот на хромозоми на герминативните клетки се нарекува хаплоиден(n), или сингл. Неговото формирање е поврзано со мејотичната клеточна делба.
Количината на ДНК во соматските клетки е означена како 2c, а во половите клетки - 1c. Генетската формула на соматските клетки е напишана како 2n2c, а половите клетки - 1n1c.
Во јадрата на некои соматски клетки, бројот на хромозоми може да се разликува од нивниот број во соматските клетки. Ако оваа разлика е поголема од една, две, три итн. хаплоидни множества, тогаш таквите ќелии се нарекуваат полиплоиден(три-, тетра-, пентаплоиден, соодветно). Во таквите клетки, метаболичките процеси обично се одвиваат многу интензивно.
Бројот на хромозоми сам по себе не е карактеристика специфична за видот, бидејќи различни организми можат да имаат еднаков број на хромозоми, но сродните може да имаат различен број. На пример, маларичниот плазмодиум и коњскиот кружен црв имаат по два хромозома, додека луѓето и шимпанзата имаат 46 и 48, соодветно.
Човечките хромозоми се поделени во две групи: автозоми и полови хромозоми (хетерохромозоми). Автосомнаво човечките соматски клетки има 22 пара, тие се исти за мажи и жени, и полови хромозомисамо еден пар, но токму тоа го одредува полот на поединецот. Постојат два вида полови хромозоми - X и Y. Клетките на телото на жените носат два Х хромозоми, а машките - X и Y.
Кариотип- ова е збир на карактеристики на хромозомскиот сет на еден организам (бројот на хромозоми, нивната форма и големина).
Условниот запис на кариотипот го вклучува вкупниот број на хромозоми, полови хромозоми и можните отстапувања во збирот на хромозоми. На пример, кариотипот на нормален маж е напишан како 46, XY, а кариотипот на нормална жена е 46, XX.
Животен циклус на клетките: интерфаза и митоза
Клетките не се појавуваат одново секој пат, тие се формираат само како резултат на поделбата на матичните клетки. По поделбата, на ќерките клетки им е потребно извесно време да формираат органели и да добијат соодветна структура која би обезбедила извршување на одредена функција. Овој временски период се нарекува созревање.
Се нарекува временскиот период од појавата на клетка како резултат на делба до нејзината поделба или смрт животниот циклус на клетката.
Во еукариотските клетки, животниот циклус е поделен на две главни фази: интерфаза и митоза.
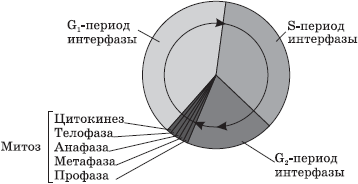
Интерфаза- ова е временски период во животниот циклус во кој клетката не се дели и нормално функционира. Интерфазата е поделена на три периоди: G 1 -, S- и G 2 -период.
G 1 -период(пресинтетички, постмитотски) е период на клеточен раст и развој во кој се јавува активна синтеза на РНК, протеини и други супстанции неопходни за целосна животна поддршка на новоформираната клетка. Кон крајот на овој период, клетката може да почне да се подготвува за дуплирање на нејзината ДНК.
ВО S-период(синтетички) се јавува самиот процес на репликација на ДНК. Единствениот дел од хромозомот кој не подлежи на репликација е центромерот, така што добиените молекули на ДНК не се разминуваат целосно, туку остануваат заедно во него, а на почетокот на поделбата хромозомот има изглед во облик на Х. Генетската формула на клетката по удвојувањето на ДНК е 2n4c. Исто така во S-периодот, центриолите на клеточниот центар се удвоени.
G 2 -период(постсинтетички, премитотски) се карактеризира со интензивна синтеза на РНК, протеини и АТП неопходни за процесот на клеточната делба, како и раздвојување на центриоли, митохондрии и пластиди. До крајот на интерфазата, хроматинот и јадрото остануваат јасно да се разликуваат, интегритетот на нуклеарната обвивка не е нарушен и органелите не се менуваат.
