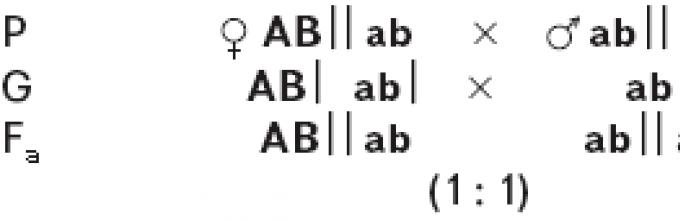
32. Особености на наследување на особина со нецелосно и целосно спојување
ПОВРЗАНО НАСЛЕДУВАЊЕ НА КАРАКТЕРИСТИКИ
Се запознавме со дихибридното вкрстување и сфативме дека независната комбинација на особини се објаснува со фактот дека разделувањето на еден пар алелни гени кои ги одредуваат соодветните особини се случува независно од другиот пар. Сепак, ова е забележано само во случај кога гените од различни парови се наоѓаат во различни парови на хромозоми и кога хибридните герминативни клетки се формираат во мејоза, татковските и мајчините хромозоми се независно комбинирани. Но, бројот на хромозоми е многу ограничен во споредба со бројот на знаци, од кои секоја се развива под контрола на специфичен ген. Така, во Дрософила се познати околу 7000 гени со четири пара хромозоми. Се претпоставува дека едно лице има најмалку 50 илјади гени со 23 пара хромозоми итн. Следи дека стотици алели мора да бидат локализирани во секој пар на хромозоми. Природно; дека постои поврзаност помеѓу гените кои се наоѓаат на истиот хромозом и кога се формираат герминативните клетки, тие мора да се пренесат заедно.
Поврзаното наследство беше откриено во 1906 година од страна на англиските генетичари В. Батсон и Р. Пунет додека го проучуваа наследството на особините кај слаткиот грашок, но тие не беа во можност да ги откријат причините за овој феномен. Природата на поврзаното наследство беше разјаснета во 1910 година од страна на научниците Т. Морган и неговите соработници К. Бриџис и А. Стуртевант. Како предмет на проучување, тие ја избраа овошната мушичка Drosophila, која се покажа дека е многу погодна за генетски експерименти. Клетките на телото Drosophila содржат 4 пара хромозоми. Се одликува со многу висока плодност - еден пар дава повеќе од сто потомци. Има голема брзина на развој - во рок од 12-15 дена по оплодувањето, од јајцето се развиваат ларва, кукличка и возрасна личност, која речиси веднаш е способна да произведе потомство. Во текот на една година може да се проучуваат повеќе од дваесет генерации. Мувите имаат сива боја, со црвени очи. се мали по големина (околу 3 mm), лесно се разредуваат во биолошки епрувети; Можете да користите лупи за да ги проучите нивните знаци. Кога гледал стотици илјади индивидуи, Морган открил многу различни мутации: имало муви со црно и жолто тело, со бели и други обоени очи, со променета форма и положба на крилјата итн. Понекогаш имало и поединци со неколку мутации одеднаш, на пример црно тело, рудиментирани крилја, цинабарски очи.
Проучувајќи го наследството на различни парови знаци за време на дихибридни и полихибридни вкрстувања, Морган и неговите колеги открија голем број примери на нивното поврзано (заедничко) наследување. Сите проучувани знаци беа поделени во четири групи за поврзување во согласност со бројот и големината на хромозомите во Drosophila. Врз основа на тоа, Морган заклучил дека гените кои ги одредуваат овие особини се наоѓаат на хромозомите. Гените лоцирани на истиот хромозом сочинуваат група за поврзување.
Генската поврзаност е заедничко наследување на гените лоцирани на истиот хромозом.Бројот на групи за поврзување одговара на хаплоидниот број на хромозоми. На пример, Drosophila има 4 групи на поврзување, луѓето имаат 23, говедата имаат 30, свињите имаат 19 итн.
Идејата за локацијата на гените во хромозомите беше изразена од Сетон во 1902 година. Понатамошното истражување спроведено од Морган покажа дека поврзаноста на гените лоцирани на истиот хромозом може да биде целосна или нецелосна.
Разликата во однесувањето на поврзаните и независно наследените гени најјасно се открива при спроведување на аналитички вкрстување. Со независно наследство на два пара знаци во хибриден Fi (АаББ)Со еднаква веројатност се формираат 4 типа гамети: АБ,Аб, aB, ab. Кога се вкрстува со целосен рецесивен (аабб) бројот на сорти на гамети во хибридот го одредува бројот на типови на потомци и истата веројатност за нивно појавување, бидејќи гаметите на рецесивна индивидуа (ab) не може да ги промени манифестациите на доминантните и рецесивни гени на хибридните гамети. Како резултат на тоа, односот на фенотиповите на потомството ќе биде 1:1:1:1. Ако двата пара алелни гени се наоѓаат во еден пар на хромозоми, тогаш за време на формирањето на герминативните клетки гените на овие алели нема да можат слободно да се комбинираат. Во овој случај, се забележува поврзано наследство.
