Верверички- висока молекуларна тежина органски соединенија, кој се состои од остатоци од α-аминокиселини.
ВО протеински составвклучува јаглерод, водород, азот, кислород, сулфур. Некои протеини формираат комплекси со други молекули кои содржат фосфор, железо, цинк и бакар.
Протеините имаат голема молекуларна тежина: албумин од јајца - 36.000, хемоглобин - 152.000, миозин - 500.000. За споредба: молекуларната тежина на алкохолот е 46, оцетна киселина- 60, бензен - 78.
Аминокиселински состав на протеини
Верверички- непериодични полимери, чии мономери се α-амино киселини. Типично, 20 видови на α-амино киселини се нарекуваат протеински мономери, иако над 170 од нив се наоѓаат во клетките и ткивата.
Во зависност од тоа дали аминокиселините можат да се синтетизираат во телото на луѓето и другите животни, тие се разликуваат: неесенцијални амино киселини- може да се синтетизира; есенцијални амино киселини- не може да се синтетизира. Есенцијални амино киселинимора да влезе во телото со храна. Растенијата ги синтетизираат сите видови амино киселини.
Во зависност од составот на аминокиселините, протеините се: целосни- го содржи целиот сет на амино киселини; неисправни- некои амино киселини недостасуваат во нивниот состав. Ако протеините се состојат само од амино киселини, тие се нарекуваат едноставно. Ако протеините содржат, покрај аминокиселините, неаминокиселинска компонента (протетска група), тие се нарекуваат комплекс. Протетската група може да биде претставена со метали (металопротеини), јаглени хидрати (гликопротеини), липиди (липопротеини), нуклеински киселини (нуклеопротеини).
Сите амино киселините содржат: 1) карбоксилна група (-COOH), 2) амино група (-NH 2), 3) радикална или R-група (остатокот од молекулата). Структурата на радикалот различни типовиамино киселини - различни. Во зависност од бројот на амино групи и карбоксилни групи вклучени во составот на амино киселините, тие се разликуваат: неутрални амино киселиниима една карбоксилна група и една амино група; основни амино киселиниима повеќе од една амино група; кисели амино киселиниима повеќе од една карбоксилна група.
Амино киселините се амфотерични соединенија , бидејќи во раствор тие можат да дејствуваат и како киселини и како бази. ВО водени раствориамино киселините постојат во различни јонски форми.
Пептидна врска
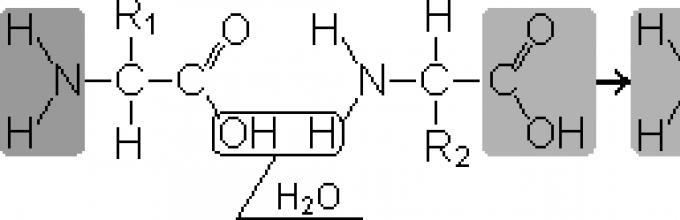
Пептиди- органски материи кои се состојат од остатоци од аминокиселини поврзани со пептидни врски.
Формирањето на пептиди се јавува како резултат на реакцијата на кондензација на амино киселините. Кога амино групата на една аминокиселина е во интеракција со карбоксилната група на друга, меѓу нив се јавува ковалентна азотно-јаглеродна врска, која се нарекува. пептид. Во зависност од бројот на аминокиселински остатоци вклучени во пептидот, постојат дипептиди, трипептиди, тетрапептидиитн. Формирањето на пептидна врска може да се повтори многу пати. Ова доведува до формирање полипептиди. На едниот крај на пептидот има слободна амино група (наречена N-терминал), а на другиот има слободна карбоксилна група (наречена C-крај).

Просторна организација на протеинските молекули
Извршувањето на одредени специфични функции од страна на протеините зависи од просторната конфигурација на нивните молекули, освен тоа, енергетски е неповолно за клетката да ги одржува протеините во расклопена форма, во форма на синџир, затоа полипептидните синџири се подложени на превиткување, стекнувајќи одредена тродимензионална структура или конформација. Има 4 нивоа просторна организацијапротеини.
Примарна структураверверица- редоследот на распоредот на амино киселинските остатоци во полипептидниот синџир што ја сочинува протеинската молекула. Врската помеѓу амино киселините е пептидна врска.

Ако протеинската молекула се состои од само 10 остатоци од аминокиселини, тогаш бројот е теоретски можни опциипротеински молекули кои се разликуваат по редоследот на алтернација на амино киселини - 10 20. Имајќи 20 амино киселини, можете да направите уште поразновидни комбинации од нив. Во човечкото тело се пронајдени околу десет илјади различни протеини, кои се разликуваат како едни од други, така и од протеините на другите организми.
Примарната структура на протеинската молекула е таа што ги одредува својствата на протеинските молекули и нејзината просторна конфигурација. Замената на само една аминокиселина со друга во полипептидниот синџир доведува до промена на својствата и функциите на протеинот. На пример, заменувањето на шестата глутаминска амино киселина во β-подединицата на хемоглобинот со валин води до фактот дека молекулата на хемоглобинот како целина не може да ја изврши својата главна функција - транспорт на кислород; Во такви случаи, лицето развива болест наречена српеста анемија.
Секундарна структура- наредено преклопување на полипептидниот синџир во спирала (изгледа како продолжена пружина). Вртежите на спиралата се зајакнати со водородни врски што се појавуваат помеѓу карбоксилните групи и амино групите. Речиси сите CO и NH групи учествуваат во формирањето на водородни врски. Тие се послаби од пептидните, но, повторени многу пати, даваат стабилност и цврстина на оваа конфигурација. На ниво на секундарна структура, постојат протеини: фиброин (свила, пајакова мрежа), кератин (коса, нокти), колаген (тетиви).



Терцијарна структура- пакување на полипептидни синџири во глобули кои произлегуваат од појавата хемиски врски(водород, јонски, дисулфид) и воспоставување на хидрофобни интеракции помеѓу радикалите на амино киселинските остатоци. Главната улога во формирањето на терциерната структура ја играат хидрофилно-хидрофобните интеракции. Во водените раствори, хидрофобните радикали имаат тенденција да се кријат од водата, групирајќи се во глобулата, додека хидрофилните радикали, како резултат на хидратација (интеракција со водните диполи), имаат тенденција да се појавуваат на површината на молекулата. Кај некои протеини, терциерната структура се стабилизира со дисулфидни ковалентни врски формирани помеѓу атомите на сулфур на два остатоци од цистеин. На ниво на терциерна структура има ензими, антитела и некои хормони.

Кватернарна структуракарактеристични за сложените протеини чии молекули се формирани од две или повеќе глобули. Подединиците се држат во молекулата со јонски, хидрофобни и електростатски интеракции. Понекогаш, за време на формирањето на кватернарна структура, се јавуваат дисулфидни врски помеѓу подединици. Најпроучен протеин со кватернарна структура е хемоглобинот. Формирана е од две α-подединици (141 остатоци од аминокиселини) и две β-подединици (146 остатоци од аминокиселини). Со секоја подединица е поврзана хем молекула која содржи железо.
Ако поради некоја причина просторната конформација на протеините отстапува од нормалата, протеинот не може да ги извршува своите функции. На пример, причината за „кравјо лудило“ (спонгиформна енцефалопатија) е абнормалната конформација на прионите, површинските протеини на нервните клетки.
Својства на протеините
Аминокиселинскиот состав и структурата на протеинската молекула го одредуваат својства. Протеините ги комбинираат основните и киселинските својства, определени со радикали на аминокиселини: колку повеќе кисели амино киселини во протеинот, толку поизразени се неговите кисели својства. Утврдена е способноста за донирање и додавање H + пуферски својства на протеините; Еден од најмоќните пуфери е хемоглобинот во црвените крвни зрнца, кој ја одржува pH вредноста на крвта на константно ниво. Постојат растворливи протеини (фибриноген), а има и нерастворливи протеини кои вршат механички функции (фиброин, кератин, колаген). Постојат протеини кои се хемиски активни (ензими), има хемиски неактивни протеини кои се отпорни на различни услови на животната средина и оние кои се крајно нестабилни.
Надворешни фактори (топлина, ултравиолетово зрачење, тешки металии нивните соли, pH промени, зрачење, дехидрација)
може да предизвика нарушување на структурната организација на протеинската молекула. Процесот на губење на тродимензионалната конформација својствена за дадена протеинска молекула се нарекува денатурација. Причината за денатурација е кршењето на врските кои стабилизираат одредена протеинска структура. Првично, најслабите врски се прекинуваат, а како што се заоструваат условите, се прекинуваат уште посилните. Затоа, прво се губат квартерните, потоа терциерните и секундарните структури. Промената во просторната конфигурација доведува до промена на својствата на протеинот и, како резултат на тоа, го оневозможува протеинот да ги извршува своите вродени биолошки функции. Ако денатурацијата не е придружена со уништување на примарната структура, тогаш тоа може да биде реверзибилна, во овој случај, се случува само-обновување на конформациската карактеристика на протеинот. На пример, мембранските рецепторни протеини се подложени на таква денатурација. Процесот на обновување на протеинската структура по денатурација се нарекува ренатурација. Ако обновувањето на просторната конфигурација на протеинот е невозможно, тогаш се нарекува денатурација неповратен.
Функции на протеините
| Функција | Примери и објаснувања |
|---|---|
| Градба | Протеините се вклучени во формирањето на клеточните и екстрацелуларните структури: тие се дел од клеточните мембрани (липопротеини, гликопротеини), косата (кератин), тетивите (колаген) итн. |
| Транспорт | Крвниот протеин хемоглобинот го прицврстува кислородот и го транспортира од белите дробови до сите ткива и органи, а од нив го пренесува јаглерод диоксидот во белите дробови; Составот на клеточните мембрани вклучува специјални протеини кои обезбедуваат активен и строго селективен трансфер на одредени супстанции и јони од клетката во надворешната средина и назад. |
| Регулаторна | Протеинските хормони учествуваат во регулирањето на метаболичките процеси. На пример, хормонот инсулин го регулира нивото на гликоза во крвта, ја промовира синтезата на гликоген и го зголемува формирањето на масти од јаглехидратите. |
| Заштитна | Како одговор на пенетрацијата на туѓи протеини или микроорганизми (антигени) во телото, се формираат специјални протеини - антитела кои можат да ги врзат и неутрализираат. Фибринот, формиран од фибриноген, помага да се запре крварењето. |
| Мотор | Контрактилните протеини актин и миозин обезбедуваат мускулна контракција кај повеќеклеточните животни. |
| Сигнал | Во површинската мембрана на клетката се вградени протеински молекули кои се способни да ја менуваат својата терциерна структура како одговор на факторите на околината, со што добиваат сигнали од надворешната средина и пренесуваат команди до клетката. |
| Складирање | Во телото на животните, протеините, по правило, не се складираат, со исклучок на албумин од јајца и млечен казеин. Но, благодарение на протеините, некои супстанции можат да се складираат во телото; на пример, за време на распаѓањето на хемоглобинот, железото не се отстранува од телото, туку се складира, формирајќи комплекс со протеинот феритин. |
| Енергија | Кога 1 g протеин се распаѓа во финални производи, се ослободува 17,6 kJ. Прво, протеините се разложуваат на амино киселини, а потоа во финалните производи - вода, јаглерод диоксид и амонијак. Меѓутоа, протеините се користат како извор на енергија само кога се трошат други извори (јаглехидрати и масти). |
| Каталитички | Една од најважните функции на протеините. Обезбедени од протеини - ензими кои ги забрзуваат биохемиските реакции што се случуваат во клетките. На пример, рибулоза бифосфат карбоксилаза ја катализира фиксацијата на CO 2 за време на фотосинтезата. |
Ензими
Ензими, или ензими, се посебна класа на протеини кои се биолошки катализатори. Благодарение на ензимите, биохемиските реакции се случуваат со огромна брзина. Брзината на ензимските реакции е десетици илјади пати (а понекогаш и милиони) поголема од брзината на реакциите што се случуваат со учество на неоргански катализатори. Супстанцијата на која делува ензимот се нарекува супстрат.
Ензимите се глобуларни протеини, структурни карактеристикиензимите можат да се поделат во две групи: едноставни и сложени. Едноставни ензимисе едноставни протеини, т.е. се состои само од амино киселини. Комплексни ензимисе сложени протеини, т.е. Покрај протеинскиот дел содржат и група од непротеинска природа - кофактор. Некои ензими користат витамини како кофактори. Молекулата на ензимот содржи посебен дел наречен активен центар. Активен центар- мал дел од ензимот (од три до дванаесет амино киселински остатоци), каде што врзувањето на супстратот или супстратите се случува за да се формира комплекс ензим-супстрат. По завршувањето на реакцијата, комплексот ензим-супстрат се распаѓа во ензимот и во производ(ите) на реакцијата. Некои ензими имаат (освен активните) алостерични центри- области на кои се прикачени регулаторите на брзината на ензимот ( алостерични ензими).

Реакциите на ензимската катализа се карактеризираат со: 1) висока ефикасност, 2) строга селективност и насока на дејство, 3) специфичност на супстратот, 4) фина и прецизна регулација. Подлогата и специфичноста на реакцијата на реакциите на ензимска катализа се објаснети со хипотезите на E. Fischer (1890) и D. Koshland (1959).
Е. Фишер (хипотеза за заклучување на клучот)сугерираше дека просторните конфигурации на активното место на ензимот и супстратот мора точно да одговараат една на друга. Подлогата се споредува со „клучот“, ензимот со „бравата“.
Д. Кошланд (хипотеза за ракавици)сугерираше дека просторната кореспонденција помеѓу структурата на подлогата и активниот центар на ензимот се создава само во моментот на нивната меѓусебна интеракција. Оваа хипотеза е исто така наречена хипотеза за индуцирана кореспонденција.
Брзината на ензимските реакции зависи од: 1) температура, 2) концентрација на ензимот, 3) концентрација на супстратот, 4) pH вредност. Треба да се нагласи дека бидејќи ензимите се протеини, нивната активност е најголема кога е физиолошки нормални услови.
Повеќето ензими можат да работат само на температури помеѓу 0 и 40°C. Во овие граници, брзината на реакција се зголемува приближно 2 пати со секое зголемување на температурата за 10 °C. На температури над 40 °C, протеинот се подложува на денатурација и ензимската активност се намалува. На температури блиску до замрзнување, ензимите се деактивираат.
Како што се зголемува количината на супстратот, брзината на ензимската реакција се зголемува додека бројот на молекули на супстратот не се изедначи со бројот на ензимските молекули. Со дополнително зголемување на количината на супстрат, брзината нема да се зголеми, бидејќи активните центри на ензимот се заситени. Зголемувањето на концентрацијата на ензимот доведува до зголемена каталитичка активност, бидејќи поголем број на молекули на супстратот се подложени на трансформации по единица време.

За секој ензим, постои оптимална pH вредност при која покажува максимална активност (пепсин - 2,0, плунковна амилаза - 6,8, панкреасна липаза - 9,0). При повисоки или пониски pH вредности, ензимската активност се намалува. Со ненадејни промени во pH вредноста, ензимот се денатурира.
Брзината на алостеричните ензими е регулирана со супстанции кои се прикачуваат на алостерични центри. Ако овие супстанции ја забрзаат реакцијата, тие се нарекуваат активатори, ако успорат - инхибитори.
Класификација на ензими
Според видот на хемиските трансформации што ги катализираат, ензимите се поделени во 6 класи:
- оксиредуктази(пренос на атоми на водород, кислород или електрони од една супстанција во друга - дехидрогеназа),
- трансферази(пренос на метил, ацил, фосфат или амино група од една супстанција во друга - трансаминаза),
- хидролази(реакции на хидролиза во кои од супстратот се формираат два производи - амилаза, липаза),
- лијази(нехидролитичко додавање на подлогата или одвојување на група атоми од него, при што може да се прекинат врските C-C, C-N, C-O, C-S - декарбоксилаза),
- изомерази(интрамолекуларно преуредување - изомераза),
- лигази(поврзување на две молекули како резултат на формирањето C-C врски, C-N, C-O, C-S - синтетаза).
Класите за возврат се поделени на подкласи и поткласи. Во тековната меѓународна класификацијаСекој ензим има специфичен код, кој се состои од четири броеви одделени со точки. Првиот број е класа, вториот е подкласа, третиот е подкласа, четвртиот е сериски бројензимот во оваа подкласа, на пример, кодот на аргиназа е 3.5.3.1.
Оди до предавања бр.2„Структура и функции на јаглени хидрати и липиди“
Оди до предавања бр.4„Структура и функции нуклеински киселиниАТП"
Краток опис:
Фрагмент од учебникот: Биолошка хемија со вежби и задачи: учебник / ед. Дописен член RAMS S.E. Северина. М.: ГЕОТАР-Медиа, 2011. - 624 стр.: ill. МОДУЛ 1: СТРУКТУРА, СВОЈСТВА И ФУНКЦИИ НА ПРОТЕИНИ
МОДУЛ 1: СТРУКТУРА, СВОЈСТВА И ФУНКЦИИ НА ПРОТЕИНИ
Структура на модулот | Теми |
Модуларна единица 1 | 1.1. Структурна организација на протеините. Фази на формирање на конформација на мајчин протеин 1.2. Основи на функционирање на протеините. Лекови како лиганди кои влијаат на функцијата на протеините 1.3. Денатурација на протеините и можност за нивно спонтано обновување |
Модуларна единица 2 | 1.4. Карактеристики на структурата и функционирањето на олигомерните протеини користејќи го примерот на хемоглобинот 1.5. Одржување на конформацијата на природните протеини во клеточни услови 1.6. Разновидност на протеини. Протеински семејства користејќи го примерот на имуноглобулините 1.7. Физичко-хемиски својства на протеините и методи за нивно одвојување |
Модуларна единица 1 СТРУКТУРНА ОРГАНИЗАЦИЈА НА МОНОМЕРНИ ПРОТЕИНИ И ОСНОВИТЕ НА НИВНО ФУНКЦИОНИРАЊЕ
Цели на учењето Бидете способни:
1. Користете знаење за структурните карактеристики на протеините и зависноста на протеинските функции од нивната структура за да ги разберете механизмите на развој на наследни и стекнати протеинопатии.
2. Објаснете ги механизмите на терапевтското дејство на некои лекови како лиганди кои комуницираат со протеините и ја менуваат нивната активност.
3. Користете знаење за структурата и конформациската лабилност на протеините за да ја разберете нивната структурна и функционална нестабилност и тенденција за денатурација при променливи услови.
4. Објаснете ја употребата на средства за денатурирање како средства за стерилизирање медицински материјали и инструменти, како и антисептици.
Знајте:
1. Нивоа на структурна организација на протеините.
2. Важноста на примарната структура на протеините, која ја одредува нивната структурна и функционална разновидност.
3. Механизмот на формирање на активниот центар во протеините и неговата специфична интеракција со лигандот, кој лежи во основата на функционирањето на протеините.
4. Примери за влијанието на егзогените лиганди (лекови, токсини, отрови) врз конформацијата и функционалната активност на протеините.
5. Причини и последици од денатурација на протеини, фактори кои предизвикуваат денатурација.
6. Примери за употреба на денатурирани фактори во медицината како антисептици и средства за стерилизирање медицински инструменти.
ТЕМА 1.1. СТРУКТУРНА ОРГАНИЗАЦИЈА НА ПРОТЕИНИ. ФАЗИ НА ФОРМИРАЊЕ НА РОДЕН
ПРОТЕИНИ КОНФОРМАЦИИ
Верверички - ова се хетерополимери молекули (т.е. се состои од различни мономери). Протеинските мономери се 20 типови на α-амино киселини поврзани едни со други со пептидни врски.
Множеството и редоследот на комбинацијата на аминокиселините во протеинот се одредува според структурата на соодветните гени во ДНК на поединците. Секој протеин, во согласност со неговата специфична структура, врши своја функција. Збир на протеини на даден организам ( протеом) ги одредува неговите фенотипски карактеристики, како и присуството на наследни болести или предиспозиција за нивниот развој.
1. Амино киселини кои ги сочинуваат протеините. Пептидна врска.
Протеините се хетерополимери изградени од мономери - 20 α-амино киселини.
Општата формула на амино киселините е претставена подолу.