Некои од телесните клетки се способни да ги извршуваат своите функции во текот на целиот животен век на телото (неврони на нашиот мозок, мускулни клетки на срцето), додека други постојат кратко време, по што умираат (интестинални епителни клетки, епидермални клетки на кожата). Следствено, телото мора постојано да се подложува на процеси на клеточна делба и формирање на нови кои би ги замениле мртвите. Се нарекуваат клетки способни за делење стебло. Во човечкото тело се наоѓаат во црвената коскена срцевина, во длабоките слоеви на епидермисот на кожата и на други места. Користејќи ги овие клетки, можете да одгледувате нов орган, да постигнете подмладување, а исто така и да го клонирате телото. Изгледите за користење на матични клетки се апсолутно јасни, но моралните и етичките аспекти на овој проблем сè уште се дискутираат, бидејќи во повеќето случаи се користат ембрионски матични клетки добиени од човечки ембриони убиени за време на абортусот.
Времетраењето на интерфазата во растителните и животинските клетки е во просек 10-20 часа, додека митозата трае околу 1-2 часа.
За време на последователните поделби во повеќеклеточните организми, ќерките ќерки стануваат сè поразновидни додека читаат информации од зголемен број гени.
Некои клетки престануваат да се делат со текот на времето и умираат, што може да се должи на завршување на одредени функции, како во случајот со епидермалните кожни клетки и крвните клетки, или поради оштетување на овие клетки од фактори на животната средина, особено патогени. Генетски програмираната клеточна смрт се нарекува апоптоза, додека случајна смрт - некроза.
Митозата е поделба на соматски клетки. Фази на митоза
Митоза- метод на индиректна поделба на соматски клетки.
За време на митозата, клетката поминува низ низа последователни фази, како резултат на што секоја клетка ќерка го добива истиот сет на хромозоми како и во матичната клетка.
Митозата е поделена на четири главни фази: профаза, метафаза, анафаза и телофаза. Профаза- најдолгиот стадиум на митоза, при кој хроматинот се кондензира, што резултира со тоа што хромозоми во облик на Х кои се состојат од два хроматиди (ќерки хромозоми) стануваат видливи. Во овој случај, јадрото исчезнува, центриолите се разминуваат до половите на клетката и почнува да се формира ахроматинско вретено (вретено на поделба) од микротубулите. На крајот на профазата, нуклеарната мембрана се распаѓа во посебни везикули.

ВО метафазаХромозомите се наредени по екваторот на клетката со нивните центромери, на кои се прикачени микротубулите на целосно формираното вретено. Во оваа фаза на поделба, хромозомите се најкомпактирани и имаат карактеристична форма, што овозможува да се проучува кариотипот.
ВО анафазаБрзата репликација на ДНК се јавува кај центромерите, како резултат на што хромозомите се делат и хроматидите се разминуваат до половите на клетката, растегнати со микротубули. Распределбата на хроматидите мора да биде апсолутно еднаква, бидејќи токму овој процес обезбедува одржување на константен број на хромозоми во клетките на телото.
На сцената телофазиќерките хромозоми се собираат на половите, околу нив од везикулите се формираат деспирални, нуклеарни мембрани, а во новоформираните јадра се појавуваат јадра.
По нуклеарна поделба, се јавува цитоплазматска поделба - цитокинеза,при што настанува повеќе или помалку рамномерна распределба на сите органели на матичната клетка.
Така, како резултат на митоза, две ќерки ќерки се формираат од една мајка клетка, од кои секоја е генетска копија на матичната клетка (2n2c).
Кај болните, оштетените, старечките клетки и специјализираните ткива на телото, може да дојде до малку поинаков процес на поделба - амитоза. Амитозанаречена директна поделба на еукариотските клетки, во која не се јавува формирање на генетски еквивалентни клетки, бидејќи клеточните компоненти се нерамномерно распоредени. Се наоѓа кај растенијата во ендоспермата, а кај животните - во црниот дроб, 'рскавицата и рожницата на окото.
