Лизозоми. Митохондриите. Пластиди
1. Која е структурата и функциите АТП?
2. Какви видови пластиди знаете?
Кога разни хранливи материи влегуваат во клетката преку фагоцитоза или пиноцитоза, тие мора да се сварат. При што верверичкимора да се разложи на поединечни амино киселини, полисахариди - до молекули на гликоза или фруктоза, липиди- на глицерол и масни киселини. За да биде возможно интрацелуларното варење, фагоцитната или пиноцитната везикула мора да се спои со лизозомот (сл. 25). Лизозомот е мал меур, со дијаметар од само 0,5-1,0 микрони, кој содржи голем сет на ензими способни да ги уништат хранливите материи. Еден лизозом може да содржи 30-50 различни ензими.

Митохондриите се постојани мембрански органели со тркалезна или прачка (често разгранета) форма. Дебелина - 0,5 микрони, должина - 5-7 микрони. Бројот на митохондриите во повеќето животински клетки е 150-1500; кај женските јајца - до неколку стотици илјади, во сперматозоидите - еден спирален митохондрион извиткан околу аксијалниот дел на флагелумот.
Главните функции на митохондриите:
1) играат улога на енергетски станици на ќелиите;
2) складирање на наследниот материјал во форма на митохондријална ДНК.
Странични функции - учество во синтезата на стероидни хормони, некои амино киселини (на пример, глутамин).
Структурата на митохондриите
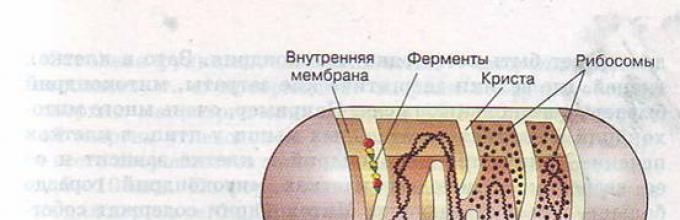
Митохондриите имаат две мембрани: надворешна (мазна) и внатрешна (формирајќи израстоци - во облик на лист (cristae) и тубуларни (тубули)).
Во митохондриите, внатрешната содржина е матрица - колоидна супстанција во која се откриени зрна со дијаметар од 20-30 nm со помош на електронски микроскоп (тие акумулираат јони на калциум и магнезиум, резерви на хранливи материи, на пример, гликоген).
Во матрицата се наоѓа апаратот за биосинтеза на протеините на органелата: 2-6 копии од кружна ДНК без хистонски протеини, рибозоми, збир на t-RNA, ензими за редупликација, транскрипција и превод на наследни информации.
Митохондриите се размножуваат со лигатура; митохондриите се карактеризираат со релативна автономија во клетката.
Лизозомите се везикули со дијаметар од 200-400 μm. (обично). Тие имаат обвивка со една мембрана, која понекогаш е покриена однадвор со влакнест протеински слој. Главната функција е интрацелуларно варење на различни хемиски соединенија и клеточни структури.
Постојат примарни (неактивни) и секундарни лизозоми (процесот на варење се одвива во нив). Секундарните лизозоми се формираат од примарните. Тие се поделени на хетеролизозоми и автолизозоми.
Во хетеролизозомите (или фаголизозомите) процесот на варење на материјалот што влегува во клетката однадвор се одвива преку активен транспорт (пиноцитоза и фагоцитоза).
Во автолизозомите (или цитолизозомите), нивните сопствени клеточни структури кои го завршиле својот живот се уништени.
Секундарните лизозоми кои престанале да варат материјал се нарекуваат резидуални тела. Тие не содржат хидролази и содржат несварен материјал.
Кога интегритетот на лизозомската мембрана е нарушен или кога клетката е заболена, хидролазите влегуваат во клетката од лизозомите и го вршат нејзиното само-варење (автолиза). Истиот процес е во основата на процесот на природна смрт на сите клетки (апоптоза).
Микротела
Микротелата формираат колективна група органели. Тие се везикули со дијаметар од 100-150 nm, ограничени со една мембрана. Тие содржат ситно зрнеста матрица и често протеински подмножества.
Постојани клеточни структури, клеточни органи кои обезбедуваат извршување на специфични функции во текот на животот на клетката - складирање и пренос на генетски информации, пренос на супстанции, синтеза и трансформација на супстанции и енергија, делење, движење итн.
На органоидите (органелите) на клетките еукариотисе однесуваат:
- хромозоми;
- клеточната мембрана;
- митохондриите;
- комплекс Голџи;
- ендоплазматичен ретикулум;
- рибозоми;
- микротубули;
- микрофиламенти;
- лизозоми.
Животинските клетки исто така содржат центриоли и микрофибрили, а растителните клетки содржат пластиди кои се единствени за нив.
Понекогаш јадрото како целина се класифицира како органели на еукариотските клетки.
Прокариотинемаат повеќето органели, тие имаат само клеточна мембрана и рибозоми, кои се разликуваат од цитоплазматските рибозоми на еукариотските клетки.
Специјализираните еукариотски клетки може да имаат сложени структури засновани на универзални органели, како што се микротубули и центриоли - главните компоненти на флагелите и цилиите. Микрофибрилите се во основата на тоно- и неврофибрилите. Специјални структури на едноклеточни организми, како што се флагели и цилии (конструирани на ист начин како и во повеќеклеточните клетки), ја вршат функцијата на органите за движење.
Почесто во современата литература термините „ органоиди "И" органели “ се користат како синоними.
Структури заеднички за животинските и растителните клетки
| Шематска илустрација | Структура | Функции |
|
| Плазма мембрана (плазмалема, клеточна мембрана) | Два слоја липиди (двослоен) помеѓу два слоја протеин | Селективно пропустлива бариера која ја регулира размената помеѓу клетката и околината |
|
| Јадро | Најголемата органела, затворена во обвивка од две мембрани, проникна нуклеарни пори. Содржи хроматин- во оваа форма, ненамотани хромозоми се во интерфаза. Исто така, содржи структура наречена јадро | Хромозомите содржат ДНК - супстанција на наследноста.ДНК се состои од гени кои ги регулираат сите видови на клеточна активност. Нуклеарната поделба е во основата на репродукцијата на клетките, а со тоа и на процесот на репродукција. Рибозомите се формираат во јадрото |
|
| Ендоплазматичен ретикулум (ЕР) | Систем на сплескани мембрански кеси - тенкови- во форма на цевки и плочи. Формира единствена единица со надворешната мембрана на нуклеарната обвивка | Ако површината на ЕР е покриена со рибозоми, тогаш се нарекува грубо.Протеинот синтетизиран на рибозомите се транспортира низ резервоарите на таков ЕР. Мазна ЕР(без рибозоми) служи како место за синтеза на липиди и стероиди |
|
| Рибозоми | Многу мали органели кои се состојат од две подчестички - големи и мали. Тие содржат протеини и РНК во приближно еднакви пропорции. Рибозомите, кои се наоѓаат во митохондриите (а исто така и во хлоропластите во растенијата), се уште помали | Местото на синтеза на протеини, каде што различни молекули во интеракција се држат во правилна положба. Рибозомите се поврзани со ЕР или лежат слободни во цитоплазмата. Може да се формираат многу рибозоми полисом (полирибозом), во која тие се нанижани на една нишка на гласник РНК |
|
| Митохондриите | Митохондрионот е опкружен со обвивка од две мембрани, внатрешната мембрана формира набори ( cristas). Содржи матрица која содржи мал број рибозоми, една кружна молекула на ДНК и фосфатни гранули | За време на аеробното дишење, оксидативната фосфорилација и трансферот на електрони се случуваат во кристаите, а ензимите вклучени во циклусот на Кребс и оксидацијата на масните киселини работат во матрицата. |
|
| Голџи апарат | Куп од сплескани мембрански кеси - тенкови. На едниот крај континуирано се формираат купишта торбички, а на другиот се врзуваат во форма на меурчиња. Купиштата може да постојат како дискретни диктјозоми, како во растителните клетки или да формираат просторна мрежа, како кај многу животински клетки | Многу клеточни материјали, како што се ензимите од ЕР, подлежат на модификација во цистерните и се транспортираат во везикули. Апаратот Голџи е вклучен во процесот на секреција, а во него се формираат лизозоми |
|
| Лизозоми | Едноставна сферична мембранска кеса (единечна мембрана) исполнета со дигестивни (хидролитички) ензими. Содржината изгледа хомогена | Врши многу функции, секогаш поврзани со распаѓање на какви било структури или молекули |