Амино киселините се разликуваат по структурата, големината и физичко-хемиските својства на радикалите прикачени на α-јаглеродниот атом. Функционалните групи на амино киселини ги одредуваат карактеристиките на својствата на различни α-амино киселини. Радикалите кои се наоѓаат во α-амино киселините може да се поделат во неколку групи:
Пролин,За разлика од другите 19 протеински мономери, тоа не е амино киселина, туку имино киселина; радикалот во пролинот е поврзан и со α-јаглеродниот атом и со имино групата
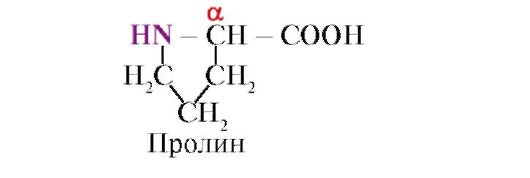
Амино киселините се разликуваат во растворливоста во вода.Ова се должи на способноста на радикалите да комуницираат со водата (хидрат).
ДО хидрофилнавклучуваат радикали кои содржат анјонски, катјонски и поларни ненаполнети функционални групи.
ДО хидрофобнивклучуваат радикали кои содржат метил групи, алифатични синџири или прстени.
2. Пептидните врски ги поврзуваат амино киселините за да формираат пептиди.За време на синтезата на пептидите, α-карбоксилната група на една аминокиселина комуницира со α-амино групата на друга аминокиселина за да формира пептидна врска:
Протеините се полипептиди, т.е. линеарни полимери на α-амино киселини поврзани со пептидна врска (сл. 1.1.)
 Ориз. 1.1. Термини кои се користат за опишување на структурата на пептидите
Ориз. 1.1. Термини кои се користат за опишување на структурата на пептидите
Мономерите на амино киселините кои ги сочинуваат полипептидите се нарекуваат остатоци од аминокиселини.Синџир од повторувачки групи - NH-CH-CO- форми пептиден 'рбет.Остатокот од аминокиселина што има слободна α-амино група се нарекува N-терминален, а оној што има слободна α-карбоксилна група се нарекува C-терминал. Пептидите се пишуваат и читаат од N-крајот до C-крајот.
Пептидната врска формирана од имино групата на пролин се разликува од другите пептидни врски: азотниот атом од пептидната група нема водород,
наместо тоа, постои врска со радикал, како резултат на што едната страна од прстенот е вклучена во пептидниот столб:

Пептидите се разликуваат по составот на аминокиселините, бројот на амино киселините и редоследот на поврзување со аминокиселините, на пример, Ser-Ala-Glu-Gis и His-Glu-Ala-Ser се два различни пептиди.
Пептидните врски се многу силни, а нивната хемиска неензимска хидролиза бара строги услови: анализираниот протеин се хидролизира во концентрирана хлороводородна киселинана температура од околу 110° 24 часа. Во жива ќелија пептидни врскиможе да се распарчи од протеолитички ензими,повикани протеазиили пептидни хидролази.
3. Примарна структура на протеините.Остатоците од амино киселините во пептидните синџири на различни протеини не се менуваат случајно, туку се распоредени по одреден редослед. Линеарната низа или редоследот на алтернација на аминокиселинските остатоци во полипептидниот синџир се нарекува примарна структура на протеинот.
Примарната структура на секој поединечен протеин е кодирана во молекулата на ДНК (во регионот наречен ген) и се реализира за време на транскрипцијата (копирање на информации на mRNA) и транслација (синтеза на примарната структура на протеинот). Следствено, примарната структура на протеините на поединечно лице е информацијата наследна пренесена од родителите на децата, која ги одредува структурните карактеристики на протеините на даден организам, од кои зависи функцијата на постоечките протеини (сл. 1.2.).
 Ориз. 1.2. Односот помеѓу генотипот и конформацијата на протеините синтетизирани во телото на поединецот
Ориз. 1.2. Односот помеѓу генотипот и конформацијата на протеините синтетизирани во телото на поединецот
Секој од околу 100.000 поединечни протеини во човечкото тело има единственпримарна структура. Молекулите од ист тип на протеин (на пример, албумин) имаат иста алтернација на амино киселински остатоци, што го разликува албуминот од кој било друг поединечен протеин.
Редоследот на остатоци од аминокиселини во пептидниот синџир може да се смета за форма на снимање информации. Оваа информација го одредува просторниот распоред на линеарниот пептиден синџир во покомпактна тридимензионална структура т.н. конформацијаверверица. Процесот на формирање на функционално активна протеинска конформација се нарекува виткање
4. Конформација на протеини.Можна е слободна ротација во пептидниот столб помеѓу азотниот атом од пептидната група и соседниот α-јаглероден атом, како и помеѓу α-јаглеродниот атом и јаглеродот од карбонилната група. Поради интеракцијата на функционалните групи на амино киселински остатоци, примарната структура на протеините може да стекне посложени просторни структури. Во глобуларните протеини, постојат две главни нивоа на преклопување на конформацијата на пептидните синџири: секундарноИ терцијарна структура.
Секундарна структура на протеиние просторна структура формирана како резултат на формирање на водородни врски помеѓу функционалните групи -C=O и -NH- на пептидниот рбет. Во овој случај, пептидниот синџир може да стекне правилни структури од два вида: α-спиралиИ β-структури.
ВО α-спиралисе формираат водородни врски помеѓу атомот на кислород од карбонилната група и водородот на амидниот азот од 4-та амино киселина од него; странични синџири на остатоци од аминокиселини
се наоѓаат по должината на периферијата на спиралата, без да учествуваат во формирањето на секундарната структура (сл. 1.3.).
Масовните радикали, или радикалите што носат еднакви полнежи, го спречуваат формирањето на α-спирала. Остатокот од пролин, кој има структура на прстен, ја прекинува α-спиралата, бидејќи поради недостаток на водород во азотниот атом во пептидниот синџир е невозможно да се формира водородна врска. Врската помеѓу азот и α-јаглеродниот атом е дел од пролинскиот прстен, така што пептидниот 'рбет станува свиткан во оваа точка.
β-Структурасе формира помеѓу линеарните региони на пептидниот столб на еден полипептиден синџир, со што се формираат преклопени структури. Може да се формираат полипептидни синџири или нивни делови паралелноили антипаралелни β-структури.Во првиот случај, N- и C-краевите на интерактивните пептидни синџири се совпаѓаат, а во вториот тие имаат спротивна насока (сл. 1.4).
Ориз. 1.3. Протеинска секундарна структура - α-спирала
 Ориз. 1.4. Паралелни и антипаралелни β-лист структури
Ориз. 1.4. Паралелни и антипаралелни β-лист структури
β-структурите се означени со широки стрелки: A - Антипаралелна β-структура. Б - Паралелни β-лист структури
Во некои протеини, β-структурите може да се формираат поради формирање на водородни врски помеѓу атомите на пептидниот столб на различни полипептидни синџири.
Се наоѓа и во протеините области со неправилни секундарниструктура, која вклучува свиоци, јамки и вртења на полипептидниот столб. Тие често се наоѓаат на места каде што се менува насоката на пептидниот синџир, на пример, кога се формира паралелна структура на β-лист.
Врз основа на присуството на α-спирали и β-структури, глобуларните протеини може да се поделат во четири категории.
Ориз. 1.5. Секундарна структура на миоглобинот (А) и хемоглобинскиот β-синџир (Б), кој содржи осум α-спирали
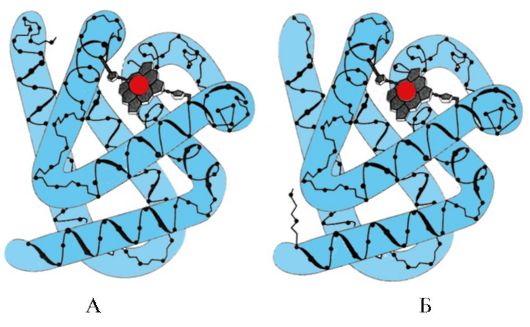
 Ориз. 1.6. Секундарна структура на доменот на триосефосфат изомераза и пируват киназа
Ориз. 1.6. Секундарна структура на доменот на триосефосфат изомераза и пируват киназа
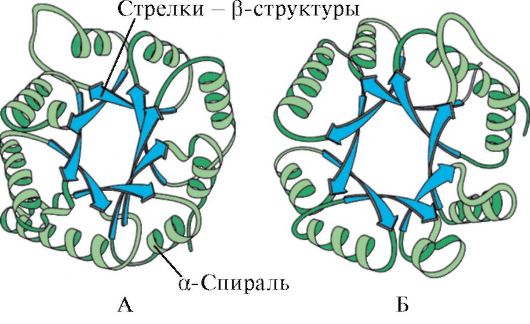
 Ориз. 1.7. Секундарна структура на константниот домен на имуноглобулинот (А) и ензимот супероксид дисмутаза (Б)
Ориз. 1.7. Секундарна структура на константниот домен на имуноглобулинот (А) и ензимот супероксид дисмутаза (Б)
ВО четврта категоријавклучени протеини кои содржат мала количина на редовни секундарни структури. Овие протеини вклучуваат мали протеини богати со цистеин или металопротеини.
Протеинска терцијарна структура- тип на конформација формирана поради интеракции помеѓу радикали на амино киселини, кои можат да се лоцираат на значително растојание едни од други во пептидниот синџир. Повеќето протеини формираат просторна структура слична на топче (глобуларни протеини).
Од хидрофобни радикали амино киселина имаат тенденција да се комбинираат преку т.н хидрофобни интеракциии интермолекуларните ван дер Валс сили, густо хидрофобно јадро се формира во внатрешноста на протеинската топка. Хидрофилните јонизирани и нејонизирани радикали главно се наоѓаат на површината на протеинот и ја одредуваат неговата растворливост во вода.
 Ориз. 1.8. Видови врски што се појавуваат помеѓу радикали на аминокиселини за време на формирањето на терциерната структура на протеинот
Ориз. 1.8. Видови врски што се појавуваат помеѓу радикали на аминокиселини за време на формирањето на терциерната структура на протеинот
1 - јонска врска- се јавува помеѓу позитивно и негативно наелектризираните функционални групи;
2 - водородна врска- се јавува помеѓу хидрофилна ненаполнета група и која било друга хидрофилна група;
3 - хидрофобни интеракции- настануваат помеѓу хидрофобните радикали;
4 - дисулфидна врска- формиран поради оксидација на SH групи на остатоци од цистеин и нивната интеракција едни со други
Остатоците од хидрофилни аминокиселини лоцирани во внатрешноста на хидрофобното јадро можат да комуницираат едни со други користејќи јонскиИ водородни врски(Сл. 1.8).
Јонските и водородните врски, како и хидрофобните интеракции, се слаби: нивната енергија не е многу повисока од енергијата на топлинското движење на молекулите на собна температура. Конформацијата на протеинот се одржува со формирање на многу такви слаби врски. Бидејќи атомите што го сочинуваат протеинот се во постојано движење, можно е да се скршат некои слаби врски и да се формираат други, што доведува до мали движења на одделни делови од полипептидниот синџир. Ова својство на протеините да ја менуваат конформацијата како резултат на кршење на некои и формирање на други слаби врски се нарекува конформациска лабилност.
Човечкото тело има системи кои поддржуваат хомеостазата- конзистентност внатрешно опкружувањево одредени прифатливи граници за здрав организам. Во услови на хомеостаза, малите промени во конформацијата не ја нарушуваат целокупната структура и функција на протеините. Функционално активната конформација на протеинот се нарекува родна конформација.Промените во внатрешната средина (на пример, концентрацијата на гликоза, јони на Ca, протони итн.) доведуваат до промени во конформацијата и нарушување на функциите на протеините.
Терциерната структура на некои протеини е стабилизирана дисулфидни врски,формирана поради интеракцијата на -SH групи од два остатоци
 Ориз. 1.9. Формирање на дисулфидна врска во протеинска молекула
Ориз. 1.9. Формирање на дисулфидна врска во протеинска молекула
цистеин (сл. 1.9). Повеќето интрацелуларни протеини немаат ковалентни дисулфидни врски во нивната терциерна структура. Нивното присуство е карактеристично за протеините што ги лачи клетката, што обезбедува нивна поголема стабилност во екстрацелуларните услови. Така, дисулфидните врски се присутни во молекулите на инсулинот и имуноглобулините.
Инсулин- протеински хормон кој се синтетизира во β-клетките на панкреасот и се лачи во крвта како одговор на зголемување на концентрацијата на гликоза во крвта. Во структурата на инсулинот, постојат две дисулфидни врски што ги поврзуваат полипептидните А и Б синџирите и една дисулфидна врска во ланецот А (сл. 1.10).
 Ориз. 1.10. Дисулфидни врски во структурата на инсулинот
Ориз. 1.10. Дисулфидни врски во структурата на инсулинот
5. Суперсекундарна структура на протеините.Во протеините со различна примарна структура и функции, тие понекогаш се откриваат слични комбинации и релативни позиции на секундарните структури,кои се нарекуваат суперсекундарна структура. Тој зазема средна позиција помеѓу секундарните и терциерните структури, бидејќи е специфична комбинација на елементи од секундарната структура во формирањето на терциерната структура на протеинот. Суперсекундарни структури имаат специфични имиња, како што се „α-хеликс-вртење-а-спирала“, „леуцин патент“, „цинк прсти“ итн. Ваквите суперсекундарни структури се карактеристични за протеините што се врзуваат за ДНК.
"Леуцин патент."Овој тип на суперсекундарна структура се користи за спојување на два протеини заедно. На површината на протеините кои се во интеракција има α-спирални региони кои содржат најмалку четири остатоци од леуцин. Остатоците од леуцин во α-спиралата се наоѓаат на шест аминокиселини една од друга. Бидејќи секое вртење на α-спиралата содржи 3,6 остатоци од аминокиселини, радикалите на леуцин се наоѓаат на површината на секое второ вртење. Остатоците од леуцин од α-спиралата на еден протеин можат да комуницираат со остатоците од леуцин на друг протеин (хидрофобни интеракции), поврзувајќи ги заедно (сл. 1.11.). Многу протеини за врзување на ДНК функционираат во олигомерни комплекси каде што поединечните подединици се поврзани една со друга со „леуцински патенти“.
 Ориз. 1.11. „Леуцин патент“ помеѓу α-спирални региони на два протеини
Ориз. 1.11. „Леуцин патент“ помеѓу α-спирални региони на два протеини
Пример за такви протеини се хистоните. Хистони- нуклеарни протеини, кои содржат голем број на позитивно наелектризирани амино киселини - аргинин и лизин (до 80%). Молекулите на хистон се комбинираат во олигомерни комплекси кои содржат осум мономери користејќи „леуцин патенти“, и покрај значителното хомонимно полнење на овие молекули.
„Цинк прст“- варијанта на суперсекундарна структура, карактеристична за протеините што се врзуваат за ДНК, има форма на издолжен фрагмент на површината на протеинот и содржи околу 20 остатоци од аминокиселини (сл. 1.12). Обликот на „продолжен прст“ е поддржан од атом на цинк врзан за четири радикали на аминокиселини - два остатоци од цистеин и два остатоци од хистидин. Во некои случаи, наместо остатоци од хистидин, има остатоци од цистеин. Два тесно поставени остатоци од цистеин се одвоени од другите два остатоци од Gisili со Cys секвенца која се состои од приближно 12 амино киселински остатоци. Овој регион на протеинот формира α-спирала, чии радикали можат конкретно да се врзат за регулаторните региони на главниот жлеб на ДНК. Индивидуална специфичност на врзување
 Ориз. 1.12. Примарната структура на регионот на протеини кои се врзуваат за ДНК кои ја формираат структурата на „цинк прст“ (буквите ги означуваат амино киселините што ја сочинуваат оваа структура)
Ориз. 1.12. Примарната структура на регионот на протеини кои се врзуваат за ДНК кои ја формираат структурата на „цинк прст“ (буквите ги означуваат амино киселините што ја сочинуваат оваа структура)
Регулаторниот протеин за врзување на ДНК зависи од секвенцата на остатоци од аминокиселини лоцирани во регионот на прстите на цинкот. Таквите структури содржат, особено, рецептори за стероидни хормони вклучени во регулирањето на транскрипцијата (читање информации од ДНК до РНК).
ТЕМА 1.2. ОСНОВИ НА ФУНКЦИОНИРАЊЕТО НА ПРОТЕИНИТЕ. ЛЕКОВИ КАКО ЛИГАНДИ КОИ ВЛИЈААТ НА ФУНКЦИЈАТА НА ПРОТЕИНИТЕ
1. Активниот центар на протеинот и неговата интеракција со лигандот.За време на формирањето на терциерната структура, на површината на функционално активен протеин се формира регион, обично во вдлабнатина, формиран од радикали на аминокиселини кои се далеку еден од друг во примарната структура. Овој регион, кој има единствена структура за даден протеин и е способен за специфична интеракција со одредена молекула или група слични молекули, се нарекува место за врзување протеин-лиганд или активно место. Лигандите се молекули кои комуницираат со протеините.
Висока специфичностИнтеракцијата на протеинот со лигандот е обезбедена со комплементарноста на структурата на активниот центар со структурата на лигандот.
Комплементарност- ова е просторна и хемиска кореспонденција на површините во интеракција. Активниот центар не само што мора просторно да одговара на лигандот вклучен во него, туку мора да се формираат и врски (јонски, водородни и хидрофобни интеракции) помеѓу функционалните групи на радикалите вклучени во активниот центар и лигандот, кои го држат лигандот. во активниот центар (сл. 1.13 ).
 Ориз. 1.13. Комплементарна интеракција на протеинот со лигандот
Ориз. 1.13. Комплементарна интеракција на протеинот со лигандот
Некои лиганди, кога се прикачени на активниот центар на протеинот, играат помошна улога во функционирањето на протеините. Таквите лиганди се нарекуваат кофактори, а протеините кои содржат непротеински дел се нарекуваат комплексни протеини(за разлика од едноставните протеини, кои се состојат само од протеинскиот дел). Непротеинскиот дел, цврсто поврзан со протеинот, се нарекува протетска група.На пример, миоглобинот, хемоглобинот и цитохромите содржат протетска група, хем, која содржи железен јон, цврсто прикачен на активниот центар. Комплексните протеини кои содржат хем се нарекуваат хемопротеини.
Кога специфични лиганди се прикачени на протеините, функцијата на овие протеини се манифестира. Така, албуминот, најважниот протеин во крвната плазма, ја покажува својата транспортна функција со прикачување на хидрофобни лиганди, како што се масни киселини, билирубин, некои лекови итн. во активниот центар (сл. 1.14).
Лигандите кои комуницираат со тридимензионалната структура на пептидниот синџир можат да бидат не само нискомолекуларни органски и неоргански молекули, туку и макромолекули:
ДНК (примери со протеини кои се врзуваат за ДНК дискутирани погоре);
Полисахариди;
 Ориз. 1.14. Врска помеѓу генотипот и фенотипот
Ориз. 1.14. Врска помеѓу генотипот и фенотипот
Уникатната примарна структура на човечките протеини, кодирана во молекулата на ДНК, се реализира во клетките во форма на единствена конформација, активна централна структура и протеински функции
Во овие случаи, протеинот препознава специфичен регион на лигандот кој е сразмерен и комплементарен на местото на врзување. Така, на површината на хепатоцитите има рецепторни протеини за хормонот инсулин, кој исто така има протеинска структура. Интеракцијата на инсулинот со рецепторот предизвикува промена во неговата конформација и активирање на сигналните системи, што доведува до складирање на хранливи материи во хепатоцитите после јадење.
Така, Функционирањето на протеините се заснова на специфичната интеракција на активниот центар на протеинот со лигандот.
2. Структура на доменот и неговата улога во функционирањето на протеините.Долгите полипептидни синџири на глобуларни протеини често се преклопуваат во неколку компактни, релативно независни региони. Тие имаат независна терциерна структура, која потсетува на онаа на глобуларните протеини и се нарекуваат домени.Поради структурата на доменот на протеините, полесно се формира нивната терциерна структура.
Во протеините на доменот, местата за врзување на лиганди често се наоѓаат помеѓу домени. Така, трипсинот е протеолитички ензим кој се произведува од егзокриниот дел на панкреасот и е неопходен за варење на протеините од храната. Има структура со два домени, а центарот на врзување на трипсин со неговиот лиганд - протеин на храна - се наоѓа во жлебот помеѓу двата домени. Во активниот центар се создаваат услови неопходни за ефективно врзување на специфично место на протеини од храна и хидролиза на неговите пептидни врски.
Различни домени во протеинот можат да се движат релативно едни на други кога активниот центар е во интеракција со лигандот (сл. 1.15).
Хексокиназа- ензим кој ја катализира фосфорилацијата на гликозата користејќи АТП. Активното место на ензимот се наоѓа во расцепот помеѓу двата домени. Кога хексокиназата се врзува за гликозата, областите што ја опкружуваат се затвораат и супстратот се заглавува, каде што се јавува фосфорилација (види Сл. 1.15).
 Ориз. 1.15. Врзување на домени на хексокиназа со гликоза
Ориз. 1.15. Врзување на домени на хексокиназа со гликоза
Во некои протеини, домените вршат независни функции со врзување за различни лиганди. Таквите протеини се нарекуваат мултифункционални.
3. Лековите се лиганди кои влијаат на функцијата на протеините.Специфична е интеракцијата на протеините со лигандите. Сепак, поради конформациската лабилност на протеинот и неговиот активен центар, можно е да се избере друга супстанција која исто така би можела да комуницира со протеинот во активниот центар или друг дел од молекулата.
Се нарекува супстанца слична по структура на природен лиганд структурен аналог на лигандотили неприроден лиганд. Исто така, комуницира со протеинот на активното место. Структурен аналог на лиганд може и да ја подобри функцијата на протеините (агонист),и намалете го (антагонист).Лигандот и неговите структурни аналози се натпреваруваат едни со други за врзување за протеинот на истото место. Таквите супстанции се нарекуваат конкурентни модулатори(регулатори) на протеинските функции. Многу лекови делуваат како инхибитори на протеини. Некои од нив се добиваат со хемиска модификација на природни лиганди. Инхибитори на протеинските функции може да бидат лекови и отрови.
Атропинот е конкурентен инхибитор на М-холинергичните рецептори.Ацетилхолинот е невротрансмитер за пренос на нервните импулси преку холинергичните синапси. За извршување на возбудувањето, пуштено во синаптичка пукнатинаацетилхолин мора да комуницира со рецепторниот протеин на постсинаптичката мембрана. Најдени се два вида холинергични рецептори:
М рецепторпокрај ацетилхолинот, селективно комуницира со мускаринот (токсин од мушички агарик). М - холинергичните рецептори се присутни на мазните мускули и, при интеракција со ацетилхолин, предизвикуваат нивна контракција;
H рецепторспецифично врзување за никотинот. Н-холинергичните рецептори се наоѓаат во синапсите на напречно-пругастите скелетни мускули.
Специфичен инхибитор М-холинергични рецепторие атропин. Го има во растенијата беладона и кокошката.
 Атропинот има функционални групи слични во структурата на ацетилхолинот и нивниот просторен распоред, затоа е конкурентен инхибитор на М-холинергичните рецептори. Имајќи предвид дека врзувањето на ацетилхолин за М-холинергичните рецептори предизвикува контракција на мазните мускули, атропинот се користи како лек кој го ублажува нивниот спазам. (антиспазмодичен).Така, познато е да се користи атропин за опуштање на очните мускули при гледање на фундусот, како и за ублажување на грчеви за време на гастроинтестинална колика. М-холинергичните рецептори се исто така присутни во централниот нервен систем(ЦНС), затоа големите дози на атропин може да предизвикаат несакана реакција од централниот нервен систем: моторна и ментална агитација, халуцинации, конвулзии.
Атропинот има функционални групи слични во структурата на ацетилхолинот и нивниот просторен распоред, затоа е конкурентен инхибитор на М-холинергичните рецептори. Имајќи предвид дека врзувањето на ацетилхолин за М-холинергичните рецептори предизвикува контракција на мазните мускули, атропинот се користи како лек кој го ублажува нивниот спазам. (антиспазмодичен).Така, познато е да се користи атропин за опуштање на очните мускули при гледање на фундусот, како и за ублажување на грчеви за време на гастроинтестинална колика. М-холинергичните рецептори се исто така присутни во централниот нервен систем(ЦНС), затоа големите дози на атропин може да предизвикаат несакана реакција од централниот нервен систем: моторна и ментална агитација, халуцинации, конвулзии.
Дитилин е конкурентен агонист на H-холинергичните рецептори, инхибирајќи ја функцијата на невромускулните синапси.
 Невромускулните синапси на скелетните мускули содржат H-холинергични рецептори. Нивната интеракција со ацетилхолин доведува до мускулни контракции. За време на некои хируршки операции, како и во ендоскопски студии, се користат лекови кои предизвикуваат релаксација на скелетните мускули (мускулни релаксанти).Тие вклучуваат дитилин, кој е структурен аналог на ацетилхолин. Се прицврстува на H-холинергичните рецептори, но за разлика од ацетилхолинот, тој многу бавно се уништува од ензимот ацетилхолинестераза. Како резултат на продолженото отворање на јонските канали и постојаната деполаризација на мембраната, се нарушува спроводливоста на нервните импулси и се јавува мускулна релаксација. Првично, овие својства биле откриени во отровот кураре, поради што таквите лекови се нарекуваат кураре-како.
Невромускулните синапси на скелетните мускули содржат H-холинергични рецептори. Нивната интеракција со ацетилхолин доведува до мускулни контракции. За време на некои хируршки операции, како и во ендоскопски студии, се користат лекови кои предизвикуваат релаксација на скелетните мускули (мускулни релаксанти).Тие вклучуваат дитилин, кој е структурен аналог на ацетилхолин. Се прицврстува на H-холинергичните рецептори, но за разлика од ацетилхолинот, тој многу бавно се уништува од ензимот ацетилхолинестераза. Како резултат на продолженото отворање на јонските канали и постојаната деполаризација на мембраната, се нарушува спроводливоста на нервните импулси и се јавува мускулна релаксација. Првично, овие својства биле откриени во отровот кураре, поради што таквите лекови се нарекуваат кураре-како.
ТЕМА 1.3. ДЕНАТУРАЦИЈА НА ПРОТЕИНИ И МОЖНОСТ ЗА НИВНА СПОНТАНА РЕНАТИВАЦИЈА
1. Бидејќи природната конформација на протеините се одржува поради слабите интеракции, промените во составот и својствата на околината што го опкружува протеинот, изложеноста на хемиски реагенси и физички фактори предизвикуваат промена во нивната конформација (својството на конформациска лабилност). Јаз големо количествоврските доведуваат до уништување на природната конформација и денатурација на протеините.
Денатурација на протеини- ова е уништување на нивната матична конформација под влијание на денатурирачки агенси, предизвикано од кинење на слабите врски кои ја стабилизираат просторната структура на протеинот. Денатурацијата е придружена со уништување на уникатната тридимензионална структура и активниот центар на протеинот и губење на неговата биолошка активност (сл. 1.16).
Сите денатурирани молекули на еден протеин добиваат случајна конформација која се разликува од другите молекули на истиот протеин. Аминокиселинските радикали кои го формираат активниот центар се испостави дека се просторно оддалечени еден од друг, т.е. специфичното место за врзување на протеинот со лигандот е уништено. За време на денатурацијата, примарната структура на протеините останува непроменета.
Примена на денатурирачки агенси во биолошките истражувања и медицината.ВО биохемиски истражувањаПред да се одредат соединенијата со мала молекуларна тежина во биолошкиот материјал, протеините обично прво се отстрануваат од растворот. За таа цел најчесто се користи трихлороцетна киселина (TCA). По додавањето на TCA во растворот, денатурираните протеини се таложат и лесно се отстрануваат со филтрација (Табела 1.1.)
Во медицината, средствата за денатурирање често се користат за стерилизирање на медицински инструменти и материјали во автоклави (средството за денатурација е висока температура) и како антисептици (алкохол, фенол, хлорамин) за третирање на контаминирани површини кои содржат патогена микрофлора.
2. Спонтана реактивација на протеини- доказ за детерминизмот на примарната структура, конформацијата и функцијата на протеините. Индивидуалните протеини се производи на еден ген кои имаат идентична аминокиселинска секвенца и ја добиваат истата конформација во клетката. Фундаменталниот заклучок дека примарната структура на протеинот веќе содржи информации за неговата конформација и функција е направен врз основа на способноста на некои протеини (особено, рибонуклеазата и миоглобинот) спонтано да се обновуваат - да ја вратат нивната родна конформација по денатурацијата.