ПОЛНА СУПЈА
Т. Морган вкрсти црни долгокрилни женки со сиви мажјаци со рудиментирани крилја. Во Drosophila, сивата боја на телото доминира над црната, а долгите крилја доминираат над рудиментираните крилја. Дозволете ни да го означиме генот за сива боја на телото ВО,алелен ген за црна боја на телото б; Longwing™ ген В, алеличен на него е генот за рудиментирани крилја v. Двата пара од овие гени се наоѓаат на истиот втор пар на хромозоми. За двата пара Карактеристики, родителските форми беа хомозиготни: женски за рецесивната карактеристика на црно тело (бб) и доминантната особина долги крила™ (В.В.), машки со доминантна сива боја. (ББ)и рецесивната особина на ембрионските крилја (w>. Гаметите на родителите при редукциската поделба добиваат од мајката хромозом со гени б И В, од татковиот - со гени ВОи v. Сите потомци од првата генерација (Fi) имаа сиво тело и долги крилја (сл. 13) и беа хетерозиготни за двата пара особини (bV/ Bv). Потоа, мажјаците беа избрани од Fi и вкрстени со женки хомозиготни за двата рецесивни гени, црните зачетоци (bv/ bv), т.е., беше извршено аналитичко вкрстување, како резултат на што, со независна комбинација на знаци, требаше да се добие потомство од четири фенотипови во еднакви размери: сиво долгокрилести, сиви со рудиментирани крилја, црни долгокрилести, црна со рудиментирани крилја, но се добиени потомци на само два фенотипа, слични на оригиналните родителски форми: црни долгокрилести и сиви кратки крилја. Во овој случај, се забележува целосна конкатенација на карактеристиките. Ова се должи на фактот дека кај хетерозиготниот маж, и генот за црно обојување и генот за долги крилја се наоѓаат на истиот хромозом од хомологен пар, а генот за сива боја и генот за рудиментирани крилја се наоѓаат на другиот хромозом.
За време на спермиогенезата за време на мејозата, хомологните хромозоми се одвојуваат во различни герминативни клетки. Се произведуваат само два вида гамети: едната со хромозом што носи гени КомерсантИ В, другиот со хромозомот на кој се наоѓаат гените ВОи v. Кога овие гамети се комбинираат со гамети на поединец со рецесивни особини, се формираат само два типа на потомство. Во целосна поврзаност, гените лоцирани на истиот хромозом секогаш се пренесуваат заедно.Целосна поврзаност досега е воспоставена само кај овошните мушички и женските свилени буби.
НЕЦОЛНА СУПЈКА
Во следниот експеримент, како и во претходниот, Морган вкрстил црни долгокрилни женки со сиви мажјаци со рудиментирани крила. Во првата генерација, сите потомци беа сиви долги крила. Потоа повторно направил аналитички крст, но од првата генерација избрал не машко, туку женско и ја прекрстил со црн мажјак со рудиментирани крилја (сл. 14). Во овој случај, потомството не се појавило од два вида, како со целосна поврзаност, туку од четири: сиво со рудиментирани крилја, црно долгокрилести, сиво долгокрилести и црни со рудиментирани крилја, но не во еднакви размери, како со независна комбинација на знаци, но со значителна доминација фенотипови слични на родителските форми. 41,5% од мувите биле сиви со рудиментирани крилја, како едниот оригинален родител, а 41,5% биле црни долгокрилести, како и другиот оригинален родител. Само 17% од потомците се родени со нова комбинација на карактеристики: 8,5% биле црни со рудиментирани крилја и 8,5% биле сиви долги крилја. Така, 83% од потомците имале комбинација на особини како првобитните родителски форми, но се појавиле и поединци со нова комбинација на особини. Затоа, спојката е нецелосни./ Се постави прашањето: зошто се појавија поединци со нова комбинација на родителски карактеристики? За да го објасни овој феномен, Морган ја искористи и ја разви теоријата хијазмотиповиБелгиски цитолог Јансенс. Во 1909 година, Јансенс забележал дека за време на спермиогенезата во саламандерот во профазата на мејозата, хомологните хромозоми се конјугираат, а потоа, на почетокот на дивергенцијата, формираат фигури во делови за размена на хромозомите.Ако поврзаните гени лежат на истиот хромозом и рекомбинацијата на овие гени се случува кај хетерозиготите за време на формирањето на гамети, тоа значи дека хомологните хромозоми ги разменувале своите делови за време на мејозата. Размената на хомологните хромозоми со нивните делови се нарекува декусација или вкрстено синновер(англискиот збор crossingover значи формирање на крст). Се повикуваат поединци со нови комбинации на знаци кои произлегуваат од вкрстување кросовери.
Да се вратиме на слика 14. Кај женските Fi, хетерозиготни за двата пара особини, гените се наоѓаат на еден од хомологните хромозоми КомерсантИ В, во другиот - гени алелични за нив ВОи v. Во профазата на редукциона поделба, кога два хомологни хромозоми се обединуваат во еден бивалентен, секој хромозом се удвојува и се состои од два хроматиди. Ќе има вкупно 4 хроматиди. Помеѓу два хроматиди на хомологни хромозоми, се јавува размена на нивните делови. Како резултат на тоа, генот Комерсант,лоциран во хроматидот на еден хомологен хромозом, може да се поврзе со генот v лоциран во хроматидот на друг хомологен хромозом, а како резултат на еден настан се формира втор хроматид, каде што гените се поврзани ВОИ В. Последователно, хроматидите ќе се одвојат и ќе се формираат вкрстени гамети со хромозоми со нова комбинација на гени. (bv И Б.В.).