|
| Микротела | Органелата не е сосема правилна сферична форма, опкружена со една мембрана. Содржината има грануларна структура, но понекогаш има кристалоиди или збирка нишки во неа | Сите микротела содржат каталаза, ензим кој го катализира разградувањето на водород пероксид. Сите тие се поврзани со оксидативни реакции |
|
| Клеточен ѕид, средна ламина, плазмодезмати |
|||
| клеточен ѕид | Цврстиот клеточен ѕид што ја опкружува клетката се состои од целулозни микрофибрили вградени во матрица која содржи други сложени полисахариди, имено хемицелулози и пектини. Во некои клетки, клеточните ѕидови се подложени на секундарно задебелување | Обезбедува механичка поддршка и заштита. Благодарение на него, се јавува тургорски притисок, што ја подобрува функцијата за поддршка. Спречува руптура на осмотските клетки. Движењето на водата и минералните соли се случува долж клеточниот ѕид. Различни модификации, како што е импрегнацијата на лигнин, обезбедуваат специјализирани функции |
|
| средна плоча | Тенок слој на пектински материи (калциум и магнезиум пектити) | Ги држи клетките заедно |
|
| плазмодезма | Тенка цитоплазматска нишка која ја поврзува цитоплазмата на две соседни клетки преку тенка пора во клеточниот ѕид. Пората е обложена со плазма мембрана Дезмотубулата поминува низ пората, често поврзана на двата краја со ЕР | Обединете ги протопластите на соседните клетки во единствен континуиран систем - симпласт, преку кој супстанциите се транспортираат помеѓу овие клетки |
|
| Хлоропласт | Голем пластид што содржи хлорофил во кој се јавува фотосинтеза. Хлоропластот е опкружен со двојна мембрана и исполнет со желатинозна строма. Стромата содржи систем на мембрани склопени во стекови, или зрна.Во него може да се депонира и скроб. Покрај тоа, стромата содржи рибозоми, кружна молекула на ДНК и капки масло | Во оваа органела се јавува фотосинтеза, односно синтеза на шеќери и други материи од CO 2 и вода поради светлосната енергија заробена од хлорофилот.Светлосната енергија се претвора во хемиска енергија |
|
| Голема централна вакуола | Кесичка формирана од една мембрана наречена тонопласт. Вакуолата содржи клеточен сок - концентриран раствор на различни материи, како што се минерални соли, шеќери, пигменти, органски киселини и ензими. Во зрелите клетки, вакуолите се обично големи | Овде се складираат различни супстанции, вклучувајќи ги и крајните производи на метаболизмот. Осмотските својства на клетката силно зависат од содржината на вакуолата. Понекогаш вакуолата функционира како лизозом |
|
Компаративни карактеристики на РНК и ДНК
| Знаци | РНК | ДНК |
| Локација во кафезот | Јадро, рибозоми, цитоплазма, митохондрии, хлоропласти | Јадро, митохондрии, хлоропласти |
| Локација во јадрото | Нуклеолус | Хромозоми |
| Структура на макромолекула | Единечен полинуклеотиден ланец | Двоен неразгранет линеарен полимер, навиен во десна спирала |
| Мономери | Рибонуклеотиди | Деоксирибонуклеотиди |
| Нуклеотиден состав | Азотна база (пурин - аденин, гванин, пиримидин - урацил, цитозин); рибоза (јаглехидрати): остаток од фосфорна киселина | Азотна база (пурин - аденин, гванин, пиримидин - тимин, цитозин); деоксирибоза (јаглехидрати): остаток од фосфорна киселина |
| Видови нуклеотиди | Аленил (A), гванил (G), уридил (U), цитидил (C) | Аленил (А), гуанил (G), тимидил (Т), цитидил (C) |
| Својства | Неспособни за само-удвојување. Лабилна | Способни за самодуплирање според принципот на комплементарност (редупликација): A-T, T-A, G-C, C-G Стабилна |
| Функции | Информативен (mRNA) - го пренесува кодот на наследни информации за примарната структура на протеинската молекула; рибозомални (rRNA) - дел од рибозомите; транспорт (тРНК) - пренесува амино киселини во рибозомите; митохондријална и пластидна РНК - се дел од рибозомите на овие органели | Хемиска основа на хромозомски генетски материјал (ген); Синтеза на ДНК, синтеза на РНК, информации за структурата на протеините |
Митохондриите се микроскопски мембрански врзани органели кои обезбедуваат енергија на клетката. Затоа, тие се нарекуваат енергетски станици (батерија) на ќелии.
Митохондриите се отсутни во клетките на едноставни организми, бактерии и ентамеба, кои живеат без употреба на кислород. Некои зелени алги, трипанозоми содржат еден голем митохондрион, а клетките на срцевиот мускул и мозокот имаат од 100 до 1000 од овие органели.
Структурни карактеристики
Митохондриите се органели со двојна мембрана; тие имаат надворешна и внатрешна мембрана, меѓумембрански простор меѓу нив и матрица.
Надворешна мембрана. Мазна е, нема набори и ја одвојува внатрешната содржина од цитоплазмата. Неговата ширина е 7 nm и содржи липиди и протеини. Важна улога игра поринот, протеин кој формира канали во надворешната мембрана. Тие обезбедуваат јонска и молекуларна размена.
Меѓумембрански простор. Големината на меѓумембранскиот простор е околу 20 nm. Супстанцијата што ја пополнува е слична во составот на цитоплазмата, со исклучок на големите молекули кои можат да навлезат овде само преку активен транспорт.

Внатрешна мембрана. Таа е изградена главно од протеини, само една третина е наменета за липидни супстанции. Голем број на протеини се транспортни протеини, бидејќи на внатрешната мембрана и недостасуваат слободно проодни пори. Формира многу израстоци - cristae, кои изгледаат како срамнети со земја сртови. Оксидацијата на органските соединенија до CO 2 во митохондриите се случува на мембраните на кристаите. Овој процес е зависен од кислород и се изведува под дејство на АТП синтетаза. Ослободената енергија се складира во форма на молекули на АТП и се користи по потреба.
Матрица– внатрешната средина на митохондриите има грануларна, хомогена структура. Во електронски микроскоп, можете да видите гранули и филаменти во топчиња кои слободно лежат помеѓу кристаите. Матрицата содржи полуавтономен систем за синтеза на протеини - ДНК, сите видови РНК, а тука се наоѓаат рибозомите. Но, сепак, повеќето од протеините се снабдуваат од јадрото, поради што митохондриите се нарекуваат полуавтономни органели.
Локација и поделба на клетките
Хондриоме група на митохондрии кои се концентрирани во една клетка. Тие се наоѓаат различно во цитоплазмата, што зависи од специјализацијата на клетките. Поставувањето во цитоплазмата зависи и од околните органели и подмножества. Во растителните клетки тие ја окупираат периферијата, бидејќи митохондриите се туркаат кон мембраната од централната вакуола. Во бубрежните епителни клетки, мембраната формира испакнатини, меѓу кои има митохондрии.
Во матичните клетки, каде што енергијата се користи подеднакво од сите органели, митохондриите се хаотично распоредени. Во специјализираните ќелии, тие се главно концентрирани во области со најголема потрошувачка на енергија. На пример, во напречно-пругастите мускули тие се наоѓаат во близина на миофибрилите. Кај сперматозоидите, тие спирално ја покриваат оската на флагелумот, бидејќи е потребна многу енергија за да се стави во движење и да се движи спермата. Протозоите кои се движат со помош на цилии, исто така, содржат голем број на митохондрии во нивната база.
Поделба. Митохондриите се способни за независна репродукција, имаат свој геном. Органелите се поделени со констрикции или прегради. Формирањето на нови митохондрии во различни клетки се разликува по фреквенција; на пример, во ткивото на црниот дроб тие се заменуваат на секои 10 дена.
Функции во ќелијата
- Главната функција на митохондриите е формирање на молекули на АТП.
- Таложење на јони на калциум.
- Учество во размена на вода.
- Синтеза на прекурсори на стероидни хормони.