Формирањето на просторни протеински структури се врши со методот на самосклопување - спонтан процес во кој полипептидниот синџир, кој има единствена примарна структура, има тенденција да усвои конформација со најниска слободна енергија во растворот. Способноста да се обноват протеините кои ја задржуваат својата примарна структура по денатурацијата беше опишана во експеримент со ензимот рибонуклеаза.
Рибонуклеазата е ензим кој ги разградува врските помеѓу поединечните нуклеотиди во молекулата на РНК. Овој топчест протеин има еден полипептиден синџир, чија терцијарна структура е стабилизирана со многу слаби и четири дисулфидни врски.
Третманот на рибонуклеазата со уреа, која ги раскинува водородните врски во молекулата, и средство за намалување, кое ги раскинува дисулфидните врски, доведува до денатурација на ензимот и губење на неговата активност.
Отстранувањето на денатурирачките агенси со дијализа доведува до обновување на конформацијата и функцијата на протеините, т.е. до повторно раѓање. (Сл. 1.17).
 Ориз. 1.17. Денатурација и ренативација на рибонуклеазата
Ориз. 1.17. Денатурација и ренативација на рибонуклеазата
А - природна конформација на рибонуклеаза, во терциерната структура на која има четири дисулфидни врски; Б - денатурирана молекула на рибонуклеаза;
Б - реактивирана молекула на рибонуклеаза со обновена структура и функција
1. Пополнете ја табелата 1.2.
Табела 1.2. Класификација на аминокиселините според поларитетот на радикалите
2. Напишете ја формулата на тетрапептидот:
Асп - Про - Фен - Лиз
а) означете ги повторувачките групи во пептидот што го формираат пептидниот столб и променливите групи претставени со радикали на аминокиселини;
б) означете ги N- и C-крајните;
в) истакнете ги пептидните врски;
г) напишете друг пептид кој се состои од исти амино киселини;
д) брои го бројот на можни варијанти на тетрапептид со сличен состав на аминокиселини.
3. Објаснете ја улогата на примарната структура на протеините користејќи го примерот на компаративна анализа на два структурно слични и еволутивно блиски пептидни хормони на неврохипофизата на цицачите - окситоцин и вазопресин (Табела 1.3).
Табела 1.3. Структура и функции на окситоцин и вазопресин
 За ова:
За ова:
а) споредете го составот и аминокиселинската секвенца на два пептида;
б) да ја најде сличноста на примарната структура на двата пептида и сличноста на нивното биолошко дејство;
в) најдете разлики во структурата на два пептида и разлики во нивните функции;
г) да донесе заклучок за влијанието на примарната структура на пептидите врз нивните функции.
4. Опишете ги главните фази на формирање на конформацијата на глобуларни протеини (секундарни, терциерни структури, концептот на суперсекундарна структура). Наведете ги видовите на врски вклучени во формирањето на протеинските структури. Кои аминокиселински радикали можат да учествуваат во формирањето на хидрофобни интеракции, јонски, водородни врски.
Наведи примери.
5. Дефинирајте го концептот на „конформациска лабилност на протеините“, наведете ги причините за неговото постоење и значење.
6. Проширете го значењето на следната фраза: „Функционирањето на протеините се заснова на нивната специфична интеракција со лигандот“, користејќи ги термините и објаснувајќи го нивното значење: конформација на протеини, активен центар, лиганд, комплементарност, протеинска функција.
7. Користејќи еден пример, објаснете што се домени и која е нивната улога во функционирањето на протеините.
ЗАДАЧИ ЗА САМОКОНТРОЛА
1. Натпревар.
Функционална група во аминокиселинскиот радикал:
А. Карбоксилна група Б. Хидроксилна група Ц Гванидинска група Д. Тиол група Е. Амино група
2. Изберете ги точните одговори.
Амино киселините со поларни ненаполнети радикали се:
A. Cis B. Asn
Б. Глу Г. Три
3. Изберете ги точните одговори.
Аминокиселински радикали:
А. Обезбедете специфичност на примарната структура Б. Учествувајте во формирањето на терциерната структура
Б. Сместени на површината на протеинот, тие влијаат на неговата растворливост D. Формираат активниот центар
D. Учествуваат во формирањето на пептидни врски
4. Изберете ги точните одговори.
Може да се формираат хидрофобни интеракции помеѓу аминокиселинските радикали:
А. Тре Леј Б. Про Три
Б. Мет Иле Г. Тир Ала Д. Вал Фен
5. Изберете ги точните одговори.
Јонските врски можат да се формираат помеѓу аминокиселинските радикали:
А. Глн Асп Б. Апр Лиз
B. Liz Glu G. Gis Asp D. Asn Apr
6. Изберете ги точните одговори.
Водородните врски можат да се формираат помеѓу радикали на аминокиселини:
А. Сер Глн Б. Цис Тре
Б. Асп Лиз Г. Глу Асп Д. Асн Тре
7. Натпревар.
Вид на врска вклучена во формирањето на протеинската структура:
А. Примарна структура Б. Секундарна структура
Б. Терцијарна структура
D. Суперсекундарна структура E. Конформација.
1. Водородни врски помеѓу атомите на пептидниот столб
2. Слаби врски помеѓу функционалните групи на аминокиселински радикали
3. Врски помеѓу α-амино и α-карбоксилни групи на амино киселини
8. Изберете ги точните одговори. Трипсин:
A. Протеолитички ензим B. Содржи два домени
Б. Хидролизира скроб
D. Активната локација се наоѓа помеѓу домените. D. Се состои од два полипептидни синџири.
9. Изберете ги точните одговори. Атропин:
A. Невротрансмитер
Б. Структурен аналог на ацетилхолин
B. Интеракции со H-холинергични рецептори
D. Ја зајакнува спроводливоста на нервните импулси преку холинергичните синапси
D. Конкурентен инхибитор на М-холинергичните рецептори
10. Изберете ги точните изјави. Во протеините:
A. Примарната структура содржи информации за структурата на нејзиното активно место
Б. Активниот центар се формира на ниво на примарната структура
Б. Конформацијата е цврсто фиксирана со ковалентни врски
D. Активното место може да комуницира со група слични лиганди
поради конформациската лабилност на протеините D. Промена животната средина, може да влијае на афинитетот на активната
центар до лиганд
1. 1-Б, 2-Г, 3-Б.
3. А, Б, Ц, Д.
7. 1-Б, 2-Д, 3-А.
8. А, Б, Ц, Д.
ОСНОВНИ ПОИМИ И ПОИМИ
1. Протеини, полипептиди, амино киселини
2. Примарни, секундарни, терциерни протеински структури
3. Конформација, конформација на мајчин протеин
4. Ковалентни и слаби врски во протеините
5. Конформациска лабилност
6. Протеинско активно место
7. Лиганди
8. Преклопување на протеини
9. Структурни аналози на лиганди
10. Протеини од домен
11. Едноставни и сложени протеини
12. Протеинска денатурација, денатурирачки агенси
13. Реактивирање на протеините
Решавајте проблеми
„Структурна организација на протеините и основата на нивното функционирање“
1. Главната функција на протеинот - хемоглобинот А (HbA) е транспортот на кислород до ткивата. Кај човечката популација познати се повеќе форми на овој протеин со изменети својства и функција - таканаречените абнормални хемоглобини. На пример, хемоглобинот S, кој се наоѓа во црвените крвни зрнца на пациенти со српеста анемија (HbS), е откриено дека има ниска растворливост во услови на низок парцијален притисок на кислород (како што е случајот со венската крв). Ова доведува до формирање на агрегати на овој протеин. Протеинот ја губи својата функција, се таложи и црвените крвни зрнца се стекнуваат неправилна форма(некои од нив формираат срп форма) и се уништуваат побрзо од вообичаеното во слезината. Како резултат на тоа, се развива српеста анемија.
Единствената разлика во примарната структура на HbA беше пронајдена во N-терминалниот регион на β-синџирот на хемоглобинот. Споредете ги N-терминалните региони на β-влакното и покажете како промените во примарната структура на протеинот влијаат на неговите својства и функции.
 За ова:
За ова:
а) напишете ги формулите на амино киселините по кои HbA се разликува и споредете ги својствата на овие амино киселини (поларитет, полнеж).
б) донесе заклучок за причината за намалување на растворливоста и нарушување на транспортот на кислород во ткивата.
2. На сликата е прикажан дијаграм на структурата на протеин кој има сврзувачки центар со лиганд (активен центар). Објаснете зошто протеинот е селективен во изборот на лиганд. За ова:
а) запомнете што е активниот центар на протеинот и разгледајте ја структурата на активниот центар на протеинот прикажан на сликата;
б) напиши ги формулите на радикалите на аминокиселините кои го сочинуваат активниот центар;
в) нацртајте лиганд кој би можел конкретно да комуницира со активното место на протеинот. Наведете ги на неа функционалните групи кои можат да формираат врски со радикалите на аминокиселините што го сочинуваат активниот центар;
г) наведете ги видовите врски што се појавуваат помеѓу лигандот и радикалите на аминокиселините на активниот центар;
д) објасни на што се заснова специфичноста на интеракцијата протеин-лиганд.
 3.
Сликата го прикажува активното место на протеинот и неколку лиганди.
3.
Сликата го прикажува активното место на протеинот и неколку лиганди.
Определете кој лиганд има најголема веројатност да комуницира со активното место на протеинот и зошто.
 Какви видови врски се јавуваат при формирањето на протеинско-лигандскиот комплекс?
Какви видови врски се јавуваат при формирањето на протеинско-лигандскиот комплекс?
4. Структурните аналози на природните протеински лиганди може да се користат како лекови за менување на активноста на протеините.
Ацетилхолинот е посредник за пренос на возбуда во невромускулните синапси. Кога ацетилхолин е во интеракција со протеини - рецептори на постсинаптичката мембрана на скелетните мускули, се отвораат јонските канали и се јавува контракција на мускулите. Дитилин е лек кој се користи во некои операции за опуштање на мускулите, бидејќи го нарушува преносот на нервните импулси преку невромускулните синапси. Објаснете го механизмот на дејство на дитилин како мускулен релаксант. За ова:
а) напишете ги формулите на ацетилхолин и дитилин и споредете ги нивните структури;
б) опишете го механизмот на релаксирачки ефект на дитилин.
5. Кај некои болести, температурата на телото на пациентот се зголемува, што се смета како заштитна реакција на телото. Сепак, високите температури се штетни за телесните протеини. Објаснете зошто на температури над 40 °C се нарушува функцијата на протеините и се јавува закана за човечкиот живот. За да го направите ова, запомнете:
1) Структурата на протеините и врските што ја држат нејзината структура во природната конформација;
2) Како се менува структурата и функцијата на протеините со зголемување на температурата?;
3) Што е хомеостаза и зошто е важна за одржување на здравјето на луѓето.
Модуларна единица 2 ОЛИГОМЕРИЧКИ ПРОТЕИНИ КАКО ЦЕЛИ НА РЕГУЛАТОРНИ ВЛИЈАНИЕ. СТРУКТУРНА И ФУНКЦИОНАЛНА РАЗЛИЧНОСТ НА ПРОТЕИНИ. МЕТОДИ ЗА РАЗДЕЛУВАЊЕ И ПРОЧИСТУВАЊЕ НА ПРОТЕИНИ
Цели на учењето Бидете способни:
1. Користете знаење за карактеристиките на структурата и функциите на олигомерните протеини за да ги разберете адаптивните механизми на регулирање на нивните функции.
2. Објаснете ја улогата на чаперони во синтезата и одржувањето на конформацијата на протеините во клеточни услови.
3. Објаснете ја разновидноста на манифестациите на животот со разновидноста на структурите и функциите на протеините синтетизирани во телото.
4. Анализирајте го односот помеѓу структурата на протеините и нивната функција користејќи примери за споредба на сродни хемопротеини - миоглобин и хемоглобин, како и претставници на петте класи на протеини од семејството на имуноглобулини.
5. Применуваат знаења за особеностите на физичките и хемиските својства на протеините за избор на методи за нивно прочистување од други протеини и нечистотии.
6. Толкувајте ги резултатите од квантитативните и квалитетен составпротеините на крвната плазма за да се потврди или разјасни клиничката дијагноза.
Знајте:
1. Карактеристики на структурата на олигомерните протеини и адаптивни механизми за регулирање на нивните функции користејќи го примерот на хемоглобинот.
2. Структурата и функциите на чапероните и нивната важност за одржување на природната конформација на протеините во клеточни услови.
3. Принципи на комбинирање на протеините во семејства врз основа на сличноста на нивната конформација и функции користејќи го примерот на имуноглобулините.
4. Методи за одвојување на протеините врз основа на карактеристиките на нивните физичко-хемиски својства.
5. Електрофореза на крвната плазма како метод за проценка на квалитативниот и квантитативниот состав на протеините.
ТЕМА 1.4. КАРАКТЕРИСТИКИ НА СТРУКТУРАТА И ФУНКЦИОНИРАЊЕТО НА ОЛИГОМЕРНИ ПРОТЕИНИ КОРИСТЕТЕ ГО ПРИМЕРОТ НА ХЕМОГЛОБИН
1. Многу протеини содржат неколку полипептидни синџири. Таквите протеини се нарекуваат олигомерни,и индивидуални синџири - протомери.Протомерите во олигомерните протеини се поврзани со многу слаби нековалентни врски (хидрофобни, јонски, водородни). Интеракција
протомери се вршат благодарение на комплементарностнивните контактни површини.
Бројот на протомери во олигомерните протеини може многу да варира: хемоглобинот содржи 4 протомери, ензимот аспартат аминотрансфераза има 12 протомери, а протеинот од вирусот на мозаикот од тутун содржи 2120 протомери поврзани со нековалентни врски. Следствено, олигомерните протеини може да имаат многу висока молекуларна тежина.
Интеракцијата на еден протомер со други може да се смета како посебен случајпротеин-лиганд интеракции, бидејќи секој протомер служи како лиганд за други протомери. Бројот и начинот на спојување на протомери во протеин се нарекуваат кватернарна протеинска структура.
Протеините можат да содржат протомери со иста или различна структура, на пример, хомодимерите се протеини кои содржат два идентични протомери, а хетеродимерите се протеини кои содржат два различни протомери.
Ако протеините содржат различни протомери, тогаш на нив може да се формираат сврзувачки центри со различни лиганди кои се разликуваат по структура. Кога лигандот се врзува за активното место, функцијата на овој протеин се манифестира. Центарот лоциран на различен протомер се нарекува алостеричен (различен од активниот). Контактирање алостеричен лиганд или ефектор,врши регулаторна функција (сл. 1.18). Интеракцијата на алостеричниот центар со ефекторот предизвикува конформациски промени во структурата на целиот олигомерен протеин поради неговата конформациска лабилност. Ова влијае на афинитетот на активното место за специфичен лиганд и ја регулира функцијата на тој протеин. Промената во конформацијата и функцијата на сите протомери за време на интеракцијата на олигомерниот протеин со најмалку еден лиганд се нарекуваат кооперативни конформациски промени. Ефекторите кои ја подобруваат протеинската функција се нарекуваат активатори,и ефектори кои ја инхибираат неговата функција - инхибитори.
Така, олигомерните протеини, како и протеините со структура на домен, имаат ново својство во споредба со мономерните протеини - способност за алостерично регулирање на функциите (регулација со прикачување на различни лиганди на протеинот). Ова може да се види со споредување на структурите и функциите на два тесно поврзани комплексни протеини, миоглобинот и хемоглобинот.
 Ориз. 1.18. Шема на структурата на димерен протеин
Ориз. 1.18. Шема на структурата на димерен протеин
2. Формирање на просторни структури и функционирање на миоглобинот.
Миоглобинот (Mb) е протеин кој се наоѓа во црвените мускули, чија главна функција е создавање резерви на О 2 неопходни за интензивна мускулна работа. Mb е комплексен протеин кој содржи протеински дел - apoMb и непротеински дел - хем. Примарната структура на apoMB ја одредува нејзината компактна топчеста конформација и структурата на активниот центар, на кој е прикачен непротеинскиот дел од миоглобинот, хем. Кислородот што доаѓа од крвта до мускулите се врзува за Fe+2 хемите во миоглобинот. Mb е мономерен протеин кој има многу висок афинитет за O 2, затоа ослободувањето на кислород од миоглобинот се случува само при интензивна мускулна работа, кога парцијалниот притисок на O 2 нагло се намалува.
Формирање на конформацијата Mv.Во црвените мускули, на рибозомите за време на транслацијата, се синтетизира примарната структура на МБ, претставена со специфична низа од 153 аминокиселински остатоци. Секундарната структура на Mb содржи осум α-спирали, наречени со латински букви од A до H, меѓу кои има неспирални региони. Терциерната структура на Mb има форма на компактна топка, во чија вдлабнатина се наоѓа активниот центар помеѓу F и E α-спиралите (сл. 1.19).
 Ориз. 1.19. Структура на миоглобинот
Ориз. 1.19. Структура на миоглобинот
3. Карактеристики на структурата и функционирањето на активниот центар на МВ.Активниот центар на Mb е формиран претежно од хидрофобни аминокиселински радикали, широко распоредени еден од друг во примарната структура (на пример, Tri 3 9 и Фен 138) Слабо растворливи во вода лиганди - хем и О 2 - се прикачуваат на активниот центар. Хемот е специфичен лиганд на apoMB (сл. 1.20), чија основа е составена од четири пиролски прстени поврзани со метенилни мостови; во центарот има атом Fe+ 2 поврзан со атомите на азот на пиролските прстени со четири координативни врски. Во активниот центар на Mb, покрај хидрофобните аминокиселински радикали, има и остатоци од две амино киселини со хидрофилни радикали - Гис Е 7(Gis 64) и ГИС Ф 8(Неговиот 93) (Сл. 1.21).
 Ориз. 1.20. Структурата на хем - не-протеинскиот дел од миоглобинот и хемоглобинот
Ориз. 1.20. Структурата на хем - не-протеинскиот дел од миоглобинот и хемоглобинот
 Ориз. 1.21. Локација на хем и О2 во активното место на апомиоглобинот и хемоглобинските протомери
Ориз. 1.21. Локација на хем и О2 во активното место на апомиоглобинот и хемоглобинските протомери
Хемот е ковалентно врзан за неговиот F8 преку атом на железо. O 2 се прицврстува на железото од другата страна на рамнината на хем. Неговиот Е 7 е неопходен за правилна ориентација на О 2 и го олеснува додавањето на кислород во Fe + 2 хем
ГИС Ф 8формира координативна врска со Fe+ 2 и цврсто го фиксира хемот во активниот центар. Гис Е 7неопходен за правилна ориентација во активниот центар на друг лиганд - O 2 за време на неговата интеракција со Fe + 2 хем. Микросредината на хем создава услови за силно, но реверзибилно врзување на O 2 со Fe + 2 и ја спречува водата да навлезе во хидрофобното активно место, што може да доведе до негова оксидација до Fe + 3.
Мономерната структура на Mb и неговиот активен центар го одредува високиот афинитет на протеинот за O 2.
4. Олигомерна структура на Hb и регулација на афинитетот на Hb кон O 2 лиганди. Човечки хемоглобини- семејство на протеини, како миоглобинот, поврзани со комплексни протеини (хемопротеини). Тие имаат тетрамерна структура и содржат два α-синџири, но се разликуваат во структурата на другите два полипептидни синџири (2α-, 2x-синџири). Структурата на вториот полипептиден синџир ги одредува карактеристиките на функционирањето на овие форми на Hb. Околу 98% од хемоглобинот во црвените крвни зрнца на возрасен е хемоглобин А(2α-, 2p-синџири).
За време на развојот на фетусот, функционираат два главни типа на хемоглобини: ембрионски Hb(2α, 2ε), кој се наоѓа на раните фазифетален развој и хемоглобин F (фетален)- (2α, 2γ), кој го заменува раниот фетален хемоглобин во шестиот месец од интраутериниот развој и дури по раѓањето се заменува со Hb A.
HB A е протеин поврзан со миоглобинот (MB) кој се наоѓа во црвените крвни зрнца на возрасен човек. Структурата на неговите поединечни протомери е слична на онаа на миоглобинот. Секундарните и терциерните структури на протомерите на миоглобинот и хемоглобинот се многу слични, и покрај фактот што во примарната структура на нивните полипептидни синџири се идентични само 24 амино киселински остатоци (секундарната структура на протомерите на хемоглобин, како миоглобинот, содржи осум α-спирали, означени со латински букви од A до H, а терциерната структура има форма на компактна топка). Но, за разлика од миоглобинот, хемоглобинот има олигомерна структура, која се состои од четири полипептидни синџири поврзани со не-ковалентни врски (Слика 1.22).
Секој протомер на Hb е поврзан со непротеински дел - хем и соседни протомери. Врската на протеинскиот дел на Hb со хем е слична на онаа на миоглобинот: во активниот центар на протеинот, хидрофобните делови на хемот се опкружени со хидрофобни аминокиселински радикали со исклучок на His F 8 и His E 7, кои се наоѓаат на двете страни на рамнината на хем и играат слична улога во функционирањето на протеинот и неговото врзување со кислородот (види структура на миоглобинот).
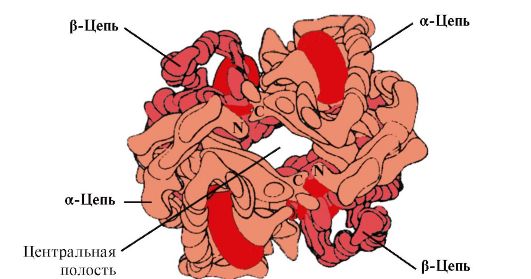 Ориз. 1.22. Олигомерна структура на хемоглобинот
Ориз. 1.22. Олигомерна структура на хемоглобинот
Освен тоа, Гис Е 7врши важна дополнителна улогаво функционирањето на Нв. Слободниот хем има 25.000 пати поголем афинитет за CO отколку за O2. CO се формира во мали количини во телото и, со оглед на неговиот висок афинитет за хем, може да го наруши транспортот на O 2 неопходен за живот на клетките. Меѓутоа, во составот на хемоглобинот, афинитетот на хем за јаглерод моноксид го надминува афинитетот за O 2 само за 200 пати поради присуството на His E 7 во активниот центар. Остатокот од оваа аминокиселина создава оптимални услови за врзување на хем со O 2 и ја ослабува интеракцијата на хем со CO.
5. Главната функција на HB е транспортот на О2 од белите дробови до ткивата.За разлика од мономерниот миоглобин, кој има многу висок афинитет за О2 и ја врши функцијата на складирање на кислород во црвените мускули, олигомерната структура на хемоглобинот обезбедува:
1) брза заситеност на HB со кислород во белите дробови;
2) способноста на HB да ослободува кислород во ткивата при релативно висок парцијален притисок од O 2 (20-40 mm Hg);
3) можност за регулирање на афинитетот на Hb кон O 2.
6. Кооперативните промени во конформацијата на протомерите на хемоглобин го забрзуваат врзувањето на O 2 во белите дробови и неговото ослободување во ткивата. Во белите дробови, високиот парцијален притисок на O 2 го промовира неговото врзување за Hb во активното место на четири протомери (2α и 2β). Активниот центар на секој протомер, како и кај миоглобинот, се наоѓа помеѓу два α-спирали (F и E) во хидрофобен џеб. Содржи непротеински дел - хем, прикачен на протеинскиот дел со многу слаби хидрофобни интеракции и една силна врска помеѓу Fe 2 + хем и His F 8 (види Сл. 1.21).
Во деоксихемоглобинот, поради оваа врска со His F 8, атомот Fe 2 + штрчи од рамнината на хем кон хистидин. Врзувањето на O 2 со Fe 2 + се случува на другата страна на хемот во регионот His E 7 со помош на единечна слободна координативна врска. Неговиот Е 7 обезбедува оптимални услови за врзување на О 2 со хем железото.
Додавањето на O 2 во атомот Fe + 2 на еден протомер предизвикува негово движење во рамнината на хем, проследено со остаток од хистидин поврзан со него
 Ориз. 1.23. Промена на конформацијата на протомерот на хемоглобин кога се комбинира со O 2
Ориз. 1.23. Промена на конформацијата на протомерот на хемоглобин кога се комбинира со O 2
Ова доведува до промена во конформацијата на сите полипептидни синџири поради нивната конформациска лабилност. Промената на конформацијата на другите синџири ја олеснува нивната интеракција со следните молекули на O 2.
Четвртата молекула O 2 се прицврстува на хемоглобинот 300 пати полесно од првата (сл. 1.24).
 Ориз. 1.24. Кооперативни промени во конформацијата на протомери на хемоглобин за време на неговата интеракција со О2
Ориз. 1.24. Кооперативни промени во конформацијата на протомери на хемоглобин за време на неговата интеракција со О2
Во ткивата, секоја следна молекула на O2 се отцепува полесно од претходната, исто така поради кооперативните промени во конформацијата на протомерите.
7. CO 2 и H+ формирани за време на катаболизмот органска материја, го намалуваат афинитетот на хемоглобинот за O 2 пропорционално на нивната концентрација. Енергијата потребна за функционирање на клетките се произведува првенствено во митохондриите за време на оксидацијата на органските материи со користење на O 2 испорачан од белите дробови преку хемоглобинот. Како резултат на оксидацијата на органските материи, се формираат финалните производи од нивното распаѓање: CO 2 и K 2 O, чија количина е пропорционална на интензитетот на тековните процеси на оксидација.
CO 2 дифундира од клетките во крвта и продира во црвените крвни зрнца, каде што, под дејство на ензимот карбанхидраза, се претвора во јаглеродна киселина. Оваа слаба киселина се дисоцира во протон и бикарбонат јон.
H+ се способни да им се придружат на Неговите радикали 14 6 во α- и β-синџирите на хемоглобинот, т.е. во области оддалечени од хем. Протонацијата на хемоглобинот го намалува неговиот афинитет за O 2, го промовира отстранувањето на O 2 од oxyHb, формирањето на deoxyHb и го зголемува снабдувањето со кислород до ткивата пропорционално на бројот на формирани протони (сл. 1.25).
Зголемувањето на количината на ослободениот кислород во зависност од зголемувањето на концентрацијата на H+ во црвените крвни зрнца се нарекува Боров ефект (именуван по данскиот физиолог Кристијан Бор, кој прв го открил овој ефект).
Во белите дробови, високиот парцијален притисок на кислород го промовира неговото врзување за деокси Hb, што го намалува афинитетот на протеинот за H +. Ослободените протони под дејство на јаглеродна киселина реагираат со бикарбонати за да формираат CO 2 и H 2 O