Останатите два хроматиди од пар хомологни хромозоми не учествуваат во вкрстувањето и ги задржуваат мајчините во оригиналната комбинација (bV) и татковски (Bv) гени. Формирањето на нови вкрстени гамети обезбеди појава на овошни мушички со нова комбинација на знаци: црни со рудиментирани крилја и сиви долги крилја. Сепак, поголемиот дел од потомството ќе биде слично на оригиналните родители (црни долги крила и сиви кратки крила). Морган заклучува дека бројот на појавувања на нови форми зависи од фреквенцијата на вкрстување, што се одредува со следнава формула:
Ако, на пример, вкупниот број на потомци е 900, а новите форми на вкрстување се 180, тогаш фреквенцијата на вкрстување ќе биде 20 %. Морган го откри тоа фреквенција на вкрстување помеѓу специфичен пар на гени- релативно константна вредност, но различна за различни парови на гени.Врз основа на ова беше заклучено дека Фреквенцијата на вкрстување може да се користи за да се процени растојанијата помеѓу гените. Мерната единица на крстот се зема еднаква на 1%.Понекогаш таа се нарекува Морганида. Количината на вкрстување зависи од растојанието помеѓу гените што се проучуваат.Колку гените се пооддалечени еден од друг, толку почесто се случува вкрстување; колку поблиску се наоѓаат, толку е помала веројатноста за премин. Утврдено е дека бројот на вкрстени индивидуи на вкупниот број на потомци никогаш не надминува 50%, бидејќи на многу големи растојанија помеѓу гените, двојното вкрстување се случува почесто и некои вкрстени поединци остануваат непознати. Тие може да се земат предвид кога се проучуваат не два пара поврзани особини, туку три или четири. Во овој случај, земајќи ги предвид двојните и тројните вкрстувања, можно е попрецизно да се процени растојанијата и зачестеноста на вкрстувањето помеѓу гените.
Блок на информации.
Поврзаност на гените.
На почетокот на 20 век (1902-1907), американскиот научник В. Сетон и германскиот ембриолог Т. Бовери откриле паралелизам во наследувањето на карактеристиките и однесувањето на хромозомите на клеточното јадро за време на процесот на гаметогенеза и оплодување . Ова ја потврди локализацијата на наследни информации во хромозомите. Утврдено е дека бројот на гени значително го надминува бројот на хромозоми. Значи, едно лице има 46 хромозоми, а има од 70.000 до 100.000 гени, како резултат на тоа, голем број гени се локализирани на секој хромозом. Гените на истиот хромозом се наследуваат заедно (поврзани).
Експериментална студија за овој феномен беше спроведена од американскиот генетичар Т. Морган и неговите соработници: А. Стуртевант, А. Бриџис и Г. Молер во 1910-1916 година. Овие студии ја потврдија хромозомската локализација на гените и ја формираа основата на хромозомската теорија на наследноста.
Основни одредби на хромозомската теорија на наследноста.
1. Секој ген зазема одредено место на хромозомот - локус.
2. Гените на хромозомот се распоредени линеарно во одредена низа.
3. Различни хромозоми содржат различен број на гени. Збирот на гени на секој нехомологен хромозом е единствен.
4. Гените на еден хромозом формираат „група за поврзување и се наследуваат заедно, т.е. поврзани.
5. Бројот на групи за поврзување е еднаков на бројот на хромозоми во хаплоидниот сет (има четири кај Drosophila, 10 кај пченката, 20 кај глувците, 23 кај луѓето).
6. Размена на алелни гени, т.е. вкрстување, може да се случи помеѓу хомологните хромозоми.
8. Посебна единица, морганидот (М), се зема како единица за растојание помеѓу гените во група за поврзување. 1М=1% преминување.
Има целосна и нецелосна поврзаност на гените.
Целосно зафат.
Во експериментите на Drosophila беше откриено дека развојот
особини кои се наследени поврзани и контролирани од гените на еден
хромозоми.
Гени за бојата на телото (б - сива и Б - црна) и должината на крилата
(v - нормални и V - кратки, вестигиални крила) се локализирани во еден пар хомологни хромозоми.
Вкрстувањето на сивите муви со нормални крила и сивите муви со заостанати крилја создава сиви хибриди со нормални крила во првата генерација
При спроведувањето на анализираното вкрстување, мажјаците беа избрани од стр, бидејќи е познато дека машките Drosophila имаат ахијазматична сперматогенеза (т.е. вкрстување не се случува и комплетноста на генската поврзаност не е нарушена на кој било начин). вкрстување, индивидуи од два фенотипа слични на оригиналните родителски форми и во еднакви количини: 1:1 фенотипско разделување.
Разгледани заедно, резултатите од двете вкрстувања нè убедуваат дека развојот на анализираните особини е контролиран од различни гени, а поврзаното наследство се објаснува со локализацијата на гените на истиот хромозом. Комплетноста на адхезијата во овој случај не е нарушена од ништо.
Оваа генска врска е завршена.
За да се проучи нецелосната поврзаност од P, беа избрани женките (генотип B||b) кај женките C за време на гаметогенезата; Затоа, дихетерозиготна единка формира дополнителни, т.е. вкрстени сорти на гамети. Веројатноста за нивно формирање се одредува со веројатноста за преминување, т.е. зависи од растојанието помеѓу гените во групата за поврзување.
Не рекомбинантни индивидуи; y Рекомбинантни индивидуи; на
забележано е дека имаат иста формација на нивните генотипови
комбинациите, како и оригиналните, вклучуваа кросовер
Преовладувањето на сивите долгокрилести и црни краткокрилни муви кај потомството укажува дека гените B и V;
b и v се навистина поврзани. Од друга страна, појавата на рекомбинантни индивидуи укажува дека во одреден број случаи има прекин на врската помеѓу гените B и V и гените B и V. Ова е резултат на вкрстување.