Молекуларната биологија е наука која ја проучува улогата на митохондриите во метаболизмот. Тие исто така го претвораат пируватот во ацетил-коензим А и бета-оксидација на масните киселини.
| Табела: структура и функции на митохондриите (накратко) | ||
|---|---|---|
| Структурни елементи | Структура | Функции |
| Надворешна мембрана | Мазна обвивка, направена од липиди и протеини | Ја одвојува внатрешната содржина од цитоплазмата |
| Меѓумембрански простор | Постојат водородни јони, протеини, микромолекули | Создава протонски градиент |
| Внатрешна мембрана | Формира испакнатини - cristae, содржи системи за транспорт на протеини | Трансфер на макромолекули, одржување на градиент на протон |
| Матрица | Локација на ензимите на Кребсовиот циклус, ДНК, РНК, рибозоми | Аеробна оксидација со ослободување на енергија, конверзија на пируват во ацетил коензим А. |
| Рибозоми | Комбинирани две подединици | Синтеза на протеини |
Сличности помеѓу митохондриите и хлоропластите
Заедничките својства на митохондриите и хлоропластите првенствено се должат на присуството на двојна мембрана.
Знаците на сличност ја вклучуваат и способноста за независно синтетизирање на протеини. Овие органели имаат своја ДНК, РНК и рибозоми.
И митохондриите и хлоропластите можат да се делат со стегање.
Тие се обединети и со способноста да произведуваат енергија; митохондриите се поспецијализирани за оваа функција, но хлоропластите произведуваат и АТП молекули за време на фотосинтетичките процеси. Така, растителните клетки имаат помалку митохондрии од животинските клетки, бидејќи хлоропластите делумно ги извршуваат функциите за нив.
Ајде накратко да ги опишеме сличностите и разликите:
- Тие се двомембрански органели;
- внатрешната мембрана формира испакнатини: кристаите се карактеристични за митохондриите, а тилакоидите се карактеристични за хлоропластите;
- имаат свој геном;
- способни да синтетизираат протеини и енергија.
Овие органели се разликуваат во нивните функции: митохондриите се наменети за синтеза на енергија, овде се јавува клеточно дишење, хлоропластите им се потребни на растителните клетки за фотосинтеза.
Содржината на статијата
КЛЕТКА,елементарна единица на живи суштества. Клетката е ограничена од другите клетки или од надворешната средина со посебна мембрана и има јадро или негов еквивалент, во кој е концентриран најголемиот дел од хемиските информации што ја контролираат наследноста. Цитологијата ја проучува структурата на клетките, а физиологијата се занимава со нивното функционирање. Науката која го проучува ткивото составено од клетки се нарекува хистологија.
Постојат едноклеточни организми чие цело тело се состои од една клетка. Оваа група вклучува бактерии и протисти (протозои и едноклеточни алги). Понекогаш тие се нарекуваат и безклеточни, но терминот едноклеточни се користи почесто. Вистинските повеќеклеточни животни (Metazoa) и растенијата (Metaphyta) содржат многу клетки.
Огромното мнозинство на ткива се составени од клетки, но има некои исклучоци. Телото на калапи од лигите (миксомицети), на пример, се состои од хомогена супстанција која не е поделена на клетки со бројни јадра. Некои животински ткива, особено срцевиот мускул, се организирани на сличен начин. Вегетативното тело (талус) на габите е формирано со микроскопски нишки - хифи, често сегментирани; секоја таква нишка може да се смета за еквивалент на клетка, иако со атипична форма.
Некои структури на телото кои не учествуваат во метаболизмот, особено лушпите, бисерите или минералната основа на коските, не се формираат од клетките, туку од производите на нивното лачење. Други, како што се дрвото, кората, роговите, косата и надворешниот слој на кожата, не се од секреторно потекло, туку се формираат од мртви клетки.
Малите организми, како што се ротиферите, се состојат од само неколку стотици клетки. За споредба: во човечкото тело има прибл. 10 14 клетки, секоја секунда 3 милиони црвени крвни зрнца умираат и се заменуваат со нови, а тоа е само една десетина милионити дел од вкупниот број на телесните клетки.
Вообичаено, големината на растителните и животинските клетки се движи од 5 до 20 микрони во дијаметар. Типична бактериска клетка е многу помала - прибл. 2 микрони, а најмалиот познат е 0,2 микрони.
Некои слободни живи клетки, како што се протозоите како што се фораминиферите, можат да достигнат неколку сантиметри; тие секогаш имаат многу јадра. Клетките на тенки растителни влакна достигнуваат должина од еден метар, а процесите на нервните клетки достигнуваат неколку метри кај големите животни. Со таква должина, волуменот на овие ќелии е мал, но површината е многу голема.
Најголемите клетки се неоплодени птичји јајца исполнети со жолчка. Најголемото јајце (и, според тоа, најголемата клетка) припаѓаше на изумрена огромна птица - apyornis ( Aepyornis). Веројатно неговата жолчка тежела прибл. 3,5 кг. Најголемото јајце меѓу живите видови му припаѓа на нојот, неговата жолчка тежи приближно. 0,5 кг.
Како по правило, клетките на големите животни и растенија се само малку поголеми од клетките на малите организми. Слонот е поголем од глушец не затоа што неговите ќелии се поголеми, туку главно затоа што самите има многу повеќе клетки. Постојат групи на животни, како што се ротифери и нематоди, во кои бројот на клетки во телото останува константен. Така, иако големите видови нематоди имаат поголем број клетки од малите, главната разлика во големината во овој случај се должи на поголемите големини на клетките.
Во рамките на даден тип на клетка, нивните големини обично зависат од плоидијата, т.е. за бројот на множества на хромозоми присутни во јадрото. Тетраплоидните клетки (со четири групи на хромозоми) се двојно поголеми по волумен од диплоидните клетки (со две групи хромозоми). Плоидноста на растението може да се зголеми со воведување на хербалниот лек колхицин во него. Бидејќи растенијата изложени на овој ефект имаат поголеми клетки, тие самите се поголеми. Сепак, овој феномен може да се забележи само кај полиплоидите од неодамнешно потекло. Во еволутивно античките полиплоидни растенија, големината на клетките е предмет на „обратна регулација“ кон нормалните вредности и покрај зголемувањето на бројот на хромозоми.
СТРУКТУРА НА КЛЕТКАТА
Едно време, клетката се сметаше за повеќе или помалку хомогена капка органска материја, која се нарекуваше протоплазма или жива супстанција. Овој термин стана застарен откако беше откриено дека клетката се состои од многу јасно различни структури наречени клеточни органели („мали органи“).
Хемиски состав.
Вообичаено, 70-80% од клеточната маса е вода, во која се раствораат различни соли и органски соединенија со мала молекуларна тежина. Најкарактеристичните компоненти на клетката се протеините и нуклеинските киселини. Некои протеини се структурни компоненти на клетката, други се ензими, т.е. катализатори кои ја одредуваат брзината и насоката на хемиските реакции што се случуваат во клетките. Нуклеинските киселини служат како носители на наследни информации, кои се реализираат во процесот на интрацелуларна синтеза на протеини.
Честопати клетките содржат одредена количина на материи за складирање кои служат како резерва за храна. Растителните клетки првенствено складираат скроб, полимерна форма на јаглени хидрати. Друг полимер на јаглени хидрати, гликоген, се складира во клетките на црниот дроб и мускулите. Често складираната храна вклучува и масти, иако некои масти извршуваат различна функција, имено, тие служат како суштински структурни компоненти. Протеините во клетките (со исклучок на семенските клетки) обично не се складираат.
Не е возможно да се опише типичниот состав на клетката, првенствено затоа што има големи разлики во количината на складирана храна и вода. Клетките на црниот дроб содржат, на пример, 70% вода, 17% протеини, 5% масти, 2% јаглени хидрати и 0,1% нуклеински киселини; останатите 6% доаѓаат од соли и органски соединенија со ниска молекуларна тежина, особено амино киселини. Растителните клетки обично содржат помалку протеини, значително повеќе јаглехидрати и нешто повеќе вода; исклучок се клетките кои се во состојба на мирување. Клетката за одмор на зрно пченица, која е извор на хранливи материи за ембрионот, содржи прибл. 12% протеини (најчесто складирани протеини), 2% масти и 72% јаглени хидрати. Количината на вода достигнува нормално ниво (70–80%) само на почетокот на ртење на зрната.
Главните делови на клетката.
Некои клетки, претежно растителни и бактериски, имаат надворешен клеточен ѕид. Во вишите растенија се состои од целулоза. Ѕидот ја опкружува самата клетка, заштитувајќи ја од механички стрес. Клетките, особено бактериските клетки, исто така можат да лачат мукозни материи, при што околу себе формираат капсула, која, како и клеточниот ѕид, има заштитна функција.