 Ориз. 1.25. Зависност на афинитетот на Hb за O 2 од концентрацијата на CO 2 и протоните (Боров ефект):
Ориз. 1.25. Зависност на афинитетот на Hb за O 2 од концентрацијата на CO 2 и протоните (Боров ефект):
А- влијание на концентрацијата на CO 2 и H+ врз ослободувањето на O 2 од комплексот со HB (Боров ефект); Б- оксигенација на деоксихемоглобин во белите дробови, формирање и ослободување на CO 2.
Добиениот CO 2 влегува во алвеоларниот простор и се отстранува со издишаниот воздух. Така, количината на кислород што се ослободува од хемоглобинот во ткивата е регулирана со производите на катаболизмот на органските супстанции: колку е поинтензивно разградувањето на супстанциите, на пример за време на физички вежби, толку е поголема концентрацијата на CO 2 и H + и толку повеќе кислород. ткивата добиваат како резултат на намалување на афинитетот на Hb за O 2.
8. Алостерична регулација на афинитетот на Hb за O2 од страна на лигандот - 2,3-бисфосфоглицерат.Во еритроцитите, алостеричниот лиганд на хемоглобинот, 2,3-бисфосфоглицерат (2,3-BPG), се синтетизира од производот на оксидација на гликоза - 1,3-бифосфоглицерат. Во нормални услови, концентрацијата на 2,3-BPG е висока и споредлива со концентрацијата на Hb. 2,3-BPG има силен негативен полнеж од -5.
 Бифосфоглицератот во ткивните капилари, врзувајќи се за деоксихемоглобинот, го зголемува ослободувањето на кислород во ткивата, намалувајќи го афинитетот на Hb за O2.
Бифосфоглицератот во ткивните капилари, врзувајќи се за деоксихемоглобинот, го зголемува ослободувањето на кислород во ткивата, намалувајќи го афинитетот на Hb за O2.
Во центарот на тетрамерната молекула на хемоглобинот е шуплина. Се формира од остатоци од аминокиселини на сите четири протомери (види Сл. 1.22). Во ткивните капилари, протонацијата на Hb (Боров ефект) доведува до прекин на врската помеѓу хем железото и О2. Во молекула
деоксихемоглобин во споредба со оксихемоглобин има дополнителни јонски врски, поврзување на протомери, како резултат на што димензиите на централната празнина се зголемуваат во споредба со оксихемоглобинот. Централната празнина е местото на прицврстување на 2,3-BPG на хемоглобинот. Поради разликата во големината на централната празнина, 2,3-BPG може да се прикачи само на деоксихемоглобинот.
2,3-BPG комуницира со хемоглобинот во место оддалечено од активните центри на протеинот и припаѓа на алостерична(регулаторни) лиганди, а централната празнина на Hb е алостеричен центар. 2,3-BPG има силен негативен полнеж и е во интеракција со пет позитивно наелектризирани групи од двата β-синџири на Hb: N-терминалната α-амино група на Val и Lys 82 His 143 радикали (сл. 1.26).
 Ориз. 1.26. BPG во централната празнина на деоксихемоглобинот
Ориз. 1.26. BPG во централната празнина на деоксихемоглобинот
BPG се врзува за три позитивно наелектризирани групи на секоја β-влакна.
Во ткивните капилари, добиениот деоксихемоглобин е во интеракција со 2,3-BPG и се формираат јонски врски помеѓу позитивно наелектризираните радикали на β-синџирите и негативно наелектризираниот лиганд, кои ја менуваат конформацијата на протеинот и го намалуваат афинитетот на Hb за O2. . Намалувањето на афинитетот на Hb за O 2 придонесува за поефикасно ослободување на O 2 во ткивото.
Во белите дробови, при висок парцијален притисок, кислородот комуницира со Hb, спојувајќи го хем железото; во овој случај, конформацијата на протеинот се менува, централната празнина се намалува и 2,3-BPG се поместува од алостеричниот центар
Така, олигомерните протеини имаат нови својства во споредба со мономерните протеини. Прицврстување на лиганди на места
просторно оддалечени едни од други (алостерични), може да предизвикаат конформациски промени во целата протеинска молекула. Поради интеракцијата со регулаторните лиганди, се јавува промена во конформацијата и адаптација на функцијата на протеинската молекула на промените на животната средина.
ТЕМА 1.5. ОДРЖУВАЊЕ НА МАТИЧНА КОНФОРМАЦИЈА НА ПРОТЕИНИ ВО КЛЕТНИ УСЛОВИ
Во клетките, при синтезата на полипептидните синџири, нивниот транспорт преку мембраните до соодветните делови на клетката, при процесот на превиткување (формирање на природната конформација) и при склопување на олигомерните протеини, како и при нивното функционирање, меѓу , во структурата на протеините се појавуваат нестабилни конформации склони кон агрегација. Хидрофобните радикали, обично скриени во протеинската молекула во природната конформација, се појавуваат на површината во нестабилна конформација и имаат тенденција да се комбинираат со групи други протеини кои се слабо растворливи во вода. Во клетките на сите познати организми, откриени се специјални протеини кои обезбедуваат оптимално превиткување на клеточните протеини, ја стабилизираат нивната природна конформација за време на функционирањето и што е најважно, ја одржуваат структурата и функциите на интрацелуларните протеини кога е нарушена хомеостазата. Овие протеини се нарекуваат "сопственици"што значи „дадилка“ на француски.
1. Молекуларни шаперони и нивната улога во спречувањето на денатурација на протеините.
Шаперони (CH) се класифицираат според масата на нивните подединици. Шаперони со висока молекуларна тежина имаат маса од 60 до 110 kDa. Меѓу нив најмногу се изучувани три класи: Ш-60, Ш-70 и Ш-90. Секоја класа вклучува семејство на сродни протеини. Така, Ш-70 вклучува протеини со молекуларна тежина од 66 до 78 kDa. Шаперони со ниска молекуларна тежина имаат молекуларна тежина од 40 до 15 kDa.
Меѓу придружниците има конститутивенпротеини, чија висока базална синтеза не зависи од стресните ефекти врз клетките на телото и индуцирана,чија синтеза во нормални услови е слаба, но нагло се зголемува под стрес. Индуцираните чаперони се нарекуваат и „протеини од топлински шок“ затоа што првпат биле откриени во клетките изложени на високи температури. Во клетките, поради високата концентрација на протеини, спонтано реактивирање на делумно денатурирани протеини е тешко. Ш-70 може да го спречи почетокот на денатурацијата и да помогне во обновувањето на природната конформација на протеините. Молекуларни чаперони-70- високо конзервирана класа на протеини кои се наоѓаат во сите делови на клетката: цитоплазма, јадро, ендоплазматичен ретикулум, митохондрии. На карбоксилниот крај на единечниот полипептиден синџир Ш-70 има регион кој е жлеб способен за интеракција со пептиди со должина
од 7 до 9 остатоци од аминокиселини збогатени со хидрофобни радикали. Таквите региони во глобуларните протеини се јавуваат приближно на секои 16 аминокиселини. Ш-70 е способен да ги заштити протеините од температурна инактивација и да ја врати конформацијата и активноста на делумно денатурираните протеини.
2. Улогата на чаперони во превиткување на протеини.За време на синтезата на протеините на рибозомот, N-терминалниот регион на полипептидот се синтетизира пред C-терминалниот регион. За да се формира природната конформација, потребна е целосна аминокиселинска секвенца на протеинот. Во процесот на синтеза на протеини, chaperones-70, поради структурата на нивниот активен центар, се способни да ги затворат областите на полипептидот кои се склони кон агрегација, збогатени со хидрофобни аминокиселински радикали додека не се заврши синтезата (Слика 1.27, A ).
 Ориз. 1.27. Учество на чаперони во превиткување на протеини
Ориз. 1.27. Учество на чаперони во превиткување на протеини
А - учество на chaperones-70 во спречување на хидрофобни интеракции помеѓу деловите на синтетизираниот полипептид; Б - формирање на природна конформација на протеинот во комплексот на чаперони
Многу високомолекуларни протеини кои имаат сложена конформација, како што е структурата на доменот, се преклопуваат во посебен простор формиран од Ш-60. Ш-60функционира како олигомерен комплекс кој се состои од 14 подединици. Тие формираат два шупливи прстени, од кои секоја се состои од седум подединици, овие прстени се поврзани едни со други. Секоја подединица Sh-60 се состои од три домени: апикална (апикална), збогатена со хидрофобни радикали свртени кон шуплината на прстенот, средно и екваторијално (сл. 1.28).
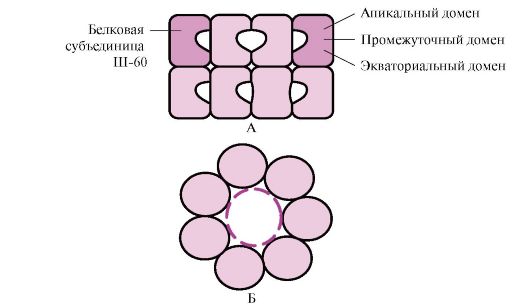 Ориз. 1.28. Структура на комплексот чаперонин кој се состои од 14 Ш-60
Ориз. 1.28. Структура на комплексот чаперонин кој се состои од 14 Ш-60
А - страничен поглед; Б - поглед одозгора
Синтетизираните протеини, кои имаат елементи на површината карактеристични за расклопените молекули, особено хидрофобните радикали, влегуваат во шуплината на придружните прстени. Во специфичното опкружување на овие шуплини, се бараат можни конформации додека не се најде единствената енергетски најповолна (сл. 1.27, Б). Формирањето на конформации и ослободувањето на протеини е придружено со АТП хидролизаво екваторијалниот регион. Вообичаено, ваквото преклопување кое зависи од чаперон бара значителна количина на енергија.
Покрај учеството во формирањето на тродимензионалната структура на протеините и обновувањето на делумно денатурираните протеини, шапероните се исто така неопходни за појава на такви основни процеси како што се склопување на олигомерни протеини, препознавање и транспорт на денатурирани протеини во лизозоми, транспорт на протеини низ мембраните и учество во регулација на активноста на протеинските комплекси.
ТЕМА 1.6. РАЗНОСТ НА ПРОТЕИНИ. ПРОТЕИНИ СЕМЕЈСТВА: ПРИМЕР НА ИМУНОГЛОБУЛИНИ
1. Протеините играат клучна улога во животот на поединечните клетки и сè повеќеклеточен организам, а нивните функции се изненадувачки разновидни. Ова се одредува според карактеристиките на примарната структура и конформациите на протеините, уникатната структура на активниот центар и способноста да се врзуваат специфични лиганди.
Само многу мал дел од сите можни варијанти на пептидни синџири може да усвои стабилна просторна структура; мнозинство
од нив може да земат многу конформации со приближно иста Гибсова енергија, но со различни својства. Избрана е примарна структура на повеќето познати протеини биолошка еволуција, обезбедува исклучителна стабилност на една од конформациите, што ги одредува особеностите на функционирањето на овој протеин.
2. Протеински семејства.Во рамките на истиот биолошки вид, замените на аминокиселинските остатоци може да доведат до појава на различни протеини кои вршат сродни функции и имаат хомологни низиамино киселини. Ваквите сродни протеини имаат неверојатно слични конформации: бројот и релативните позиции на α-спиралите и/или β-структурите, а повеќето вртења и свиоци на полипептидните синџири се слични или идентични. Протеините со хомологни региони на полипептидниот синџир, слична конформација и сродни функции се класифицирани во фамилии на протеини. Примери на протеински семејства: серински протеинази, имуноглобулинско семејство, миоглобинско семејство.
Серински протенази- семејство на протеини кои вршат функција на протеолитички ензими. Тие вклучуваат дигестивни ензими - химотрипсин, трипсин, еластаза и многу фактори на згрутчување на крвта. Овие протеини имаат идентични амино киселини во 40% од нивните позиции и многу слична конформација (сл. 1.29).