Пример за целосна поврзаност на гените кај луѓето е наследството на Rh факторот. Тоа е предизвикано од три пара C, D, K, тесно поврзани еден со друг, затоа наследувањето на припадноста на резус се јавува според видот на монохибридното вкрстување. Друг пример за тесна поврзаност на гените кај луѓето е наследувањето на катаракта и полидактилија. Гените за хемофилија и далтонизам се локализирани на Х-хромозомот на растојание од 9,8 морганиди (М), т.е.
се подложени на вкрстување и затоа се наследуваат како нецелосно поврзани. Автозомните гени за Rh факторот и обликот на еритроцитите, лоцирани на растојание од 3 М едни од други, се исто така пример за нецелосно поврзување.
Во 1909 година, Ф. Јансенс, додека ја проучувал мејозата кај водоземците, открил хијазмати (вкрстувања) на хромозоми, кои се цитолошки доказ за вкрстување. Од тоа време, направени се многу обиди да се објасни механизмот на овој феномен. Постојат неколку теории за вкрстување. Најчести се две хипотези. Планирање на наставата 10-то одделение
Тема: „Целосно и нецелосно поврзување на гените. Генетски карти на хромозоми“.
Цел на часот:
да ги запознае учениците со процесот на целосно и нецелосно поврзување на гените; и формираат идеја за генетските карти на хромозомите.
Образовни задачи:
Откријте ја суштината на феноменот на поврзаното наследување на гените.
Да се развие знаење за главните одредби на законот на Т. Морган.
Воведување на принципот на составување генетски карти : Развијте го логичното размислување на учениците.
Опрема, визуелни помагала табели за општа биологија кои го илустрираат поврзаното наследство на гените и особините, презентација за часот, задачи за консолидирање на нов материјал.
Тип на лекција: Лекција за учење нов материјал.
Методи:
објаснувачко - илустративно. Напредокот на лекцијата:
Јас
Организациски момент Проверка на персоналот на учениците и визуелната подготвеност на часот за часот.
II
Тестирање на знаењето на учениците
Фронтална анкета:
1. Наведете три закони на Г. Мендел?
4. Дали цртите секогаш можат јасно да се поделат на доминантни и рецесивни?
5. Какво име доби оваа појава?
6. Дали е секогаш можно од фенотипот да се определи кои гени содржи дадена индивидуа? Наведи пример.
7. Дали е можно да се утврди генотипот на индивидуи кои не се разликуваат по фенотип? Кој метод се користи за ова?
8. Кои карактеристики го карактеризираат дихибридното вкрстување?
Браво! Ја завршивте оваа фаза на работа
III Учење нов материјал:
Поврзано наследство на гени
Г. Мендел го следеше наследството на седум пара особини во грашокот. Многу истражувачи, повторувајќи ги експериментите на Мендел, ги потврдија законите што тој ги откри. Беше признаено дека овие закони се од универзална природа. Меѓутоа, во 1906 година, англиските генетичари В. Потомците секогаш ги повторувале карактеристиките на нивните родителски форми. Постепено, се акумулираа докази за исклучоци од третиот закон на Мендел. Стана јасно дека не сите гени се карактеризираат со независна дистрибуција во потомството и слободна комбинација.
Секој организам има различни морфолошки, физиолошки, биохемиски и други карактеристики и својства, а секоја карактеристика или својство е контролирана од еден или повеќе гени локализирани во хромозомите.
Меѓутоа, ако бројот на гени во еден организам е огромен и може да изнесува десетици илјади, тогаш бројот на хромозоми е релативно мал и, по правило, се мери во неколку десетици. Затоа, стотици и илјадници алелни гени кои формираат поврзувачки групи се локализирани во секој пар на хромозоми.
Воспоставена е целосна кореспонденција помеѓу бројот на групи за поврзување и бројот на парови на хромозомите. На пример, пченката има збир на хромозоми 2n = 20 и 10 групи за поврзување, а Drosophila има 2n = 8 и 4 поврзувачки групи, односно бројот на групи за поврзување е еднаков на хаплоидниот сет на хромозоми.
Законот Томас Морган
Гените локализирани на истиот хромозом се пренесуваат заедно, а нивниот начин на наследување се разликува од наследувањето на гените локализирани во различни парови на хомологни хромозоми.
Така, на пример, со независна дистрибуција на хромозоми, дихибридАаБб формира четири типа гамети (АБ , aB , Аб , ab ), и под услов на целосна поврзаност, истиот дихибрид ќе даде само два вида гамети (АБ Иab ), бидејќи овие гени се наоѓаат на истиот хромозом.
Развојот на проблемот со поврзаното наследување на гените припаѓа на школата на Т. Морган (1866–1945). Ако Мендел ги спроведе своите експерименти на грашок, тогаш за Морган главниот предмет беше овошната мушичка Дрософила. Мувата раѓа бројни подмладоци на секои две недели на температура од 25 °C. Машкото и женското јасно се разликуваат - мажјакот има помал и потемни стомак. Покрај тоа, тие се разликуваат по бројни карактеристики и можат да се репродуцираат во епрувети на евтин хранлив медиум.
Проучување на моделите на наследувањегени локализирани на истиот хромозом , Морган заклучи дека тиесе наследени поврзани . Ова е закон на Т. Морган.