Токму со уништувањето на клеточните ѕидови е поврзана смртта на многу бактерии под влијание на пеницилин. Факт е дека во внатрешноста на бактериската клетка концентрацијата на соли и нискомолекуларни соединенија е многу висока, и затоа, во отсуство на зајакнувачки ѕид, приливот на вода во клетката предизвикан од осмотскиот притисок може да доведе до нејзино кинење. Пеницилинот, кој го спречува формирањето на неговиот ѕид за време на растот на клетките, доведува до кинење на клетките (лиза).
Клеточните ѕидови и капсулите не учествуваат во метаболизмот и често може да се одвојат без да се убие клетката. Така, тие може да се сметаат за надворешни помошни делови на ќелијата. На животинските клетки обично им недостасуваат клеточни ѕидови и капсули.
Самата клетка се состои од три главни дела. Под клеточниот ѕид, доколку е присутна, се наоѓа клеточната мембрана. Мембраната опкружува хетероген материјал наречен цитоплазма. Тркалезно или овално јадро е потопено во цитоплазмата. Подолу ќе ја разгледаме подетално структурата и функциите на овие делови од клетката.
КЛЕТОЧНАТА МЕМБРАНА
Клеточната мембрана е многу важен дел од клетката. Ги држи сите клеточни компоненти заедно и ги разграничува внатрешните и надворешните средини. Покрај тоа, модифицираните набори на клеточната мембрана формираат многу органели на клетката.
Клеточната мембрана е двоен слој на молекули (бимолекуларен слој или двослој). Тоа се главно молекули на фосфолипиди и други супстанции поврзани со нив. Молекулите на липидите имаат двојна природа, што се манифестира во тоа како тие се однесуваат во однос на водата. Главите на молекулите се хидрофилни, т.е. имаат афинитет за вода, а нивните јаглеводородни опашки се хидрофобни. Затоа, кога се мешаат со вода, липидите формираат филм на неговата површина сличен на филм со масло; Покрај тоа, сите нивни молекули се ориентирани на ист начин: главите на молекулите се во водата, а јаглеводородните опашки се над нејзината површина.
Во клеточната мембрана има два такви слоја и во секој од нив главите на молекулите се свртени кон надвор, а опашките се свртени во внатрешноста на мембраната, еден кон друг, со што не доаѓаат во контакт со вода. Дебелината на таквата мембрана е прибл. 7 nm. Покрај главните липидни компоненти, содржи големи протеински молекули кои можат да „пловат“ во липидниот двослој и се наредени така што едната страна е свртена кон внатрешноста на клетката, а другата е во контакт со надворешната средина. Некои протеини се наоѓаат само на надворешната или само на внатрешната површина на мембраната или се само делумно потопени во липидниот двослој.
Главната функција на клеточната мембрана е да го регулира транспортот на супстанции во и надвор од клетката. Бидејќи мембраната физички е нешто слична на маслото, супстанциите што се растворливи во масло или органски растворувачи, како што е етер, лесно минуваат низ неа. Истото важи и за гасовите како што се кислородот и јаглерод диоксидот. Во исто време, мембраната е практично непропустлива за повеќето супстанции растворливи во вода, особено шеќери и соли. Благодарение на овие својства, тој е во состојба да одржува хемиска средина во клетката која се разликува од надвор. На пример, во крвта концентрацијата на натриумовите јони е висока, а јоните на калиум се ниски, додека во интрацелуларната течност овие јони се присутни во спротивен однос. Слична ситуација е типична за многу други хемиски соединенија.
Очигледно е дека клетката, сепак, не може целосно да се изолира од околината, бидејќи мора да ги прими супстанциите неопходни за метаболизмот и да се ослободи од нејзините финални производи. Покрај тоа, липидниот двослој не е целосно непропустлив дури и за супстанции растворливи во вода, и т.н. Протеините кои формираат канали создаваат пори или канали кои можат да се отвораат и затвораат (во зависност од промените во конформацијата на протеините) и, во отворена состојба, да спроведат одредени јони (Na +, K +, Ca 2+) долж градиентот на концентрацијата . Следствено, разликата во концентрациите внатре и надвор од клетката не може да се одржува само поради малата пропустливост на мембраната. Всушност, содржи протеини кои вршат функција на молекуларна „пумпа“: тие транспортираат одредени супстанции и во и надвор од клетката, работејќи против градиентот на концентрацијата. Како резултат на тоа, кога концентрацијата на, на пример, аминокиселините внатре во клетката е висока и ниска надвор, аминокиселините сепак можат да течат од надворешната средина во внатрешната. Овој трансфер се нарекува активен транспорт и користи енергија обезбедена од метаболизмот. Мембранските пумпи се многу специфични: секоја од нив е способна да транспортира или само јони на одреден метал, или амино киселина или шеќер. Специфични се и мембранските јонски канали.
Таквата селективна пропустливост е физиолошки многу важна, а нејзиното отсуство е првиот доказ за клеточна смрт. Ова е лесно да се илустрира со примерот на цвекло. Ако жив корен од цвекло е потопен во ладна вода, тој го задржува својот пигмент; ако цвеклото се варат, клетките умираат, стануваат лесно пропустливи и го губат пигментот, со што водата станува црвена.
Клетката може да „голта“ големи молекули како што се протеините. Под влијание на одредени протеини, доколку се присутни во течноста што ја опкружува клетката, се случува инвагинација во клеточната мембрана, која потоа се затвора, формирајќи везикула - мала вакуола која содржи молекули на вода и протеини; По ова, мембраната околу вакуолата пука, а содржината влегува во клетката. Овој процес се нарекува пиноцитоза (буквално „пиење на клетката“) или ендоцитоза.
Поголемите честички, како што се честичките од храната, можат да се апсорбираат на сличен начин за време на т.н. фагоцитоза. Типично, вакуолата формирана за време на фагоцитозата е поголема, а храната се вари со лизозомални ензими во внатрешноста на вакуолата пред да пукне околната мембрана. Овој тип на исхрана е типичен за протозои, како што се амебите, кои јадат бактерии. Сепак, способноста за фагоцитоза е карактеристична и за цревните клетки на долните животни и за фагоцитите, еден од типовите на бели крвни зрнца (леукоцити) на 'рбетниците. Во вториот случај, значењето на овој процес не е во исхраната на самите фагоцити, туку во нивното уништување на бактерии, вируси и други странски материјали штетни за телото.
Функциите на вакуолите може да бидат различни. На пример, протозоите кои живеат во свежа вода доживуваат постојан осмотски прилив на вода, бидејќи концентрацијата на соли внатре во клетката е многу поголема отколку надвор. Тие се способни да лачат вода во посебна екскреторна (контрактилна) вакуола, која периодично ја турка нејзината содржина надвор.
Растителните клетки често имаат една голема централна вакуола која ја зафаќа речиси целата клетка; цитоплазмата формира само многу тенок слој помеѓу клеточниот ѕид и вакуолата. Една од функциите на таквата вакуола е акумулација на вода, овозможувајќи и на клетката брзо да се зголеми во големина. Оваа способност е особено неопходна во периодот кога растителните ткива растат и формираат влакнести структури.
Во ткивата, на места каде што клетките се цврсто поврзани, нивните мембрани содржат бројни пори формирани од протеини кои продираат во мембраната - т.н. конектори. Порите на соседните клетки се лоцирани една спроти друга, така што нискомолекуларните супстанции можат да минуваат од клетка до клетка - овој хемиски комуникациски систем ја координира нивната витална активност. Еден пример за таква координација е повеќе или помалку синхроната поделба на соседните клетки забележана во многу ткива.
ЦИТОПЛАЗАМ
Цитоплазмата содржи внатрешни мембрани кои се слични на надворешната мембрана и формираат органели од различни видови. Овие мембрани може да се сметаат како набори на надворешната мембрана; понекогаш внатрешните мембрани се интегрални со надворешната, но честопати внатрешниот набор е одврзан и контактот со надворешната мембрана е прекинат. Сепак, дури и ако се одржува контакт, внатрешната и надворешната мембрана не се секогаш хемиски идентични. Особено, составот на мембранските протеини се разликува во различни клеточни органели.
Ендоплазматичен ретикулум.
Мрежа на внатрешни мембрани која се состои од тубули и везикули се протега од површината на клетката до јадрото. Оваа мрежа се нарекува ендоплазматичен ретикулум. Често е забележано дека тубулите се отвораат на површината на клетката, а ендоплазматскиот ретикулум на тој начин ја игра улогата на микроциркулаторниот апарат преку кој надворешната средина може директно да комуницира со целата содржина на клетката. Оваа интеракција е пронајдена во некои клетки, особено во мускулните клетки, но сè уште не е јасно дали е универзална. Во секој случај, всушност се случува транспорт на голем број супстанции низ овие тубули од еден дел на клетката до друг.