Мејоза. Фази на мејоза
Мејозае метод на индиректна поделба на примарните герминативни клетки (2n2c), што резултира со формирање на хаплоидни клетки (1n1c), најчесто герминативни клетки.
За разлика од митозата, мејозата се состои од две последователни клеточни делби, од кои на секоја и претходи интерфаза. Првата поделба на мејозата (мејоза I) се нарекува редукционист, бидејќи во овој случај бројот на хромозоми е преполовен, а втората поделба (мејоза II) - равенка, бидејќи во неговиот процес бројот на хромозоми е зачуван.

Интерфаза Iпродолжува како интерфаза на митозата. Мејоза Iе поделена на четири фази: профаза I, метафаза I, анафаза I и телофаза I. Б профаза IСе случуваат два важни процеси: конјугација и вкрстување. Конјугација- Ова е процес на фузија на хомологни (спарени) хромозоми по целата должина. Паровите на хромозоми формирани за време на конјугацијата се зачувани до крајот на метафазата I.
Премин преку- меѓусебна размена на хомологни региони на хомологни хромозоми. Како резултат на вкрстување, хромозомите добиени од телото од двајцата родители добиваат нови комбинации на гени, што предизвикува појава на генетски разновидни потомци. На крајот од профазата I, како и во профазата на митоза, јадрото исчезнува, центриолите се разминуваат до половите на клетката и јадрената мембрана се распаѓа.

ВО метафаза Iпарови на хромозоми се порамнети по екваторот на клетката, а микротубулите на вретеното се прикачени на нивните центромери.
ВО анафаза IЦели хомологни хромозоми, составени од два хроматиди, се разминуваат до половите.
ВО телофаза IНуклеарни мембрани се формираат околу кластери на хромозоми на половите на клетката и се формираат јадра.
Цитокинеза Iобезбедува раздвојување на цитоплазмите на ќерките клетки.
Ќерките клетки (1n2c) формирани како резултат на мејозата I се генетски хетерогени, бидејќи нивните хромозоми, случајно дисперзирани на клеточните полови, содржат различни гени.
Компаративни карактеристики на митоза и мејоза
| Потпишете | Митоза | Мејоза | |
| Кои клетки почнуваат да се делат? | Соматски (2n) | Примарни герминативни клетки (2n) | |
| Број на поделби | 1 | 2 | |
| Колку и какви клетки се формираат при делбата? | 2 соматски (2n) | 4 сексуални (n) | |
| Интерфаза | Подготовка на клетката за поделба, ДНК удвојување | Многу кратко, не се случува удвојување на ДНК | |
| Фази | Мејоза I | Мејоза II | |
| Профаза | Може да дојде до кондензација на хромозом, исчезнување на јадрото, распаѓање на нуклеарната мембрана, конјугација и вкрстување | Хромозомска кондензација, исчезнување на јадрото, распаѓање на нуклеарната мембрана | |
| Метафаза | По должината на екваторот се наоѓаат парови на хромозоми, се формира вретено | Хромозомите се редат долж екваторот, се формира вретено | |
| Анафаза | Хомологните хромозоми од два хроматиди се движат кон половите | Хроматидите се движат кон половите | |
| Телофаза | Хромозомите се деспирани, се формираат нови нуклеарни мембрани и јадра | Хромозомите се деспирани, се формираат нови нуклеарни мембрани и јадра | |
Интерфаза IIмногу кратко, бидејќи во него не се случува удвојување на ДНК, односно нема S-период.
Мејоза IIисто така поделени во четири фази: профаза II, метафаза II, анафаза II и телофаза II. ВО профаза IIсе случуваат истите процеси како во профазата I, со исклучок на конјугација и вкрстување.
ВО метафаза IIхромозомите се наоѓаат долж екваторот на клетката.
ВО анафаза IIхромозомите се поделени на центромерите, а хроматидите се протегаат кон половите.