Премин преку - процес на размена на парцелиза време на во профаза I . Покрај мејотичното, исто така е опишано . Хромозомот е поделен на овие делови во одредени точки, исти за еден вид, што може да биде дефиниција на видот на генетско ниво, локацијата на овие точки се определува со еден ген.
Со оглед на тоа што преминувањето воведува пречки во сликата , успешно се користеше за мапирање на „групи за поврзување“ (хромозоми). Изводливоста на мапирањето беше заснована на претпоставката дека почестото вкрстување се случува помеѓу две , колку подалеку овие гени се лоцирани еден од друг во групата за поврзување и толку почесто ќе се забележат отстапувања од поврзаното наследување.
Целосна и делумна спојка
За да се одреди типот на наследување на два пара гени (поврзани или независни), неопходно е да се спроведе аналитичко вкрстување и, врз основа на неговите резултати, да се донесе заклучок за природата на наследувањето на гените. Дозволете ни да разгледаме три можни варијанти на резултатите од анализата.
1) Независно наследство .
Ако, како резултат на аналитичко вкрстување, меѓу хибридите се формираат четири класи на фенотипови, тоа значи дека гените се наследуваат независно.
2) Целосна генска врска .

Со целосна генска поврзаностА
ИВО
според резултатите од анализираниот премин, тие констатирале -
Постојат две фенотипски класи на хибриди кои целосно ги копираат нивните родители.
3) Нецелосна генска врска .

Во случај на нецелосно поврзување на генитеА ИВО Кога се анализираат вкрстувањата, се појавуваат четири фенотипови, од кои два имаат нова комбинација на гени:Аб ‖ ab ; aB ‖ ab . Појавата на такви форми укажува на тоа дека дихибридот со гаметиАБ │ иab │ формира вкрстени гаметиАб │ иaB │. Појавата на такви гамети е можна само како резултат на размена на делови од хомологни хромозоми, односно во процес на вкрстување. Бројот на вкрстени гамети е значително помал од не-кросовер гамети.
Фреквенцијата на вкрстување е пропорционална на растојанието помеѓу гените. Колку поблиску се наоѓаат гените на хромозомот, толку е поблиска врската меѓу нив и поретко се раздвојуваат за време на вкрстувањето. И обратно, колку гените се подалеку еден од друг, толку е послаба врската меѓу нив и почесто тие се вкрстуваат. Следствено, растојанието помеѓу гените на хромозомите може да се процени според фреквенцијата на вкрстување.
Генетски карти
Генетското мапирање обично значи одредување на положбата на генот во однос на другите гени.
Да ја разгледаме постапката за составување генетски карти.
1. Воспоставување на групата за поврзување (односно определување на хромозомот во кој е локализиран генот). За да го направите ова, неопходно е да имате барем еден ген за маркери во секоја поврзана група.
2. Наоѓање на локацијата на генот што се проучува во хромозомот. За да го направите ова, мутантната форма се вкрстува со нормална и се зема предвид резултатот од вкрстувањето.
3. Определување на растојанието помеѓу поврзаните гени, што овозможува да се изготват генетски карти на хромозомите, кои укажуваат на редоследот на гените во хромозомите и нивните релативни растојанија едни од други. Колку е поголема фреквенцијата на вкрстување, толку е поголемо растојанието помеѓу гените. Доколку се утврди дека помеѓу поврзаните гениА ИВО фреквенцијата на вкрстување е 10%, а помеѓу генитеВО ИСО – 20%, тогаш е очигледно дека растојаниетоСонцето 2 пати повеќе одАБ . Растојанието помеѓу гените се изразува во единици што одговараат на 1% вкрстување. Овие единици се нарекуваат морганиди.
Така, генетските карти се составуваат врз основа на податоците за фреквенцијата на вкрстување.
IV Консолидација на знаењето
Решавање на генетски проблем
Жена Drosophila хетерозиготна за рецесивни гени за темна боја на телото и минијатурни крилја била вкрстена со мажјак кој имал темно тело и минијатурни крилја. Од овој премин е добиено следново:
– 244 муви со темни тела и минијатурни крилја;
– 20 муви со сива боја на каросеријата и минијатурни крилја;
– 15 муви со темна боја на телото и нормални крила;
– 216 муви со сива боја на телото и нормални крилја.
Врз основа на дадените податоци, утврди дали овие два пара гени се поврзани или не. Како се поврзани гените?
Со оглед на :
А
– сиво тело
А
– темно тело
ВО
– нормални крилја
б
– минијатурни крилја
Природата на генското наследувањеА ИВО - ?
Решение
Резултатите од разделувањето меѓу хибридите (две фенотипски класи се доминантни и ги повторуваат фенотипски и генотипски родителските форми, а другите две класи на фенотипови се претставени со мал број индивидуи) укажуваат на нецелосна поврзаност на гените§41, стр.161-164. Одговори на прашањата стр.165.
Задача бр. 1. Дихетерозиготно растение грашок со мазни семиња и ластари било вкрстено со растение со збрчкани семиња без ластари. Познато е дека и двата доминантни гени (мазни семиња и присуството на ластари) се локализирани на истиот хромозом; Направете дијаграм за решавање на проблемот. Определете ги генотиповите на родителите, фенотиповите и генотиповите на потомството, односот на поединци со различни генотипови и фенотипови. Кој закон се манифестира во овој случај?