Ситни тела наречени рибозоми ја покриваат површината на ендоплазматскиот ретикулум, особено во близина на јадрото. Дијаметар на рибозомот прибл. 15 nm, тие се состојат од половина протеини, половина рибонуклеински киселини. Нивната главна функција е синтеза на протеини; гласник РНК и амино киселини поврзани со трансфер РНК се прикачени на нивната површина. Областите на ретикулумот покриени со рибозоми се нарекуваат груб ендоплазматичен ретикулум, а оние без нив се нарекуваат мазни. Покрај рибозомите, различни ензими се адсорбираат на ендоплазматскиот ретикулум или на друг начин се прикачени на него, вклучувајќи ги и ензимските системи кои обезбедуваат употреба на кислород за формирање на стероли и за неутрализација на одредени отрови. Под неповолни услови, ендоплазматскиот ретикулум брзо се дегенерира, и затоа неговата состојба служи како чувствителен индикатор за здравјето на клетките.
Голџи апарат.
Апаратот Голџи (комплекс Голџи) е специјализиран дел од ендоплазматскиот ретикулум, кој се состои од наредени рамни мембрански кеси. Тој е вклучен во лачењето на протеините од клетката (во него се случува пакување на секретираните протеини во гранули) и затоа е особено развиен во клетките кои вршат секреторна функција. Важните функции на апаратот Голџи, исто така, вклучуваат прицврстување на јаглехидратни групи на протеини и употреба на овие протеини за изградба на клеточната мембрана и лизозомската мембрана. Кај некои алги, целулозните влакна се синтетизираат во апаратот Голџи.
Лизозоми
- Тоа се мали меурчиња опкружени со една мембрана. Тие пукаат од апаратот Голџи и можеби од ендоплазматскиот ретикулум. Лизозомите содржат различни ензими кои разградуваат големи молекули, особено протеини. Поради нивното деструктивно дејство, овие ензими се, како да се, „заклучени“ во лизозомите и се ослободуваат само кога е потребно. Така, за време на интрацелуларното варење, ензимите се ослободуваат од лизозомите во дигестивните вакуоли. Лизозомите се исто така неопходни за уништување на клетките; на пример, за време на трансформацијата на полноглавецот во возрасна жаба, ослободувањето на лизозомалните ензими обезбедува уништување на клетките на опашката. Во овој случај, ова е нормално и корисно за телото, но понекогаш таквото уништување на клетките е патолошко. На пример, кога се вдишува азбестната прашина, таа може да навлезе во клетките на белите дробови, а потоа пукаат лизозомите, се развиваат уништување на клетките и пулмонална болест.
Митохондриите и хлоропластите.
Митохондриите се релативно големи структури слични на вреќички со прилично сложена структура. Тие се состојат од матрица опкружена со внатрешна мембрана, меѓумембрански простор и надворешна мембрана. Внатрешната мембрана е превиткана во набори наречени cristae. Кластери на протеини се наоѓаат на кристаите. Многу од нив се ензими кои ја катализираат оксидацијата на производите за распаѓање на јаглени хидрати; други катализираат реакции на синтеза и оксидација на масти. Помошните ензими вклучени во овие процеси се раствораат во митохондријалната матрица.
Оксидацијата на органските материи се јавува во митохондриите, заедно со синтезата на аденозин трифосфат (ATP). Распаѓањето на АТП за да се формира аденозин дифосфат (ADP) е придружено со ослободување на енергија, која се троши на различни витални процеси, на пример, за синтеза на протеини и нуклеински киселини, транспорт на супстанции во и надвор од клетката, пренос на нервните импулси или мускулната контракција. Така, митохондриите се енергетски станици кои го обработуваат „горивото“ - мастите и јаглехидратите - во форма на енергија што може да ја користи клетката, а со тоа и телото како целина.
Растителните клетки исто така содржат митохондрии, но главниот извор на енергија за нивните клетки е светлината. Светлината енергија ја користат овие клетки за производство на АТП и синтетизирање на јаглехидрати од јаглерод диоксид и вода. Хлорофилот, пигмент кој акумулира светлосна енергија, се наоѓа во хлоропластите. Хлоропластите, како и митохондриите, имаат внатрешни и надворешни мембрани. Од израстоците на внатрешната мембрана за време на развојот на хлоропластите, произлегуваат таканаречените хлоропласти. тилакоидни мембрани; вторите формираат срамнети со земја вреќи, собрани во купишта како колона од монети; овие купишта, наречени грана, содржат хлорофил. Покрај хлорофилот, хлоропластите ги содржат сите други компоненти неопходни за фотосинтеза.
Некои специјализирани хлоропласти не вршат фотосинтеза, но имаат други функции, како што се складирање на скроб или пигменти.
Релативна автономија.
Во некои аспекти, митохондриите и хлоропластите се однесуваат како автономни организми. На пример, исто како и самите клетки, кои произлегуваат само од клетки, митохондриите и хлоропластите се формираат само од претходно постоечки митохондрии и хлоропласти. Ова беше докажано во експериментите на растителни клетки, во кои формирањето на хлоропласти беше потиснато од антибиотикот стрептомицин, и на клетките на квасецот, каде што формирањето на митохондриите беше потиснато со други лекови. По ваквите ефекти, клетките никогаш не ги вратија органелите што недостасуваа. Причината е што митохондриите и хлоропластите содржат одредена количина на сопствен генетски материјал (ДНК) кој кодира дел од нивната структура. Ако оваа ДНК се изгуби, што се случува кога формирањето на органели е потиснато, тогаш структурата не може да се рекреира. Двата типа на органели имаат свој систем за синтетизирање на протеини (рибозоми и трансферни РНК), кој е нешто различен од главниот систем за синтеза на протеини на клетката; познато е, на пример, дека системот за синтеза на протеини на органели може да се потисне со помош на антибиотици, додека тие немаат ефект врз главниот систем.
ДНК на органела е одговорна за најголемиот дел од екстрахромозомското, или цитоплазматското наследување. Екстрахромозомската наследност не ги почитува менделовите закони, бидејќи кога клетката се дели, ДНК од органели се пренесува до ќерките клетки на различен начин од хромозомите. Студијата за мутации кои се јавуваат во ДНК на органелата и во хромозомската ДНК покажа дека ДНК на органелата е одговорна само за мал дел од структурата на органелите; повеќето од нивните протеини се кодирани во гените лоцирани на хромозомите.
Делумната генетска автономија на органелите што се разгледуваат и карактеристиките на нивните системи за синтеза на протеини послужија како основа за претпоставката дека митохондриите и хлоропластите потекнуваат од симбиотски бактерии кои се населиле во клетките пред 1-2 милијарди години. Модерен пример за таква симбиоза се малите фотосинтетички алги кои живеат во клетките на некои корали и мекотели. Алгите обезбедуваат кислород на своите домаќини и добиваат хранливи материи од нив.
Фибриларни структури.
Цитоплазмата на клетката е вискозна течност, така што површинскиот напон би предизвикал да се очекува клетката да биде сферична, освен ако клетките не се цврсто спакувани. Сепак, ова обично не се забележува. Многу протозои имаат густи облоги или мембрани кои и даваат на клетката специфична, не-сферична форма. Сепак, дури и без мембрана, клетките можат да одржат несферична форма поради фактот што цитоплазмата е структурирана со помош на бројни, прилично крути, паралелни влакна. Вторите се формираат од шупливи микротубули, кои се состојат од протеински единици организирани во спирала.
Некои протозои формираат псевдоподија - долги, тенки цитоплазматски проекции со кои ја фаќаат храната. Псевдоподиите ја задржуваат својата форма поради ригидноста на микротубулите. Ако хидростатичкиот притисок се зголеми на приближно 100 атмосфери, микротубулите се распаѓаат и клетката добива облик на капка. Кога притисокот се враќа во нормала, микротубулите повторно се собираат и клетката формира псевдоподија. Многу други клетки реагираат на сличен начин на промените во притисокот, што го потврдува учеството на микротубулите во одржувањето на клеточната форма. Склопувањето и распаѓањето на микротубулите, неопходни за брзо менување на обликот на клетката, се случува дури и во отсуство на промени во притисокот.
Микротубулите формираат и фибриларни структури кои служат како органи за движење на клетките. Некои клетки имаат проекции слични на камшици наречени флагели или цилии - нивното тепање обезбедува движење на клетката во вода. Ако клетката е неподвижна, овие структури туркаат вода, честички од храна и други честички кон или подалеку од клетката. Флагелите се релативно големи, и обично клетката има само една, понекогаш и неколку, флагели. Цилиите се многу помали и ја покриваат целата површина на клетката. Иако овие структури се карактеристични главно за наједноставните, тие можат да бидат присутни и во високо организирани форми. Во човечкото тело, сите респираторни патишта се обложени со цилии. Малите честички кои влегуваат во нив обично се фатени од слузта на површината на клетката, а цилиите ги туркаат заедно со слузта надвор и на тој начин ги штитат белите дробови. Машките репродуктивни клетки на повеќето животни и некои пониски растенија се движат со помош на флагелум.
Постојат и други видови на клеточно движење. Еден од нив е амебоидно движење. Амебата, како и некои клетки на повеќеклеточни организми, „течат“ од место до место, т.е. се движат поради струјата на содржината на ќелијата. Постојана струја на материја постои и во растителните клетки, но тоа не повлекува движење на клетката како целина. Најпроучен тип на клеточно движење е контракцијата на мускулните клетки; се врши со лизгање на фибрили (протеински нишки) релативно едни на други, што доведува до скратување на клетката.
КОР
Јадрото е опкружено со двојна мембрана. Многу тесниот (околу 40 nm) простор помеѓу две мембрани се нарекува перинуклеарен. Нуклеарните мембрани минуваат во мембраните на ендоплазматскиот ретикулум, а перинуклеарниот простор се отвора во ретикуларниот простор. Обично нуклеарната мембрана има многу тесни пори. Очигледно, преку нив се транспортираат големи молекули, како што е гласник РНК, која се синтетизира на ДНК и потоа влегува во цитоплазмата.
Најголемиот дел од генетскиот материјал се наоѓа во хромозомите на клеточното јадро. Хромозомите се состојат од долги синџири на двоверижна ДНК, на кои се прикачени основни (т.е., алкални) протеини. Понекогаш хромозомите имаат неколку идентични ДНК нишки кои лежат една до друга - таквите хромозоми се нарекуваат политен (мулти-верижна). Бројот на хромозоми варира меѓу видовите. Диплоидните клетки на човечкото тело содржат 46 хромозоми или 23 пара.
Во клетка која не се дели, хромозомите се прикачени на една или повеќе точки на нуклеарната мембрана. Во нивната нормална незавиткана состојба, хромозомите се толку тенки што не се видливи под светлосен микроскоп. На одредени места (пресеци) на еден или повеќе хромозоми, се формира густо тело, кое е присутно во јадрата на повеќето клетки - т.н. јадро. Во јадрата, се јавува синтеза и акумулација на РНК што се користи за изградба на рибозоми, како и некои други видови на РНК.
ДЕЛБА НА КЛЕТКИ
Иако сите клетки произлегуваат од поделбата на претходната клетка, не сите продолжуваат да се делат. На пример, нервните клетки во мозокот, откако ќе се формираат, не се делат. Нивниот број постепено се намалува; Оштетеното мозочно ткиво не може да се опорави преку регенерација. Ако клетките продолжат да се делат, тогаш тие се карактеризираат со клеточен циклус кој се состои од две главни фази: интерфаза и митоза.
Самата интерфаза се состои од три фази: G 1, S и G 2. Подолу е нивното времетраење, типично за растителни и животински клетки.
G 1 (4-8 часа). Оваа фаза започнува веднаш по раѓањето на клетката. За време на фазата G 1, клетката, со исклучок на хромозомите (кои не се менуваат), ја зголемува својата маса. Ако клетката дополнително не се дели, таа останува во оваа фаза.
S (6–9 часа). Клеточната маса продолжува да се зголемува, а настанува удвојување (дуплирање) на хромозомската ДНК. Сепак, хромозомите остануваат единечни во структурата, иако двојно зголемени во масата, бидејќи по две копии од секој хромозом (хроматиди) сè уште се поврзани едни со други по целата должина.
Г2. Масата на клетката продолжува да се зголемува додека не биде приближно двојно поголема од нејзината првобитна маса, а потоа се јавува митоза.
Откако ќе се удвојат хромозомите, секоја од ќерките клетки треба да добие целосен сет на хромозоми. Едноставната клеточна делба не може да го постигне ова - овој резултат се постигнува преку процес наречен митоза. Без да навлегуваме во детали, почетокот на овој процес треба да се смета за усогласување на хромозомите во екваторијалната рамнина на клетката. Потоа секој хромозом се дели надолжно на два хроматиди, кои почнуваат да се разминуваат во спротивни насоки, станувајќи независни хромозоми. Како резултат на тоа, комплетен сет на хромозоми се наоѓа на двата краја на клетката. Клетката потоа се дели на два, а секоја клетка ќерка добива целосен сет на хромозоми.
Следното е опис на митозата во типична животинска клетка. Обично се дели на четири фази.
I. Профаза. Посебна клеточна структура - центриолот - се удвојува (понекогаш ова удвојување се случува во S-периодот на интерфазата), а двете центриоли почнуваат да се разминуваат до спротивните полови на јадрото. Нуклеарната мембрана е уништена; во исто време, специјалните протеини се комбинираат (агрегат), формирајќи микротубули во форма на нишки. Центриолите, сега сместени на спротивните полови на клетката, имаат организационен ефект врз микротубулите, кои како резултат се наредени радијално, формирајќи структура која потсетува на астер цвет („ѕвезда“). Други нишки на микротубули се протегаат од една центриола до друга, формирајќи т.н. вретено на фисија. Во тоа време, хромозомите се во спирална состојба, налик на пружина. Тие се јасно видливи во светлосен микроскоп, особено по боење. Во профаза, хромозомите се поделени, но хроматидите сè уште остануваат врзани во парови во зоната на центромерот - хромозомска органела слична по функција на центриолот. Центромерите, исто така, имаат организационен ефект врз филаментите на вретеното, кои сега се протегаат од центриола до центромер и од неа до друга центриола.
II. Метафаза. Хромозомите, до овој момент распоредени по случаен избор, почнуваат да се движат, како да се нацртани со вретено навои прикачени на нивните центромери и постепено се редат во иста рамнина во одредена положба и на еднакво растојание од двата пола. Центромерите кои лежат во иста рамнина заедно со хромозомите го формираат т.н. екваторијална плоча. Центромерите што ги поврзуваат паровите на хроматиди се делат, по што сестринските хромозоми се целосно одвоени.
III. Анафаза. Хромозомите на секој пар се движат во спротивни насоки кон половите, како да се влечат со нишки на вретеното. Во овој случај, нишки се формираат и помеѓу центромерите на спарените хромозоми.
IV. Телофаза. Штом хромозомите се приближуваат до спротивните полови, самата клетка почнува да се дели по рамнината во која се наоѓала екваторијалната плоча. Како резултат на тоа, се формираат две клетки. Нишките на вретеното се уништуваат, хромозомите се одмотуваат и стануваат невидливи, а околу нив се формира нуклеарна мембрана. Клетките се враќаат во G 1 фаза од интерфазата. Целиот процес на митоза трае околу еден час.
Деталите за митозата варираат донекаде кај различните типови на клетки. Типична растителна клетка формира вретено, но нема центриоли. Кај габите, митозата се јавува внатре во јадрото, без претходно распаѓање на нуклеарната мембрана.
Поделбата на самата клетка, наречена цитокинеза, нема строга врска со митозата. Понекогаш една или повеќе митози се јавуваат без клеточна делба; Како резултат на тоа, се формираат мултинуклеарни клетки, кои често се наоѓаат во алгите. Ако јадрото се отстрани од јајцето на морскиот еж преку микроманипулација, вретеното продолжува да се формира и јајцето продолжува да се дели. Ова покажува дека присуството на хромозоми не е неопходен услов за клеточната делба.
Репродукцијата со митоза се нарекува асексуална репродукција, вегетативна репродукција или клонирање. Нејзиниот најважен аспект е генетскиот: со таква репродукција, нема дивергенција на наследните фактори кај потомството. Добиените ќерки ќерки генетски се сосема исти како и матичната клетка. Митозата е единствениот начин на саморепродукција кај видовите кои немаат сексуална репродукција, како што се многу едноклеточни организми. Сепак, дури и кај видовите со сексуална репродукција, телесните клетки се делат преку митоза и доаѓаат од една клетка, оплодената јајце клетка, и затоа сите се генетски идентични. Високите растенија можат да се размножуваат бесполово (со користење на митоза) со садници и ластари (добро познат пример се јагодите).