Ориз. 1.29. Просторни структуриеластаза (А) и химотрипсин (Б)
Некои замени на амино киселини доведоа до промени во специфичноста на супстратот на овие протеини и појава на функционална разновидност во семејството.
3. Семејство на имуноглобулини.Во функционирањето на имунолошкиот систем, протеините од суперфамилијата на имуноглобулини играат огромна улога, која вклучува три фамилии на протеини:
Антитела (имуноглобулини);
Т-лимфоцитни рецептори;
Протеини од главниот комплекс на хистокомпатибилност - MHC класи 1 и 2 (Главен комплекс на хистокомпатибилност).
Сите овие протеини имаат структура на домен, се состојат од хомологни домени слични на имунолошкиот систем и вршат слични функции: тие комуницираат со туѓи структури, растворени во крвта, лимфата или меѓуклеточната течност (антитела) или лоцирани на површината на клетките (сопствени или странски).
4. Антитела- специфични протеини произведени од Б-лимфоцитите како одговор на влегувањето на туѓа структура во телото, наречени антиген.
Карактеристики на структурата на антителата
Наједноставните молекули на антитела се состојат од четири полипептидни синџири: два идентични лесни - L, кои содржат околу 220 амино киселини и две идентични тешки - H, кои се состојат од 440-700 амино киселини. Сите четири синџири во молекулата на антителото се поврзани со многу не-ковалентни врски и четири дисулфидни врски (сл. 1.30).
Лесните синџири на антитела се состојат од два домени: променлив домен (VL), кој се наоѓа во N-терминалниот регион на полипептидниот синџир и константен домен (CL), кој се наоѓа на C-крајот. Тешките синџири обично имаат четири домени: една променлива (VH), лоцирана на N-крајот, и три константни домени (CH1, CH2, CH3) (види Сл. 1.30). Секој домен на имуноглобулин има надградба на β-лист во која два цистеински остатоци се поврзани со дисулфидна врска.
Помеѓу двата константни домени CH1 и CH2 постои регион кој содржи голем бројостатоци од пролин, кои го спречуваат формирањето на секундарна структура и интеракцијата на соседните H-синџири во овој сегмент. Овој регион на шарката му дава флексибилност на молекулата на антителото. Помеѓу променливите домени на тешкиот и лесниот синџир постојат две идентични места за врзување на антигенот (активни места за врзување антигени), затоа таквите антитела често се нарекуваат бивалентни.Не целата аминокиселинска секвенца на променливите региони на двата синџири е вклучена во врзувањето на антигенот со антителото, туку само 20-30 аминокиселини лоцирани во хиперпроменливите региони на секој синџир. Токму овие региони ја одредуваат уникатната способност на секој тип на антитела да комуницира со соодветниот комплементарен антиген.
Антителата се една од одбранбените линии на телото од навлегување на туѓи организми. Нивното функционирање може да се подели во две фази: првата фаза е препознавање и врзување на антигенот на површината на туѓите организми, што е можно поради присуството на места за врзување на антиген во структурата на антителата; втората фаза е започнување на процесот на инактивација и уништување на антигенот. Специфичноста на втората фаза зависи од класата на антитела. Постојат пет класи на тешки синџири, кои се разликуваат една од друга во структурата на константните домени: α, δ, ε, γ и μ, според кои се разликуваат пет класи на имуноглобулини: A, D, E, G и M.
Структурните карактеристики на тешките синџири им даваат на регионите на шарките и на C-терминалните региони на тешките синџири конформативна карактеристика на секоја класа. Откако антигенот се врзува за антитело, конформационите промени во константните домени го одредуваат патот за отстранување на антигенот.
 Ориз. 1. 30. Структура на домен на IgG
Ориз. 1. 30. Структура на домен на IgG
Имуноглобулини М
Имуноглобулините М имаат две форми.
Мономерна форма- 1-ва класа на антитела произведени со развој на Б-лимфоцити. Последователно, многу Б-клетки се префрлаат на производство на други класи на антитела, но со истото место за врзување на антигенот. IgM е вграден во мембраната и делува како рецептор за препознавање на антиген. Интеграцијата на IgM во клеточната мембрана е можна поради присуството на 25 хидрофобни амино киселински остатоци во опашката на регионот.
Секреторен облик на IgMсодржи пет мономерни подединици поврзани една со друга со дисулфидни врски и дополнителен полипептиден J синџир (сл. 1.31). Тешките синџири на мономери од оваа форма не содржат хидрофобна опашка. Пентамерот има 10 места за врзување на антигенот и затоа е ефикасен во препознавање и отстранување на антигенот кој прв влегува во телото. Секреторната форма на IgM е главната класа на антитела секретирани во крвта за време на примарниот имунолошки одговор. Врзувањето на IgM за антиген ја менува конформацијата на IgM и го индуцира неговото врзување за првата протеинска компонента на системот на комплементот (системот на комплементот е збир на протеини вклучени во уништувањето на антигенот) и активирање на овој систем. Ако антигенот се наоѓа на површината на микроорганизмот, системот на комплемент предизвикува нарушување на интегритетот клеточната мембранаи смрт на бактериската клетка.
Имуноглобулини Г
Квантитативно, оваа класа на имуноглобулини преовладува во крвта (75% од сите ИГ). IgG - мономери, главната класа на антитела секретирани во крвта за време на секундарен имунолошки одговор. По интеракцијата на IgG со површинските антигени на микроорганизмите, комплексот антиген-антитела е во состојба да ги врзува и активира протеините на системот на комплементот или може да комуницира со специфични рецептори на макрофаги и неутрофили. Интеракцијата со фагоцитите води
 Ориз. 1.31. Структура на секреторната форма на IgM
Ориз. 1.31. Структура на секреторната форма на IgM
до апсорпција на комплекси антиген-антитела и нивно уништување во клеточните фагозоми. IgG е единствената класа на антитела кои можат да навлезат во плацентарната бариера и да обезбедат интраутерина заштита на фетусот од инфекции.
Имуноглобулини А
Главната класа на антитела присутни во секретите (млеко, плунка, секрети на респираторниот тракт и цревниот тракт). IgA се секретира првенствено во димерна форма, каде што мономерите се поврзани еден со друг преку дополнителен J синџир (сл. 1.32).
IgA не е во интеракција со системот на комплементот и фагоцитните клетки, но со врзување за микроорганизмите, антителата го спречуваат нивното прицврстување за епителните клетки и пенетрацијата во телото.
Имуноглобулини Е
Имуноглобулините Е се претставени со мономери во кои тешките ε-синџири содржат, како μ-синџирите на имуноглобулините М, една променлива и четири константни домени. По секрецијата, IgE се врзува со него
 Ориз. 1.32. Структура на IgA
Ориз. 1.32. Структура на IgA
Ц-терминални региони со соодветни рецептори на површината на мастоцитите и базофилите. Како резултат на тоа, тие стануваат рецептори за антигени на површината на овие клетки (сл. 1.33).
 Ориз. 1.33. Интеракција на IgE со антиген на површината на мастоцитите
Ориз. 1.33. Интеракција на IgE со антиген на површината на мастоцитите
Откако антигенот се врзува за соодветните места за врзување на антигенот на IgE, клетките добиваат сигнал за биолошки лачење активни супстанции(хистамин, серотонин), кои во голема мера се одговорни за развојот на воспалителната реакција и за манифестацијата на алергиски реакции како што се астма, уртикарија, поленска треска.
Имуноглобулини Д
Имуноглобулините Д се наоѓаат во многу мали количини во серумот; тие се мономери. Тешките δ синџири имаат една променлива и три константни домени. IgD делуваат како рецептори за Б-лимфоцитите; другите функции сè уште се непознати. Интеракцијата на специфични антигени со рецепторите на површината на Б-лимфоцитите (IgD) доведува до пренос на овие сигнали во клетката и активирање на механизмите кои обезбедуваат пролиферација на даден лимфоцитен клон.
ТЕМА 1.7. ФИЗИЧКИ И ХЕМИСКИ СВОЈСТВА НА ПРОТЕИНИ И МЕТОДИ ЗА НИВНО РАЗДЕЛУВАЊЕ
1. Индивидуалните протеини се разликуваат по физички и хемиски својства:
Облик на молекули;
Молекуларна тежина;
Вкупниот полнеж, чија големина зависи од односот на анјонските и катјонските групи на амино киселини;
Односот на поларните и неполарните аминокиселински радикали на површината на молекулите;
Степени на отпорност на различни денатурирачки агенси.
2. Растворливоста на протеините зависиза својствата на протеините наведени погоре, како и за составот на медиумот во кој се раствора протеинот (рН вредности, состав на сол, температура, присуство на други органски материи кои можат да комуницираат со протеинот). Количината на полнење на протеинските молекули е еден од факторите кои влијаат на нивната растворливост. Кога полнењето во изоелектричната точка ќе се изгуби, протеините полесно се агрегираат и таложат. Ова е особено типично за денатурирани протеини, во кои на површината се појавуваат хидрофобни аминокиселински радикали.
На површината на протеинската молекула има и позитивно и негативно наелектризирани радикали на аминокиселини. Бројот на овие групи, а со тоа и вкупниот полнеж на протеините, зависи од рН на медиумот, т.е. однос на концентрации на H+ - и OH - групи. Во кисела срединаЗголемувањето на концентрацијата на H+ доведува до потиснување на дисоцијацијата на карбоксилните групи -COO - + H+ > - COOH и намалување на негативниот полнеж на протеините. Во алкална средина, врзувањето на вишокот OH - со протони формирани при дисоцијација на амино групи -NH 3 + + OH - - NH 2 + H 2 O со формирање на вода, доведува до намалување на позитивниот полнеж на протеините . рН вредноста на која протеинот има нето нула полнеж се нарекува изоелектрична точка (IEP).Во IET, бројот на позитивно и негативно наелектризираните групи е ист, т.е. протеинот е во изоелектрична состојба.
3. Одвојување на поединечни протеини.Карактеристиките на структурата и функционирањето на телото зависат од збирот на протеини синтетизирани во него. Проучувањето на структурата и својствата на протеините е невозможно без нивна изолација од клетката и прочистување од други протеини и органски молекули. Фази на изолација и прочистување на поединечни протеини:
Уништување на клеткитеткивото што се проучува и се добива хомогенат.
Одвојување на хомогенатот на фракциисо центрифугирање, добивајќи нуклеарна, митохондријална, цитосолна или друга фракција која го содржи саканиот протеин.
Селективна термичка денатурација- краткотрајно загревање на протеински раствор, при што може да се отстранат дел од денатурираните протеински нечистотии (ако протеинот е релативно термостабилен).
Солење надвор.Различни протеини се таложат при различни концентрации на сол во растворот. Со постепено зголемување на концентрацијата на сол, можно е да се добијат голем број посебни фракции со доминантна содржина на изолиран протеин во една од нив. Амониум сулфат најчесто се користи за фракционирање на протеини. Протеините со најмала растворливост се таложат при ниски концентрации на сол.
Гел филтрација- метод за просејување на молекули низ отечени Sephadex гранули (тридимензионални полисахаридни синџири на декстран кои имаат пори). Брзината со која протеините минуваат низ колона исполнета со Sephadex ќе зависи од нивната молекуларна тежина: колку е помала масата на протеинските молекули, толку полесно тие продираат во гранулите и остануваат таму подолго од поголема маса, толку побрзо елуираат од колоната.
Ултрацентрифугирање- метод кој вклучува ставање протеини во цевка за центрифуга во роторот на ултрацентрифуга. Кога роторот се ротира, стапката на седиментација на протеините е пропорционална на нивната молекуларна тежина: фракциите на потешките протеини се наоѓаат поблиску до дното на епрувета, полесните - поблиску до површината.
Електрофореза- метод заснован на разлики во брзината на движење на протеините во електрично поле. Оваа вредност е пропорционална на полнењето на протеините. Електрофорезата на протеините се изведува на хартија (во овој случај, брзината на движење на протеините е пропорционална само со нивното полнење) или во гел од полиакриламид со одредена големина на порите (брзината на движење на протеините е пропорционална на нивниот полнеж и молекуларна тежина) .
Хроматографија за размена на јони- метод на фракционирање базиран на врзување на јонизирани групи на протеини со спротивно наелектризирани групи на јонско менувачки смоли (нерастворливи полимерни материјали). Јачината на врзувањето на протеинот со смолата е пропорционална на полнењето на протеинот. Протеините адсорбирани во полимерот за размена на јони може да се измијат со зголемување на концентрациите на растворите на NaCl; колку е помал протеинскиот полнеж, толку е помала концентрацијата на NaCl потребна за да се измие протеинот врзан за јонските групи на смолата.
Афинитетна хроматографија- најспецифичен метод за изолирање на поединечни протеини Лиганд на протеин е ковалентно врзан за инертен полимер. Кога протеинскиот раствор се пренесува низ колона со полимер, само протеинот специфичен за даден лиганд се адсорбира на колоната поради комплементарното врзување на протеинот со лигандот.
Дијализа- метод кој се користи за отстранување на соединенија со мала молекуларна тежина од раствор на изолиран протеин. Методот се заснова на неможноста на протеините да поминат низ полупропустлива мембрана, за разлика од супстанциите со мала молекуларна тежина. Се користи за прочистување на протеините од нискомолекуларни нечистотии, на пример, соли по солење.
ЗАДАЧИ ЗА ВОННАСТАВНА РАБОТА
1. Пополнете ја табелата. 1.4.
Табела 1.4. Компаративна анализаструктура и функции на сродни протеини - миоглобин и хемоглобин
а) запомнете ја структурата на активниот центар на Mb и Hb. Каква улога играат хидрофобните аминокиселински радикали во формирањето на активните центри на овие протеини? Опишете ја структурата на активниот центар на Mb и Hb и механизмите на прицврстување на лигандите кон него. Каква улога играат остатоците His F 8 и His E 7 во функционирањето на активниот центар на Mv iHv?
б) какви нови својства во споредба со мономерниот миоглобин има тесно поврзаниот олигомерен протеин, хемоглобинот? Објаснете ја улогата на кооперативните промени во конформацијата на протомерите во молекулата на хемоглобинот, ефектот на концентрациите на CO 2 и протонот врз афинитетот на хемоглобинот за кислород, како и улогата на 2,3-BPG во алостеричната регулација на функцијата на Hb .
2. Карактеризирајте ги молекуларните чаперони, обрнувајќи внимание на односот помеѓу нивната структура и функција.
3. Кои протеини се групирани во семејства? Користејќи го примерот на семејството на имуноглобулини, идентификувајте слични структурни карактеристики и сродни функции на протеините од оваа фамилија.
4. Често се потребни прочистени индивидуални протеини за биохемиски и медицински цели. Објаснете кој физички и хемиски својствапротеините се базираат на методите што се користат за нивно одвојување и прочистување.
ЗАДАЧИ ЗА САМОКОНТРОЛА
1. Изберете ги точните одговори.
Функции на хемоглобинот:
А. Транспорт на О 2 од белите дробови до ткивото Б. Транспорт на H + од ткивата до белите дробови
Б. Одржување константна pH на крвта D. Транспорт на CO 2 од белите дробови до ткивата
Д. Транспорт на CO 2 од ткивата до белите дробови
2. Изберете ги точните одговори. Лигандα - протомер Hb е:А. Хеме
Б. Кислород
B. CO G. 2,3-BPG
D. β-протомер
3. Изберете ги точните одговори.
Хемоглобин за разлика од миоглобинот:
A. Има кватернарна структура
B. Секундарната структура е претставена само со α-спирали
Б. Припаѓа на комплексни протеини
D. Станува во интеракција со алостеричен лиганд D. Ковалентно врзан за хем
4. Изберете ги точните одговори.
Афинитетот на Hb за O2 се намалува:
A. Кога се додава една молекула O 2 B. Кога се отстранува една молекула O 2
B. При интеракција со 2,3-BPG
D. Кога е прикачен на протомери H + D. Кога концентрацијата на 2,3-BPG се намалува
5. Натпревар.
Видовите HB се карактеризираат со:
А. Во деокси форма формира фибриларни агрегати Б. Содржи два α- и два δ-синџири
B. Доминантна форма на Hb кај возрасни еритроцити D. Содржи хем со Fe + 3 во активниот центар
D. Содржи два α- и два γ-синџири 1. HbA 2.
6. Натпревар.
Hb лиганди:
A. Се врзува за Hb во алостеричниот центар
Б. Има многу висок афинитет за активното место на Hb
Б. Со спојување, го зголемува афинитетот на Hb за O 2 G. Оксидира Fe+ 2 до Fe+ 3
D. Форми ковалентна врскасо gisF8
7. Изберете ги точните одговори.
Шеперони:
A. Протеини присутни во сите делови на клетката
Б. Синтезата се зголемува под стрес
Б. Учествуваат во хидролиза на денатурирани протеини
D. Учествувајте во одржувањето на природната конформација на протеините
D. Тие создаваат органели во кои се формира конформацијата на протеините.
8. Натпревар. Имуноглобулини:
A. Секреторната форма е пентамерна.
Б. Класа Ig која продира во плацентарната бариера
B. Ig - рецептор на мастоцитите
D. Главната класа на Ig присутна во секретите на епителните клетки. D. Б-лимфоцитен рецептор, чие активирање обезбедува клеточна пролиферација
9. Изберете ги точните одговори.
Имуноглобулини Е:
A. Произведени од макрофаги B. Тие имаат тешки ε синџири.
B. Вграден во мембраната на Т-лимфоцитите
D. Дејствуваат како мембрански антигенски рецептори на мастоцитите и базофилите
D. Одговорен за алергиски реакции
10. Изберете ги точните одговори.
Методот за одвојување на протеините се заснова на разликите во нивната молекуларна тежина:
A. Гел филтрација
Б. Ултрацентрифугација
Б. Електрофореза со полиакриламид гел Д. Хроматографија за размена на јони
Д. Афинитетна хроматографија
11. Одбери го точниот одговор.
Методот за одвојување на протеините се заснова на разликите во нивната растворливост во вода:
А. Гел филтрација Б. Солење надвор
Б. Јонска размена на хроматографија Г. Афинитетна хроматографија
D. Електрофореза во полиакриламид гел
СТАНДАРДИ НА ОДГОВОРИ НА „ЗАДАЧИ ЗА САМОКОНТРОЛА“
1. А, Б, Ц, Д
2. А, Б, Ц, Д
5. 1-Б, 2-А, 3-Г
6. 1-Б, 2-Б, 3-А
7. А, Б, Д, Д
8. 1-G; 2-Б, 3-Б
ОСНОВНИ ПОИМИ И ПОИМИ
1. Олигомерни протеини, протомер, кватернарна структура на протеините
2. Кооперативни промени во конформацијата на протомерот
3. Боров ефект
4. Алостерична регулација на протеинските функции, алостеричен центар и алостеричен ефектор
5. Молекуларни шаперони, протеини од топлински шок
6. Фамилии на протеини (серин протеази, имуноглобулини)
7. Однос IgM-, G-, E-, A-структура-функција
8. Вкупен полнеж на протеини, изоелектрична точка на протеини
9. Електрофореза
10. Солење надвор
11. Гел филтрација
12. Јонска размена на хроматографија
13. Ултрацентрифугирање
14. Афинитетна хроматографија
15. Електрофореза на протеините во крвната плазма
ЗАДАЧИ ЗА РАБОТА ВО УЧИЛНИЦА
1. Споредете ги зависностите на степените на заситеност на хемоглобинот (Hb) и миоглобинот (Mb) со кислород од неговиот парцијален притисок во ткивата
 Ориз. 1.34. Зависност од сатурација Mv иNHкислород од неговиот парцијален притисок
Ориз. 1.34. Зависност од сатурација Mv иNHкислород од неговиот парцијален притисок
Ве молиме имајте предвид дека обликот на кривите за заситеност со протеински кислород е различен: за миоглобинот - хипербола, за хемоглобинот - сигмоиден облик.
1. споредете ги вредностите на парцијалниот притисок на кислородот при кој Mb и Hb се заситени со O 2 за 50%. Кој од овие протеини има поголем афинитет за O 2?
2. Кои структурни карактеристики на Mb го одредуваат неговиот висок афинитет за O 2?
3. Кои структурни карактеристики на HB му дозволуваат да ослободува О2 во капиларите на ткивата во мирување (при релативно висок парцијален притисок на О2) и нагло да го зголеми ова ослободување во работните мускули? Кое својство на олигомерните протеини го обезбедува овој ефект?
4. Пресметајте колкаво количество на O 2 (во%) оксигениран хемоглобин им дава на мускулите кои се одмораат и работат?
5. донесува заклучоци за односот помеѓу структурата на протеинот и неговата функција.
2. Количината на кислород што се ослободува од хемоглобинот во капиларите зависи од интензитетот на катаболичките процеси во ткивата (Боров ефект). Како промените во ткивниот метаболизам го регулираат афинитетот на Hb за O2? Ефект на CO 2 и H + врз афинитетот на Hb за O 2
 1. опишете го Боров ефект.
1. опишете го Боров ефект.
2. во која насока се одвива процесот прикажан на дијаграмот:
а) во капиларите на белите дробови;
б) во ткивните капилари?
3. Кое е физиолошкото значење на Боровиот ефект?
4. Зошто интеракцијата на Hb со H+ на места оддалечени од хем го менува афинитетот на протеинот за O 2?
3. Афинитетот на Hb за O2 зависи од концентрацијата на неговиот лиганд - 2,3-бисфосфоглицерат, кој е алостеричен регулатор на афинитетот на Hb за O2. Зошто интеракцијата на лигандот на место оддалечено од активното место влијае на функцијата на протеините? Како 2,3-BPG го регулира афинитетот на Hb за O2? За да го решите проблемот, одговорете на следниве прашања:
1. каде и од што се синтетизира 2,3-бисфосфоглицерат (2,3-BPG)? Напишете ја неговата формула, означете го полнењето на оваа молекула.
2. Со која форма на хемоглобин (окси или деокси) делува БПГ и зошто? Во кој дел од молекулата на Hb се јавува интеракцијата?
3. во која насока се случува процесот прикажан на дијаграмот?
а) во ткивните капилари;
б) во капиларите на белите дробови?
4. каде концентрацијата на комплексот треба да биде поголема
Nv-2,3-BFG:
а) во капиларите на мускулите во мирување,
б) во капиларите на работните мускули (обезбедена е иста концентрација на БПГ во еритроцитите)?
5. Како ќе се промени афинитетот на HB за кислород кога човекот се прилагодува на условите на голема надморска височина, доколку се зголеми концентрацијата на BPG во еритроцитите? Кое е физиолошкото значење на овој феномен?
4. Уништувањето на 2,3-BPG при складирање на зачувана крв ги нарушува функциите на HB. Како ќе се промени афинитетот на HB за O 2 во зачуваната крв ако концентрацијата на 2,3-BPG во еритроцитите може да се намали од 8 на 0,5 mmol/l. Дали е можно да се трансфузира таква крв на сериозно болни пациенти ако концентрацијата на 2,3-BPG се врати не порано од три дена? Дали е можно да се обноват функциите на црвените крвни зрнца со додавање на 2,3-BPG во крвта?
5. Запомнете ја структурата на наједноставните имуноглобулински молекули. Каква улога играат имуноглобулините во имунолошкиот систем? Зошто Igs често се нарекуваат двовалентни? Како структурата на Igs е поврзана со нивната функција? (Опишете користејќи пример на класа на имуноглобулини.)
Физичко-хемиски својства на протеините и методи на нивно одвојување.
6. Како нето полнежот на протеинот влијае на неговата растворливост?
а) определи го вкупниот полнеж на пептидот на pH 7
Ала-Глу-Тре-Про-Асп-Лиз-Цис
б) како ќе се промени полнењето на овој пептид при pH >7, pH<7, рН <<7?
в) која е изоелектричната точка на протеинот (IEP) и во која средина лежи?
IET на овој пептид?
г) при која pH вредност ќе биде забележана најмала растворливост на овој пептид.
7. Зошто киселото млеко, за разлика од свежото млеко, „замрсува“ кога се вари (т.е. млечниот протеин казеин таложи)? Во свежото млеко, молекулите на казеин имаат негативен полнеж.
8. Гел-филтрација се користи за одвојување на поединечни протеини. Мешавина која содржи протеини A, B, C со молекуларна тежина еднаква на 160.000, 80.000 и 60.000, соодветно, беше анализирана со гел-филтрација (сл. 1.35). Надојдените гел гранули се пропустливи за протеини со молекуларна тежина помала од 70.000. Кој принцип лежи во основата на овој метод на сепарација? Кој графикон правилно ги одразува резултатите од фракционирањето? Наведете го редоследот по кој протеините A, B и C се ослободуваат од колоната.
 Ориз. 1.35. Користење на филтрација со гел за одвојување на протеини
Ориз. 1.35. Користење на филтрација со гел за одвојување на протеини
9. На сл. 1.36, А покажува дијаграм на електрофореза на хартија на протеини од крвниот серум на здрава личност. Релативните количини на протеински фракции добиени со овој метод се: албумини 54-58%, α1-глобулини 6-7%, α2-глобулини 8-9%, β-глобулини 13%, γ-глобулини 11-12%.
 Ориз. 1.36 Електрофореза на хартија на протеини од крвната плазма на здрава личност (А) и пациент (Б)
Ориз. 1.36 Електрофореза на хартија на протеини од крвната плазма на здрава личност (А) и пациент (Б)
I - γ-глобулини; II - β-глобулини; III -α 2 -глобулин; IV -α 2 -глобулин; V - албумини
Многу болести се придружени со квантитативни промени во составот на серумските протеини (диспротеинемија). Природата на овие промени се зема предвид при поставување на дијагноза и проценка на тежината и фазата на болеста.
1. Зошто протеините се сметаат за полимери?
Одговори. Протеините се полимери, односно молекули изградени како синџири од повторувачки мономерни единици, или подединици, кои се состојат од амино киселини поврзани во одредена низа со пептидна врска. Тие се основните и неопходните компоненти на сите организми.
Постојат едноставни протеини (протеини) и сложени протеини (протеиди). Протеините се протеини чии молекули содржат само протеински компоненти. Кога тие се целосно хидролизирани, се формираат амино киселини.
Протеидите се сложени протеини чии молекули значително се разликуваат од протеинските молекули со тоа што, покрај самата протеинска компонента, тие содржат компонента со мала молекуларна тежина од непротеинска природа.
2. Кои функции на протеините ги знаете?
Одговори. Протеините ги извршуваат следните функции: градежни, енергетски, каталитички, заштитни, транспортни, контрактилни, сигнални и други.
Прашања по § 11
1. Кои материи се нарекуваат протеини?
Одговори. Протеините или протеините се биолошки полимери чии мономери се амино киселини. Сите амино киселини имаат амино група (-NH2) и карбоксилна група (-COOH) и се разликуваат по структурата и својствата на радикалите. Амино киселините се поврзани една со друга со пептидни врски, поради што протеините се нарекуваат и полипептиди.
Одговори. Протеинските молекули можат да добијат различни просторни форми - конформации, кои претставуваат четири нивоа на нивната организација. Линеарната низа на амино киселини во полипептидниот синџир ја претставува примарната структура на протеинот. Тој е единствен за секој протеин и ја одредува неговата форма, својства и функции.
3. Како се формираат секундарни, терциерни и кватернарни протеински структури?
Одговори. Секундарната структура на протеинот се формира со формирање на водородни врски помеѓу -CO- и -NH- групите. Во овој случај, полипептидниот синџир се превртува во спирала. Спиралата може да добие конфигурација на глобули, бидејќи се појавуваат различни врски помеѓу радикалите на аминокиселините во спиралата. Глобулата е терциерна структура на протеинот. Ако неколку глобули се спојат во еден комплексен комплекс, се јавува кватернарна структура. На пример, хемоглобинот во човечката крв е формиран од четири глобули.
4. Што е денатурација на протеини?
Одговори. Повреда на природната структура на протеинот се нарекува денатурација. Под влијание на голем број фактори (хемиски, радиоактивни, температурни итн.), кватернарните, терциерните и секундарните структури на протеинот можат да бидат уништени. Ако ефектот на факторот престане, протеинот може да ја врати својата структура. Ако ефектот на факторот се зголеми, примарната структура на протеинот - полипептидниот синџир - исто така се уништува. Ова е неповратен процес - протеинот не може да ја врати својата структура
5. На која основа протеините се делат на едноставни и сложени?
Одговори. Едноставните протеини се состојат исклучиво од амино киселини. Комплексните протеини може да вклучуваат и други органски материи: јаглехидрати (тогаш наречени гликопротеини), масти (липопротеини), нуклеински киселини (нуклеопротеини).
6. Кои функции на протеините ги знаете?
Одговори. Конструктивна (пластична) функција. Протеините се структурна компонента на биолошките мембрани и клеточните органели, а исто така се дел од потпорните структури на телото, косата, ноктите и крвните садови. Ензимска функција. Протеините служат како ензими, односно биолошки катализатори кои ја забрзуваат брзината на биохемиските реакции десетици и стотици милиони пати. Пример е амилазата, која го разложува скробот во моносахариди. Контрактилна (моторна) функција. Се изведува со специјални контрактилни протеини кои обезбедуваат движење на клетките и интрацелуларните структури. Благодарение на нив, хромозомите се движат за време на клеточната делба, а флагелите и цилиите ги движат клетките на протозои. Контрактилните својства на протеините актин и миозин се во основата на мускулната функција. Транспортна функција. Протеините се вклучени во транспортот на молекули и јони во телото (хемоглобинот носи кислород од белите дробови до органите и ткивата, серумскиот албумин е вклучен во транспортот на масни киселини). Заштитна функција. Се состои од заштита на телото од оштетување и инвазија на странски протеини и бактерии. Протеините на антителата произведени од лимфоцитите создаваат одбрана на телото од туѓа инфекција; тромбинот и фибринот се вклучени во формирањето на згрутчување на крвта, а со тоа му помагаат на телото да избегне големи загуби на крв. Регулаторна функција. Се изведува од хормонски протеини. Тие учествуваат во регулирањето на клеточната активност и сите витални процеси на телото. Така, инсулинот го регулира нивото на шеќер во крвта и го одржува на одредено ниво. Сигнална функција. Протеините вградени во клеточната мембрана се способни да ја променат нивната структура како одговор на иритација. Така, сигналите се пренесуваат од надворешната средина во ќелијата. Енергетска функција. Исклучително ретко се реализира од протеини. Со целосно разградување на 1 g протеин, може да се ослободи 17,6 kJ енергија. Сепак, протеините се многу вредно соединение за телото. Затоа, разградувањето на протеините обично се случува кај амино киселините, од кои се градат нови полипептидни синџири. Хормонските протеини ја регулираат активноста на клетката и сите животни процеси на телото. Така, во човечкото тело, соматотропинот е вклучен во регулирањето на растот на телото, инсулинот го одржува нивото на гликоза во крвта на константно ниво.
7. Каква улога играат хормонските протеини?
Одговори. Регулаторната функција е вродена во хормонските протеини (регулатори). Тие регулираат различни физиолошки процеси. На пример, најпознатиот хормон е инсулинот, кој го регулира нивото на гликоза во крвта. Кога нема доволно инсулин во телото, се јавува болест позната како дијабетес мелитус.
8. Каква функција вршат ензимските протеини?
Одговори. Ензимите се биолошки катализатори, односно ги забрзуваат хемиските реакции стотици милиони пати. Ензимите имаат строга специфичност за супстанцијата што реагира. Секоја реакција се катализира од свој ензим.
9. Зошто протеините ретко се користат како извор на енергија?
Одговори. Протеинските мономери на амино киселини се вредни суровини за изградба на нови протеински молекули. Затоа, целосно разградување на полипептидите во неоргански супстанции ретко се случува. Следствено, енергетската функција, која се состои во ослободување енергија при целосно распаѓање, ја вршат протеините доста ретко.
Белата од јајце е типичен протеин. Дознајте што ќе се случи со него ако е изложен на вода, алкохол, ацетон, киселина, алкали, растително масло, висока температура итн.
Одговори. Како резултат на ефектот на висока температура врз белката од јајцето, ќе дојде до денатурација на протеините. Кога се изложени на алкохол, ацетон, киселини или алкалии, се случува приближно истото: протеинот се коагулира. Ова е процес во кој терциерната и кватернарната структура на протеинот е нарушена поради прекин на водородните и јонските врски.
Во водата и растителното масло, протеинот ја задржува својата структура.
Сомелете ја кртулата од суров компир до каша. Земете три епрувети и во секоја ставете мала количина сечкани компири.
Првата епрувета ставете ја во замрзнувачот на фрижидерот, втората на долната полица од фрижидерот и третата во тегла со топла вода (t = 40 °C). По 30 минути, извадете ги епруветите и истурете мала количина водород пероксид во секоја од нив. Набљудувајте што се случува во секоја епрувета. Објаснете ги вашите резултати
Одговори. Овој експеримент ја илустрира активноста на ензимот каталаза во жива клетка на водород пероксид. Како резултат на реакцијата, се ослободува кислород. Динамиката на ослободување на меурчиња може да се користи за да се процени активноста на ензимот.
Искуството ни овозможи да ги забележиме следните резултати:
Активноста на каталазата зависи од температурата:
1. Епрувета 1: нема меурчиња - тоа е затоа што при ниски температури ќелиите на компирот колабирале.
2. Епрувета 2: има мал број меурчиња - бидејќи активноста на ензимот на ниски температури е мала.
3. Епрувета 3: има многу меурчиња, температурата е оптимална, каталазата е многу активна.
Капкајте неколку капки вода во првата епрувета со компири, неколку капки киселина (оцет за маса) во втората и алкали во третата.
Набљудувајте што се случува во секоја епрувета. Објаснете ги вашите резултати. Извлечете заклучоци.
Одговори. Кога се додава вода, ништо не се случува, кога се додава киселина, се јавува одредено затемнување, кога се додава алкали, се јавува „пенење“ - алкална хидролиза.
Како што знаете, протеините се основа за потеклото на животот на нашата планета. Но, токму коацерватната капка, составена од молекули на пептид, стана основа за потеклото на живите суштества. Ова е несомнено, бидејќи анализата на внатрешниот состав на кој било претставник на биомасата покажува дека овие супстанции се присутни во сè: растенија, животни, микроорганизми, габи, вируси. Покрај тоа, тие се многу разновидни и макромолекуларни по природа.
Овие структури имаат четири имиња, сите се синоними:
- протеини;
- протеини;
- полипептиди;
- пептиди.
Протеински молекули
Нивниот број е навистина безброј. Во овој случај, сите протеински молекули може да се поделат во две големи групи:
- едноставно - се состои само од секвенци на аминокиселини поврзани со пептидни врски;
- комплекс - структурата и структурата на протеинот се карактеризираат со дополнителни протолитички (протетски) групи, исто така наречени кофактори.
Во исто време, сложените молекули имаат и своја класификација.
Градација на комплексни пептиди
- Гликопротеините се тесно поврзани соединенија на протеини и јаглени хидрати. Протетските групи на мукополисахариди се вткаени во структурата на молекулата.
- Липопротеините се сложено соединение од протеини и липиди.
- Металопротеини - метални јони (железо, манган, бакар и други) делуваат како протетска група.
- Нуклеопротеините се врската помеѓу протеините и нуклеинските киселини (ДНК, РНК).
- Фосфопротеини - конформација на протеин и остаток на ортофосфорна киселина.
- Хромопротеините се многу слични на металопротеините, меѓутоа, елементот што е дел од протетската група е цел комплекс во боја (црвено - хемоглобин, зелено - хлорофил и така натаму).
Во секоја разгледана група, структурата и својствата на протеините се различни. Функциите што ги извршуваат исто така варираат во зависност од видот на молекулата.