А - мазни семиња, а - збрчкани семиња
Б - присуство на антени, б - без антени
AB/ab
ab/ab
AB/ab
ab/ab
мазна
семиња,
мустаќи
збрчкана.
семиња,
без мустаќи
50%
50%
Ако не се случи вкрстување, тогаш дихетерозиготниот родител произведува само два вида гамети (целосна поврзаност).
Гените лоцирани на истиот хромозом се група спојката. Генска поврзаност- Ова е заедничко наследство на гени лоцирани на истиот хромозом. Бројот на групи за поврзување одговара на хаплоидниот број на хромозоми. Врската на гените лоцирани на истиот хромозом може да биде целосна или нецелосна. Целосно зафат: Морган вкрсти црни долгокрилни женки со сиви мажјаци со рудиментирани крилја. Во Drosophila, сивата боја на телото доминира над црната, а долгите крилја доминираат над рудиментираните крилја. Сиво тело - А, црно тело a; долгокрилести - Б, рудиментирани крилја - в. За време на спермиогенезата за време на мејозата, хомологните хромозоми се одвојуваат во различни герминативни клетки. 1) AA//AB*av//av=4AB//av; 2) AB//av*AB//av=AB//AB, AB//av, ab//AV, ab//av. Ако гените се најдат во автозомите, тогаш со целосна поврзаност во F1 ќе има униформност во фенотипот, а во F2 - 3:1, без разлика во колку карактеристики се разликуваат родителите, бидејќи се проучува еден пар на хромозоми.
Феноменот на нецелосно поврзување во наследувањето на особините
Како резултат на вкрстувањето, потомците имале комбинација на карактеристики, како првобитните родителски форми, но поединците се појавувале и со нова комбинација на карактеристики - спојката нецелосна. Б – сива, в – црна, V – нормална, v – рудиментирана. Bv||Bv*bV||bV=Bv||bV; женките од првата генерација беа вкрстени со мажјаци-анализатори: BV//bV*bv//bv=Bv//bv,bV//bv – не кросовер. Bv//bV*bv//bv=2bv//bv, 2BV//bv – кросовер. Размената на хомологните хромозоми со нивните делови се нарекува вкрстување или вкрстување.Се повикуваат поединци со нови комбинации на знаци кои произлегуваат од вкрстување кросовери.Бројот на појавувања на нови форми зависи од фреквенцијата на вкрстување, која се одредува со следната формула: Фреквенција на вкрстување = (Број на форми на вкрстување) 100/ Вкупен број на потомци. Мерната единица на крстот се зема еднаква на 1%. Таа се вика морганида. Количината на вкрстување зависи од растојанието помеѓу гените што се проучуваат. Колку гените се пооддалечени еден од друг, толку почесто се случува вкрстување; колку поблиску се наоѓаат, толку е помала веројатноста за премин.
Мапи на хромозомите. Пример за нивната конструкција
Карта на хромозомите- план на локацијата на гените на хромозомот. Гените се наоѓаат на хромозомите во линеарна низа на одредени растојанија едни од други. Феноменот на инхибиција на вкрстување во една област со вкрстување во друга се нарекува мешање.Колку е помало растојанието што ги раздвојува трите гени, толку е поголема интерференцијата. Земајќи го предвид линеарното распоредување на гените во хромозомот, земајќи го вкрстувањето преку фреквенцијата како единица за растојание, Морган ја составил првата карта на локацијата на гените во еден од хромозомите на Drosophila: сh___13.6___ y___28.2___b. Кога се конструираат мапи, тие не го означуваат растојанието помеѓу гените, туку растојанието до секој ген од нултата точка на почетокот на хромозомот. Доминантниот алел е означен со голема буква, а рецесивниот алел со мала буква. По конструирањето на генетските карти, се појави прашањето дали локацијата на гените на хромозомот, конструирана врз основа на фреквенцијата на вкрстување, одговара на вистинската локација. Секој хромозом долж неговата должина има специфични обрасци на дискови, што овозможува да се разликуваат неговите различни делови едни од други. Материјалот за тестирање беа хромозоми во кои, како резултат на мутација, се појавија различни хромозомски преуредувања: недостасуваа поединечни дискови, или тие беа превртени или двојно зголемени. Физичките растојанија помеѓу гените на генетската карта не соодветствуваат сосема на утврдените цитолошки растојанија. Сепак, ова не ја намалува вредноста на генетските карти на хромозоми за предвидување на веројатноста за појава на поединци со нови комбинации на особини. Врз основа на анализа на резултатите од бројните експерименти со Дрософила, Т. Морган формулираше теорија на хромозомитенаследноста, чија суштина е следново: 1) гените се наоѓаат во хромозомите, лоцирани линеарно во нив на одредено растојание едни од други; 2) гените лоцирани на истиот хромозом припаѓаат на иста поврзана група. Бројот на групи за поврзување одговара на хаплоидниот број на хромозоми; H) особини чии гени се наоѓаат на истиот хромозом се наследени поврзани; 4) кај потомците на хетерозиготни родители, може да се појават нови комбинации на гени лоцирани во истиот пар на хромозоми како резултат на вкрстување за време на процесот на мејоза. Фреквенцијата на вкрстување зависи од растојанието помеѓу гените; 5) врз основа на линеарното распоредување на гените во хромозомот и фреквенцијата на вкрстување како показател за растојанието помеѓу гените, може да се конструираат мапи на хромозомите.
Бисексуалност, интерсексуалност, гиандроморфизам, химеризам според полови хроми. Улогата на хормоните и условите на околината во развојот на половите карактеристики
Секој зигот има х-хроми и автозоми, т.е. има и женски и машки гени, т.е. генетски секој организам бисексуалец(бисексуалец). Интерсекс– хермафродити – индивидуи со развиени и женски и машки карактеристики. 2 типа: точно - имаат женски и машки гонади поради нерамнотежа на гените; условно - имаат жлезди од еден пол, а надворешните полови карактеристики на другиот пол поради нерамнотежа на хормоните. Понекогаш се наоѓа кај инсекти и животни гиандроморфи– едниот дел од телото има женски карактеристики, а другиот машки. Причини: Женскиот зигот е поделен на 2 бластомери. Еден од нив изгубил еден х-лејм. Од овој бластомер ќе се развие машката половина од телото. Химеризамполовина од хромозомите xx/xy се наоѓаат кај мултипари животни, кај бикови - кога истиот организам содржи xx хромозоми, а репродукцијата на xy хромозомите е нарушена. Со нормално хранење, мажјаците растат, а ако во храната се додадат женски полови хормони, растат женките (рибното пржење). Ако ларвата на морскиот црв се закачи на дното на морето, тоа е женка, ако е закачена за пробосцис на женка, тоа е мажјак.
Видови на определување пол кај животните. Примарен и секундарен однос на половите. Проблемот на родовата регулација
Определувањеобезбедува формирање на еднаков број мажи и жени, што е неопходно за нормална саморепродукција на видот. Видови: 1) епигамно - полот на поединецот се одредува за време на онтогенезата и зависи од надворешната средина. 2) прогамно - сексот се одредува при гаметогенезата кај родителите на поединецот. 3) сингамно - полот се одредува во моментот на фузија на гамети. Сооднос на примарен и секундарен пол:половиот сооднос, мачката се одредува во моментот на фузија на гамети, наречен основно, секогаш 1:1. Секоја промена во односот на половите, било пред или по раѓањето, се нарекува секундарно.Обично по раѓањето се префрла во корист на женскиот пол, така што кај многу животински видови и луѓе се раѓаат повеќе мажи отколку жени: зајаци - 57%, луѓе - 51%, птици - 59%. Проблем со родовата регулација:е од големо економско значење. На пример: во млекарството и одгледувањето на живина што носи јајца, пожелни се женките, а каде главниот производ е месото, подобри се мажјаците. Проблемот е да се оддели спермата на x- и y-фракции. Методи: 1) електрофореза - x - сперматозоидите имаат негативен полнеж - се движат кон катодата, а y - сперматозоидите се движат кон анодата. 80% гаранција. 2) Метод на седиментација - x - спермата е погуста и се смирува, а y - останува на врвот. 3) Користење на сет на киселини за промена на pH на женскиот репродуктивен тракт за да се создадат услови само за x - или само за y-. 4) Партеногенеза: геногенеза - добивање женки - ооцитот се озрачува со рендгенски зраци. прв ред, со што се одложува дивергенцијата на хром-м, се формира јајце со диплоиден сет на хром-м, а женката се развива во мачка без оплодување. Андрогенеза - добивање на мажјаци - јадрото на јајце клетката се убива со рендгенски зраци, потоа две сперматозоиди продираат во него, јадрата се спојуваат, давајќи диплоиден сет, ќе има мажјак. 5) Начин на делење на сперматозоидите на фракции врз основа на количината на ДНК во спермата. 6) Колку се помлади родителите, толку е поголема веројатноста да се родат машки. 7) Колку повеќе сперматозоиди во женскиот генитален тракт, толку е поголема веројатноста за раѓање на маж. 8) Колку подолго се чува спермата - женската. 9) Хранење со живина: ако додадете Ca во храната на петелот, тогаш тоа е женско, а ако К, тоа е машко. 10) Во секоја популација важи законот за рамнотежа, т.е. Соодносот на половите има тенденција да биде 1:1.
Поврзано наследување на особини.Анализата на истовременото наследување на неколку особини кај Дрософила, спроведена од Т. Морган, покажа дека резултатите од аналитичкото вкрстување на хибридите на Fi понекогаш се разликуваат од оние што се очекуваат во случај на нивно независно наследување. Кај потомците на ваквите крстови, наместо слободно да се комбинираат особините од различни парови, забележана е тенденција да се наследуваат претежно родителски комбинации на особини. Ова наследство на особини беше наречено поврзани.Поврзаното наследство се објаснува со локацијата на соодветните гени на истиот хромозом. Како дел од второто, тие се пренесуваат од генерација на генерација на клетки и организми, зачувувајќи ја комбинацијата на родителски алели.
Зависноста на поврзаното наследување на особините од локализацијата на гените на еден хромозом дава основа да се сметаат хромозомите како посебни групи на спојката.
При анализа на вкрстени мажјаци од F 1, се појавија само два типа на потомство, слично на родителите во комбинацијата на варијанти на анализираните знаци (сива боја на телото и нормални крилја или црна боја на телото и кратки крилја) во сооднос 1:1. Ова покажува дека мажјаците F 1 произведуваат само два вида гамети со еднаква веројатност, кои ги вклучуваат оригиналните родителски комбинации на алели на гените кои ги контролираат именуваните особини (BV или bv).
При анализа на вкрстувањето на женките F 1, се појавија четири типа на потомци со сите можни комбинации на карактеристики. Во исто време, потомство со родителски комбинации на особини биле пронајдени кај 83%. 17% од потомците имале нови комбинации на знаци (сива боја на телото и кратки крилја или црна боја на телото и нормални крилја). Може да се види дека кај овие вкрстувања постои и тенденција за поврзано наследување или на доминантни или на рецесивни особини (83%). Делумното нарушување на поврзаноста (17% од потомците) беше објаснето со процесот на вкрстување - размена на соодветните делови од хомологни хромозоми во профазата I на мејозата (види Сл. 3.72).
Од резултатите од вкрстувањето произлегува дека женките Drosophila формираат четири типа гамети, од кои повеќето (83%) не се вкрстени ((BV) и (bv)), 17% од гаметите што ги формираат се појавуваат како резултат на вкрстување над и носат нови комбинации на алели на анализираните гени ((Bv ) и (bV)). Разликите забележани кога мажјаците и женките од F 1 се вкрстуваат со рецесивни хомозиготни партнери се објаснети со фактот дека од причини кои се слабо разбрани, вкрстувањето не се случува кај Drosophila мажите. Како резултат на тоа, мажјаците кои се дихетерозиготни за гени лоцирани на истиот хромозом формираат два вида гамети. Кај женките се јавува вкрстување и доведува до формирање на не-кросовер и вкрстувачки гамети, по два вида од секоја. Затоа, четири фенотипови се појавуваат кај потомците на анализирачкиот крст, од кои два имаат нови комбинации на особини во споредба со родителите.
Ориз. 6.14. Поврзано наследување на особини
(боја на телото и должина на крилата во Drosophila):
објаснувачко - илустративно.- преминување на чисти линии, II, III -анализирање на вкрстувања помеѓу мажјаци и жени од F 1.
Студијата за наследување на други комбинации на особини покажа дека процентот на вкрстени потомци за секој пар на особини е секогаш ист, но тој варира за различни парови. Ова набљудување доведе до заклучок дека гените на хромозомите се распоредени во линеарен редослед. Погоре беше забележано дека хромозомот е поврзана група на одредени гени. Хомологните хромозоми се идентични групи на поврзување кои се разликуваат една од друга самоалели на поединечни гени. За време на конјугацијата, хомолозите се приближуваат меѓу себе со нивните алелни гени, а при вкрстувањето тие разменуваат соодветни региони. Како резултат на тоа, вкрстените хромозоми се појавуваат со нов сет на алели. Фреквенцијата со која се случува размена на место помеѓу два дадени гени зависи од растојанието меѓу нив (правило на Т. Морган).Процентот на вкрстени гамети индиректно го одразува растојанието помеѓу гените. Ова растојание обично се изразува во сантшморганиди.Еден центиморганид се зема како растојание помеѓу гените на кое се формираат 1% од вкрстено потомство (кросовер гамети).
Како што се зголемува растојанието помеѓу гените, се зголемува веројатноста за вкрстување во областа меѓу нив во клетките прекурсори на гамети. Бидејќи два од четирите хроматиди присутни во бивалентот се вклучени во чинот на вкрстување, дури и ако се случи размена помеѓу гените на даден пар во сите претходници на гамети, процентот на вкрстени герминативни клетки не може да надмине 50. Сепак, таквата ситуација е можна само теоретски. Во пракса, како што се зголемува растојанието помеѓу гените, се зголемува можноста за неколку вкрстувања кои се случуваат истовремено во дадена област (види Сл. 5.9). Бидејќи секој втор вкрстување води до обновување на претходната комбинација на алели во хромозомот, со зголемување на растојанието, бројот на вкрстени гамети може да не се зголемува, туку да се намалува. Од ова произлегува дека процентот на вкрстени гамети е показател за вистинското растојание помеѓу гените само кога тие се наоѓаат доволно блиску, кога е исклучена можноста за втор премин.
Нарушувањето на поврзаното наследување на родителските алели како резултат на вкрстување ни овозможува да зборуваме за нецелосна спојказа разлика од целосна спојка,забележано, на пример, кај машки Drosophila.
Употребата на аналитичко вкрстување во експериментите на Т. Морган покажа дека со негова помош е можно да се одреди не само составот на парови на не-алелни гени, туку и природата на нивното заедничко наследување. Во случај на поврзано наследување на особини, врз основа на резултатите од анализата на вкрстувањето, може да се одреди и растојанието помеѓу гените во хромозомот.

Ориз. 6.15. Дијаграм што го објаснува нискиот процент на вкрстени гамети
(во однос на овие два гена)
Плус ги означува клетките прекурсори на гамети во кои се случило вкрстување во областа помеѓу овие два гена; вкрстените гамети се поцрнети
Генетски карти на хромозоми- ова е дијаграм на релативната положба и релативните растојанија помеѓу гените на одредени хромозоми лоцирани во иста поврзана група.
За прв пат, Т. Морган и неговите колеги укажаа на можноста за конструирање на генетски карти на хромозоми. Тие експериментално покажаа дека врз основа на феноменот на генска поврзаност и вкрстување, можно е да се конструираат генетски карти на хромозоми. Способноста за мапирање се заснова на конзистентноста на процентот на вкрстување помеѓу одредени гени. Составени се генетски карти на хромозоми за многу видови организми: инсекти (дрозофила, комарец, бубашваби и др.), габи (квасец, аспергилус), бактерии и вируси.
Човечките генетски карти се користат во медицината за дијагностицирање на голем број тешки наследни болести кај луѓето. Студиите на еволутивниот процес ги споредуваат генетските карти на различни видови живи организми.