Сексуалната репродукција на организмите се врши со помош на специјализирани клетки, т.н. гамети - ооцити (јајца) и сперматозоиди (сперматозоиди). Гаметите се спојуваат за да формираат една клетка - зигот. Секоја гамета е хаплоидна, т.е. има еден сет на хромозоми. Во комплетот, сите хромозоми се различни, но секој хромозом на јајце клетката одговара на еден од хромозомите на спермата. Според тоа, зиготот веќе содржи пар хромозоми кои одговараат еден на друг, кои се нарекуваат хомологни. Хомологните хромозоми се слични бидејќи имаат исти гени или нивни варијанти (алели) кои одредуваат специфични карактеристики. На пример, еден од спарените хромозоми може да има ген кој ја кодира крвната група А, а другиот може да има варијанта што ја кодира крвната група Б. Хромозомите на зиготот кои потекнуваат од јајце клетката се мајчини, а оние кои потекнуваат од спермата се татковски.
Како резултат на повторени митотични поделби, од добиениот зигот произлегува или повеќеклеточен организам или бројни слободни живи клетки, како што се случува кај протозоите кои имаат сексуална репродукција и кај едноклеточните алги.
За време на формирањето на гамети, диплоидниот сет на хромозоми присутни во зиготот мора да се намали за половина. Доколку тоа не се случеше, тогаш во секоја генерација спојувањето на гамети ќе доведе до удвојување на множеството хромозоми. Намалување на хаплоиден број на хромозоми се јавува како резултат на намалување на поделба - т.н. мејоза, која е варијанта на митоза.
Расцепување и рекомбинација.
Особеноста на мејозата е тоа што за време на клеточната делба, екваторијалната плоча се формира од парови хомологни хромозоми, а не од дупликати поединечни хромозоми, како во митозата. Спарените хромозоми, од кои секој останува сингл, се разминуваат до спротивните полови на клетката, клетката се дели и како резултат на тоа, клетките ќерки добиваат половина од множеството хромозоми во споредба со зиготот.
На пример, да претпоставиме дека хаплоидниот сет се состои од два хромозома. Во зиготот (и соодветно во сите клетки на организмот што произведува гамети) се присутни мајчините хромозоми А и Б и татковските хромозоми А" и Б". За време на мејозата тие можат да се поделат на следниов начин:
Најважното нешто во овој пример е фактот дека кога хромозомите се разминуваат, оригиналниот мајчин и татковски сет не е нужно формиран, но можна е рекомбинација на гените, како кај гаметите AB" и A"B на горниот дијаграм.
Сега да претпоставиме дека хромозомскиот пар АА“ содржи два алели - аИ б– ген кој ги одредува крвните групи А и Б. Слично, парот хромозоми „ББ“ содржи алели мИ nдруг ген кој ги одредува крвните групи М и Н. Одвојувањето на овие алели може да се одвива на следниов начин:
Очигледно, добиените гамети можат да содржат која било од следниве комбинации на алели на двата гени: сум, бн, bmили ан.
Ако има повеќе хромозоми, тогаш парови алели ќе се одделат независно според истиот принцип. Ова значи дека истите зиготи можат да произведат гамети со различни комбинации на генски алели и да доведат до различни генотипови кај потомството.
Мејотичка поделба.
И двата примера го илустрираат принципот на мејоза. Всушност, мејозата е многу покомплексен процес, бидејќи вклучува две последователни поделби. Главната работа во мејозата е дека хромозомите се удвојуваат само еднаш, додека клетката се дели двапати, како резултат на што бројот на хромозоми се намалува и диплоидниот сет се претвора во хаплоиден.
За време на профазата на првата поделба, хомологните хромозоми се конјугираат, односно се спојуваат во парови. Како резултат на овој многу прецизен процес, секој ген завршува спроти својот хомолог на друг хромозом. И двата хромозома потоа се удвојуваат, но хроматидите остануваат поврзани едни со други со заеднички центромер.
Во метафазата, четирите поврзани хроматиди се редат за да ја формираат екваторијалната плоча, како да се еден дупликат хромозом. Спротивно на она што се случува во митозата, центромерите не се делат. Како резултат на тоа, секоја клетка ќерка добива пар хроматиди кои сè уште се поврзани со центромерот. За време на втората поделба, хромозомите, веќе индивидуални, повторно се редат, формирајќи, како во митозата, екваторијална плоча, но нивното удвојување не се случува при оваа поделба. Центромерите потоа се делат и секоја клетка ќерка добива по еден хроматид.
Цитоплазматска поделба.
Како резултат на две мејотски поделби на диплоидна клетка, се формираат четири клетки. Кога се формираат машките репродуктивни клетки, се добиваат четири сперматозоиди со приближно иста големина. Кога се формираат јајца, поделбата на цитоплазмата се случува многу нерамномерно: една клетка останува голема, додека другите три се толку мали што речиси целосно се окупирани од јадрото. Овие мали клетки, т.н. поларните тела служат само за сместување на вишокот хромозоми формирани како резултат на мејоза. Најголемиот дел од цитоплазмата неопходна за зиготот останува во една клетка - јајцето.
Конјугација и вкрстување.
За време на конјугацијата, хроматидите на хомологните хромозоми можат да се скршат и потоа да се спојат во нов редослед, разменувајќи ги деловите на следниов начин:

Оваа размена на делови од хомологни хромозоми се нарекува вкрстување. Како што е прикажано погоре, вкрстувањето води до појава на нови комбинации на алели на поврзани гени. Значи, ако оригиналните хромозоми имале комбинации АБИ ab, потоа по преминувањето ќе содржат АбИ aB. Овој механизам за појава на нови комбинации на гени го надополнува ефектот на независно сортирање на хромозомите што се јавува за време на мејозата. Разликата е во тоа што вкрстувањето ги одвојува гените на истиот хромозом, додека независното сортирање ги одвојува само гените на различни хромозоми.
НАМЕНИ ГЕНЕРАЦИИ
ПРИМИТИВНИ КЛЕТКИ: ПРОКАРИОТИ
Сето горенаведено се однесува на клетките на растенијата, животните, протозоите и едноклеточните алги, колективно наречени еукариоти. Еукариотите еволуирале од поедноставна форма, прокариоти, кои сега се претставени со бактерии, вклучувајќи архебактерии и цијанобактерии (последниве порано се нарекувале сино-зелени алги). Во споредба со еукариотските клетки, прокариотските клетки се помали и имаат помалку клеточни органели. Тие имаат клеточна мембрана, но немаат ендоплазматичен ретикулум, а рибозомите слободно лебдат во цитоплазмата. Митохондриите се отсутни, но оксидативните ензими обично се прикачени на клеточната мембрана, која на тој начин станува еквивалент на митохондриите. На прокариотите им недостасуваат и хлоропласти, а хлорофилот, доколку е присутен, е присутен во форма на многу мали гранули.
Прокариотите немаат јадро затворено со мембрана, иако локацијата на ДНК може да се идентификува по нејзината оптичка густина. Еквивалент на хромозом е влакно на ДНК, обично кружно, со многу помалку прикачени протеини. ДНК синџирот е прикачен на клеточната мембрана во една точка. Не постои митоза кај прокариотите. Се заменува со следниот процес: ДНК се удвојува, по што клеточната мембрана почнува да расте помеѓу соседните точки на прицврстување на две копии од молекулата на ДНК, кои како резултат постепено се разминуваат. Клетката на крајот се дели помеѓу точките на прицврстување на молекулите на ДНК, формирајќи две клетки, секоја со своја копија од ДНК.
ДИФЕРЕНЦИЈА НА КЛЕТКИ
Повеќеклеточните растенија и животни еволуирале од едноклеточни организми чии клетки, по делењето, останале заедно за да формираат колонија. Првично, сите клетки беа идентични, но понатамошната еволуција доведе до диференцијација. Како прво, се диференцираат соматските клетки (т.е. телесните клетки) и герминативните клетки. Понатамошната диференцијација стана посложена - се појавија се повеќе и повеќе различни типови на клетки. Онтогенезата - индивидуалниот развој на повеќеклеточниот организам - го повторува во општи термини овој еволутивен процес (филогенија).