Хемиска структура на протеините
Од оваа гледна точка, протеините се долг, масивен ланец на остатоци од аминокиселини поврзани едни со други со специфични врски наречени пептидни врски. Гранките наречени радикали се протегаат од страничните структури на киселините. Оваа молекуларна структура е откриена од Е. Фишер на почетокот на 21 век.
Подоцна, протеините, структурата и функциите на протеините беа подетално проучувани. Стана јасно дека има само 20 амино киселини кои ја формираат структурата на пептидот, но тие можат да се комбинираат на различни начини. Оттука и разновидноста на полипептидните структури. Покрај тоа, во процесот на живот и извршување на нивните функции, протеините се способни да подлежат на голем број хемиски трансформации. Како резултат на тоа, тие ја менуваат структурата и се појавува сосема нов тип на врска.
За да ја прекинете пептидната врска, односно да го нарушите протеинот и структурата на синџирите, треба да изберете многу строги услови (високи температури, киселини или алкалии, катализатор). Ова се должи на високата јачина во молекулата, имено во групата на пептиди.

Откривањето на структурата на протеините во лабораторија се врши со помош на реакцијата на биурет - изложеност на свежо преципитираниот полипептид (II). Комплексот на пептидната група и бакарниот јон дава светла виолетова боја.
Постојат четири главни структурни организации, од кои секоја има свои структурни карактеристики на протеините.
Нивоа на организација: примарна структура
Како што споменавме погоре, пептидот е низа од остатоци од аминокиселини со или без инклузии, коензими. Значи, примарната е структурата на молекула која е природна, природна, вистински аминокиселини поврзани со пептидни врски, и ништо повеќе. Тоа е, полипептид со линеарна структура. Покрај тоа, структурните карактеристики на протеините од овој тип се дека таквата комбинација на киселини е одлучувачка за извршување на функциите на протеинската молекула. Благодарение на присуството на овие карактеристики, можно е не само да се идентификува пептид, туку и да се предвидат својствата и улогата на сосема нов, сè уште неоткриен. Примери на пептиди со природна примарна структура се инсулин, пепсин, химотрипсин и други.

Секундарна конформација
Структурата и својствата на протеините во оваа категорија се разликуваат малку. Таквата структура може да се формира првично по природа или кога примарната е изложена на тешка хидролиза, температура или други услови.
Оваа конформација има три варијанти:
- Мазни, правилни, стереорегуларни свиоци, изградени од остатоци од аминокиселини, кои се извртуваат околу главната оска на врската. Тие се држат заедно само од оние што произлегуваат помеѓу кислородот на една пептидна група и водородот од друга. Покрај тоа, структурата се смета за точна поради фактот што вртењата се повторуваат рамномерно на секои 4 врски. Таквата структура може да биде или левак или деснак. Но, кај повеќето познати протеини доминира декстророторниот изомер. Ваквите конформации обично се нарекуваат алфа структури.
- Составот и структурата на протеините од следниот тип се разликуваат од претходниот по тоа што водородните врски не се формираат помеѓу остатоците во непосредна близина на едната страна од молекулата, туку помеѓу значително оддалечените и на прилично големо растојание. Поради оваа причина, целата структура има форма на неколку брановидни полипептидни синџири слични на змии. Постои една карактеристика што протеинот мора да ја покаже. Структурата на аминокиселините на гранките треба да биде што е можно пократка, како глицин или аланин, на пример. Овој тип на секундарна конформација се нарекува бета листови поради нивната способност да се држат заедно за да формираат заедничка структура.
- Биологијата се однесува на третиот тип на протеинска структура како сложени, хетерогено расфрлани, нарушени фрагменти кои немаат стереорегуларност и се способни да ја менуваат структурата под влијание на надворешни услови.
Не се идентификувани примери на протеини кои природно имаат секундарна структура.

Терцијарно образование
Ова е прилично сложена конформација наречена „глобула“. Што е овој протеин? Неговата структура се заснова на секундарната структура, меѓутоа, се додаваат нови типови на интеракции помеѓу атомите на групите, а целата молекула изгледа како да се превиткува, со што се фокусира на фактот дека хидрофилните групи се насочени во топката, а хидрофобните оние однадвор.
Ова го објаснува полнењето на протеинската молекула во колоидните раствори на вода. Какви видови на интеракции се присутни овде?
- Водородни врски - остануваат непроменети помеѓу истите делови како и во секундарната структура.
- интеракции - се јавуваат кога полипептидот се раствора во вода.
- Јонските атракции се формираат помеѓу различно наелектризираните групи на остатоци од аминокиселини (радикали).
- Ковалентни интеракции - може да се формираат помеѓу специфични кисели места - молекули на цистеин, или подобро кажано, нивните опашки.
Така, составот и структурата на протеините со терциерна структура може да се опишат како полипептидни синџири преклопени во глобули кои ја задржуваат и стабилизираат нивната конформација поради различни видови хемиски интеракции. Примери за такви пептиди: фосфоглицерат кеназа, tRNA, алфа-кератин, свилен фиброин и други.
Кватернарна структура
Ова е една од најкомплексните глобули што ги формираат протеините. Структурата и функциите на протеините од овој тип се многу повеќеслојни и специфични.
Каква е оваа конформација? Тоа се неколку (во некои случаи десетици) големи и мали полипептидни синџири кои се формираат независно еден од друг. Но потоа, поради истите интеракции што ги разгледавме за терциерната структура, сите овие пептиди се извртуваат и се испреплетуваат еден со друг. На овој начин се добиваат сложени конформациски глобули кои можат да содржат метални атоми, липидни групи и јаглехидрати. Примери за такви протеини: ДНК полимераза, протеинската обвивка на вирусот на тутунот, хемоглобинот и други.

Сите пептидни структури што ги испитавме имаат свои методи за идентификација во лабораторија, засновани на современи можности за користење на хроматографија, центрифугирање, електронска и оптичка микроскопија и високи компјутерски технологии.
Извршени функции
Структурата и функциите на протеините се тесно поврзани едни со други. Односно, секој пептид игра специфична улога, единствена и специфична. Има и такви кои се способни да извршат неколку значајни операции одеднаш во една жива клетка. Сепак, можно е да се изразат во генерализирана форма главните функции на протеинските молекули во живите организми:
- Обезбедување на движење. Едноклеточните организми или органели или некои типови на клетки се способни за движење, контракција и движење. Ова го обезбедуваат протеините кои ја сочинуваат структурата на нивниот моторен апарат: цилии, флагели и цитоплазматска мембрана. Ако зборуваме за клетки неспособни за движење, тогаш протеините можат да придонесат за нивната контракција (мускулен миозин).
- Нутритивна или резервна функција. Тоа е акумулација на протеински молекули во јајцата, ембрионите и семките на растенијата за дополнително да се надополнат хранливите материи кои недостасуваат. Кога се разградуваат, пептидите произведуваат амино киселини и биолошки активни супстанции кои се неопходни за нормален развој на живите организми.
- Енергетска функција. Покрај јаглехидратите, протеините исто така можат да му дадат сила на телото. Со распаѓањето на 1 g пептид се ослободуваат 17,6 kJ корисна енергија во форма на аденозин трифосфорна киселина (ATP), која се троши на витални процеси.
- Сигнализацијата се состои од внимателно следење на тековните процеси и пренесување сигнали од клетки до ткива, од нив до органи, од вторите до системите итн. Типичен пример е инсулинот, кој строго ја фиксира количината на гликоза во крвта.
- Функција на рецепторот. Се изведува со менување на конформацијата на пептидот на едната страна од мембраната и вклучување на другиот крај во реструктуирањето. Во исто време, сигналот и потребните информации се пренесуваат. Најчесто, таквите протеини се вградени во цитоплазматските мембрани на клетките и вршат строга контрола врз сите супстанции што минуваат низ него. Тие исто така даваат информации за хемиските и физичките промени во животната средина.
- Транспортна функција на пептиди. Се изведува преку канални протеини и транспортерни протеини. Нивната улога е очигледна - транспортирање на потребните молекули до места со мала концентрација од делови со висока концентрација. Типичен пример е транспортот на кислород и јаглерод диоксид низ органите и ткивата преку протеинот хемоглобин. Тие исто така вршат испорака на соединенија со мала молекуларна тежина преку клеточната мембрана во внатрешноста.
- Структурна функција. Една од најважните функции што ја извршуваат протеините. Структурата на сите клетки и нивните органели е обезбедена со пептиди. Тие, како рамка, ја поставуваат формата и структурата. Покрај тоа, тие го поддржуваат и го менуваат доколку е потребно. Затоа, за раст и развој, сите живи организми бараат протеини во нивната исхрана. Таквите пептиди вклучуваат еластин, тубулин, колаген, актин, кератин и други.
- Каталитичка функција. Се изведува од ензими. Бројни и разновидни, тие ги забрзуваат сите хемиски и биохемиски реакции во телото. Без нивно учество, обично јаболко во желудникот би можело да се свари за само два дена, а најверојатно ќе изгние во тој процес. Под влијание на каталаза, пероксидаза и други ензими, овој процес се случува за два часа. Во принцип, благодарение на оваа улога на протеините се вршат анаболизмот и катаболизмот, односно пластика и

Заштитна улога
Постојат неколку видови на закани од кои протеините се дизајнирани да го заштитат телото.
Прво, трауматски реагенси, гасови, молекули, супстанции од различен спектар на дејство. Пептидите се способни да комуницираат хемиски со нив, претворајќи ги во безопасна форма или едноставно неутрализирајќи ги.
Второ, физичката закана од рани - ако протеинот фибриноген не се трансформира навреме во фибрин на местото на повредата, тогаш крвта нема да се згрутчи, што значи дека нема да дојде до блокада. Потоа, напротив, ќе ви треба пептид плазмин, кој може да го раствори згрутчувањето и да ја врати проодноста на садот.
Трето, закана за имунитетот. Структурата и значењето на протеините кои формираат имунолошка одбрана се исклучително важни. Антитела, имуноглобулини, интерферони - сето тоа се важни и значајни елементи на човечкиот лимфен и имунолошки систем. Секоја странска честичка, штетна молекула, мртов дел од клетка или цела структура е предмет на итно испитување од пептидното соединение. Затоа човек може самостојно, без помош на лекови, секојдневно да се заштити од инфекции и едноставни вируси.
Физички својства
Структурата на клеточниот протеин е многу специфична и зависи од извршената функција. Но, физичките својства на сите пептиди се слични и се сведуваат на следните карактеристики.
- Тежината на молекулата е до 1.000.000 Далтони.
- Колоидните системи се формираат во воден раствор. Таму структурата добива полнеж што може да варира во зависност од киселоста на околината.
- Кога се изложени на сурови услови (зрачење, киселина или алкали, температура итн.) тие се способни да преминат на други нивоа на конформации, односно денатурација. Овој процес е неповратен во 90% од случаите. Сепак, постои и обратно поместување - ренатурација.
Ова се главните својства на физичките карактеристики на пептидите.
Станува збор за високомолекуларни органски соединенија, биополимери, изградени од 20 видови на остатоци од L-?-аминокиселини поврзани во одредена низа во долги синџири. Молекуларната тежина на протеините варира од 5 илјади до 1 милион. Името „белки“ за прв пат е дадено на супстанцијата на птичји јајца, која се коагулира кога се загрева во бела нерастворлива маса. Терминот подоцна беше проширен на други супстанции со слични својства изолирани од животни и растенија.
Ориз. 1. Најсложени биополимери се протеините. Нивните макромолекули се состојат од мономери, кои се амино киселини. Секоја аминокиселина има две функционални групи: карбоксилна група и амино група. Целата разновидност на протеините се создава како резултат на различни комбинации на 20 амино киселини.
Протеините доминираат над сите други соединенија присутни во живите организми, обично сочинуваат повеќе од половина од нивната сува тежина. Се претпоставува дека во природата има неколку милијарди индивидуални протеини (на пример, повеќе од 3 илјади различни протеини се присутни само во бактеријата E. coli).
Протеините играат клучна улога во животните процеси на секој организам. Протеините вклучуваат ензими, со чие учество се случуваат сите хемиски трансформации во клетката (метаболизам); тие го контролираат дејството на гените; со нивно учество се реализира дејството на хормоните, се врши трансмембрански транспорт, вклучително и генерирање на нервни импулси. Тие се составен дел на имунолошкиот систем (имуноглобулини) и системот за коагулација, ја формираат основата на коскеното и сврзното ткиво и се вклучени во трансформацијата и искористувањето на енергијата.
Историја на истражување на протеини
Првите обиди да се изолираат протеините беа направени уште во 18 век. До почетокот на 19 век, се појавија првите дела за хемиско проучување на протеините. Француските научници Жозеф Луис Геј-Лусак и Луис Жак Тенар се обидоа да го утврдат елементарниот состав на протеините од различни извори, што го означи почетокот на систематските аналитички студии, благодарение на кои беше заклучено дека сите протеини се слични во множеството елементи вклучени во нивниот состав. Во 1836 година, холандскиот хемичар G. J. Mulder ја предложил првата теорија за структурата на протеинските супстанции, според која сите протеини имаат одреден хипотетички радикал (C 40 H 62 N 10 O 12), поврзан во различни пропорции со атоми на сулфур и фосфор. Тој го нарече овој радикален „протеин“ (од грчкиот протеин - прво, главен). Теоријата на Мулдер придонесе за зголемен интерес за проучување на протеините и подобрување на методите на хемијата на протеините. Развиени се техники за изолирање на протеини со екстракција со раствори на неутрални соли и за прв пат се добиени протеини во кристална форма (некои растителни протеини). За да ги анализираат протеините, тие почнаа да го користат нивното прелиминарно варење со киселини и алкалии.
Во исто време, зголеменото внимание почна да се посветува на проучувањето на функцијата на протеините. Јенс Јакоб Берзелиус бил првиот што во 1835 година предложил да ја играат улогата на биокатализатори. Наскоро беа откриени протеолитички ензими - пепсин (Т. Шван, 1836) и трипсин (Л. Корвисарт, 1856), кои привлекоа внимание на физиологијата на варењето и анализата на производите формирани за време на распаѓањето на хранливите материи. Понатамошните студии за структурата на протеините и работата на хемиската синтеза на пептидите резултираа со појава на хипотеза за пептиди, според која сите протеини се изградени од амино киселини. До крајот на 19 век, повеќето од амино киселините кои ги сочинуваат протеините беа проучувани.
На почетокот на 20 век, германскиот хемичар Емил Херман Фишер бил првиот што ги користел методите на органска хемија за проучување на протеините и докажал дека протеините се состојат од β-амино киселини поврзани едни со други со амид (пептидна) врска. Подоцна, благодарение на употребата на физичко-хемиски методи на анализа, беше утврдена молекуларната маса на многу протеини, беше воспоставена сферична форма на глобуларните протеини, спроведена е рендгенска дифракциона анализа на амино киселини и пептиди и беа направени методи на хроматографска анализа. развиена (види хроматографија).
Првиот протеински хормон беше изолиран (Фредерик Грант Бантинг, Џон Џејмс Рикард Меклаод, 1922), беше докажано присуството на гама глобулини во антителата и опишана е ензимската функција на мускулниот протеин миозин (Владимир Александрович Енгелхард, М. Н. Љубимова, 1939). . За прв пат се добиени ензими во кристална форма - уреаза (J.B. Saliner, 1926), пепсин (J.H. Nortron, 1929), лизозим (E.P. Abraham, Robert Robinson, 1937).

Ориз. 2. Шема на тродимензионална структура на ензимот лизозим. Кругови - амино киселини; нишки - пептидни врски; засенчените правоаголници се дисулфидни врски. Видливи се спирализирани и издолжени делови од полипептидниот синџир.
Во 1950-тите, беше докажана организација на три нивоа на протеинските молекули - присуство на примарна, секундарна и терцијарна структура; создаде автоматски аминокиселински анализатор (Стенфорд Мур, Вилијам Хауард Стајн, 1950). Во 60-тите, беа направени обиди за хемиска синтеза на протеини (инсулин, рибонуклеаза). Значително се подобрени методите за анализа на дифракција на Х-зраци; беше создаден уред - секвенсер (П. Едман, Г. Бег, 1967), што овозможи да се одреди низата на амино киселини во полипептиден синџир. Последица на ова беше воспоставувањето на структурата на неколку стотици протеини од различни извори. Меѓу нив се протеолитички ензими (пепсин, трипсин, химотрипсин, субтилизин, карбоксипептидази), миоглобини, хемоглобини, цитохроми, лизозими, имуноглобулини, хистони, невротоксини, вирусни обвивни протеини, протеин-пепти. Како резултат на тоа, се појавија предуслови за решавање на итни проблеми во ензимологијата, имунологијата, ендокринологијата и другите области од биолошката хемија.
На крајот на 20 век, постигнат е значителен напредок во проучувањето на улогата на протеините во синтезата на матрицата на биополимерите, разбирањето на механизмите на нивното дејство во различните животни процеси на организмите и воспоставувањето на врската помеѓу нивната структура и функција. Од големо значење беше подобрувањето на методите на истражување и појавата на нови методи за одвојување на протеините и пептидите.
Развојот на ефективен метод за анализа на низата на нуклеотиди во нуклеинските киселини овозможи значително да се поедностави и забрза одредувањето на амино киселинската секвенца во протеините. Се покажа дека ова е можно бидејќи редоследот на аминокиселините во протеинот се одредува според низата на нуклеотиди во генот што го кодира овој протеин (фрагмент). Следствено, знаејќи го распоредот на нуклеотидите во овој ген и генетскиот код, може точно да се предвиди по кој редослед се наоѓаат амино киселините во полипептидниот синџир на протеинот. Заедно со напредокот во структурната анализа на протеините, постигнати се значајни резултати во проучувањето на нивната просторна организација, механизмите на формирање и дејство на супрамолекуларните комплекси, вклучувајќи ги рибозомите и другите клеточни органели, хроматинот, вирусите итн.
Структура на протеини
Речиси сите протеини се изградени од 20 α-амино киселини кои припаѓаат на серијата L и се исти кај скоро сите организми. Амино киселините во протеините се поврзани една со друга со пептидна врска -CO-NH-, која е формирана од карбоксилната и -амино групата на соседните амино киселински остатоци: две амино киселини формираат дипептид во кој терминалниот карбоксил (-COOH) а амино групата (H2N-) остануваат слободни, на која може да се додадат нови амино киселини за да се формира полипептиден синџир.
Делот од синџирот на кој се наоѓа терминалот H 2 N-група се нарекува N-терминал, а делот спроти него се нарекува C-терминал. Огромната разновидност на протеини е одредена од редоследот на распоредот и бројот на остатоци од аминокиселини што ги содржат. Иако не постои јасна разлика, кратките синџири обично се нарекуваат пептиди или олигопептиди (од олиго...), а полипептидите (протеини) обично се сфаќаат како синџири што се состојат од 50 или повеќе. Најчести протеини се оние кои содржат 100-400 остатоци од аминокиселини, но има и такви чии молекули се формираат од 1000 или повеќе остатоци. Протеините може да се состојат од неколку полипептидни синџири. Во таквите протеини, секој полипептиден синџир се нарекува подединица.
Просторна структура на протеините