ВО телофаза IIНуклеарни мембрани и јадра се формираат околу кластери на ќерки хромозоми.
По цитокинеза IIГенетската формула на сите четири ќерки клетки е 1n1c, но сите тие имаат различен сет на гени, што е резултат на вкрстување и случајна комбинација на хромозоми на мајчините и татковските организми во клетките ќерки.
Развој на герминативните клетки кај растенијата и животните
Гаметогенеза(од грчки гамета- сопруга, гамети- сопруг и генеза- потекло, појава) е процес на формирање на зрели герминативни клетки.
Бидејќи за сексуалната репродукција најчесто се потребни две индивидуи - женско и машко, кои произведуваат различни полови клетки - јајца и сперматозоиди, тогаш процесите на формирање на овие гамети мора да бидат различни.
Природата на процесот во значителна мера зависи од тоа дали се јавува во растителна или животинска клетка, бидејќи кај растенијата се јавува само митоза при формирање на гамети, а кај животните се јавува и митоза и мејоза.
Развој на герминативните клетки во растенијата.Кај ангиоспермите, формирањето на машки и женски репродуктивни клетки се случува во различни делови на цветот - стомачните и пестиците, соодветно.
Пред формирањето на машките репродуктивни клетки - микрогаметогенеза(од грчки микро- мал) - се случува микроспорогенеза, односно формирање на микроспори во прашниците на стоманите. Овој процес е поврзан со мејотичната поделба на матичната клетка, што резултира со четири хаплоидни микроспори. Микрогаметогенезата е поврзана со митотична поделба на микроспората, давајќи машки гаметофит од две клетки - голем вегетативна(сифоногени) и плитки генеративен. По поделбата, машкиот гаметофит се покрива со густи мембрани и формира поленов зрно. Во некои случаи, дури и за време на процесот на созревање на поленот, а понекогаш и само по пренесувањето на стигмата на пестикот, генеративната клетка се дели митотички за да формира две неподвижни машки герминативни клетки - сперматозоиди. По опрашувањето, од вегетативната клетка се формира поленска цевка, преку која сперматозоидите продираат во јајниците на пестикот за оплодување.

Развојот на женски герминативни клетки во растенијата се нарекува мегагаметогенеза(од грчки мега- големо). Се јавува во јајниците на пестикот, на кој му претходи мегаспорогенеза, како резултат на што се формираат четири мегаспори од матичната клетка на мегаспората што лежи во јадрото преку мејотична делба. Еден од мегаспорите се дели митотички три пати, давајќи му на женскиот гаметофит - ембрионска кеса со осум јадра. Со последователното одвојување на цитоплазмите на ќерките клетки, една од добиените клетки станува јајце, на чии страни лежат таканаречените синергиди, на спротивниот крај на ембрионската кеса се формираат три антиподи, а во центарот , како резултат на фузија на две хаплоидни јадра, се формира диплоидна централна клетка.

Развој на герминативните клетки кај животните.Кај животните, постојат два процеси на формирање на герминативни клетки - сперматогенеза и оогенеза.
Сперматогенеза(од грчки сперматозоиди, сперматозоиди- семе и генеза- потекло, појава) е процес на формирање на зрели машки герминативни клетки - сперматозоиди. Кај луѓето се јавува во тестисите или тестисите и е поделен на четири периоди: репродукција, раст, созревање и формирање.

ВО сезоната на парењеисконските герминативни клетки се делат митотички, што резултира со формирање на диплоиди сперматогонија. ВО период на растсперматогонијата акумулира хранливи материи во цитоплазмата, се зголемува во големина и се претвора во примарни сперматоцити, или Сперматоцити од прв ред. Само после ова тие влегуваат во мејоза ( период на созревање), како резултат на што се формираат првите две секундарен сперматоцит, или Сперматоцити од втор ред, а потоа - четири хаплоидни клетки со уште доста голема количина на цитоплазма - сперматиди. ВО период на формирањетие ја губат речиси целата своја цитоплазма и формираат флагелум, претворајќи се во сперма.
Сперма, или живи, - многу мали подвижни машки репродуктивни клетки со глава, врат и опашка.
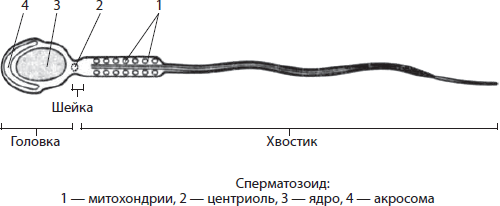
ВО главата, покрај јадрото, е акросома- модифициран Golgi комплекс кој обезбедува растворање на мембраните на јајцето за време на оплодувањето. ВО грлото на маткатасе центриолите на клеточниот центар и основата конска опашкаформираат микротубули кои директно го поддржуваат движењето на спермата. Содржи и митохондрии, кои на сперматозоидите и обезбедуваат АТП енергија за движење.
Огенеза(од грчки ОН- јајце и генеза- потекло, појава) е процес на формирање на зрели женски герминативни клетки - јајце клетки. Кај луѓето се јавува во јајниците и се состои од три периоди: репродукција, раст и созревање. Периодите на репродукција и раст, слични на оние во сперматогенезата, се јавуваат за време на интраутериниот развој. Во овој случај, диплоидните клетки се формираат од примарните герминативни клетки како резултат на митоза. оогонија, кои потоа се претвораат во диплоидна примарна ооцити, или Ооцити од прв ред. Мејозата и последователната цитокинеза се јавуваат во период на созревање, се карактеризираат со нерамномерна поделба на цитоплазмата на матичната клетка, така што како резултат на тоа најпрво се добива секундарен ооцит, или Ооцит од втор ред, И првото поларно тело, а потоа од секундарниот ооцит - јајце клетката, која ја задржува целата залиха на хранливи материи и второто поларно тело, додека првото поларно тело е поделено на две. Поларните тела зафаќаат вишок генетски материјал.
Кај луѓето, јајцата се произведуваат со интервал од 28-29 дена. Циклусот поврзан со созревањето и ослободувањето на јајцата се нарекува менструален.
Јајце- голема женска репродуктивна клетка која носи не само хаплоиден сет на хромозоми, туку и значително снабдување со хранливи материи за последователниот развој на ембрионот.

Јајцето кај цицачите е покриено со четири мембрани, кои ја намалуваат веројатноста за оштетување од различни фактори. Дијаметарот на јајцето кај луѓето достигнува 150-200 микрони, додека кај ној може да биде неколку сантиметри.
Клеточната делба е основа за раст, развој и репродукција на организмите. Улогата на митозата и мејозата
Ако кај едноклеточните организми клеточната делба води до зголемување на бројот на поединци, т.е. репродукција, тогаш кај повеќеклеточните организми овој процес може да има различно значење. Така, поделбата на ембрионските клетки, почнувајќи од зиготот, е биолошка основа на меѓусебно поврзаните процеси на раст и развој. Слични промени се забележани кај луѓето за време на адолесценцијата, кога бројот на клетки не само што се зголемува, туку се јавува и квалитативна промена во телото. Репродукцијата на повеќеклеточните организми се заснова и на клеточната делба, на пример, при бесполово размножување, благодарение на овој процес се обновува цел дел од организмот, а при сексуалната репродукција, во процесот на гаметогенеза, се формираат полови клетки, кои последователно доведуваат до појава на нов организам. Треба да се напомене дека главните методи за поделба на еукариотската клетка - митоза и мејоза - имаат различни значења во животниот циклус на организмите.
Како резултат на митозата, постои рамномерна распределба на наследниот материјал помеѓу ќерките клетки - точни копии на мајката. Без митоза, постоењето и растот на повеќеклеточни организми кои се развиваат од една клетка, зиготот, би било невозможно, бидејќи сите клетки на таквите организми мора да ги содржат истите генетски информации.
Во текот на процесот на делба, ќерките клетки стануваат сè поразновидни во структурата и функциите, што е поврзано со активирање на се повеќе и повеќе нови групи на гени во нив поради меѓуклеточната интеракција. Така, митозата е неопходна за развој на организмот.
Овој метод на клеточна делба е неопходен за процесите на бесполово размножување и регенерација (реставрација) на оштетените ткива, како и органи.
Мејозата, пак, ја обезбедува постојаноста на кариотипот за време на сексуалната репродукција, бидејќи го преполовува збирот на хромозоми пред сексуалната репродукција, што потоа се обновува како резултат на оплодувањето. Покрај тоа, мејозата води до појава на нови комбинации на родителски гени поради вкрстување и случајна комбинација на хромозоми во ќерките клетки. Благодарение на ова, потомството се покажува како генетски разновидно, што обезбедува материјал за природна селекција и е материјална основа за еволуција. Промената на бројот, обликот и големината на хромозомите, од една страна, може да доведе до појава на различни отстапувања во развојот на организмот, па дури и до негова смрт, а од друга страна, може да доведе до појава на поединци. повеќе прилагодени на животната средина.
Така, клетката е единица за раст, развој и репродукција на организмите.
резиме на други презентации„Методи на настава по биологија“ - Училишна зоологија. Запознавање на учениците со употребата на научни зоолошки податоци. Морално образование. Дополнителен благослов на кокошарникот. Избор на методи. Животни процеси. Аквариумска риба. Исхрана. Еколошка едукација. Материјалноста на животните процеси. Негативни резултати. Внимание студенти. Задолжителна форма. Гледајќи мали животни. Цели и задачи на биологијата. Приказна.
„Учење засновано на проблем на часови по биологија“ - Знаење. Нови учебници. Патот до решението. Проблем. Семинари. Што е задача? Албрехт Дирер. Учење базирано на проблем на часовите по биологија. Нестандардни лекции. Што се подразбира под учење засновано на проблем? Квалитетот на животот. Биологијата како академски предмет. Прашање. Лекција за решавање проблеми. Намален интерес за темата. Лабораториски часови засновани на проблем.
„Критичко размислување на часови по биологија“ - Технологија на „критичко размислување“. Користење на технологијата на „развој на критичко размислување“. Табела за лекцијата. Мотивација за учење. Екосистеми. Значењето на „развивање критичко размислување“. Знаци на технологија. RKM технологија. Структура на лекцијата. Главни насоки. Историја на технологијата. Педагошки технологии. Правила на технологијата. Задачи по биологија. Фотосинтеза. Техники кои се користат во различни фази од часот.
„Часови по биологија со интерактивна табла“ - Електронски учебници. Придобивки за студентите. Интерактивната табла помага да се пренесат информации до секој ученик. Дидактички задачи. Решавање на биолошки проблеми. Предности на работа со интерактивни табли. Работа со презентации. Работете на споредување на предмети. Подвижни предмети. Користење на табели. Користење на интерактивна табла во процесот на настава на ученици. Придобивки за наставниците.
„Пристап на системска активност во биологијата“ - Семинарски прашања. Метод на активност. Дриопитекус. Вонземски пат од човечко потекло. Лизозоми. Хемиска организација. Гимносперми. Метаболизам. Анализатори. Пристап со системска активност во наставата по биологија. Хромозоми. Цитоплазма. Слепило. Должина на увото. Класификација на лице. Скелет на цицач. Патеки на човековата еволуција. Митоза. Површен комплекс. Проблематично прашање. Нуклеолус. Нуклеарна обвивка.
„Компјутер по биологија“ - Колаборативна активност на студенти. Семејства на ангиосперми. Интерактивен тренинг. Модели на учење. Пример за систем за оценување. Прашања за наставна картичка. Пример на карта за инструкции. Истражувачите. Микрогрупи. Интерактивни технологии за учење. Рингишпил. Интерактивни технологии за учење. Интерактивни пристапи во часовите по биологија. Групна форма на работа. Задачи за групи „истражувачи“.