Физиолошки, клетките делумно се разликуваат со подобрување на една или друга карактеристика заедничка за сите клетки. На пример, контрактилната функција е засилена во мускулните клетки, што може да биде резултат на подобрување на механизмот што врши амебоиди или други видови на движење во помалку специјализирани клетки. Сличен пример се тенкоѕидните коренски клетки со нивните процеси, т.н. коренски влакна, кои служат за апсорпција на соли и вода; до еден или друг степен, оваа функција е вродена во сите клетки. Понекогаш специјализацијата е поврзана со стекнување нови структури и функции - пример е развојот на локомоторен орган (флагелум) во спермата.
Диференцијацијата на клеточно или ткивно ниво е проучена во некои детали. Знаеме, на пример, дека понекогаш се случува автономно, т.е. еден тип на клетка може да се претвори во друг без оглед на тоа каков тип на клетки се соседните клетки. Сепак, т.н ембрионската индукција е феномен во кој еден вид ткиво ги стимулира клетките од друг тип да се разликуваат во дадена насока.
Во општиот случај, диференцијацијата е неповратна, т.е. високо диференцираните клетки не можат да се трансформираат во друг тип на клетка. Сепак, тоа не е секогаш случај, особено кај растителните клетки.
Разликите во структурата и функцијата на крајот се одредуваат од тоа какви видови на протеини се синтетизираат во клетката. Бидејќи синтезата на протеините е контролирана од гените, а збирот на гени е ист во сите клетки на телото, диференцијацијата мора да зависи од активирањето или инактивирањето на одредени гени во различни типови на клетки. Регулирањето на генската активност се јавува на ниво на транскрипција, т.е. формирање на гласник РНК користејќи ДНК како шаблон. Само транскрибираните гени произведуваат протеини. Синтетизираните протеини можат да ја блокираат транскрипцијата, но понекогаш и да ја активираат. Покрај тоа, бидејќи протеините се производи на гени, некои гени можат да ја контролираат транскрипцијата на други гени. Хормоните, особено стероидите, исто така се вклучени во регулирањето на транскрипцијата. Многу активните гени може да се дуплираат (удвојуваат) многу пати за да се произведе повеќе гласник РНК.
Развојот на малигни тумори често се смета за посебен случај на клеточна диференцијација. Сепак, појавата на малигните клетки е резултат на промените во структурата на ДНК (мутација), а не на процесите на транскрипција и транслација во протеин на нормална ДНК.
МЕТОДИ ЗА ПРОУЧУВАЊЕ НА КЛЕТКИТЕ
Светлосен микроскоп.
Во проучувањето на формата и структурата на клетките, првата алатка беше светлосниот микроскоп. Неговата моќ на разрешување е ограничена со димензии споредливи со брановата должина на светлината (0,4-0,7 μm за видлива светлина). Сепак, многу елементи на клеточната структура се многу помали по големина.
Друга тешкотија е што повеќето клеточни компоненти се проѕирни и имаат индекс на рефракција речиси ист како водата. За да се подобри видливоста, често се користат бои кои имаат различни афинитети за различни клеточни компоненти. Боењето се користи и за проучување на клеточната хемија. На пример, некои бои преференцијално се врзуваат за нуклеинските киселини и со тоа ја откриваат нивната локализација во клетката. Мал дел од бои - наречени интравитални бои - може да се користат за обојување на живите клетки, но обично клетките мора прво да се фиксираат (со користење на протеински коагулациони супстанции) пред да можат да се обојат. Цм. ХИСТОЛОГИЈА.
Пред тестирањето, клетките или парчињата ткиво обично се вградуваат во парафин или пластика, а потоа се сечат на многу тенки делови со помош на микротом. Овој метод е широко користен во клиничките лаборатории за да се идентификуваат клетките на туморот. Покрај конвенционалната светлосна микроскопија, развиени се и други оптички методи за проучување на клетките: флуоресцентна микроскопија, фазна контрастна микроскопија, спектроскопија и анализа на дифракција на Х-зраци.
Електронски микроскоп.
Електронскиот микроскоп има резолуција од прибл. 1-2 nm. Ова е доволно за проучување на големи протеински молекули. Обично е потребно да се обои и да се направи контраст на предметот со метални соли или метали. Поради оваа причина, а исто така и поради тоа што предметите се испитуваат во вакуум, само убиените клетки може да се проучуваат со помош на електронски микроскоп.
Авторадиографија.
Доколку во медиумот се додаде радиоактивен изотоп кој се апсорбира од клетките за време на метаболизмот, неговата интрацелуларна локализација потоа може да се открие со помош на авторрадиографија. Со овој метод, тенки делови од клетки се ставаат на филм. Филмот потемнува под оние места каде што се наоѓаат радиоактивни изотопи.
Центрифугирање.
За биохемиско проучување на клеточните компоненти, клетките мора да се уништат - механички, хемиски или ултразвук. Ослободените компоненти се суспендирани во течноста и може да се изолираат и прочистат со центрифугирање (најчесто во градиент на густина). Вообичаено, таквите прочистени компоненти задржуваат висока биохемиска активност.
Клеточни култури.
Некои ткива може да се поделат на поединечни клетки, така што клетките остануваат живи и често се способни да се репродуцираат. Овој факт дефинитивно ја потврдува идејата за клетката како жива единица. Сунѓерот, примитивен повеќеклеточен организам, може да се одвои во клетки со триење низ сито. По некое време, овие клетки повторно се поврзуваат и формираат сунѓер. Животинските ембрионски ткива може да се натераат да се дисоцираат со помош на ензими или други средства кои ги ослабуваат врските помеѓу клетките.
Американскиот ембриолог R. Harrison (1879–1959) беше првиот што покажа дека ембрионските, па дури и некои зрели клетки можат да растат и да се размножуваат надвор од телото во соодветна средина. Оваа техника, наречена клеточно одгледување, била усовршена од францускиот биолог А. Карел (1873–1959). Растителните клетки можат да се одгледуваат и во култура, но во споредба со животинските клетки тие формираат поголеми грутки и се поцврсто прицврстени една за друга, така што ткивата се формираат како што расте културата, наместо поединечни клетки. Во клеточната култура, цело возрасно растение, како што е морков, може да се одгледува од една клетка.
Микрохирургија.
Со помош на микроманипулатор, одделни делови од ќелијата може да се отстранат, додадат или изменат на некој начин. Голема клетка на амеба може да се подели на три главни компоненти - клеточната мембрана, цитоплазмата и јадрото, а потоа овие компоненти може повторно да се соберат за да формираат жива клетка. На овој начин може да се добијат вештачки клетки составени од компоненти на различни видови амеби.
Ако се земе предвид дека се чини возможно да се синтетизираат некои клеточни компоненти вештачки, тогаш експериментите во склопување на вештачки клетки може да бидат првиот чекор кон создавање нови форми на живот во лабораторија. Бидејќи секој организам се развива од една клетка, методот на производство на вештачки клетки во принцип дозволува изградба на организми од даден тип, ако во исто време се користат компоненти малку различни од оние што се наоѓаат во постоечките клетки. Меѓутоа, во реалноста, не е потребна целосна синтеза на сите клеточни компоненти. Структурата на повеќето, ако не и на сите, компоненти на клетката е одредена од нуклеинските киселини. Така, проблемот со создавањето на нови организми се сведува на синтеза на нови видови нуклеински киселини и нивна замена на природните нуклеински киселини во одредени клетки.
Фузија на клетки.
Друг тип на вештачки клетки може да се добие со спојување на клетки од ист или различен вид. За да се постигне фузија, клетките се изложени на вирусни ензими; во овој случај, надворешните површини на две клетки се залепени заедно, а мембраната меѓу нив е уништена и се формира клетка во која две групи на хромозоми се затворени во едно јадро. Можно е да се спојат клетки од различни типови или во различни фази на поделба. Користејќи го овој метод, беше можно да се добијат хибридни клетки на глушец и кокошка, човек и глушец и човек и жаба. Таквите клетки се хибридни само на почетокот, а по бројните клеточни делби ги губат повеќето од хромозомите од едниот или од другиот тип. Финалниот производ станува, на пример, во суштина клетка од глушец без присутна или присутна само трага од човечки гени. Од особен интерес е спојувањето на нормалните и малигните клетки. Во некои случаи, хибридите стануваат малигни, во други не, т.е. двете својства можат да се манифестираат и како доминантни и како рецесивни. Овој резултат не е неочекуван, бидејќи малигнитет може да бидат предизвикани од различни фактори и има сложен механизам.
Литература:
Хем А., Кормак Д. Хистологија, том 1. М., 1982 година
Албертс Б., Бреј Д., Луис Ј., Раф М., Робертс К., Вотсон Џ. Молекуларна клеточна биологија, том 1. М., 1994 година