Ориз. 3. Протеините во сите организми се состојат од 20 видови на амино киселини. Секој протеин се карактеризира со одреден асортиман и квантитативен сооднос на амино киселини. Во протеинските молекули, аминокиселините се поврзани една со друга со пептидни врски (- CO - NH -) во линеарна низа, што ја сочинува таканаречената примарна структура на протеинот. Горна линија - слободни амино киселини со странични групи R1, R2, R3; крајна линија - амино киселините се поврзани со пептидни врски.
Полипептидниот синџир е способен спонтано да формира и одржува посебна просторна структура. Врз основа на обликот на протеинските молекули, протеините се поделени на фибриларни и глобуларни. Во глобуларните протеини, еден или повеќе полипептидни синџири се превиткуваат во компактна сферична структура, или топка. Обично овие протеини се многу растворливи во вода. Тие ги вклучуваат речиси сите ензими, протеини за транспорт на крв и многу протеини за складирање. Фибриларните протеини се молекули слични на нишки кои се држат заедно со вкрстени врски и формираат долги влакна или слоевити структури. Имаат висока механичка сила, нерастворливи во вода и вршат главно структурни и заштитни функции. Типични претставници на таквите протеини се кератините за коса и волна, свилен фиброин и колаген на тетивите.
Редоследот на ковалентно поврзани амино киселини во полипептидниот синџир се нарекува амино киселинска секвенца или примарна структура на протеините. Примарната структура на секој протеин, кодирана од соодветниот ген, е константна и ги носи сите информации потребни за формирање на структури на повисоко ниво. Потенцијалниот број на протеини што може да се формираат од 20 амино киселини е практично неограничен.
Како резултат на интеракцијата на страничните групи на амино киселински остатоци, поединечни релативно мали делови од полипептидниот синџир добиваат една или друга конформација (тип на преклопување), позната како секундарна структура на протеините. Неговите најкарактеристични елементи се периодично повторувачката α-спирала и β-структурата. Секундарната структура е многу стабилна. Бидејќи во голема мера е одредена од амино киселинската секвенца на соодветниот протеински регион, станува возможно да се предвиди со одреден степен на веројатност. Терминот „?-хеликс“ беше воведен од американскиот биохемичар, физичар и хемичар Линус Карл Полинг, кој го опиша распоредот на полипептидниот синџир во протеинот?-кератин во форма на десна спирала (?-спиралата може да да се спореди со телефонски кабел). За секое вртење на таква спирала во протеинот има 3,6 остатоци од аминокиселини. Ова значи дека -C=O групата од една пептидна врска формира водородна врска со -NH групата на друга пептидна врска, четири амино киселински остатоци оддалечени од првата. Во просек, секој α-спирален регион вклучува до 15 амино киселини, што одговара на 3-4 вртења на спиралата. Но, во секој поединечен протеин, должината на спиралата може многу да се разликува од оваа вредност. Во пресек, α-спиралата има форма на диск, од кој страничните синџири на амино киселини се насочени кон надвор.
Структура или? - преклопен слој, може да се формира од неколку делови од полипептидниот синџир. Овие делови се испружени и поставени паралелно еден на друг, поврзани едни со други со водородни врски што се јавуваат помеѓу пептидните врски. Тие можат да бидат ориентирани во исти или спротивни насоки (насоката на движење по полипептидниот синџир обично се смета дека е од N-крајот до C-крајот). Во првиот случај, преклопениот слој се нарекува паралелен, во вториот - антипаралелен. Вториот се формира кога пептидниот синџир нагло се врти назад, формирајќи свиок (?-свиткување). Дали страничните синџири на амино киселините се ориентирани нормално на рамнината? -Слој.
Релативна содржина? -спирални пресеци и? -Структурите може да варираат во голема мера меѓу различните протеини. Постојат протеини со доминација на α-спирали (околу 75% од амино киселините во миоглобинот и хемоглобинот), а главниот тип на преклопување на синџирот во многу фибриларни протеини (вклучувајќи свилен фиброин, β-кератин) е α-хеликс. -структура. Регионите на полипептидниот синџир кои не можат да се класифицираат во ниту една од гореопишаните конформации се нарекуваат поврзувачки јамки. Нивната структура е одредена главно од интеракциите помеѓу страничните синџири на амино киселините, а во молекулата на кој било протеин се вклопува на строго дефиниран начин.
Терциерната структура се нарекувапросторна структура на глобуларни протеини. Но, често овој концепт се однесува на методот на преклопување на полипептидниот синџир во вселената, карактеристичен за секој специфичен протеин. Терциерната структура е формирана од полипептидниот синџир на протеин спонтано, очигледно, по одредена патека(и) на коагулација со прелиминарно формирање на секундарни структурни елементи. Ако стабилноста на секундарната структура се должи на водородни врски, тогаш терциерната структура е фиксирана со различен систем на не-ковалентни интеракции: водородни, јонски, интермолекуларни интеракции, како и хидрофобни контакти помеѓу страничните синџири на неполарните амино киселински остатоци.
Кај некои протеини, терциерната структура дополнително се стабилизира со формирање на дисулфидни врски (-S-S- врски) помеѓу остатоците од цистеин. Како по правило, во внатрешноста на протеинската топка има странични синџири на хидрофобни амино киселини собрани во јадрото (нивниот трансфер во протеинската топка е термодинамички поволен), а на периферијата има хидрофилни остатоци и некои хидрофобни. Протеинската топка е опкружена со неколку стотици молекули на хидратациона вода, која е неопходна за стабилноста на протеинската молекула и често е вклучена во неговото функционирање. Терциерната структура е мобилна, нејзините поединечни делови можат да се поместат, што доведува до конформациски транзиции кои играат значајна улога во интеракцијата на протеинот со другите молекули.
Терциерната структура е основата на функционалните својства на протеинот. Го одредува формирањето на ансамбли на функционални групи во протеинот - активни центри и врзувачки зони, им ја дава потребната геометрија, овозможува создавање на внатрешно опкружување, што е предуслов за појава на многу реакции и обезбедува интеракција со други протеини. .
Терциерната структура на протеините јасно одговара на нејзината примарна структура; веројатно постои уште недешифриран стереохемиски код кој ја одредува природата на преклопувањето на протеините. Сепак, еден ист метод на уредување на просторот обично не одговара на една примарна структура, туку на цело семејство на структури во кои само мал дел (до 20-30%) од остатоците од амино киселини може да се совпаднат, но во одредени места во синџирот сличноста на амино киселинските остатоци е зачувана. Резултатот е формирање на големи семејства на протеини кои се карактеризираат со слична терцијарна и повеќе или помалку слична примарна структура и, по правило, заедничка функција. Тоа се, на пример, протеини на организми од различни видови кои имаат иста функција и се еволутивно поврзани: миоглобини и хемоглобини, трипсин, химотрипсин, еластаза и други животински протеинази.

Ориз. 4. Како резултат на комбинацијата на неколку протеински макромолекули со терциерна структура, кватернарна протеинска структура се формира во комплексен комплекс. Пример за такви сложени протеини е хемоглобинот, кој се состои од четири макромолекули.
Често, особено кај големите протеини, преклопувањето на полипептидниот синџир се случува преку формирање од поединечни делови од синџирот на повеќе или помалку автономни елементи на просторната структура - домени кои можат да имаат функционална автономија, одговорни за една или друга биолошка активност на протеини. Така, N-терминалните домени на протеините за коагулација на крвта обезбедуваат нивно прицврстување на клеточната мембрана.
Постојат многу протеини чии молекули се ансамбл од глобули (подединици) кои се држат заедно со хидрофобни интеракции, водородни или јонски врски. Таквите комплекси се нарекуваат олигомерни, мултимерни или подединици протеини. Распоредот на подединици во функционално активен протеински комплекс се нарекува кватернарна структура на протеинот. Некои протеини се способни да формираат структури од повисоки редови, на пример, мултиензимски комплекси, проширени структури (протеини на обвивката на бактериофагот), супрамолекуларни комплекси кои функционираат како една целина (на пример, рибозоми или компоненти на митохондријалниот респираторен синџир).
Кватернарната структура овозможува создавање на молекули со необични геометрии. Така, феритинот, формиран од 24 подединици, има внатрешна празнина, благодарение на која протеинот успева да врзе до 3000 железни јони. Покрај тоа, кватернарната структура овозможува извршување на неколку различни функции во една молекула. Триптофан синтетаза комбинира ензими одговорни за неколку последователни фази на синтезата на аминокиселината триптофан.
Методи за проучување на структурата на протеините
Примарната структура на протеините ги одредува сите други нивоа на организација на протеинската молекула. Затоа, кога се проучува биолошката функција на различни протеини, важно е знаењето за оваа структура. Првиот протеин за кој беше воспоставена аминокиселинската секвенца беше панкреасниот хормон, инсулин. Оваа работа, која траеше 11 години, ја изврши англискиот биохемичар Фредерик Сангер (1954). Тој ја утврди локацијата на 51 аминокиселина во молекулата на хормонот и покажа дека таа се состои од 2 синџири поврзани со дисулфидни врски. Подоцна, поголемиот дел од работата за воспоставување на примарната структура на протеините беше автоматизирана.
Со развојот на методите на генетски инженеринг, стана можно дополнително да се забрза овој процес со одредување на примарната структура на протеините во согласност со резултатите од анализата на нуклеотидната секвенца во гените што ги кодираат овие протеини. Секундарната и терциерната структура на протеините се изучува со користење на прилично сложени физички методи, на пример, кружен дихроизам или рендгенска дифракциона анализа на протеинските кристали. Терциерната структура за прв пат беше воспоставена од англискиот биохемичар Џон Каудри Кендру (1957) за мускулниот протеин миоглобин.

Ориз. 5. Модел на молекулата на миоглобинот (просторна конфигурација на молекулата)
Денатурација на протеини
Релативно слабите врски одговорни за стабилизирање на секундарните, терциерните и кватернерните структури на протеинот лесно се уништуваат, што е придружено со губење на неговата биолошка активност. Уништувањето на првобитната (матична) протеинска структура, наречена денатурација, се случува во присуство на киселини и бази, со загревање, промени во јонската јачина и други влијанија. Како по правило, денатурираните протеини се слабо или воопшто не се растворливи во вода. Со краткотраен ефект и брза елиминација на денатурираните фактори, можна е ренатурација на протеините со целосно или делумно враќање на првобитната структура и биолошките својства.
Класификација на протеини
Сложеноста на структурата на протеинските молекули и екстремната разновидност на функции што тие ги извршуваат го отежнуваат создавањето на унифицирана и јасна класификација на нив, иако обидите да се направи тоа постојано се правени од крајот на 19 век. Врз основа на нивниот хемиски состав, протеините се делат на едноставни и сложени (понекогаш наречени протеиди). Молекулите на првите се состојат само од амино киселини. Покрај самиот полипептиден синџир, комплексните протеини содржат непротеински компоненти претставени со јаглехидрати (гликопротеини), липиди (липопротеини), нуклеински киселини (нуклеопротеини), метални јони (металопротеини), фосфатна група (фосфопротеини), пигменти (хромопротеини), итн.
Во зависност од функциите што ги извршуваат, се разликуваат неколку класи на протеини. Најразновидна и најспецијализирана класа се состои од протеини со каталитичка функција - ензими кои имаат способност да ги забрзуваат хемиските реакции што се случуваат во живите организми. Во овој капацитет, протеините учествуваат во сите процеси на синтеза и разградување на различни соединенија за време на метаболизмот, во биосинтезата на протеините и нуклеинските киселини, регулирањето на развојот и диференцијацијата на клетките. Транспортните протеини имаат способност селективно да ги врзуваат масните киселини, хормоните и другите органски и неоргански соединенија и јони, а потоа да ги транспортираат со струја до саканата локација (на пример, хемоглобинот е вклучен во трансферот на кислород од белите дробови до сите клетки на телото). Транспортните протеини исто така вршат активен транспорт на јони, липиди, шеќери и амино киселини низ биолошките мембрани.
Структурните протеини вршат потпорна или заштитна функција; тие учествуваат во формирањето на клеточниот скелет. Најчести меѓу нив се колагенот на сврзното ткиво, кератинот, ноктите и пердувите, еластинот на васкуларните клетки и многу други. Во комбинација со липидите, тие се структурна основа на клеточните и интрацелуларните мембрани.
Голем број на протеини вршат заштитна функција. На пример, имуноглобулините (антитела) на 'рбетниците, кои имаат способност да ги врзуваат странските патогени микроорганизми и супстанции, да ги неутрализираат нивните патогени ефекти врз телото и да го спречат размножувањето на клетките. Фибриногенот и тромбинот се вклучени во процесот на згрутчување на крвта. Многу протеински супстанции кои се излачуваат од бактерии, како и компоненти на некои безрбетници, се класифицирани како токсини.
Некои протеини (регулаторни) се вклучени во регулирањето на физиолошката активност на телото како целина, поединечни органи, клетки или процеси. Тие ја контролираат транскрипцијата на гените и синтезата на протеините; тие вклучуваат пептидно-протеински хормони кои се излачуваат од ендокрините жлезди. Протеините за складирање на семиња обезбедуваат хранливи материи за почетните фази на развојот на ембрионот. Тие исто така вклучуваат казеин, албумин од белка од јајце (овалбумин) и многу други. Благодарение на протеините, мускулните клетки стекнуваат способност да се контрахираат и на крајот обезбедуваат движење на телото. Примери за такви контрактилни протеини се актинот и миозинот на скелетните мускули, како и тубулинот, кои се компоненти на цилиите и флагелите на едноклеточните организми; Тие исто така обезбедуваат дивергенција на хромозомите за време на клеточната делба.
Рецепторските протеини се цел на хормоните и другите биолошки активни соединенија. Со нивна помош, клетката согледува информации за состојбата на надворешното опкружување. Тие играат важна улога во пренесувањето на нервната возбуда и во ориентираното движење на клетките (хемотаксија). Трансформацијата и искористувањето на енергијата што влегува во телото, како и енергијата, се случува и со учество на протеини од биоенергетскиот систем (на пример, визуелниот пигмент родопсин, цитохромите на респираторниот синџир). Исто така, има многу протеини со други, понекогаш прилично невообичаени функции (на пример, плазмата на некои риби на Антарктикот содржи протеини кои имаат антифризни својства).
Биосинтеза на протеини
Сите информации за структурата на одреден протеин се „складираат“ во соодветните гени во форма на низа нуклеотиди и се имплементираат во процесот на синтеза на шаблоните. Прво, информациите се пренесуваат (читаат) од молекулата на ДНК до гласник РНК (мРНК) со помош на ензимот ДНК зависна РНК полимераза, а потоа во рибозомот на мРНК, како на матрица во согласност со генетскиот код, со учество на транспорт РНК доставување амино киселини, формирање се јавува полипептид синџир.
Синтетизираните полипептидни синџири кои излегуваат од рибозомот, спонтано се преклопуваат, ја добиваат конформациската карактеристика на протеинот и можат да бидат предмет на пост-транслациска модификација. Страничните синџири на поединечни амино киселини можат да претрпат модификации (хидроксилација, фосфорилација, итн.). Затоа, на пример, хидроксипролин и хидроксилизин се наоѓаат во колагенот (види). Модификацијата може да биде придружена и со прекин на полипептидните врски. На овој начин, на пример, доаѓа до формирање на активна инсулинска молекула која се состои од два синџири поврзани со дисулфидни врски.

Ориз. 6. Општа шема на биосинтеза на протеини.
Важноста на протеините во исхраната
Протеините се најважните компоненти на животинската и човечката храна. Хранливата вредност на протеините се одредува според нивната содржина на есенцијални амино киселини, кои не се произведуваат во самото тело. Во овој поглед, растителните протеини се помалку вредни од животинските: тие се посиромашни со лизин, метионин и триптофан и потешко се вари во гастроинтестиналниот тракт. Недостатокот на есенцијални амино киселини во храната доведува до сериозни нарушувања на метаболизмот на азот.
Протеините се разградуваат на слободни амино киселини, кои по апсорпцијата во цревата влегуваат и се дистрибуираат до сите клетки. Некои од нив се разградуваат на едноставни соединенија со ослободување на енергија, кои се користат за различни потреби на клетката, а некои одат во синтеза на нови протеини карактеристични за даден организам. (Р. А. Матвеева, Енциклопедија Кирил и Методиј)
Набројување на протеини
- амилоид - амилоид;
- анјонски - анјонски;
- антивирус - антивирусно;
- автоимуни - автоимуни;
- автологен - автологичен;
- бактериски - бактериски;
- Бенс Џонс протеин;
- вирусно индуцирано - вирусно индуцирано;
- вирусна - вирусна;
- вирусни неструктурни - вирусни неструктурни;
- вирусни структурни - вирусни структурни;
- вирус-специфичен - вирус специфичен;
- висока молекуларна тежина - висока молекуларна тежина;
- што содржи хем - хем;
- хетерологно - странско;
- хибрид - хибрид;
- гликолизиран - гликозиран;
- топчеста - топчеста;
- денатуриран - денатуриран;
- што содржи железо - железо;
- жолчка - жолчка;
- животински протеини - животински протеини;
- заштитно - одбранбено;
- имун - имун;
- имуноген - имунолошки релевантен;
- врзување на калциум;
- кисело - кисело;
- корпускуларен - корпускуларен;
- мембрана - мембрана;
- миелом - миелом;
- микрозомално - микрозомално;
- млечен протеин - млечен протеин;
- моноклонален - моноклонален имуноглобулин;
- мускулен протеин - мускулен протеин;
- мајчин - мајчин;
- нонхистон - нехистон;
- неисправни - делумно;
- нерастворлив - нерастворлив;
- несварлив - нерастворлив;
- неензимски - неензимски;
- ниска молекуларна тежина - мала молекуларна тежина;
- нов протеин - нов протеин;
- општо - целина;
- онкоген - онкопротеин;
- протеин на главната фаза - анјонски;
- протеин на акутна фаза (воспаление) - протеин на акутна фаза;
- храна - храна;
- протеин на крвната плазма - плазма протеин;
- плацентарна - плацента;
- разврзување - разврзување;
- протеин на регенерирачки нерв;
- регулаторна - регулаторна;
- рекомбинација - рекомбинантна;
- рецептор - рецептор;
- рибозомално - рибозомално;
- обврзувачки - обврзувачки;
- секреторен протеин - секреторен протеин;
- C-реактивен - C-реактивен;
- протеин од сурутка - протеин од сурутка, лактопротеин;
- ткиво - ткиво;
- токсичен - токсичен;
- химеричен - химеричен;
- целина - целина;
- цитосолен - цитозолен;
- алкален протеин - анјонски протеин;
- егзоген - егзоген;
- ендогени - ендогени протеин.
Прочитајте повеќе за протеините во литературата:
- Volkenshtein M.V., Molecules and, M., 1965, ch. 3 - 5;
- Гауровиц Ф., Хемија и функции на протеините, транс. од англиски, Москва, 1965;
- Sissakyan N. M. и Gladilin K. L., Биохемиски аспекти на синтезата на протеини, во книгата: Напредоци во биолошката хемија, том 7, М., 1965, стр. 3;
- Степанов В.М. Молекуларна биологија. Структура и функција на протеините. М., 1996;
- Шамин А.Н., Развој на хемија на протеини, М., 1966;
- Протеини и пептиди. М., 1995-2000 година. Т. 1-3;
- Биосинтеза на протеини и нуклеински киселини, ед. А. С. Спирина, М., 1965;
- Вовед во молекуларна биологија, транс. од англиски, М., 1967 г
- Молекули и клетки. [саб. чл.], транс. од англиски, М., 1966, стр. 7 - 27, 94 - 106;
- Основи на биохемија: Превод од англиски М., 1981. Т. 1;
- Проблемот со протеините. М., 1995. Т. 1-5;
- Протеините. Њујорк, 1975-79. 3 ед. V. 1-4.
Најдете нешто друго интересно:







