Wymiana substancji wchodzących do komórki lub uwalnianych przez nią na zewnątrz, a także wymiana różnych sygnałów z mikro- i makrośrodowiskiem odbywa się poprzez zewnętrzną błonę komórki. Jak wiadomo, błona komórkowa jest dwuwarstwą lipidową, w której osadzone są różne cząsteczki białka, które działają jak wyspecjalizowane receptory, kanały jonowe, urządzenia aktywnie transportujące lub usuwające różne substancje chemiczne, kontakty międzykomórkowe itp. W zdrowych komórkach eukariotycznych fosfolipidy są rozmieszczone w błona jest asymetryczna: powierzchnia zewnętrzna składa się ze sfingomieliny i fosfatydylocholiny, powierzchnia wewnętrzna - z fosfatydyloseryny i fosfatydyloetanoloaminy. Utrzymanie takiej asymetrii wymaga nakładów energetycznych. Dlatego w przypadku uszkodzenia komórki, infekcji lub głodu energetycznego zewnętrzna powierzchnia błony zostaje wzbogacona w nietypowe dla niej fosfolipidy, co staje się sygnałem dla innych komórek i enzymów o uszkodzeniu komórki z odpowiednią reakcją na to. Najważniejszą rolę odgrywa rozpuszczalna forma fosfolipazy A2, która rozkłada kwas arachidonowy i tworzy lizoformy z wymienionych fosfolipidów. Kwas arachidonowy jest ogniwem ograniczającym tworzenie mediatorów stanu zapalnego, takich jak eikozanoidy, a cząsteczki ochronne - pentraksyny (białko C-reaktywne (CRP), prekursory białek amyloidowych) - przyłączają się do lizoform w błonie, po czym następuje aktywacja dopełniacza systemu wzdłuż szlaku klasycznego i niszczenia komórek.
Struktura membrany pomaga zachować cechy środowiska wewnętrznego komórki, jej różnice w stosunku do środowiska zewnętrznego. Zapewnia to selektywna przepuszczalność błony komórkowej i istnienie w niej aktywnych mechanizmów transportu. Ich zakłócenie na skutek bezpośredniego uszkodzenia np. przez tetrodotoksynę, ouabainę, tetraetyloamoniak lub w przypadku niedostatecznego dopływu energii do odpowiednich „pomp” prowadzi do zaburzenia składu elektrolitowego komórki, zmian w jej metabolizmie, zakłócenia określonych funkcji - skurczu, przewodzenia impulsów wzbudzenia itp. Zaburzenie komórkowych kanałów jonowych (wapniowych, sodowych, potasowych i chlorkowych) u człowieka może być również uwarunkowane genetycznie poprzez mutację genów odpowiedzialnych za strukturę tych kanałów. Tak zwane kanałopatie powodują dziedziczne choroby układu nerwowego, mięśniowego i trawiennego. Nadmierne przedostawanie się wody do komórki może prowadzić do jej pęknięcia – cytolizy – w wyniku perforacji błony podczas aktywacji dopełniacza lub ataku cytotoksycznych limfocytów i komórek NK.
Błona komórkowa ma wbudowanych wiele receptorów – struktur, które w połączeniu z odpowiednimi specyficznymi cząsteczkami sygnalizacyjnymi (ligandami) przekazują sygnał wewnątrz komórki. Dzieje się to poprzez różne kaskady regulacyjne składające się z enzymatycznie aktywnych cząsteczek, które są kolejno aktywowane i ostatecznie przyczyniają się do realizacji różnych programów komórkowych, takich jak wzrost i proliferacja, różnicowanie, ruchliwość, starzenie się i śmierć komórki. Kaskady regulacyjne są dość liczne, jednak ich liczba nie została jeszcze w pełni ustalona. System receptorów i związane z nimi kaskady regulacyjne istnieją także wewnątrz komórki; tworzą specyficzną sieć regulacyjną z punktami koncentracji, dystrybucji i wyboru dalszej ścieżki sygnału w zależności od stanu funkcjonalnego komórki, etapu jej rozwoju i jednoczesnego działania sygnałów z innych receptorów. Efektem tego może być zahamowanie lub wzmocnienie sygnału, kierując go inną ścieżką regulacyjną. Zarówno aparat receptorowy, jak i szlaki przekazywania sygnału poprzez kaskady regulacyjne, np. do jądra, mogą zostać zakłócone w wyniku defektu genetycznego, który występuje jako defekt wrodzony na poziomie organizmu lub na skutek mutacji somatycznej w określonym typie komórki. Mechanizmy te mogą zostać uszkodzone przez czynniki zakaźne, toksyny, a także ulec zmianie w procesie starzenia. Końcowym etapem tego może być zaburzenie funkcji komórki, procesów jej proliferacji i różnicowania.
Na powierzchni komórek znajdują się również cząsteczki, które odgrywają ważną rolę w procesach interakcji międzykomórkowych. Mogą do nich należeć białka adhezyjne komórek, antygeny zgodności tkankowej, antygeny specyficzne tkankowo, różnicujące itp. Zmiany w składzie tych cząsteczek powodują zaburzenie oddziaływań międzykomórkowych i mogą powodować aktywację odpowiednich mechanizmów eliminacji takich komórek, gdyż stanowią one pewne zagrożenie dla integralności organizmu jako rezerwuar infekcji, zwłaszcza wirusowych, lub jako potencjalne inicjatory wzrostu nowotworu.
Naruszenie zaopatrzenia energetycznego komórki
Źródłem energii w komórce jest pożywienie, po rozkładzie którego energia uwalniana jest na substancje końcowe. Głównym miejscem wytwarzania energii są mitochondria, w których substancje utleniane są za pomocą enzymów łańcucha oddechowego. Głównym dostawcą energii jest utlenianie, gdyż w wyniku glikolizy z tej samej ilości substratów utleniania (glukozy) uwalnia się nie więcej niż 5% energii w porównaniu z utlenianiem. Około 60% energii uwalnianej podczas utleniania jest akumulowane poprzez fosforylację oksydacyjną w wysokoenergetycznych fosforanach (ATP, fosforan kreatyny), reszta jest rozpraszana w postaci ciepła. Następnie wysokoenergetyczne fosforany są wykorzystywane przez komórkę do takich procesów, jak praca pompy, synteza, podział, ruch, wydzielanie itp. Istnieją trzy mechanizmy, których uszkodzenie może spowodować zakłócenie w zaopatrzeniu komórki w energię: pierwszy to mechanizm syntezy enzymów metabolizmu energetycznego, drugi to mechanizm fosforylacji oksydacyjnej, trzeci to mechanizm wykorzystania energii.
Zakłócenie transportu elektronów w mitochondrialnym łańcuchu oddechowym lub rozłączenie utleniania i fosforylacji ADP z utratą potencjału protonowego, siła napędowa wytwarzania ATP, prowadzi do osłabienia fosforylacji oksydacyjnej w taki sposób, że większość energii jest rozpraszana w postaci ciepła i zmniejsza się liczba związków wysokoenergetycznych. Rozdzielenie utleniania i fosforylacji pod wpływem adrenaliny jest wykorzystywane przez komórki organizmów homeotermicznych do zwiększenia produkcji ciepła przy jednoczesnym utrzymaniu stałej temperatury ciała podczas chłodzenia lub jej zwiększeniu w czasie gorączki. W tyreotoksykozie obserwuje się istotne zmiany w strukturze mitochondriów i metabolizmie energetycznym. Zmiany te są początkowo odwracalne, jednak po pewnym czasie stają się nieodwracalne: mitochondria ulegają fragmentacji, rozpadają się lub pęcznieją, tracą cristae, zamieniają się w wakuole, a ostatecznie gromadzą substancje takie jak szklista, ferrytyna, wapń, lipofuscyna. U pacjentów chorych na szkorbut mitochondria łączą się, tworząc chondriosfery, prawdopodobnie w wyniku uszkodzenia błony przez związki nadtlenkowe. Do znacznych uszkodzeń mitochondriów dochodzi pod wpływem promieniowania jonizującego podczas transformacji komórki normalnej w komórkę złośliwą.
Mitochondria są potężnym magazynem jonów wapnia, gdzie ich stężenie jest o kilka rzędów wielkości wyższe niż w cytoplazmie. Kiedy mitochondria ulegają uszkodzeniu, wapń przedostaje się do cytoplazmy, powodując aktywację proteinaz, co powoduje uszkodzenie struktur wewnątrzkomórkowych i zaburzenie funkcji odpowiedniej komórki, na przykład przykurcze wapniowe, a nawet „śmierć wapnia” w neuronach. W wyniku zakłócenia zdolności funkcjonalnych mitochondriów gwałtownie wzrasta powstawanie wolnorodnikowych związków nadtlenkowych, które charakteryzują się bardzo dużą reaktywnością i przez to uszkadzają ważne składniki komórki – kwasy nukleinowe, białka i lipidy. Zjawisko to obserwuje się podczas tzw. stresu oksydacyjnego i może mieć negatywne konsekwencje dla bytu komórki. Zatem uszkodzeniu zewnętrznej błony mitochondriów towarzyszy uwolnienie do cytoplazmy substancji zawartych w przestrzeni międzybłonowej, przede wszystkim cytochromu C i niektórych innych substancji biologicznie czynnych, które wyzwalają reakcje łańcuchowe powodujące zaprogramowaną śmierć komórki – apoptozę. Uszkodzając mitochondrialne DNA, reakcje wolnorodnikowe zniekształcają informację genetyczną niezbędną do tworzenia niektórych enzymów łańcucha oddechowego, które są produkowane specyficznie w mitochondriach. Prowadzi to do jeszcze większego zakłócenia procesów oksydacyjnych. Ogólnie rzecz biorąc, własny aparat genetyczny mitochondriów, w porównaniu z aparatem genetycznym jądra, jest słabiej chroniony przed szkodliwymi wpływami, które mogą zmienić zakodowaną w nim informację genetyczną. W rezultacie dysfunkcja mitochondriów występuje przez całe życie, na przykład podczas procesu starzenia, podczas transformacji złośliwej komórki, a także na tle dziedzicznych chorób mitochondrialnych związanych z mutacją mitochondrialnego DNA w jaju. Obecnie opisano ponad 50 mutacji mitochondrialnych powodujących dziedziczne choroby zwyrodnieniowe układu nerwowego i mięśniowego. Są przekazywane dziecku wyłącznie od matki, ponieważ mitochondria plemników nie są częścią zygoty, a zatem nowego organizmu.
Naruszenie przechowywania i przekazywania informacji genetycznej
Jądro komórkowe zawiera większość informacji genetycznej i tym samym zapewnia jej normalne funkcjonowanie. Poprzez selektywną ekspresję genów koordynuje aktywność komórek podczas interfazy, przechowuje informację genetyczną oraz odtwarza i przekazuje materiał genetyczny podczas podziału komórki. Replikacja DNA i transkrypcja RNA zachodzą w jądrze. Różne czynniki chorobotwórcze, takie jak promieniowanie ultrafioletowe i jonizujące, utlenianie wolnych rodników, chemikalia, wirusy, mogą uszkodzić DNA. Szacuje się, że każda komórka zwierzęcia stałocieplnego potrzebuje 1 dnia. traci ponad 10 000 baz. Tutaj powinniśmy dodać naruszenia podczas kopiowania podczas podziału. Jeśli te uszkodzenia będą się utrzymywać, komórka nie będzie w stanie przetrwać. Ochrona polega na istnieniu potężnych systemów naprawczych, takich jak endonukleaza ultrafioletowa, systemy naprawy replikacji i rekombinacji, które zastępują uszkodzenia DNA. Defekty genetyczne w układach naprawczych powodują rozwój chorób spowodowanych zwiększoną wrażliwością na czynniki uszkadzające DNA. Jest to xeroderma pigmentosum, a także niektóre zespoły przyspieszonego starzenia, którym towarzyszy zwiększona tendencja do rozwoju nowotworów złośliwych.
System regulacji procesów replikacji DNA, transkrypcji informacyjnego RNA (mRNA) i translacji informacji genetycznej z kwasów nukleinowych na strukturę białek jest dość złożony i wielopoziomowy. Oprócz kaskad regulacyjnych wyzwalających działanie czynników transkrypcyjnych o łącznej liczbie ponad 3000, które aktywują określone geny, istnieje także wielopoziomowy system regulacyjny, w którym pośredniczą małe cząsteczki RNA (interferujący RNA; RNAi). Genom człowieka, składający się z około 3 miliardów zasad purynowych i pirymidynowych, zawiera jedynie 2% genów strukturalnych odpowiedzialnych za syntezę białek. Pozostałe zapewniają syntezę regulatorowych RNA, które jednocześnie z czynnikami transkrypcyjnymi aktywują lub blokują pracę genów strukturalnych na poziomie DNA w chromosomach lub wpływają na procesy translacji informacyjnego RNA (mRNA) podczas tworzenia cząsteczki polipeptydu w cytoplazma. Naruszenie informacji genetycznej może wystąpić zarówno na poziomie genów strukturalnych, jak i regulacyjnej części DNA, z odpowiednimi objawami w postaci różnych chorób dziedzicznych.
Ostatnio wiele uwagi poświęca się zmianom w materiale genetycznym, które zachodzą w trakcie indywidualnego rozwoju organizmu i wiążą się z hamowaniem lub aktywacją niektórych odcinków DNA i chromosomów na skutek ich metylacji, acetylacji i fosforylacji. Zmiany te utrzymują się przez długi czas, czasami przez całe życie organizmu od embriogenezy do starości i nazywane są dziedzicznością epigenomiczną.
Proliferacji komórek ze zmienioną informacją genetyczną zapobiegają także systemy (czynniki) kontrolujące cykl mitotyczny. Oddziałują one z zależnymi od cyklin kinazami białkowymi i ich podjednostkami katalitycznymi – cyklinami – blokując przejście komórki przez pełny cykl mitotyczny, zatrzymując podział na granicy fazy presyntetycznej i syntetycznej (blok G1/S) do czasu zakończenia naprawy DNA, a jeśli jest to niemożliwe, inicjują komórki zaprogramowanej śmierci. Do czynników tych zalicza się gen p53, którego mutacja powoduje utratę kontroli nad proliferacją transformowanych komórek; obserwuje się go w prawie 50% ludzkich nowotworów. Drugi punkt kontrolny cyklu mitotycznego znajduje się na granicy G2/M. Tutaj prawidłowe rozmieszczenie materiału chromosomowego pomiędzy komórkami potomnymi w mitozie lub mejozie jest kontrolowane za pomocą zestawu mechanizmów kontrolujących wrzeciono komórkowe, centrum i centromery (kinetochory). Nieskuteczność tych mechanizmów prowadzi do zaburzenia rozmieszczenia chromosomów lub ich części, co objawia się brakiem jakiegokolwiek chromosomu w jednej z komórek potomnych (aneuploidia), obecnością dodatkowego chromosomu (poliploidia), oddzieleniem się chromosomu część chromosomu (delecja) i jej przeniesienie na inny chromosom (translokacja). Procesy takie bardzo często obserwuje się podczas proliferacji komórek nowotworowo zdegenerowanych i transformowanych. Jeśli nastąpi to podczas mejozy z komórkami rozrodczymi, prowadzi to albo do śmierci płodu we wczesnym etapie rozwoju embrionalnego, albo do narodzin organizmu z chorobą chromosomalną.
Niekontrolowana proliferacja komórek podczas wzrostu nowotworu następuje w wyniku mutacji w genach kontrolujących proliferację komórek i nazywa się je onkogenami. Spośród ponad 70 obecnie znanych onkogenów większość należy do składników regulacji wzrostu komórek, niektóre są reprezentowane przez czynniki transkrypcyjne regulujące aktywność genów, a także czynniki hamujące podział i wzrost komórek. Kolejnym czynnikiem ograniczającym nadmierną ekspansję (rozprzestrzenianie się) proliferujących komórek jest skracanie końców chromosomów – telomerów, które w wyniku oddziaływania czysto sterycznego nie są w stanie w pełni replikować, dlatego po każdym podziale komórki telomery ulegają skróceniu o pewna część baz. Zatem proliferujące komórki dorosłego organizmu po określonej liczbie podziałów (zwykle od 20 do 100 w zależności od rodzaju organizmu i jego wieku) wyczerpują długość telomerów i dalsza replikacja chromosomów zatrzymuje się. Zjawisko to nie występuje w nabłonku plemnika, enterocytach i komórkach embrionalnych ze względu na obecność enzymu telomerazy, który przywraca długość telomerów po każdym podziale. W większości komórek dorosłych organizmów telomeraza jest blokowana, ale niestety w komórkach nowotworowych ulega aktywacji.
Połączenie jądra z cytoplazmą oraz transport substancji w obu kierunkach odbywa się poprzez pory w błonie jądrowej przy udziale specjalnych systemów transportowych zużywających energię. W ten sposób do jądra transportowane są substancje energetyczne i plastyczne, cząsteczki sygnalizacyjne (czynniki transkrypcyjne). Odwrotny przepływ przenosi do cytoplazmy cząsteczki mRNA i przenosi RNA (tRNA), rybosomy niezbędne do syntezy białek w komórce. Ta sama droga transportu substancji jest również charakterystyczna dla wirusów, w szczególności takich jak HIV. Przenoszą swój materiał genetyczny do jądra komórki gospodarza z jego dalszym włączeniem do genomu gospodarza i przeniesieniem nowo powstałego wirusowego RNA do cytoplazmy w celu dalszej syntezy białek nowych cząstek wirusa.
Naruszenie procesów syntezy
Procesy syntezy białek zachodzą w cysternach retikulum endoplazmatycznego, ściśle powiązanych z porami w błonie jądrowej, przez które rybosomy, tRNA i mRNA przedostają się do retikulum endoplazmatycznego. Prowadzi się tu syntezę łańcuchów polipeptydowych, które następnie uzyskują ostateczną postać w ziarnistej siatce śródplazmatycznej i kompleksie blaszkowym (kompleks Golgiego), gdzie ulegają modyfikacji potranslacyjnej i łączą się z cząsteczkami węglowodanów i lipidów. Nowo powstałe cząsteczki białka nie pozostają w miejscu syntezy, lecz w wyniku złożonego regulowanego procesu tzw proteinkineza, są aktywnie przenoszone do izolowanej części komórki, gdzie będą pełnić zamierzoną funkcję. W tym przypadku bardzo ważnym krokiem jest ustrukturyzowanie przenoszonej cząsteczki w odpowiednią konfigurację przestrzenną zdolną do pełnienia swojej nieodłącznej funkcji. Strukturyzacja ta zachodzi za pomocą specjalnych enzymów lub na matrycy wyspecjalizowanych cząsteczek białka - chaperonów, które pomagają cząsteczce białka, nowo utworzonej lub zmienionej pod wpływem czynników zewnętrznych, uzyskać prawidłową trójwymiarową strukturę. W przypadku niekorzystnego wpływu na komórkę, gdy istnieje możliwość zakłócenia struktury cząsteczek białka (na przykład wraz ze wzrostem temperatury ciała, procesem zakaźnym, zatruciem), wzrasta stężenie białek opiekuńczych w komórce ostro. Dlatego takie cząsteczki są również nazywane białka stresu, Lub białka szoku cieplnego. Naruszenie struktury cząsteczki białka prowadzi do powstania chemicznie obojętnych konglomeratów, które odkładają się w komórce lub na zewnątrz niej podczas amyloidozy, choroby Alzheimera itp. Czasami wstępnie ustrukturyzowana podobna cząsteczka może służyć jako matryca, w tym W takim przypadku, jeśli pierwotna struktura wystąpi nieprawidłowo, wszystkie kolejne cząsteczki również będą wadliwe. Sytuacja taka ma miejsce w tak zwanych chorobach prionowych (trzask u owiec, wściekłych krów, kuru, choroba Creutzfeldta-Jakoba u ludzi), gdy defekt jednego z białek błonowych komórki nerwowej powoduje późniejsze gromadzenie się obojętnych mas wewnątrz komórki i zakłócenie jego funkcji życiowych.
Zakłócenie procesów syntezy w komórce może nastąpić na różnych jej etapach: transkrypcja RNA w jądrze, translacja polipeptydów w rybosomach, modyfikacja potranslacyjna, hipermetylacja i glikozylacja cząsteczki beżowej, transport i dystrybucja białek w komórce oraz ich usuwanie na zewnątrz. W tym przypadku można zaobserwować wzrost lub spadek liczby rybosomów, rozpad polirybosomów, ekspansję cystern ziarnistej siateczki śródplazmatycznej, utratę rybosomów oraz tworzenie pęcherzyków i wakuoli. Zatem po zatruciu blady perkoz enzym polimeraza RNA ulega uszkodzeniu, co zakłóca transkrypcję. Toksyna błonicza inaktywując czynnik wydłużania, zakłóca procesy translacyjne, powodując uszkodzenie mięśnia sercowego. Przyczyną zakłócenia syntezy niektórych określonych cząsteczek białka mogą być czynniki zakaźne. Na przykład wirusy opryszczki hamują syntezę i ekspresję cząsteczek antygenu MHC, co pozwala im częściowo uniknąć kontroli immunologicznej; prątki dżumy - syntezę mediatorów ostrego stanu zapalnego. Pojawienie się nietypowych białek może zatrzymać ich dalszy rozkład i doprowadzić do gromadzenia się materiału obojętnego, a nawet toksycznego. Można to w pewnym stopniu ułatwić poprzez zakłócenie procesów rozkładu.
Zakłócenie procesów rozkładu
Równolegle z syntezą białka w komórce następuje ciągły jego rozpad. W normalnych warunkach ma to istotne znaczenie regulacyjne i kształtujące, na przykład podczas aktywacji nieaktywnych form enzymów, hormonów białkowych i białek cyklu mitotycznego. Normalny wzrost i rozwój komórek wymaga dokładnie kontrolowanej równowagi pomiędzy syntezą i degradacją białek i organelli. Jednak w procesie syntezy białek, na skutek błędów w działaniu aparatu syntetyzującego, nieprawidłowej struktury cząsteczki białka i jej uszkodzenia przez czynniki chemiczne i bakteryjne, stale powstaje dość duża liczba wadliwych cząsteczek. Według niektórych szacunków ich udział stanowi około jednej trzeciej wszystkich syntetyzowanych białek.
Komórki ssaków mają kilka głównych sposoby niszczenia białek: poprzez proteazy lizosomalne (hydrolazy pentydowe), proteinazy zależne od wapnia (endopeptydazy) i układ proteasomów. Ponadto istnieją również wyspecjalizowane proteinazy, takie jak kaspazy. Główną organellą, w której zachodzi degradacja substancji w komórkach eukariotycznych, jest lizosom, który zawiera liczne enzymy hydrolityczne. W wyniku procesów endocytozy i różnego rodzaju autofagii w lizosomach i fagolizosomach niszczone są zarówno wadliwe cząsteczki białek, jak i całe organelle: uszkodzone mitochondria, odcinki błony komórkowej, niektóre białka zewnątrzkomórkowe oraz zawartość ziarnistości wydzielniczych.
Ważnym mechanizmem degradacji białek jest proteasom, multikatalityczna struktura proteinazowa o złożonej budowie, zlokalizowana w cytozolu, jądrze, siateczce śródplazmatycznej i na błonie komórkowej. Ten układ enzymatyczny jest odpowiedzialny za rozkładanie uszkodzonych białek, a także zdrowych białek, które muszą zostać usunięte, aby komórka mogła normalnie funkcjonować. W tym przypadku białka przeznaczone do zniszczenia są wstępnie łączone ze specyficznym polipeptydem, ubikwityną. Jednakże nieubikwitynowane białka mogą również zostać częściowo zniszczone w proteasomach. Rozpad cząsteczek białek w proteasomach na krótkie polipeptydy (przetwarzanie) i ich późniejsza prezentacja wraz z cząsteczkami MHC typu I jest ważnym ogniwem w kontroli immunologicznej homeostazy antygenowej w organizmie. Kiedy funkcja proteasomu jest osłabiona, gromadzą się uszkodzone i niepotrzebne białka, co towarzyszy starzeniu się komórek. Naruszenie degradacji białek zależnych od cyklin prowadzi do zakłócenia podziału komórek, degradacji białek wydzielniczych - do rozwoju cystofibrozy. I odwrotnie, wzrost funkcji proteasomów towarzyszy wyczerpywaniu się organizmu (AIDS, nowotwór).
Przy genetycznie uwarunkowanych zaburzeniach degradacji białek organizm nie jest zdolny do życia i umiera we wczesnych stadiach embriogenezy. Jeśli rozkład tłuszczów lub węglowodanów zostanie zakłócony, pojawiają się choroby spichrzeniowe (tezauryzmoza). W takim przypadku wewnątrz komórki gromadzi się nadmierna ilość niektórych substancji lub produktów ich niepełnego rozkładu - lipidów, polisacharydów - co znacznie uszkadza funkcję komórki. Najczęściej obserwuje się to w komórkach nabłonka wątroby (hepatocytach), neuronach, fibroblastach i makrofagocytach.
Nabyte zaburzenia procesów rozkładu substancji mogą powstać w wyniku procesów patologicznych (na przykład zwyrodnienia białek, tłuszczów, węglowodanów i barwników) i towarzyszy im tworzenie niezwykłych substancji. Zaburzenia w układzie proteolizy lizosomalnej prowadzą do zmniejszonej adaptacji podczas postu lub wzmożonego stresu, a także do wystąpienia niektórych dysfunkcji endokrynologicznych - obniżonego poziomu insuliny, tyreoglobuliny, cytokin i ich receptorów. Upośledzona degradacja białek spowalnia tempo gojenia się ran, powoduje rozwój miażdżycy i wpływa na odpowiedź immunologiczną. Z niedotlenieniem, zmianami wewnątrzkomórkowego pH, uszkodzeniem popromiennym, charakteryzującym się zwiększoną peroksydacją lipidów błonowych, a także pod wpływem substancji lizosomotropowych - endotoksyny bakteryjne, metabolity toksycznych grzybów (sporofusaryna), kryształy tlenku krzemu - stabilność błony lizosomalnej W wyniku tych zmian do cytoplazmy uwalniane są aktywowane enzymy lizosomalne, co powoduje zniszczenie struktur komórkowych i ich śmierć.
§ 2. Główne składniki komórki eukariotycznej
Komórki eukariotyczne (ryc. 8 i 9) są zorganizowane znacznie bardziej złożone niż komórki prokariotyczne. Są bardzo zróżnicowane pod względem wielkości (od kilku mikrometrów do kilku centymetrów), kształtu i cech strukturalnych (ryc. 10).
Ryż. 8. Budowa komórki eukariotycznej. Uogólniony schemat
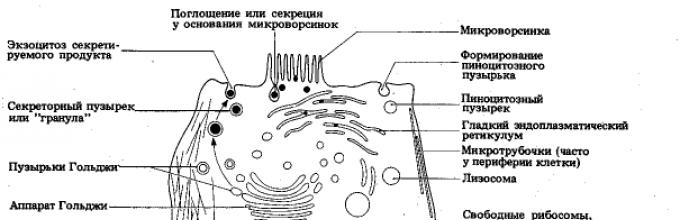
Ryż. 9. Budowa komórki według mikroskopii elektronowej
Ryż. 10. Różne komórki eukariotyczne: 1 – nabłonkowe; 2 – krew (e – erytroza, / – białaczka); 3 – chrząstka; 4 – kości; 5 – mięśnie gładkie; 6 – tkanka łączna; 7 – komórki nerwowe; 8 – włókno mięśni prążkowanych
Jednakże ogólna organizacja i obecność podstawowych składników są takie same we wszystkich komórkach eukariotycznych (ryc. 11).
Ryż. 11. Komórka eukariotyczna (schemat)
Plazmalemma (zewnętrzna błona komórkowa). Podstawą błony komórkowej, podobnie jak innych błon komórkowych (na przykład mitochondriów, plastydów itp.), Jest warstwa lipidów, która ma dwa rzędy cząsteczek (ryc. 12). Ponieważ cząsteczki lipidów są polarne (jeden biegun jest hydrofilowy, czyli przyciągany przez wodę, a drugi hydrofobowy, czyli odpychany przez wodę), są one ułożone w określonej kolejności. Hydrofilowe końce cząsteczek jednej warstwy skierowane są w stronę środowiska wodnego – do cytoplazmy komórki, a drugiej warstwy – na zewnątrz komórki – w stronę substancji międzykomórkowej (w organizmach wielokomórkowych) lub środowiska wodnego (w organizmach jednokomórkowych) ).
Ryż. 12. Budowa błony komórkowej według modelu płynnej mozaiki. Białka i glikoproteiny są zanurzone w podwójnej warstwie cząsteczek lipidów, których hydrofilowe końce (okręgi) są skierowane na zewnątrz, a hydrofobowe (faliste linie) skierowane są w głąb błony
Cząsteczki białek są mozaikowo osadzone w dwucząsteczkowej warstwie lipidów. Na zewnątrz komórki zwierzęcej cząsteczki polisacharydów łączą się z lipidami i cząsteczkami białek plazmalemy, tworząc glikolipidy i glikoproteiny.
To kruszywo tworzy warstwę glikokaliks. Związany z nim funkcja receptora błony plazmatyczne (patrz poniżej); może także gromadzić różne substancje wykorzystywane przez komórkę. Ponadto glikokaliks zwiększa stabilność mechaniczną plazmalemy.
W komórkach roślin i grzybów znajduje się także ściana komórkowa, która pełni rolę wspierającą i ochronną. U roślin składa się z celulozy, u grzybów z chityny.
Zewnętrzna błona komórkowa pełni szereg funkcji, m.in.:
♦ mechaniczny(wspieranie, budowanie form);
♦ transport barierowy(selektywna przepuszczalność różnych substancji: wnikanie do komórki niezbędnych substancji i usuwanie niepotrzebnych i szkodliwych);
♦ chwytnik(oznaczenie różnych substancji chemicznych znajdujących się w pobliżu komórki; percepcja sygnałów w postaci hormonów; rozpoznawanie „obcego” białka przez komórki układu odpornościowego itp.).
Wymiana substancji między komórką a środowiskiem odbywa się na różne sposoby - pasywny i aktywny.
Cząsteczki wody i różnych jonów biernie (w wyniku dyfuzji, osmozy), bez wydatkowania energii przez komórkę, przedostają się przez specjalne pory - są to transport pasywny. Dostarczane są makrocząsteczki, takie jak białka, polisacharydy, a nawet całe komórki fagocytoza I pinocytoza ze zużyciem energii – transport aktywny.
W wyniku fagocytozy pochłaniane są całe komórki lub duże cząstki (na przykład pomyśl o odżywianiu się ameby lub fagocytozie przez ochronne komórki krwi bakterii). Podczas pinocytozy wchłaniane są małe cząsteczki lub kropelki ciekłej substancji. Wspólne dla obu procesów jest to, że wchłonięte substancje są otoczone wklęsłą błoną zewnętrzną, tworząc wakuolę, która następnie przemieszcza się w głąb cytoplazmy komórki.
Egzocytoza jest procesem (będącym także transportem aktywnym) przeciwnym do fagocytozy i pinocytozy (ryc. 13). Za jego pomocą można usunąć niestrawione resztki pokarmu z pierwotniaków lub substancje biologicznie czynne powstałe w komórce wydzielniczej.
Cytoplazma. Cytoplazma to zawartość komórki ograniczona przez plazmalemmę, z wyjątkiem jądra. Zawiera substancja podstawowa (hialoplazma), organelle I inkluzje.
Hialoplazma- lepka ciecz, która może znajdować się w jednym lub drugim stanie Zola(płyn) lub żel(galaretowaty).
Jeśli to konieczne, cytoplazma jest zdolna do odwracalnego przejścia z jednego stanu do drugiego. Na przykład podczas ruchu ameboidów (pamiętajcie sekcję „pierwotniaki” z kursu zoologii), podczas tworzenia pseudopoda, zachodzą szybkie przejścia cytoplazmy z żelu do zolu i odwrotnie. Wynika to z obecności w cytoplazmie dużej liczby cząsteczek białka nitkowatego aktina. Kiedy łączą się ze sobą, tworząc trójwymiarową sieć, cytoplazma znajduje się w stanie żelowym, a gdy sieć rozpada się, znajduje się w stanie zolu.
Hialoplazma zawiera różne substancje - enzymy, białka, węglowodany, tłuszcze i inne, organiczne i mineralne. Zachodzą tu różnorodne procesy chemiczne – rozkład substancji, ich synteza i modyfikacje (zmiany).
Organoidy. Są to stałe składniki komórki o określonej strukturze i funkcjach, zlokalizowane w jej cytoplazmie. W dalszej części omówimy organoidy ogólnego przeznaczenia, nieodłącznie związany ze wszystkimi typami komórek wszystkich eukariontów. Są one związane z zapewnieniem funkcji życiowych tego ostatniego. Organoidy specjalnego przeznaczenia występuje tylko w komórkach określonego (wysoce wyspecjalizowanego) typu - na przykład miofibryli w komórkach mięśniowych.
Organelle ogólnego przeznaczenia mają tę samą strukturę niezależnie od tego, do jakich komórek i organizmów należą. Ale wśród nich są grupy z membraną (retikulum endoplazmatyczne, aparat Golgiego, mitochondria, plastydy, lizosomy, wakuole), jak i bezmembranowe ( rybosomy, centrum komórkowe) Struktura.
Siateczka śródplazmatyczna (ER). EPS składa się z błon i jest złożonym, rozgałęzionym systemem kanalików i cystern, który przenika całą cytoplazmę komórki (ryc. 14). Istnieją dwa rodzaje EPS - surowy I gładki. Rybosomy są przyczepione do błon szorstkich (od strony cytoplazmatycznej), natomiast nie występują na błonach gładkich.
Ryż. 14. Siateczka śródplazmatyczna
Siateczka śródplazmatyczna pełni w komórce eukariotycznej szereg ważnych funkcji:
♦ ograniczające(podział objętości wewnętrznej ogniwa na różne przestrzenie reakcyjne);
♦ udział w syntezie substancji organicznych(rybosomy znajdują się na błonach szorstkiego ER, a kompleksy enzymatyczne znajdują się na gładkich, zapewniając syntezę lipidów, węglowodanów itp.);
♦ udział w tworzeniu elementów aparatu Golgiego, lizosomów;
♦ transport substancji.
Aparat Golgiego. Aparat Golgiego (AG) jest systemem czołgi(płaskie wakuole) i bąbelki(pęcherzyki), zlokalizowane w bliskiej odległości od jądra komórkowego, które powstają w wyniku EPS w wyniku oddzielenia się jego małych fragmentów (ryc. 15). Kiedy te fragmenty się łączą, powstają nowe cysterny aparatu Golgiego, a z EPS transportowane są różne substancje, które biorą udział w składaniu złożonych związków organicznych (białka + węglowodany, białka + lipidy itp.), które są usuwane wraz z pomoc AG poza komórką. Te biologicznie aktywne substancje są albo usuwane z komórki (przy użyciu wakuoli wydzielniczych w wyniku egzocytozy), albo wchodzą w skład lizosomów (patrz poniżej) utworzonych przez AG.
Ryż. 15. Aparat Golgiego:
Aparat Golgiego spełnia następujące funkcje:
♦ synteza substancje biologicznie czynne wytwarzane przez komórkę;
♦ wydzielanie (usuwanie z komórki) różnych substancji(hormony, enzymy, substancje, z których zbudowana jest ściana komórkowa itp.);
♦ udział w tworzeniu lizosomów.
Mitochondria. Wszystkie typy komórek eukariotycznych mają mitochondria (ryc. 16). Wyglądają jak okrągłe korpusy lub pręty, rzadziej - nici. Ich rozmiary wahają się od 1 do 7 mikronów. Liczba mitochondriów w komórce waha się od kilkuset do kilkudziesięciu tysięcy (w przypadku dużych pierwotniaków).
Ryż. 16. Mitochondria. Powyżej - mitochondria (a) w kanałach moczowych widoczne w mikroskopie świetlnym. Poniżej znajduje się trójwymiarowy model organizacji mitochondriów: 1 – cristae; 2 – membrana zewnętrzna; 3 – membrana wewnętrzna; 4 – macierz
Mitochondrium zbudowane jest z dwóch błon - zewnętrzny I wewnętrzny, pomiędzy którymi się znajduje przestrzeń międzybłonowa. Wewnętrzna membrana tworzy wiele wgłębień - cristae, które są albo płytkami, albo rurkami. Ta organizacja zapewnia ogromny obszar błony wewnętrznej. Zawiera enzymy, które zapewniają przemianę energii zawartej w substancjach organicznych (węglowodany, lipidy) w energię ATP, niezbędną do życia komórki. Dlatego funkcją mitochondriów jest uczestnictwo energia procesy komórkowe. Dlatego duża liczba mitochondriów jest nieodłączna na przykład w komórkach mięśniowych, które wykonują dużo pracy.
Plastydy. W komórkach roślinnych znajdują się specjalne organelle - plastydy, które często mają wrzecionowaty lub zaokrąglony kształt, czasem bardziej złożony. Istnieją trzy rodzaje plastydów - chloroplasty (ryc. 17), chromoplasty i leukoplasty.
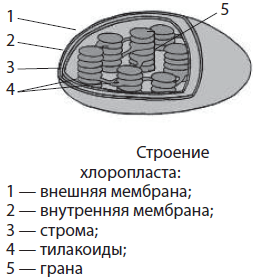
Chloroplasty różnią się zielonym kolorem, co wynika z pigmentu - chlorofil, zapewnienie procesu fotosynteza, czyli synteza substancji organicznych z wody (H 2 O) i dwutlenku węgla (CO 2) przy wykorzystaniu energii światła słonecznego. Chloroplasty występują głównie w komórkach liści (u roślin wyższych). Tworzą je dwie równoległe do siebie membrany, otaczające zawartość chloroplastów - zrąb. Błona wewnętrzna tworzy liczne spłaszczone worki - tylakoidy, które są ułożone (jak stos monet) – ziarna - i leżeć w zrębie. To tylakoidy zawierają chlorofil.
Chromoplasty określają żółty, pomarańczowy i czerwony kolor wielu kwiatów i owoców, w komórkach których są obecne w dużych ilościach. Głównymi pigmentami w ich składzie są karoteny. Funkcjonalnym celem chromoplasty jest przyciąganie zwierząt kolorem, zapewnienie zapylenia kwiatów i rozsiewanie nasion.
Ryż. 17. Plastydy: a – chloroplasty w komórkach liścia Elodea, widoczne w mikroskopie świetlnym; b – schemat wewnętrznej struktury chloroplastu z granami, które są stosami płaskich worków ułożonych prostopadle do powierzchni chloroplastu; c – bardziej szczegółowy schemat przedstawiający rurki zespalające łączące poszczególne komory wentylatora
Leukoplasty to bezbarwne plastydy zawarte w komórkach podziemnych części roślin (na przykład bulw ziemniaka), nasionach i rdzeniu łodyg. W leukoplastach skrobia powstaje głównie z glukozy i gromadzi się w organach spichrzowych roślin.
Plastydy jednego typu mogą przekształcić się w inny. Na przykład, gdy liście zmieniają kolor jesienią, chloroplasty przekształcają się w chromoplasty.
Lizosomy. Organelle te wyglądają jak pęcherzyki otoczone błoną o średnicy do 2 mikronów. Zawierają kilkadziesiąt enzymów rozkładających białka, kwasy nukleinowe, polisacharydy i lipidy. Funkcją lizosomów jest udział w procesach wewnątrzkomórkowego rozkładu złożonych związków organicznych (na przykład składników odżywczych lub substancji „zużytych” składników komórkowych). Lizosomy łączą się z wakuolami fagocytarnymi (lub pinocytowymi), tworząc wakuolę trawienną.
Tworzenie lizosomów następuje w wyniku pączkowania z cystern aparatu Golgiego.
Rybosomy. Rybosomy (ryc. 18) są obecne w komórkach zarówno eukariontów, jak i prokariotów, ponieważ pełnią ważną funkcję w biosynteza białek(patrz rozdział 5). Każda komórka zawiera dziesiątki, setki tysięcy (aż do kilku milionów) tych małych okrągłych organelli.
Ryż. 18. Schemat budowy rybosomu znajdującego się na błonie retikulum endoplazmatycznego: 1 – podjednostka mała; 2 – tRNA; 3 – aminoacylo-tRNA; 4 – aminokwas; 5 – duża podjednostka; 6 – błona siateczki śródplazmatycznej; 7 – zsyntetyzowany łańcuch polipeptydowy
Rybosom składa się z dwóch nierównych podjednostek (części). Tworzą się one oddzielnie i łączą, „obejmując” informacyjny RNA, podczas syntezy cząsteczki białka. Rybosomy zawierają różne białka i rybosomalne RNA.
Inkluzje komórkowe. Taką nazwę nadano nietrwałym składnikom komórki, występującym w głównej substancji cytoplazmy w postaci ziaren, granulek lub kropelek. Inkluzje mogą być otoczone błoną lub nie.
Funkcjonalnie istnieją trzy typy inkluzji: rezerwować składniki odżywcze(skrobia, glikogen, tłuszcze, białka), wtrącenia wydzielnicze(substancje charakterystyczne dla wytwarzanych przez nie komórek gruczołowych - hormony gruczołów dokrewnych itp.) oraz włączenie specjalnego przeznaczenia(w wysoce wyspecjalizowanych komórkach, np. hemoglobinie w czerwonych krwinkach).
§ 3. Organizacja jądra komórkowego. Chromosomy
Jądro komórkowe (patrz ryc. 8 i 9) ma ogromne znaczenie w życiu komórki, ponieważ służy jako repozytorium dziedzicznej informacji zawartej w chromosomach (patrz poniżej).
Jądro jest otoczone otoczką jądrową oddzielającą jego zawartość (karioplazma) z cytoplazmy. Powłoka składa się z dwóch membran oddzielonych szczeliną. Obydwa są przesiąknięte licznymi porami, dzięki czemu możliwa jest wymiana substancji pomiędzy jądrem a cytoplazmą. W jądrze komórkowym większości eukariontów jest od 1 do 7 jąderka. Związane są z nimi procesy syntezy RNA i tRNA.
Główne składniki jądra – chromosomy, utworzone z cząsteczki DNA i różnych białek. W mikroskopie świetlnym są one wyraźnie widoczne jedynie w okresie podziału komórki (mitoza, mejoza). W niedzielącej się komórce chromosomy wyglądają jak długie cienkie nitki rozmieszczone w całej objętości jądra.
Podczas podziału komórki nici chromosomalne tworzą gęste spirale, w wyniku czego stają się widoczne (przy użyciu konwencjonalnego mikroskopu) w postaci pręcików, „szpilek do włosów”. Cała objętość informacji genetycznej jest rozdzielona pomiędzy chromosomami jądra. W trakcie ich badania zidentyfikowano następujące wzorce:
♦ w jądrach komórek somatycznych (tj. komórek organizmu niepłciowych) wszystkie osobniki tego samego gatunku zawierają tę samą liczbę chromosomów, które tworzą zestaw chromosomów(ryc. 19);
Ryż. 19. Chromosomy różnych gatunków roślin i zwierząt przedstawione w tej samej skali: 1,2 – ameba; 3,4 – okrzemki; 5–8, 18,19 – algi zielone; 9 – muchomor; 10 – lipa; 11–12 – Drosophila; 13 – łosoś; 14 – skerda (rodzina Asteraceae); 15 – roślina z rodziny aroidów; 16 – motyl kokoryczkowy; 17 – owad z rodziny szarańczy; 20 – błąd nartnika; 21 – błąd kwiatowy; 22 – ambystoma płazów; 23 – aloes (rodzina liliowatych)
♦ każdy gatunek charakteryzuje się własnym zestawem chromosomów w zależności od ich liczby (na przykład osoba ma 46 chromosomów, mucha Drosophila ma 8, glista ma 4, rak ma 196, koń ma 66 i kukurydza ma 104);
♦ chromosomy w jądrach komórek somatycznych można grupować w pary tzw chromosomy homologiczne w oparciu o ich podobieństwo (pod względem struktury i funkcji);
♦ w jądrach komórek rozrodczych (gametach) każda para homologicznych chromosomów zawiera tylko jeden, tj. całkowity zestaw chromosomów jest o połowę mniejszy niż komórki somatyczne;
♦ nazywa się pojedynczy zestaw chromosomów w komórkach rozrodczych haploidalny i jest oznaczony literą n, a somatycznie - diploidalny(2n).
Z powyższego jasno wynika, że każda para homologicznych chromosomów powstaje w wyniku połączenia chromosomów ojcowskich i matczynych podczas zapłodnienia, tj. fuzji komórek płciowych (gamet). I odwrotnie, podczas tworzenia komórek rozrodczych z każdej pary homologicznych chromosomów tylko jeden dostaje się do gamet.
Chromosomy różny pary homologiczne różnią się wielkością i kształtem (ryc. 20 i 21).
Ryż. 20. Budowa i rodzaje chromosomów: a – wygląd 1 – centromer; 2 – krótkie ramię; 3 – długie ramię); struktura wewnętrzna tego samego chromosomu (1 – centromer; – cząsteczki DNA); c – rodzaje chromosomów (1 – jednoramienny; różnoramienny; 3 – równoramienny: X – ramię, V – centromer)
Ryż. 21. Chromosom składa się z DNA i białek. Cząsteczka DNA ulega replikacji. Dwie identyczne podwójne helisy DNA pozostają połączone w regionie centromeru. Kopie te stają się oddzielnymi chromosomami później podczas podziału komórki
W ciele chromosomów są zwężenie pierwotne (zwane centromerem), do którego przymocowane są nici wrzeciona rozszczepienia. Dzieli chromosom na dwa ramię Chromosomy mogą być równoramienne, nierówne lub jednoramienne.
Rozdział 5. Metabolizm
§ 1. Metabolizm jako jedność asymilacji i dysymilacji
Wszystkie komórki i organizmy żywe są układami otwartymi, to znaczy znajdują się w stanie ciągłej wymiany energii i substancji z otoczeniem. W przyrodzie nieożywionej istnieją systemy otwarte, ale ich istnienie różni się jakościowo od organizmów żywych. Rozważmy ten przykład: płonący kawałek rodzimej siarki znajduje się w stanie wymiany ze środowiskiem. Podczas spalania O 2 jest pochłaniany, a uwalniany jest SO 2 i energia (w postaci ciepła). Jednak w tym przypadku kawałek siarki jako ciało fizyczne ulega zniszczeniu i traci swoją pierwotną strukturę.
Dla organizmów żywych wymiana z otoczeniem okazuje się warunkiem zachowania i utrzymania ich organizacji strukturalnej poprzez samoodnowę wszystkich substancji i składników, z których się składają.
Metabolizm (metabolizm) to zespół procesów zachodzących w organizmach żywych (konsumpcja, przemiana, gromadzenie i uwalnianie substancji i energii), które zapewniają ich żywotną aktywność, rozwój, wzrost, reprodukcję. Podczas procesu metabolicznego cząsteczki tworzące komórki są rozkładane i syntetyzowane; odnowa struktur komórkowych i substancji międzykomórkowej.
Metabolizm opiera się na wzajemnie powiązanych procesach asymilacja(anabolizm) i dysymilacja(katabolizm). Podczas asymilacji (wymiany plastycznej) z prostych powstają substancje złożone. To dzięki temu powstają w komórce wszystkie substancje organiczne niezbędne do budowy jej elementów strukturalnych, układów enzymatycznych itp. Asymilacja zawsze następuje z wydatkiem energii.
Podczas dysymilacji (metabolizmu energetycznego) złożone substancje organiczne rozkładają się na prostsze lub nieorganiczne. W tym przypadku uwalniana jest energia, którą komórka wykorzystuje do wykonywania różnych procesów zapewniających jej żywotną aktywność (synteza i transport substancji, praca mechaniczna itp.).
Wszystkie żywe organizmy można podzielić na dwie grupy: autotrofy I heterotrofy, które różnią się źródłami energii i substancjami niezbędnymi do zapewnienia swoich funkcji życiowych.
Autotrofy– organizmy syntetyzujące związki organiczne z substancji nieorganicznych wykorzystując energię światła słonecznego (np fototrofy– rośliny, sinice) lub energię uzyskaną z utleniania substancji mineralnych (nieorganicznych) (np chemotrofy– bakterie siarkowe, bakterie żelazowe itp.). Dzięki temu są w stanie samodzielnie wytwarzać substancje niezbędne do ich życiowej aktywności.
§ 2. Dysymilacja u organizmów beztlenowych i tlenowych
Organizmy można podzielić na dwie grupy i ze względu na charakter dysymilacji - aeroby I beztlenowce. Aeroby (z greckiego. Demon– powietrze) wymagają do życia wolnego tlenu. U beztlenowców (gr. AC– cząstka ujemna) nie jest to konieczne. W nich dysymilacja odbywa się poprzez fermentację - beztlenowy, enzymatyczny rozkład materii organicznej z utworzeniem prostszych substancji organicznych i uwolnieniem energii. Na przykład:
♦ fermentacja mlekowa:
C 6H 12O 6 + 2H 3PO 4 + 2ADP → 2PH + 2ATP + 2H 2O;
♦ fermentacja alkoholowa:
C 6 H 12 O 6 + 2PH + 2ADP → 2C 2 H 5 OH + 2CO 2 + 2ATP + 2H 2 O.
Substancje powstające podczas fermentacji są organiczne i dlatego nadal zawierają dużo energii.
Ryż. 22. Zależności między asymilacją i dysymilacją u organizmów autotroficznych i heterotroficznych
U organizmów tlenowych podczas procesu oddychania w mitochondriach następuje całkowity rozkład substancji organicznych (przy użyciu O 2) na ubogie w energię produkty końcowe CO 2 i H 2 O i uwalniana jest znacznie większa ilość energii:
C 6 H 12 0 6 (glukoza) + 0 2 > 6C0 2 + 6H 2 0 + energia (dzięki czemu syntetyzuje się 38 cząsteczek ATP).
Rozważmy w formie uogólnionych schematów metabolizm autotroficznych i heterotroficznych organizmów tlenowych (ryc. 22).
Asymilacja. Najważniejszymi procesami są fotosynteza I biosynteza białek.
Zapraszamy do zapoznania się z materiałami i.
: błona celulozowa, błona, cytoplazma z organellami, jądro, wakuole z sokiem komórkowym.Główną cechą komórki roślinnej jest obecność plastydów.
Funkcje błony komórkowej- określa kształt komórki, chroni przed czynnikami środowiskowymi.
Membrana plazmowa- cienki film, składający się z oddziałujących na siebie cząsteczek lipidów i białek, oddziela zawartość wewnętrzną od środowiska zewnętrznego, zapewnia transport wody, minerałów i substancji organicznych do komórki na drodze osmozy i transportu aktywnego, a także usuwa produkty przemiany materii.
Cytoplazma- wewnętrzne półpłynne środowisko komórki, w którym znajduje się jądro i organelle, zapewnia połączenia między nimi i uczestniczy w podstawowych procesach życiowych.
Siateczka endoplazmatyczna- sieć rozgałęzionych kanałów w cytoplazmie. Bierze udział w syntezie białek, lipidów i węglowodanów oraz w transporcie substancji. Rybosomy to ciała zlokalizowane w ER lub cytoplazmie, składające się z RNA i białka i biorące udział w syntezie białek. EPS i rybosomy stanowią pojedynczy aparat do syntezy i transportu białek.
Mitochondria- organelle oddzielone od cytoplazmy dwiema błonami. Utleniają się w nich substancje organiczne i syntetyzują cząsteczki ATP przy udziale enzymów. Zwiększenie powierzchni błony wewnętrznej, na której znajdują się enzymy, z powodu cristae. ATP jest substancją organiczną bogatą w energię.
Plastydy(chloroplastów, leukoplasty, chromoplasty), ich zawartość w komórce jest główną cechą organizmu roślinnego. Chloroplasty to plastydy zawierające zielony pigment chlorofil, który pochłania energię świetlną i wykorzystuje ją do syntezy substancji organicznych z dwutlenku węgla i wody. Chloroplasty oddzielone są od cytoplazmy dwiema błonami, licznymi naroślami - grana na błonie wewnętrznej, w której znajdują się cząsteczki chlorofilu i enzymy.
Kompleks Golgiego- układ wnęk oddzielony od cytoplazmy błoną. Akumulacja w nich białek, tłuszczów i węglowodanów. Przeprowadzanie syntezy tłuszczów i węglowodanów na błonach.
Lizosomy- ciała oddzielone od cytoplazmy pojedynczą błoną. Zawarte w nich enzymy przyspieszają rozkład cząsteczek złożonych na proste: białek na aminokwasy, węglowodanów złożonych na proste, lipidów na glicerol i kwasy tłuszczowe, a także niszczą martwe części komórki i całe komórki.
Wakuole- wnęki w cytoplazmie wypełnione sokiem komórkowym, miejsce gromadzenia rezerwowych składników odżywczych i substancji szkodliwych; regulują zawartość wody w komórce.
Rdzeń- główna część komórki, pokryta na zewnątrz dwumembranową, perforowaną otoczką jądrową. Substancje dostają się do rdzenia i są z niego usuwane przez pory. Chromosomy są nośnikami dziedzicznej informacji o cechach organizmu, głównych strukturach jądra, z których każda składa się z jednej cząsteczki DNA połączonej z białkami. Jądro jest miejscem syntezy DNA, mRNA i rRNA.

Obecność błony zewnętrznej, cytoplazmy z organellami i jądra z chromosomami.
Błona zewnętrzna lub plazmatyczna- oddziela zawartość komórki od otoczenia (inne komórki, substancja międzykomórkowa), składa się z cząsteczek lipidów i białek, zapewnia komunikację między komórkami, transport substancji do komórki (pinocytoza, fagocytoza) i na zewnątrz komórki.
Cytoplazma- wewnętrzne półpłynne środowisko komórki, które zapewnia komunikację między jądrem a znajdującymi się w nim organellami. Główne procesy życiowe zachodzą w cytoplazmie.
Organelle komórkowe:
1) retikulum endoplazmatyczne (ER)- układ kanalików rozgałęzionych, bierze udział w syntezie białek, lipidów i węglowodanów, w transporcie substancji w komórce;
2) rybosomy- ciała zawierające rRNA znajdują się na ER i cytoplazmie i uczestniczą w syntezie białek. EPS i rybosomy to pojedynczy aparat do syntezy i transportu białek;
3) mitochondria- „elektrownie” komórki, oddzielone od cytoplazmy dwiema błonami. Wewnętrzna tworzy cristae (fałdy), zwiększając jej powierzchnię. Enzymy na cristae przyspieszają utlenianie substancji organicznych i syntezę bogatych w energię cząsteczek ATP;
4) Kompleks Golgiego- grupa wnęk ograniczonych błoną cytoplazmy, wypełnionych białkami, tłuszczami i węglowodanami, które są albo wykorzystywane w procesach życiowych, albo usuwane z komórki. Błony kompleksu przeprowadzają syntezę tłuszczów i węglowodanów;
5) lizosomy- ciała wypełnione enzymami przyspieszają rozkład białek na aminokwasy, lipidów na glicerol i kwasy tłuszczowe, polisacharydów na monosacharydy. W lizosomach martwe części komórki, całe komórki, ulegają zniszczeniu.
Inkluzje komórkowe- nagromadzenie rezerwowych składników odżywczych: białek, tłuszczów i węglowodanów.
Rdzeń- najważniejsza część komórki. Jest pokryty podwójną błoną z porami, przez które niektóre substancje wnikają do jądra, a inne do cytoplazmy. Chromosomy są głównymi strukturami jądra, nośnikami dziedzicznej informacji o cechach organizmu. Przenoszona jest podczas podziału komórki macierzystej na komórki potomne, a wraz z komórkami rozrodczymi na organizmy potomne. Jądro jest miejscem syntezy DNA, mRNA i rRNA.
Ćwiczenia:
Wyjaśnij, dlaczego organelle nazywane są wyspecjalizowanymi strukturami komórkowymi?
Odpowiedź: organelle nazywane są wyspecjalizowanymi strukturami komórkowymi, ponieważ pełnią ściśle określone funkcje, informacja dziedziczna jest przechowywana w jądrze, ATP jest syntetyzowany w mitochondriach, fotosynteza zachodzi w chloroplastach itp.
Jeśli masz pytania dotyczące cytologii, możesz się skontaktować
Teoria do zadania 5 z Unified State Exam z biologii
Struktura komórkowa. Związek między strukturą i funkcjami części i organelli komórki jest podstawą jej integralności
Struktura komórkowa
Budowa komórek prokariotycznych i eukariotycznych
Głównymi składnikami strukturalnymi komórek są błona plazmatyczna, cytoplazma i aparat dziedziczny. W zależności od cech organizacji wyróżnia się dwa główne typy komórek: prokariotyczne i eukariotyczne. Główną różnicą między komórkami prokariotycznymi a komórkami eukariotycznymi jest organizacja ich dziedzicznego aparatu: u prokariotów znajduje się on bezpośrednio w cytoplazmie (ten obszar cytoplazmy nazywa się nukleoid) i nie jest od niego oddzielony strukturami błonowymi, podczas gdy u eukariontów większość DNA jest skoncentrowana w jądrze otoczonym podwójną błoną. Ponadto informacja genetyczna komórek prokariotycznych, zlokalizowana w nukleoidzie, jest zapisana w kolistej cząsteczce DNA, podczas gdy u eukariotów cząsteczki DNA są otwarte.
W odróżnieniu od eukariotów cytoplazma komórek prokariotycznych zawiera również niewielką liczbę organelli, natomiast komórki eukariotyczne charakteryzują się znacznym zróżnicowaniem tych struktur.
Budowa i funkcje błon biologicznych
Struktura biomembrany. Błony otaczające komórki i organelle błonowe komórek eukariotycznych mają wspólny skład chemiczny i strukturę. Należą do nich lipidy, białka i węglowodany. Lipidy błonowe reprezentowane są głównie przez fosfolipidy i cholesterol. Większość białek błonowych to białka złożone, takie jak glikoproteiny. Węglowodany nie występują w błonie samodzielnie, są związane z białkami i lipidami. Grubość membran wynosi 7-10 nm.
Zgodnie z obecnie ogólnie przyjętym modelem mozaiki płynnej budowy błony, lipidy tworzą podwójną warstwę, czyli dwuwarstwa lipidowa, w którym hydrofilowe „głowy” cząsteczek lipidów są zwrócone na zewnątrz, a hydrofobowe „ogony” są ukryte wewnątrz błony. Te „ogony” dzięki swojej hydrofobowości zapewniają oddzielenie faz wodnych środowiska wewnętrznego komórki od jej otoczenia. Białka są powiązane z lipidami poprzez różnego rodzaju interakcje. Niektóre białka znajdują się na powierzchni błony. Takie białka nazywane są peryferyjny, Lub powierzchowny. Inne białka są częściowo lub całkowicie zanurzone w błonie - są to całka, Lub zanurzone białka. Białka błonowe pełnią funkcje strukturalne, transportowe, katalityczne, receptorowe i inne.
Błony nie przypominają kryształów, ich składniki są w ciągłym ruchu, w wyniku czego pomiędzy cząsteczkami lipidów pojawiają się szczeliny - pory, przez które różne substancje mogą przedostawać się lub wychodzić z komórki.
Błony biologiczne różnią się umiejscowieniem w komórce, składem chemicznym i funkcjami. Głównymi rodzajami membran są plazma i wewnętrzne. Membrana plazmowa zawiera około 45% lipidów (w tym glikolipidów), 50% białek i 5% węglowodanów. Łańcuchy węglowodanów wchodzące w skład złożonych białek – glikoprotein i złożonych lipidów – glikolipidów wystają ponad powierzchnię błony. Glikoproteiny osocza są niezwykle specyficzne. Służą na przykład do wzajemnego rozpoznawania komórek, w tym plemnika i komórki jajowej.
Na powierzchni komórek zwierzęcych łańcuchy węglowodanowe tworzą cienką warstwę powierzchniową - glikokaliks. Jest wykrywany w prawie wszystkich komórkach zwierzęcych, ale stopień jego ekspresji jest zróżnicowany (10-50 µm). Glikokaliks zapewnia bezpośrednią komunikację między komórką a środowiskiem zewnętrznym, gdzie zachodzi trawienie zewnątrzkomórkowe; Receptory znajdują się w glikokaliksie. Oprócz plazmalemy komórki bakterii, roślin i grzybów są również otoczone błonami komórkowymi.
Membrany wewnętrzne komórki eukariotyczne oddzielają różne części komórki, tworząc osobliwe „przedziały” - przegródki, co sprzyja rozdzieleniu różnych procesów metabolicznych i energetycznych. Mogą różnić się składem chemicznym i funkcjami, ale ich ogólny plan strukturalny pozostaje taki sam.
Funkcje membrany:
- Ograniczające. Ideą jest to, że oddzielają przestrzeń wewnętrzną komórki od środowiska zewnętrznego. Błona jest półprzepuszczalna, to znaczy tylko te substancje, których potrzebuje komórka, mogą przez nią swobodnie przechodzić i istnieją mechanizmy transportu niezbędnych substancji.
- Chwytnik. Związany jest przede wszystkim z percepcją sygnałów środowiskowych i przekazywaniem tych informacji do komórki. Za tę funkcję odpowiadają specjalne białka receptorowe. Białka błonowe odpowiadają także za rozpoznawanie komórek na zasadzie „przyjaciel czy wróg”, a także za tworzenie połączeń międzykomórkowych, z których najlepiej zbadanymi są synapsy komórek nerwowych.
- Katalityczny. Na membranach zlokalizowane są liczne kompleksy enzymatyczne, w wyniku czego zachodzą na nich intensywne procesy syntetyczne.
- Transformacja energii. Związany z powstawaniem energii, jej magazynowaniem w postaci ATP i zużyciem.
- Podział na przedziały. Błony wyznaczają również przestrzeń wewnątrz komórki, oddzielając w ten sposób materiały wyjściowe reakcji i enzymy, które mogą przeprowadzić odpowiednie reakcje.
- Tworzenie kontaktów międzykomórkowych. Pomimo tego, że grubość membrany jest tak mała, że nie można jej odróżnić gołym okiem, z jednej strony stanowi ona dość niezawodną barierę dla jonów i cząsteczek, zwłaszcza rozpuszczalnych w wodzie, a z drugiej , zapewnia ich transport do i z komórki.
- Transport.

Transport membranowy. Ze względu na to, że komórki jako elementarne układy biologiczne są układami otwartymi, aby zapewnić metabolizm i energię, utrzymać homeostazę, wzrost, drażliwość i inne procesy, wymagany jest transfer substancji przez błonę - transport membranowy. Obecnie transport substancji przez błonę komórkową dzieli się na aktywny, pasywny, endo- i egzocytozę.
Transport pasywny- Jest to rodzaj transportu, który odbywa się bez zużycia energii od wyższych do niższych stężeń. Rozpuszczalne w lipidach małe niepolarne cząsteczki (O 2, CO 2) z łatwością przenikają do wnętrza komórki prosta dyfuzja. Te nierozpuszczalne w lipidach, w tym małe cząstki naładowane, są wychwytywane przez białka nośnikowe lub przechodzą przez specjalne kanały (glukoza, aminokwasy, K+, PO 4 3-). Ten rodzaj transportu pasywnego nazywa się ułatwiona dyfuzja. Woda dostaje się do komórki przez pory w fazie lipidowej, a także przez specjalne kanały wyłożone białkami. Nazywa się transport wody przez membranę przez osmozę.

Osmoza jest niezwykle ważna w życiu komórki, ponieważ jeśli zostanie umieszczona w roztworze o większym stężeniu soli niż w roztworze komórkowym, wówczas woda zacznie opuszczać komórkę i objętość zawartości żywej zacznie się zmniejszać. W komórkach zwierzęcych cała komórka kurczy się, a w komórkach roślinnych cytoplazma pozostaje w tyle za ścianą komórkową, co nazywa się plazmoliza. Kiedy komórkę umieszcza się w roztworze mniej stężonym niż cytoplazma, transport wody następuje w przeciwnym kierunku – do wnętrza komórki. Istnieją jednak ograniczenia rozciągliwości błony cytoplazmatycznej i komórka zwierzęca ostatecznie pęka, podczas gdy komórka roślinna nie pozwala na to ze względu na silną ścianę komórkową. Nazywa się zjawisko wypełnienia całą wewnętrzną przestrzenią komórki zawartością komórkową deplazmoliza. Przygotowując leki, zwłaszcza do podawania dożylnego, należy wziąć pod uwagę wewnątrzkomórkowe stężenie soli, ponieważ może to prowadzić do uszkodzenia komórek krwi (w tym celu stosuje się roztwór soli fizjologicznej o stężeniu 0,9% chlorku sodu). Jest to nie mniej ważne w przypadku hodowli komórek i tkanek, a także narządów zwierząt i roślin.

Transport aktywny postępuje z wydatkowaniem energii ATP od niższego stężenia substancji do wyższego. Odbywa się to za pomocą specjalnych białek pompujących. Białka pompują K + , Na + , Ca 2+ i inne jony przez błonę, co sprzyja transportowi niezbędnych substancji organicznych, a także powstawaniu impulsów nerwowych itp.
Endocytoza- jest to aktywny proces wchłaniania substancji przez komórkę, podczas którego błona tworzy wgłębienia, a następnie tworzą się pęcherzyki błonowe - fagosomy, które zawierają wchłonięte obiekty. Następnie pierwotny lizosom łączy się z fagosomem i tworzy się lizosom wtórny, Lub fagolizosom, Lub wakuola trawienna. Zawartość pęcherzyka jest trawiona przez enzymy lizosomalne, a produkty rozpadu są wchłaniane i przyswajane przez komórkę. Niestrawione pozostałości są usuwane z komórki w drodze egzocytozy. Istnieją dwa główne typy endocytozy: fagocytoza i pinocytoza.
Fagocytoza to proces wychwytywania przez powierzchnię komórki i absorpcji cząstek stałych przez komórkę, oraz pinocytoza- płyny. Fagocytoza zachodzi głównie w komórkach zwierzęcych (zwierzęta jednokomórkowe, leukocyty ludzkie), zapewnia ich odżywianie i często chroni organizm. Przez pinocytozę białka, kompleksy antygen-przeciwciało są wchłaniane podczas reakcji immunologicznych itp. Jednak wiele wirusów przedostaje się do komórki również poprzez pinocytozę lub fagocytozę. W komórkach roślinnych i grzybowych fagocytoza jest praktycznie niemożliwa, ponieważ są one otoczone trwałymi błonami komórkowymi.

Egzocytoza- proces odwrotny do endocytozy. W ten sposób z wakuoli trawiennych uwalniane są niestrawione resztki pokarmu i usuwane są substancje niezbędne do życia komórki i całego organizmu. Na przykład przekazywanie impulsów nerwowych następuje w wyniku uwolnienia przekaźników chemicznych przez neuron wysyłający impuls - mediatorzy, a w komórkach roślinnych w ten sposób wydzielane są węglowodany pomocnicze błony komórkowej.
Ściany komórkowe komórek roślinnych, grzybów i bakterii. Na zewnątrz błony komórka może wydzielać silną strukturę - Błona komórkowa, Lub Ściana komórkowa.
U roślin podstawą ściany komórkowej jest celuloza, pakowane w wiązki po 50-100 cząsteczek. Przestrzenie między nimi wypełnione są wodą i innymi węglowodanami. Ściana komórkowa roślin jest przesiąknięta kanalikami - plazmodesmy, przez które przechodzą błony retikulum endoplazmatycznego. Plazmodesmy wykonują transport substancji między komórkami. Jednakże transport substancji, takich jak woda, może również odbywać się wzdłuż samych ścian komórkowych. Z biegiem czasu w ścianie komórkowej roślin gromadzą się różne substancje, w tym garbniki czy substancje tłuszczopodobne, co prowadzi do zdrewnienia lub suberyzacji samej ściany komórkowej, wyparcia wody i śmierci zawartości komórek. Pomiędzy ścianami komórkowymi sąsiednich komórek roślinnych znajdują się galaretowate przekładki - środkowe płytki, które spajają je i cementują organizm rośliny jako całość. Ulegają zniszczeniu dopiero w procesie dojrzewania owoców i opadania liści.

Tworzą się ściany komórkowe komórek grzybów chityna- węglowodan zawierający azot. Są dość mocne i stanowią zewnętrzny szkielet komórki, ale nadal, podobnie jak u roślin, zapobiegają fagocytozie.
U bakterii ściana komórkowa zawiera węglowodany z fragmentami peptydowymi - murein jednak jego zawartość różni się znacznie pomiędzy różnymi grupami bakterii. Inne polisacharydy mogą być również wydzielane na ścianie komórkowej, tworząc otoczkę śluzową, która chroni bakterie przed wpływami zewnętrznymi.
Błona określa kształt komórki, służy jako podpora mechaniczna, pełni funkcję ochronną, zapewnia właściwości osmotyczne komórki, ograniczając rozciąganie żywej zawartości i zapobiegając pękaniu komórki, które zwiększa się na skutek przedostawania się wody . Ponadto woda i substancje w niej rozpuszczone pokonują ścianę komórkową przed wejściem do cytoplazmy lub odwrotnie, po jej opuszczeniu, przy czym woda transportowana jest przez ściany komórkowe szybciej niż przez cytoplazmę.
Cytoplazma
Cytoplazma- To jest wewnętrzna zawartość komórki. Zanurzone są w nim wszystkie organelle komórkowe, jądro i różne produkty odpadowe.
Cytoplazma łączy ze sobą wszystkie części komórki i zachodzą w niej liczne reakcje metaboliczne. Cytoplazma jest oddzielona od środowiska i podzielona na przedziały błonami, to znaczy komórki mają strukturę błonową. Może występować w dwóch stanach – zolu i żelu. Sol- jest to półpłynny, galaretowaty stan cytoplazmy, w którym procesy życiowe przebiegają najintensywniej, oraz żel- gęstszy, galaretowaty stan, który utrudnia zachodzenie reakcji chemicznych i transport substancji.
Nazywa się płynną część cytoplazmy bez organelli hialoplazma. Hialoplazma, czyli cytozol, to roztwór koloidalny, w którym znajduje się rodzaj zawiesiny dość dużych cząstek, np. białek, otoczonych dipolami cząsteczek wody. Wytrącanie tej zawiesiny nie następuje ze względu na to, że mają one ten sam ładunek i odpychają się.
Organoidy
Organoidy- Są to stałe elementy komórki, które pełnią określone funkcje.
W zależności od cech strukturalnych dzieli się je na membranowe i niemembranowe. Membrana Organelle z kolei dzielą się na jednobłonowe (retikulum endoplazmatyczne, kompleks Golgiego i lizosomy) lub dwubłonowe (mitochondria, plastydy i jądro). Nie membranowe Organellami są rybosomy, mikrotubule, mikrofilamenty i centrum komórkowe. Spośród wymienionych organelli tylko rybosomy są nieodłączne od prokariotów.
Budowa i funkcje jądra. Rdzeń- duża organella z podwójną błoną leżąca w środku komórki lub na jej obrzeżach. Wymiary jądra mogą wynosić od 3-35 mikronów. Kształt jądra jest najczęściej kulisty lub elipsoidalny, ale zdarzają się również jądra w kształcie pręta, wrzecionowate, fasolowe, klapowane, a nawet segmentowane. Niektórzy badacze uważają, że kształt jądra odpowiada kształtowi samej komórki.
Większość komórek ma jedno jądro, ale na przykład w komórkach wątroby i serca mogą być ich dwa, a w wielu neuronach - do 15. Włókna mięśni szkieletowych zwykle zawierają wiele jąder, ale nie są to komórki w pełnym tego słowa znaczeniu, gdyż powstają w wyniku fuzji kilku komórek.
Rdzeń jest otoczony otoczka nuklearna, a jego wewnętrzna przestrzeń jest wypełniona sok nuklearny, Lub nukleoplazma (karioplazma), w którym są zanurzeni chromatyna I jąderko. Jądro pełni tak ważne funkcje, jak przechowywanie i przekazywanie informacji dziedzicznych, a także kontrolowanie życia komórki.

Rolę jądra w przekazywaniu informacji dziedzicznej przekonująco udowodniono w eksperymentach z zieloną algą Acetabularia. W pojedynczej gigantycznej komórce, osiągającej długość 5 cm, wyróżnia się czapkę, łodygę i ryzoid. Ponadto zawiera tylko jedno jądro zlokalizowane w ryzoidzie. W latach trzydziestych XX wieku I. Hemmerling przeszczepił jądro jednego gatunku panewki o kolorze zielonym do ryzoidu innego gatunku, o kolorze brązowym, z którego usunięto jądro. Po pewnym czasie na roślinie z przeszczepionym jądrem wyrosła nowa czapeczka, podobnie jak glony dawcy jądra. Jednocześnie kapelusz lub łodyga oddzielona od ryzoidu i niezawierająca jądra po pewnym czasie obumiera.

Koperta nuklearna utworzone przez dwie membrany - zewnętrzną i wewnętrzną, pomiędzy którymi jest przestrzeń. Przestrzeń międzybłonowa łączy się z wnęką szorstkiej siateczki śródplazmatycznej, a zewnętrzna błona jądra może przenosić rybosomy. Otoczka jądrowa jest przesiąknięta licznymi porami wyłożonymi specjalnymi białkami. Transport substancji odbywa się przez pory: niezbędne białka (w tym enzymy), jony, nukleotydy i inne substancje dostają się do jądra, a cząsteczki RNA, zużyte białka i podjednostki rybosomów opuszczają je. Zatem funkcjami otoczki jądrowej jest oddzielenie zawartości jądra od cytoplazmy, a także regulacja metabolizmu między jądrem a cytoplazmą.
Nukleoplazma nazywana zawartością jądra, w której zanurzona jest chromatyna i jąderko. Jest to roztwór koloidalny, chemicznie przypominający cytoplazmę. Enzymy nukleoplazmy katalizują wymianę aminokwasów, nukleotydów, białek itp. Nukleoplazma jest połączona z hialoplazmą poprzez pory jądrowe. Funkcje nukleoplazmy, podobnie jak hialoplazmy, polegają na zapewnieniu wzajemnego połączenia wszystkich składników strukturalnych jądra i przeprowadzaniu szeregu reakcji enzymatycznych.
Chromatyna zwany zbiorem cienkich włókien i granulek zanurzonych w nukleoplazmie. Można go wykryć jedynie poprzez barwienie, ponieważ współczynniki załamania światła chromatyny i nukleoplazmy są w przybliżeniu takie same. Nazywa się nitkowaty składnik chromatyny euchromatyna i granulowany - heterochromatyna. Euchromatyna jest słabo zwarta, ponieważ odczytywane są z niej informacje dziedziczne, podczas gdy bardziej spiralna heterochromatyna jest genetycznie nieaktywna.
Chromatyna jest strukturalną modyfikacją chromosomów w niedzielącym się jądrze. Zatem chromosomy są stale obecne w jądrze, jedynie ich stan zmienia się w zależności od funkcji, jaką w danym momencie pełni jądro.
Skład chromatyny obejmuje głównie białka nukleoproteinowe (deoksyrybonukleoproteiny i rybonukleoproteiny), a także enzymy, z których najważniejsze są związane z syntezą kwasów nukleinowych i niektórych innych substancji.
Funkcje chromatyny polegają, po pierwsze, na syntezie specyficznych dla danego organizmu kwasów nukleinowych, które kierują syntezą określonych białek, a po drugie, na przenoszeniu właściwości dziedzicznych z komórki macierzystej na komórki potomne, w tym celu nici chromatyny są pakowane w chromosomy podczas procesu podziału.
Jądro- ciało kuliste, dobrze widoczne pod mikroskopem, o średnicy 1-3 mikronów. Powstaje na odcinkach chromatyny, w których kodowana jest informacja o strukturze rRNA i białek rybosomalnych. Często w jądrze znajduje się tylko jedno jąderko, ale w komórkach, w których zachodzą intensywne procesy życiowe, mogą znajdować się dwa lub więcej jąder. Funkcje jąderek to synteza rRNA i składanie podjednostek rybosomalnych poprzez połączenie rRNA z białkami pochodzącymi z cytoplazmy.
Mitochondria- organelle dwubłonowe o kształcie okrągłym, owalnym lub w kształcie pręcika, chociaż występują również organelle w kształcie spirali (w nasieniu). Średnica mitochondriów dochodzi do 1 µm, a długość do 7 µm. Przestrzeń wewnątrz mitochondriów wypełniona jest matrixem. Matryca- To jest główna substancja mitochondriów. Zanurzona jest w nim okrągła cząsteczka DNA i rybosomy. Zewnętrzna błona mitochondriów jest gładka i nieprzepuszczalna dla wielu substancji. Wewnętrzna membrana ma występy - święta, zwiększając powierzchnię membran, aby mogły zachodzić reakcje chemiczne. Na powierzchni błony znajdują się liczne kompleksy białkowe tworzące tzw. łańcuch oddechowy, a także enzymy syntetazy ATP w kształcie grzyba. Etap oddychania tlenowego zachodzi w mitochondriach, podczas którego syntetyzowany jest ATP.

Plastydy- duże organelle z podwójną błoną, charakterystyczne tylko dla komórek roślinnych. Wewnętrzna przestrzeń plastydów jest wypełniona zrąb, Lub matryca. Zrąb zawiera mniej lub bardziej rozwinięty system pęcherzyków błonowych - tylakoidy, które są zebrane w stosy - ziarna, a także własną kolistą cząsteczkę DNA i rybosomy. Istnieją cztery główne typy plastydów: chloroplasty, chromoplasty, leukoplasty i proplastydy.
Chloroplasty- są to zielone plastydy o średnicy 3-10 mikronów, wyraźnie widoczne pod mikroskopem. Występują wyłącznie w zielonych częściach roślin – liściach, młodych łodygach, kwiatach i owocach. Chloroplasty mają zazwyczaj kształt owalny lub elipsoidalny, ale mogą mieć również kształt miseczki, spirali lub nawet płatków. Liczba chloroplastów w komórce wynosi średnio od 10 do 100 sztuk. Jednak np. u niektórych glonów może to być jeden, mieć znaczne wymiary i złożony kształt – wtedy tak się nazywa chromatofor. W innych przypadkach liczba chloroplastów może osiągnąć kilkaset, a ich rozmiary są niewielkie. Kolor chloroplastów wynika z głównego pigmentu fotosyntezy - chlorofil, chociaż zawierają również dodatkowe pigmenty - karotenoidy. Karotenoidy stają się zauważalne dopiero jesienią, kiedy chlorofil zawarty w starzejących się liściach ulega rozkładowi. Główną funkcją chloroplastów jest fotosynteza. Jasne reakcje fotosyntezy zachodzą na błonach tylakoidów, do których przyczepione są cząsteczki chlorofilu, natomiast ciemne reakcje zachodzą w zrębie, gdzie znajdują się liczne enzymy.
Chromoplasty- Są to plastydy żółte, pomarańczowe i czerwone zawierające pigmenty karotenoidowe. Kształt chromoplasty może również znacznie się różnić: mogą być rurowe, kuliste, krystaliczne itp. Chromoplasty nadają kolor kwiatom i owocom roślin, przyciągając zapylacze oraz dystrybutorów nasion i owoców.
Leukoplasty- Są to białe lub bezbarwne plastydy, przeważnie okrągłe lub owalne. Występują powszechnie w niefotosyntetycznych częściach roślin, np. w skórce liści, bulwach ziemniaka itp. Magazynują składniki odżywcze, najczęściej skrobię, ale u niektórych roślin mogą to być białka lub olej.
Plastydy powstają w komórkach roślinnych z proplastidów, które są już obecne w komórkach tkanki edukacyjnej i są małymi ciałami z podwójną błoną. Na wczesnych etapach rozwoju różne rodzaje plastydów są w stanie przekształcać się w siebie: pod wpływem światła leukoplasty bulwy ziemniaka i chromoplasty korzenia marchwi zmieniają kolor na zielony.
Plastydy i mitochondria nazywane są półautonomicznymi organellami komórkowymi, ponieważ mają własne cząsteczki DNA i rybosomy, przeprowadzają syntezę białek i dzielą się niezależnie od podziału komórki. Cechy te tłumaczy się ich pochodzeniem od jednokomórkowych organizmów prokariotycznych. Jednak „niezależność” mitochondriów i plastydów jest ograniczona, ponieważ ich DNA zawiera zbyt mało genów do swobodnej egzystencji, natomiast reszta informacji jest kodowana w chromosomach jądra, co pozwala mu kontrolować te organelle.
Siateczka śródplazmatyczna (ER), Lub retikulum endoplazmatyczne (ER), to organella jednobłonowa, będąca siecią wnęk i kanalików błonowych zajmujących do 30% zawartości cytoplazmy. Średnica kanalików EPS wynosi około 25-30 nm. Istnieją dwa rodzaje EPS – szorstki i gładki. Słaby XPS przenosi rybosomy, gdzie zachodzi synteza białek. Gładki XPS brakuje rybosomów. Jego funkcją jest synteza lipidów i węglowodanów, a także transport, magazynowanie i neutralizacja substancji toksycznych. Jest szczególnie rozwinięta w tych komórkach, w których zachodzą intensywne procesy metaboliczne, np. w komórkach wątroby – hepatocytach – i włóknach mięśni szkieletowych. Substancje syntetyzowane w ER transportowane są do aparatu Golgiego. Montaż błon komórkowych zachodzi również w ER, ale ich tworzenie kończy się w aparacie Golgiego.
Aparat Golgiego, Lub Kompleks Golgiego, jest organellą jednobłonową utworzoną przez system odłączonych od nich płaskich cystern, kanalików i pęcherzyków. Jednostką strukturalną aparatu Golgiego jest dyktyosom- stos zbiorników, z którego jednego bieguna pochodzą substancje z EPS, a z przeciwnego bieguna, po przejściu pewnych przemian, są pakowane w pęcherzyki i wysyłane do innych części komórki. Średnica zbiorników wynosi około 2 mikronów, a średnica małych pęcherzyków wynosi około 20-30 mikronów. Główne funkcje kompleksu Golgiego to synteza niektórych substancji i modyfikacja (zmiana) białek, lipidów i węglowodanów pochodzących z ER, ostateczne tworzenie błon, a także transport substancji w całej komórce, odnowa jej struktur i tworzenie lizosomów. Aparat Golgiego otrzymał swoją nazwę na cześć włoskiego naukowca Camillo Golgiego, który jako pierwszy odkrył tę organellę (1898).

Lizosomy- małe organelle jednobłonowe o średnicy do 1 μm, które zawierają enzymy hydrolityczne biorące udział w trawieniu wewnątrzkomórkowym. Błony lizosomów są słabo przepuszczalne dla tych enzymów, dlatego lizosomy spełniają swoje funkcje bardzo dokładnie i celowo. Biorą więc czynny udział w procesie fagocytozy, tworząc wakuole trawienne, a w przypadku głodu lub uszkodzenia niektórych części komórki trawią je, nie wpływając na inne. Niedawno odkryto rolę lizosomów w procesach śmierci komórek.
Wakuola to wnęka w cytoplazmie komórek roślinnych i zwierzęcych, ograniczona błoną i wypełniona cieczą. W komórkach pierwotniaków znajdują się wakuole trawienne i kurczliwe. Te pierwsze biorą udział w procesie fagocytozy, rozkładając składniki odżywcze. Te ostatnie zapewniają utrzymanie równowagi wodno-solnej poprzez osmoregulację. U zwierząt wielokomórkowych występują głównie wakuole trawienne.
W komórkach roślinnych zawsze obecne są wakuole, otoczone specjalną błoną i wypełnione sokiem komórkowym. Błona otaczająca wakuolę ma podobny skład chemiczny, strukturę i funkcje jak błona plazmatyczna. Sok komórkowy to wodny roztwór różnych substancji nieorganicznych i organicznych, w tym soli mineralnych, kwasów organicznych, węglowodanów, białek, glikozydów, alkaloidów itp. Wakuola może zajmować do 90% objętości komórki i wypychać jądro na obwód. Ta część komórki pełni funkcje magazynujące, wydalnicze, osmotyczne, ochronne, lizosomalne i inne, ponieważ gromadzi składniki odżywcze i produkty przemiany materii, zapewnia dopływ wody oraz utrzymuje kształt i objętość komórki, a także zawiera enzymy rozkładające wiele składników komórkowych. Ponadto biologicznie czynne substancje wakuoli mogą uniemożliwiać wielu zwierzętom spożywanie tych roślin. U wielu roślin wzrost komórek następuje w wyniku pęcznienia wakuoli poprzez wydłużanie.
Wakuole występują także w komórkach niektórych grzybów i bakterii, lecz u grzybów pełnią jedynie funkcję osmoregulacji, natomiast u sinic utrzymują pływalność i biorą udział w procesie asymilacji azotu z powietrza.
Rybosomy- małe organelle niebłonowe o średnicy 15-20 mikronów, składające się z dwóch podjednostek - dużej i małej. Podjednostki rybosomów eukariotycznych są składane w jąderku, a następnie transportowane do cytoplazmy. Rybosomy u prokariotów, mitochondriów i plastydów są mniejsze niż rybosomy u eukariontów. Podjednostki rybosomów obejmują rRNA i białka.

Liczba rybosomów w komórce może sięgać kilkudziesięciu milionów: w cytoplazmie, mitochondriach i plastydach są w stanie wolnym, a na szorstkim ER - w stanie związanym. Biorą udział w syntezie białek, w szczególności realizują proces translacji – biosyntezy łańcucha polipeptydowego na cząsteczce mRNA. Wolne rybosomy syntetyzują białka hialoplazmy, mitochondriów, plastydów i własne białka rybosomalne, podczas gdy rybosomy przyłączone do szorstkiego ER przeprowadzają translację białek w celu usunięcia z komórek, złożenia błony oraz utworzenia lizosomów i wakuoli.
Rybosomy można znaleźć pojedynczo w hialoplazmie lub połączyć w grupy podczas jednoczesnej syntezy kilku łańcuchów polipeptydowych na jednym mRNA. Takie grupy rybosomów nazywane są polirybosomy, Lub polisomy.

Mikrotubule- Są to cylindryczne, puste w środku organelle niebłonowe, które przenikają całą cytoplazmę komórki. Ich średnica wynosi około 25 nm, grubość ścianki wynosi 6-8 nm. Tworzą je liczne cząsteczki białka tubulina, które najpierw tworzą 13 nitek przypominających koraliki, a następnie łączą się w mikrotubulę. Mikrotubule tworzą siateczkę cytoplazmatyczną, która nadaje komórce kształt i objętość, łączy błonę komórkową z innymi częściami komórki, zapewnia transport substancji w komórce, bierze udział w ruchu komórki i składników wewnątrzkomórkowych, a także w podział materiału genetycznego. Są częścią centrum komórkowego i organelli ruchu - wici i rzęsek.
mikrofilamenty, Lub mikrowątki, są również organellami niebłonowymi, mają jednak kształt nitkowaty i są utworzone nie przez tubulinę, ale aktyna. Biorą udział w procesach transportu błonowego, rozpoznawania międzykomórkowego, podziału cytoplazmy komórkowej i jej ruchu. W komórkach mięśniowych interakcja mikrofilamentów aktyny z włóknami miozyny pośredniczy w skurczu.
Mikrotubule i mikrofilamenty tworzą wewnętrzny szkielet komórki - cytoszkielet. Jest to złożona sieć włókien, która zapewnia mechaniczne wsparcie błony komórkowej, determinuje kształt komórki, lokalizację organelli komórkowych i ich ruch podczas podziału komórki.

Centrum komórek- organella niebłonowa zlokalizowana w komórkach zwierzęcych w pobliżu jądra; nie ma go w komórkach roślinnych. Jego długość wynosi około 0,2-0,3 mikrona, a średnica 0,1-0,15 mikrona. Centrum komórki tworzą dwa centriole, leżące we wzajemnie prostopadłych płaszczyznach, i promienna kula z mikrotubul. Każda centriola składa się z dziewięciu grup mikrotubul, zebranych w grupy po trzy, tj. Trójki. Centrum komórkowe bierze udział w procesach składania mikrotubul, podziale materiału dziedzicznego komórki, a także w tworzeniu wici i rzęsek.

Organelle ruchu. Wici I rzęsy Są to narośle komórkowe pokryte plazmalemmą. Podstawą tych organelli jest dziewięć par mikrotubul rozmieszczonych wzdłuż obwodu i dwie wolne mikrotubule pośrodku. Mikrotubule są połączone różnymi białkami, zapewniając ich skoordynowane odchylenie od osi - oscylację. Oscylacje są zależne od energii, to znaczy, że na ten proces zużywana jest energia wysokoenergetycznych wiązań ATP. Przywrócenie utraconej wici i rzęsek jest funkcją ciała podstawowe, Lub kinetosomy zlokalizowane u ich podstawy.
Długość rzęsek wynosi około 10-15 nm, a długość wici 20-50 µm. Ze względu na ściśle ukierunkowane ruchy wici i rzęsek następuje nie tylko ruch zwierząt jednokomórkowych, plemników itp., Ale także oczyszczanie dróg oddechowych i ruch jaja przez jajowody, ponieważ wszystkie te części ludzkiego ciała pokryte są nabłonkiem rzęskowym.

Inkluzje
Inkluzje- Są to nietrwałe składniki komórki, które powstają i zanikają w trakcie jej życia. Należą do nich zarówno substancje rezerwowe, na przykład ziarna skrobi lub białka w komórkach roślinnych, granulki glikogenu w komórkach zwierząt i grzybów, wolutyna w bakteriach, krople tłuszczu we wszystkich typach komórek, jak i produkty przemiany materii, w szczególności resztki jedzenia niestrawione w wyniku fagocytozy, tworząc tzw. ciała resztkowe.
Związek między strukturą i funkcjami części i organelli komórki jest podstawą jej integralności
Każda z części komórki z jednej strony stanowi odrębną strukturę o określonej budowie i funkcjach, z drugiej zaś jest składnikiem bardziej złożonego układu zwanego komórką. Większość dziedzicznych informacji komórki eukariotycznej koncentruje się w jądrze, ale samo jądro nie jest w stanie zapewnić jej wdrożenia, ponieważ wymaga to przynajmniej cytoplazmy, która działa jako główna substancja, oraz rybosomów, na których zachodzi ta synteza . Większość rybosomów zlokalizowana jest na ziarnistej siateczce śródplazmatycznej, skąd białka najczęściej transportowane są do kompleksu Golgiego, a następnie po modyfikacji do tych części komórki, dla których są przeznaczone lub wydalane. Błonowe opakowania białek i węglowodanów można wbudować w błony organelli i błonę cytoplazmatyczną, zapewniając ich ciągłą odnowę. Od kompleksu Golgiego oddzielają się także lizosomy i wakuole, które pełnią ważne funkcje. Na przykład bez lizosomów komórki szybko zamieniłyby się w rodzaj wysypiska cząsteczek i struktur odpadowych.
Zajście wszystkich tych procesów wymaga energii wytwarzanej przez mitochondria, a u roślin przez chloroplasty. I chociaż te organelle są stosunkowo autonomiczne, ponieważ mają własne cząsteczki DNA, niektóre z ich białek są nadal kodowane przez genom jądrowy i syntetyzowane w cytoplazmie.
Zatem komórka jest nierozerwalną jednością swoich elementów składowych, z których każdy pełni swoją unikalną funkcję.
Metabolizm i konwersja energii to właściwości organizmów żywych. Metabolizm energetyczny i plastyczny, ich związek. Etapy metabolizmu energetycznego. Fermentacja i oddychanie. Fotosynteza, jej znaczenie, rola kosmiczna. Fazy fotosyntezy. Jasne i ciemne reakcje fotosyntezy, ich związek. Chemosynteza. Rola bakterii chemosyntetycznych na Ziemi
Metabolizm i przemiana energii - właściwości organizmów żywych
Komórkę można porównać do miniaturowej fabryki chemicznej, w której zachodzą setki i tysiące reakcji chemicznych.
Metabolizm- zespół przemian chemicznych mających na celu zachowanie i samoreprodukcję układów biologicznych.
Obejmuje przyjmowanie substancji do organizmu podczas odżywiania i oddychania, metabolizmu wewnątrzkomórkowego lub metabolizm, a także izolację końcowych produktów przemiany materii.
Metabolizm jest nierozerwalnie związany z procesami przemiany jednego rodzaju energii w inny. Przykładowo w procesie fotosyntezy energia świetlna magazynowana jest w postaci energii wiązań chemicznych złożonych cząsteczek organicznych, a w procesie oddychania jest uwalniana i zużywana na syntezę nowych cząsteczek, pracę mechaniczną i osmotyczną, rozpraszane w postaci ciepła itp.
Zachodzenie reakcji chemicznych w organizmach żywych zapewniają katalizatory biologiczne o charakterze białkowym - enzymy, Lub enzymy. Podobnie jak inne katalizatory, enzymy przyspieszają zachodzenie reakcji chemicznych w komórce dziesiątki i setki tysięcy razy, a czasami nawet je umożliwiają, ale nie zmieniają charakteru ani właściwości końcowego produktu(ów) reakcji i nie nie zmieniać siebie. Enzymy mogą być zarówno białkami prostymi, jak i złożonymi, które oprócz części białkowej zawierają także część niebiałkową - kofaktor (koenzym). Przykładami enzymów są amylaza ślinowa, która rozkłada polisacharydy podczas długotrwałego żucia i pepsyna, która zapewnia trawienie białek w żołądku.
Enzymy różnią się od katalizatorów niebiałkowych wysoką specyficznością działania, znacznym wzrostem szybkości reakcji za ich pomocą, a także możliwością regulowania działania poprzez zmianę warunków reakcji lub oddziaływanie z nimi różnych substancji. Ponadto warunki, w których zachodzi kataliza enzymatyczna, znacznie różnią się od tych, w których zachodzi kataliza nieenzymatyczna: optymalna temperatura dla funkcjonowania enzymów w organizmie człowieka wynosi 37°C $, ciśnienie powinno być zbliżone do atmosferycznego, a temperatura $pH$ środowiska może znacznie się wahać. Zatem amylaza wymaga środowiska zasadowego, a pepsyna wymaga środowiska kwaśnego.
Mechanizm działania enzymów polega na zmniejszaniu energii aktywacji substancji (substratów), które wchodzą w reakcję w wyniku tworzenia pośrednich kompleksów enzym-substrat.

Metabolizm energetyczny i plastyczny, ich związek
Metabolizm składa się z dwóch procesów zachodzących jednocześnie w komórce: metabolizmu plastycznego i energetycznego.
Metabolizm plastyczny (anabolizm, asymilacja) to zestaw reakcji syntezy obejmujących wydatek energii ATP. W procesie metabolizmu plastycznego syntetyzowane są substancje organiczne niezbędne komórce. Przykładami reakcji wymiany plastycznej są fotosynteza, biosynteza białek i replikacja DNA (samoduplikacja).
Metabolizm energii (katabolizm, dysymilacja) to zespół reakcji rozkładających złożone substancje na prostsze. W wyniku metabolizmu energetycznego energia jest uwalniana i magazynowana w postaci ATP. Najważniejszymi procesami metabolizmu energetycznego są oddychanie i fermentacja.
Wymiana plastiku i energii jest ze sobą nierozerwalnie związana, ponieważ w procesie wymiany plastycznej powstają substancje organiczne, do czego potrzebna jest energia ATP, a w procesie wymiany energii substancje organiczne ulegają rozkładowi i uwalniana jest energia, która następnie zostanie wykorzystana na procesy syntezy .
Organizmy otrzymują energię w procesie odżywiania, a uwalniają ją i przekształcają w dostępną formę głównie w procesie oddychania. Zgodnie z metodą żywienia wszystkie organizmy dzielą się na autotrofy i heterotrofy. Autotrofy zdolny do samodzielnej syntezy substancji organicznych z nieorganicznych, oraz heterotrofy używaj wyłącznie przygotowanych substancji organicznych.
Etapy metabolizmu energetycznego
Pomimo złożoności reakcji metabolizmu energii, umownie dzieli się go na trzy etapy: przygotowawczy, beztlenowy (beztlenowy) i tlenowy (tlenowy).
NA etap przygotowawczy cząsteczki polisacharydów, lipidów, białek, kwasów nukleinowych rozkładają się na prostsze, na przykład glukozę, glicerynę i kwasy tłuszczowe, aminokwasy, nukleotydy itp. Ten etap może zachodzić bezpośrednio w komórkach lub w jelitach, skąd rozbite Substancje puchowe są dostarczane przez krwioobieg.
Etap beztlenowy metabolizmowi energetycznemu towarzyszy dalszy rozkład monomerów związków organicznych na jeszcze prostsze produkty pośrednie, na przykład kwas pirogronowy lub pirogronian. Nie wymaga obecności tlenu, a dla wielu organizmów żyjących w błocie bagiennym czy w jelitach człowieka jest to jedyny sposób na pozyskanie energii. Beztlenowy etap metabolizmu energetycznego zachodzi w cytoplazmie.
Rozszczepieniu beztlenowemu mogą ulegać różne substancje, ale dość często substratem reakcji jest glukoza. Nazywa się proces jego beztlenowego rozszczepiania glikoliza. Podczas glikolizy cząsteczka glukozy traci cztery atomy wodoru, czyli ulega utlenieniu i powstają dwie cząsteczki kwasu pirogronowego, dwie cząsteczki ATP i dwie cząsteczki zredukowanego nośnika wodoru $NADH + H^(+)$:
$C_6H_(12)O_6 + 2H_3PO_4 + 2ADP + 2NAD → 2C_3H_4O_3 + 2ATP + 2NADH + H^(+) + 2H_2O$.
Tworzenie ATP z ADP następuje w wyniku bezpośredniego przeniesienia anionu fosforanowego z prefosforylowanego cukru i nazywa się to fosforylacja substratu.
Etap aerobowy wymiana energii może zachodzić tylko w obecności tlenu, natomiast związki pośrednie powstające podczas rozszczepienia beztlenowego ulegają utlenieniu do produktów końcowych (dwutlenek węgla i woda) i uwalnia się większość energii zmagazynowanej w wiązaniach chemicznych związków organicznych. Zamienia się w energię wysokoenergetycznych wiązań 36 cząsteczek ATP. Ten etap jest również nazywany oddychanie tkankowe. W przypadku braku tlenu związki pośrednie przekształcają się w inne substancje organiczne w procesie zwanym fermentacja.
Oddech
Mechanizm oddychania komórkowego schematycznie przedstawiono na ryc.

Oddychanie tlenowe zachodzi w mitochondriach, przy czym kwas pirogronowy najpierw traci jeden atom węgla, czemu towarzyszy synteza jednego redukującego równoważnika $NADH + H^(+)$ i cząsteczki acetylokoenzymu A (acetylo-CoA):
$C_3H_4O_3 + NAD + H~CoA → CH_3CO~CoA + NADH + H^(+) + CO_2$.
Acetylo-CoA w macierzy mitochondrialnej bierze udział w łańcuchu reakcji chemicznych, których całość nazywa się cykl Krebsa (cykl kwasu trikarboksylowego, cykl kwasu cytrynowego). Podczas tych przemian powstają dwie cząsteczki ATP, acetylo-CoA ulega całkowitemu utlenieniu do dwutlenku węgla, a jego jony wodorowe i elektrony zostają dodane do nośników wodoru $NADH + H^(+)$ i $FADH_2$. Nośniki transportują protony i elektrony wodoru do wewnętrznych błon mitochondriów, tworząc cristae. Za pomocą białek nośnikowych protony wodoru pompowane są do przestrzeni międzybłonowej, a elektrony przenoszone są przez tzw. łańcuch oddechowy enzymów znajdujących się na wewnętrznej błonie mitochondriów i odprowadzane na atomy tlenu:
$O_2+2e^(-)→O_2^-$.
Należy zauważyć, że niektóre białka łańcucha oddechowego zawierają żelazo i siarkę.
Z przestrzeni międzybłonowej protony wodoru są transportowane z powrotem do macierzy mitochondrialnej za pomocą specjalnych enzymów - syntaz ATP, a uwolniona w tym przypadku energia jest zużywana na syntezę 34 cząsteczek ATP z każdej cząsteczki glukozy. Proces ten nazywa się fosforylacja oksydacyjna. W macierzy mitochondrialnej protony wodoru reagują z rodnikami tlenowymi, tworząc wodę:
$4H^(+)+O_2^-→2H_2O$.
Zbiór reakcji oddychania tlenowego można wyrazić w następujący sposób:
$2C_3H_4O_3 + 6O_2 + 36H_3PO_4 + 36ADP → 6CO_2 + 38H_2O + 36ATP.$
Ogólne równanie oddychania wygląda następująco:
$C_6H_(12)O_6 + 6O_2 + 38H_3PO_4 + 38ADP → 6CO_2 + 40H_2O + 38ATP.$
Fermentacja
W przypadku braku tlenu lub jego niedoboru następuje fermentacja. Fermentacja jest ewolucyjnie wcześniejszą metodą pozyskiwania energii niż oddychanie, jest jednak mniej korzystna energetycznie, ponieważ podczas fermentacji powstają substancje organiczne, które wciąż są bogate w energię. Istnieje kilka głównych rodzajów fermentacji: kwas mlekowy, alkoholowy, kwas octowy itp. Zatem w mięśniach szkieletowych przy braku tlenu podczas fermentacji kwas pirogronowy jest redukowany do kwasu mlekowego, podczas gdy wcześniej utworzone równoważniki redukujące są zużywane i tylko pozostają dwie cząsteczki ATP:
$2C_3H_4O_3 + 2NADH + H^(+) → 2C_3H_6O_3 + 2NAD$.
Podczas fermentacji za pomocą drożdży kwas pirogronowy w obecności tlenu przekształca się w alkohol etylowy i tlenek węgla (IV):
$C_3H_4O_3 + NADH + H^(+) → C_2H_5OH + CO_2 + NAD^(+)$.
Podczas fermentacji za pomocą mikroorganizmów z kwasu pirogronowego można również tworzyć kwas octowy, masłowy, mrówkowy itp.
ATP powstający w wyniku metabolizmu energetycznego zużywany jest w komórce do różnego rodzaju prac: chemicznych, osmotycznych, elektrycznych, mechanicznych i regulacyjnych. Praca chemiczna obejmuje biosyntezę białek, lipidów, węglowodanów, kwasów nukleinowych i innych niezbędnych związków. Praca osmotyczna obejmuje procesy wchłaniania przez komórkę i usuwania z niej substancji znajdujących się w przestrzeni zewnątrzkomórkowej w stężeniach większych niż w samej komórce. Praca elektryczna jest ściśle powiązana z pracą osmotyczną, ponieważ w wyniku ruchu naładowanych cząstek przez membrany powstaje ładunek membranowy i nabywane są właściwości pobudliwości i przewodności. Praca mechaniczna polega na ruchu substancji i struktur wewnątrz komórki, a także komórki jako całości. Praca regulacyjna obejmuje wszystkie procesy mające na celu koordynację procesów zachodzących w komórce.
Fotosynteza, jej znaczenie, rola kosmiczna
Fotosynteza to proces zamiany energii świetlnej na energię wiązań chemicznych związków organicznych przy udziale chlorofilu.
W wyniku fotosyntezy rocznie powstaje około 150 miliardów ton materii organicznej i około 200 miliardów ton tlenu. Proces ten zapewnia obieg węgla w biosferze, zapobiegając gromadzeniu się dwutlenku węgla, a tym samym zapobiegając efektowi cieplarnianemu i przegrzaniu Ziemi. Substancje organiczne powstałe w wyniku fotosyntezy nie są w całości zużywane przez inne organizmy, znaczna ich część na przestrzeni milionów lat utworzyła złoża minerałów (węgiel kamienny i brunatny, ropa naftowa). Od niedawna jako paliwo zaczęto stosować także olej rzepakowy („biodiesel”) i alkohol otrzymywany z resztek roślinnych. Ozon powstaje z tlenu pod wpływem wyładowań elektrycznych, tworząc ekran ozonowy chroniący całe życie na Ziemi przed niszczycielskim działaniem promieni ultrafioletowych.
Nasz rodak, wybitny fizjolog roślin K. A. Timiryazev (1843-1920), nazwał rolę fotosyntezy „kosmiczną”, ponieważ łączy ona Ziemię ze Słońcem (przestrzenią), zapewniając dopływ energii do planety.
Fazy fotosyntezy. Jasne i ciemne reakcje fotosyntezy, ich związek
W 1905 roku angielski fizjolog roślin F. Blackman odkrył, że tempo fotosyntezy nie może rosnąć w nieskończoność, pewne czynniki je ograniczają. Na tej podstawie postawił hipotezę, że istnieją dwie fazy fotosyntezy: światło I ciemny. Przy małym natężeniu światła szybkość reakcji świetlnych wzrasta proporcjonalnie do wzrostu natężenia światła, a ponadto reakcje te nie zależą od temperatury, gdyż nie wymagają zajścia enzymów. Reakcje świetlne zachodzą na błonach tylakoidów.
Natomiast szybkość reakcji ciemnych wzrasta wraz ze wzrostem temperatury, jednak po osiągnięciu progu temperatury 30°C wzrost ten ustaje, co wskazuje na enzymatyczny charakter tych przemian zachodzących w zrębie. Należy zauważyć, że światło ma również pewien wpływ na reakcje ciemne, mimo że nazywa się je reakcjami ciemnymi.

Lekka faza fotosyntezy zachodzi na błonach tylakoidów zawierających kilka rodzajów kompleksów białkowych, z których głównymi są fotosystemy I i II oraz syntaza ATP. Do fotosystemów zaliczają się kompleksy pigmentowe, które oprócz chlorofilu zawierają także karotenoidy. Karotenoidy wychwytują światło w obszarach widma, w których nie robi tego chlorofil, a także chronią chlorofil przed zniszczeniem przez światło o dużym natężeniu.
Oprócz kompleksów pigmentowych fotosystemy obejmują także szereg białek akceptorowych elektronów, które sekwencyjnie przenoszą między sobą elektrony z cząsteczek chlorofilu. Sekwencja tych białek nazywa się łańcuch transportu elektronów w chloroplastach.
Z fotosystemem II związany jest także specjalny kompleks białek, który zapewnia uwalnianie tlenu podczas fotosyntezy. Ten kompleks uwalniający tlen zawiera jony manganu i chloru.
W faza jasna Kwanty świetlne, czyli fotony, padając na cząsteczki chlorofilu znajdujące się na błonach tylakoidów, wprowadzają je w stan wzbudzony, charakteryzujący się wyższą energią elektronów. W tym przypadku wzbudzone elektrony z chlorofilu fotosystemu I przenoszone są poprzez łańcuch pośredników na nośnik wodoru NADP, który przyłącza protony wodoru, zawsze obecny w roztworze wodnym:
$NADP + 2e^(-) + 2H^(+) → NADPH + H^(+)$.
Zredukowany $NADPH + H^(+)$ zostanie następnie użyty w fazie ciemnej. Elektrony z chlorofilu fotosystemu II są również przenoszone wzdłuż łańcucha transportu elektronów, ale wypełniają „dziury elektronowe” chlorofilu fotosystemu I. Braki elektronowe w chlorofilu fotosystemu II uzupełniane są poprzez usuwanie cząsteczek wody, które zachodzi przy udziale wspomnianego już kompleksu uwalniającego tlen. W wyniku rozkładu cząsteczek wody, tzw fotoliza powstają protony wodoru i uwalnia się tlen cząsteczkowy, będący produktem ubocznym fotosyntezy:
$H_2O → 2H^(+) + 2e^(-) + (1)/(2)O_2$.
Informacja genetyczna w komórce. Geny, kod genetyczny i jego właściwości. Macierzowy charakter reakcji biosyntezy. Biosynteza białek i kwasów nukleinowych
Informacja genetyczna w komórce
Rozmnażanie własnego rodzaju jest jedną z podstawowych właściwości istot żywych. Dzięki temu zjawisku istnieje podobieństwo nie tylko między organizmami, ale także między poszczególnymi komórkami, a także ich organellami (mitochondriami i plastydami). Materialną podstawą tego podobieństwa jest transfer informacji genetycznej zakodowanej w sekwencji nukleotydowej DNA, który odbywa się poprzez procesy replikacji DNA (samoduplikacji). Wszystkie cechy i właściwości komórek i organizmów realizowane są dzięki białkom, których strukturę określa przede wszystkim sekwencja nukleotydów DNA. Dlatego też biosynteza kwasów nukleinowych i białek odgrywa ogromne znaczenie w procesach metabolicznych. Jednostką strukturalną informacji dziedzicznej jest gen.
Geny, kod genetyczny i jego właściwości
Informacja dziedziczna w komórce nie jest monolityczna, jest podzielona na osobne „słowa” - geny.
Gen jest elementarną jednostką informacji genetycznej.
Prace nad programem „Human Genome”, prowadzonym jednocześnie w kilku krajach i zakończonym na początku tego stulecia, dały nam zrozumienie, że człowiek ma tylko około 25-30 tysięcy genów, ale informacja z większości naszego DNA nigdy nie jest czytany, ponieważ zawiera ogromną liczbę nic nie znaczących fragmentów, powtórzeń i genów kodujących cechy, które utraciły znaczenie dla człowieka (ogon, owłosienie na ciele itp.). Ponadto odszyfrowano szereg genów odpowiedzialnych za rozwój chorób dziedzicznych, a także geny docelowe leków. Praktyczne zastosowanie wyników uzyskanych w trakcie realizacji tego programu odłożono jednak do czasu rozszyfrowania genomów większej liczby osób i wyjaśnienia, czym się różnią.
Nazywa się geny kodujące pierwotną strukturę białka, rybosomu lub transferowego RNA strukturalny oraz geny zapewniające aktywację lub tłumienie odczytywania informacji z genów strukturalnych - regulacyjne. Jednak nawet geny strukturalne zawierają regiony regulatorowe.
Dziedziczna informacja organizmów jest szyfrowana w DNA w postaci pewnych kombinacji nukleotydów i ich sekwencji - kod genetyczny. Jego właściwości to: potrójność, specyficzność, uniwersalność, redundancja i brak nakładania się. Ponadto w kodzie genetycznym nie ma znaków interpunkcyjnych.
Każdy aminokwas jest kodowany w DNA przez trzy nukleotydy - tryplet, na przykład metionina jest kodowana przez triplet TAC, to znaczy kod jest tripletowy. Natomiast każda trójka koduje tylko jeden aminokwas, co stanowi o jej specyficzności, czyli jednoznaczności. Kod genetyczny jest uniwersalny dla wszystkich żywych organizmów, co oznacza, że dziedziczna informacja o ludzkich białkach może zostać odczytana przez bakterie i odwrotnie. Wskazuje to na jedność pochodzenia świata organicznego. Jednak 64 kombinacje trzech nukleotydów odpowiadają tylko 20 aminokwasom, w wyniku czego jeden aminokwas może być kodowany przez 2-6 tripletów, czyli kod genetyczny jest zbędny lub zdegenerowany. Trzy trojaczki nie mają odpowiednich aminokwasów, tak się je nazywa kodony stop, gdyż wskazują na koniec syntezy łańcucha polipeptydowego.
Sekwencja zasad w tripletach DNA i kodowane przez nie aminokwasy
*Kodon stop, wskazujący koniec syntezy łańcucha polipeptydowego.
Skróty nazw aminokwasów:
Ala - alanina
Arg – arginina
Asn – asparagina
Asp - kwas asparaginowy
Val - walina
Jego - histydyna
Gly - glicyna
Gln - glutamina
Glu - kwas glutaminowy
Ile - izoleucyna
Leu - leucyna
Liz - lizyna
Met - metionina
Pro - prolina
Ser - seryna
Tyr - tyrozyna
Tre - treonina
Trzy - tryptofan
Fen – fenyloalanina
Cis - cysteina
Jeśli zaczniemy czytać informację genetyczną nie od pierwszego nukleotydu w triplecie, ale od drugiego, to nie tylko przesunie się ramka odczytu, ale tak zsyntetyzowane białko będzie zupełnie inne nie tylko pod względem sekwencji nukleotydów, ale także strukturę i właściwości. Pomiędzy trójkami nie ma znaków interpunkcyjnych, zatem nie ma przeszkód w przesunięciu ramki odczytu, co otwiera przestrzeń dla występowania i utrzymywania się mutacji.
Macierzowy charakter reakcji biosyntezy
Komórki bakteryjne potrafią podwajać się co 20-30 minut, a komórki eukariotyczne - codziennie, a nawet częściej, co wymaga dużej szybkości i dokładności replikacji DNA. Ponadto każda komórka zawiera setki i tysiące kopii wielu białek, zwłaszcza enzymów, dlatego „fragmentarna” metoda ich wytwarzania jest niedopuszczalna dla ich reprodukcji. Bardziej postępową metodą jest stemplowanie, które pozwala uzyskać wiele dokładnych kopii produktu, a także obniżyć jego koszt. Do stemplowania wymagana jest matryca, z której wykonany jest wycisk.
W komórkach zasada syntezy matryc polega na tym, że nowe cząsteczki białek i kwasów nukleinowych syntetyzowane są zgodnie z programem osadzonym w strukturze istniejących wcześniej cząsteczek tych samych kwasów nukleinowych (DNA lub RNA).
Biosynteza białek i kwasów nukleinowych
Replikacja DNA. DNA jest dwuniciowym biopolimerem, którego monomerami są nukleotydy. Gdyby biosynteza DNA przebiegała na zasadzie fotokopii, nieuchronnie powstałyby liczne zniekształcenia i błędy w informacjach dziedzicznych, co ostatecznie doprowadziłoby do śmierci nowych organizmów. Dlatego proces podwajania DNA przebiega inaczej, w sposób półkonserwatywny: cząsteczka DNA rozwija się i na każdym z łańcuchów syntetyzowany jest nowy łańcuch zgodnie z zasadą komplementarności. Nazywa się proces samoreprodukcji cząsteczki DNA, zapewniający dokładne kopiowanie informacji dziedzicznej i jej przekazywanie z pokolenia na pokolenie replikacja(od łac. replikacja- powtórzenie). W wyniku replikacji powstają dwie absolutnie dokładne kopie cząsteczki macierzystego DNA, z których każda niesie jedną kopię cząsteczki macierzystego DNA.

Proces replikacji jest w rzeczywistości niezwykle złożony, ponieważ bierze w nim udział wiele białek. Niektóre z nich rozwijają podwójną helisę DNA, inne rozrywają wiązania wodorowe pomiędzy nukleotydami komplementarnych łańcuchów, jeszcze inne (np. enzym polimeraza DNA) selekcjonują nowe nukleotydy w oparciu o zasadę komplementarności itp. Dwie cząsteczki DNA tworzą się jako wynik replikacji rozdziela się na dwie części podczas podziału nowo utworzone komórki potomne.
Błędy w procesie replikacji zdarzają się niezwykle rzadko, jeśli jednak wystąpią, są bardzo szybko eliminowane zarówno przez polimerazy DNA, jak i specjalne enzymy naprawcze, gdyż każdy błąd w sekwencji nukleotydów może prowadzić do nieodwracalnej zmiany w strukturze i funkcjach białka i ostatecznie niekorzystnie wpływają na żywotność nowej komórki lub nawet osobnika.
Biosynteza białek. Jak ujął to w przenośni wybitny filozof XIX wieku F. Engels: „Życie jest formą istnienia ciał białkowych”. O strukturze i właściwościach cząsteczek białek decyduje ich struktura pierwotna, czyli sekwencja aminokwasów zakodowanych w DNA. Od dokładności odwzorowania tej informacji zależy nie tylko istnienie samego polipeptydu, ale także funkcjonowanie komórki jako całości, dlatego ogromne znaczenie ma proces syntezy białek. Wydaje się, że jest to najbardziej złożony proces syntezy w komórce, gdyż bierze w nim udział aż trzysta różnych enzymów i innych makrocząsteczek. Poza tym płynie z dużą prędkością, co wymaga jeszcze większej precyzji.
Istnieją dwa główne etapy biosyntezy białek: transkrypcja i translacja.
Transkrypcja(od łac. transkrypcja- przepisywanie) to biosynteza cząsteczek mRNA na matrycy DNA.

Ponieważ cząsteczka DNA zawiera dwa antyrównoległe łańcuchy, odczytanie informacji z obu łańcuchów doprowadziłoby do powstania zupełnie różnych mRNA, dlatego ich biosynteza jest możliwa tylko na jednym z łańcuchów, co nazywa się kodowaniem, czyli kodogennym, w przeciwieństwie do drugiego, niekodujące lub niekodogenne. Za proces przepisywania odpowiada specjalny enzym, polimeraza RNA, który selekcjonuje nukleotydy RNA zgodnie z zasadą komplementarności. Proces ten może zachodzić zarówno w jądrze, jak i w organellach posiadających własne DNA - mitochondriach i plastydach.
Cząsteczki mRNA syntetyzowane podczas transkrypcji podlegają złożonemu procesowi przygotowania do translacji (mitochondrialne i plastydowe mRNA mogą pozostać wewnątrz organelli, gdzie następuje drugi etap biosyntezy białek). W procesie dojrzewania mRNA przyłączają się do niego pierwsze trzy nukleotydy (AUG) oraz ogon nukleotydów adenylowych, których długość decyduje o tym, ile kopii białka można zsyntetyzować na danej cząsteczce. Dopiero wtedy dojrzałe mRNA opuszczają jądro przez pory jądrowe.
Równolegle w cytoplazmie zachodzi proces aktywacji aminokwasów, podczas którego aminokwas łączy się z odpowiednim wolnym tRNA. Proces ten jest katalizowany przez specjalny enzym i wymaga ATP.
Audycja(od łac. audycja- transfer) to biosynteza łańcucha polipeptydowego na matrycy mRNA, podczas której informacja genetyczna ulega translacji na sekwencję aminokwasów łańcucha polipeptydowego.

Drugi etap syntezy białek najczęściej zachodzi w cytoplazmie, na przykład na szorstkim ER. Do jego wystąpienia konieczna jest obecność rybosomów, aktywacja tRNA, podczas której przyłączają one odpowiednie aminokwasy, obecność jonów Mg2+, a także optymalne warunki środowiskowe (temperatura, pH, ciśnienie itp.).
Aby rozpocząć nadawanie ( inicjacja) do gotowej do syntezy cząsteczki mRNA przyłącza się małą podjednostkę rybosomu, po czym zgodnie z zasadą komplementarności do pierwszego kodonu (AUG) wybiera się tRNA niosący aminokwas metioninę. Dopiero potem przyłącza się duża podjednostka rybosomu. W złożonym rybosomie znajdują się dwa kodony mRNA, z których pierwszy jest już zajęty. Do sąsiadującego z nim kodonu dodaje się drugi tRNA, również niosący aminokwas, po czym za pomocą enzymów tworzy się wiązanie peptydowe pomiędzy resztami aminokwasów. Rybosom przesuwa jeden kodon mRNA; pierwszy tRNA uwolniony od aminokwasu powraca do cytoplazmy po następnym aminokwasie, a fragment przyszłego łańcucha polipeptydowego wisi jakby na pozostałym tRNA. Do nowego kodonu znajdującego się w obrębie rybosomu przyłączany jest kolejny tRNA, proces się powtarza i krok po kroku łańcuch polipeptydowy wydłuża się, tj. wydłużenie.
Koniec syntezy białek ( zakończenie) następuje w momencie napotkania w cząsteczce mRNA określonej sekwencji nukleotydowej, która nie koduje aminokwasu (kodon stop). Następnie następuje rozdział rybosomu, mRNA i łańcucha polipeptydowego, a nowo zsyntetyzowane białko uzyskuje odpowiednią strukturę i jest transportowane do części komórki, gdzie będzie pełnić swoje funkcje.
Translacja jest procesem bardzo energochłonnym, ponieważ energia jednej cząsteczki ATP jest zużywana na przyłączenie jednego aminokwasu do tRNA, a kilka innych jest zużywanych do przemieszczania rybosomu wzdłuż cząsteczki mRNA.
Aby przyspieszyć syntezę niektórych cząsteczek białka, do cząsteczki mRNA można kolejno dołączyć kilka rybosomów, które tworzą pojedynczą strukturę - polisom.
Komórka jest jednostką genetyczną żywej istoty. Chromosomy, ich budowa (kształt i wielkość) oraz funkcje. Liczba chromosomów i ich stałość gatunkowa. Komórki somatyczne i rozrodcze. Cykl życia komórki: interfaza i mitoza. Mitoza to podział komórek somatycznych. Mejoza. Fazy mitozy i mejozy. Rozwój komórek rozrodczych u roślin i zwierząt. Podział komórek jest podstawą wzrostu, rozwoju i rozmnażania organizmów. Rola mejozy i mitozy
Komórka jest jednostką genetyczną żywej istoty.
Pomimo tego, że kwasy nukleinowe są nośnikiem informacji genetycznej, realizacja tej informacji poza komórką jest niemożliwa, co łatwo udowodnić na przykładzie wirusów. Organizmy te, często zawierające jedynie DNA lub RNA, nie mogą rozmnażać się samodzielnie, w tym celu muszą korzystać z dziedzicznego aparatu komórkowego. Nie mogą nawet przeniknąć do komórki bez pomocy samej komórki, chyba że poprzez zastosowanie mechanizmów transportu przez błonę lub uszkodzenie komórki. Większość wirusów jest niestabilna i ginie już po kilku godzinach wystawienia na działanie otwartego powietrza. W związku z tym komórka jest jednostką genetyczną żywej istoty, która ma minimalny zestaw składników do przechowywania, zmiany i wdrażania informacji dziedzicznej, a także jej przekazywania potomkom.
Większość informacji genetycznej komórki eukariotycznej zlokalizowana jest w jądrze. Osobliwością jego organizacji jest to, że w przeciwieństwie do DNA komórki prokariotycznej, cząsteczki DNA eukariontów nie są zamknięte i tworzą złożone kompleksy z białkami - chromosomami.
Chromosomy, ich budowa (kształt i wielkość) oraz funkcje
Chromosom(z greckiego chrom- kolor, kolorystyka i soma- ciało) to struktura jądra komórkowego, która zawiera geny i niesie pewne dziedziczne informacje o cechach i właściwościach organizmu.
Czasami okrągłe cząsteczki DNA prokariotów nazywane są również chromosomami. Chromosomy są zdolne do samopowielania, mają strukturalną i funkcjonalną indywidualność i zachowują ją przez pokolenia. Każda komórka przenosi wszystkie dziedziczne informacje o ciele, ale tylko niewielka część w niej działa.
Podstawą chromosomu jest dwuniciowa cząsteczka DNA wypełniona białkami. U eukariontów białka histonowe i niehistonowe oddziałują z DNA, podczas gdy u prokariotów białka histonowe są nieobecne.
Chromosomy najlepiej widać pod mikroskopem świetlnym podczas podziału komórki, kiedy w wyniku zagęszczenia przyjmują wygląd ciał w kształcie pręcików oddzielonych pierwotnym przewężeniem - centromer — na ramionach. Na chromosomie może również występować zwężenie wtórne, co w niektórych przypadkach oddziela tzw satelita. Końce chromosomów nazywane są telomery. Telomery zapobiegają sklejaniu się końców chromosomów i zapewniają ich połączenie z błoną jądrową w niedzielącej się komórce. Na początku podziału chromosomy są podwojone i składają się z dwóch chromosomów potomnych - chromatyda, zapinany na centromerze.

Ze względu na kształt chromosomy dzielą się na chromosomy o równych ramionach, o nierównych ramionach i o kształcie pręta. Rozmiary chromosomów znacznie się różnią, ale przeciętny chromosom ma wymiary 5 $×$ 1,4 mikrona.
W niektórych przypadkach chromosomy w wyniku licznych duplikacji DNA zawierają setki i tysiące chromatyd: takie gigantyczne chromosomy nazywane są polietylen. Występują w gruczołach ślinowych larw Drosophila, a także w gruczołach trawiennych glisty.
Liczba chromosomów i ich stałość gatunkowa. Komórki somatyczne i rozrodcze
Według teorii komórkowej komórka jest jednostką struktury, aktywności życiowej i rozwoju organizmu. Zatem tak ważne funkcje żywych istot, jak wzrost, rozmnażanie i rozwój organizmu, są zapewniane na poziomie komórkowym. Komórki organizmów wielokomórkowych można podzielić na komórki somatyczne i rozrodcze.
Komórki somatyczne- są to wszystkie komórki organizmu powstałe w wyniku podziału mitotycznego.
Badanie chromosomów pozwoliło ustalić, że komórki somatyczne organizmu każdego gatunku biologicznego charakteryzują się stałą liczbą chromosomów. Na przykład dana osoba ma ich 46. Nazywa się zestawem chromosomów komórek somatycznych diploidalny(2n) lub podwójne.

Komórki płciowe, Lub gamety, to wyspecjalizowane komórki wykorzystywane do rozmnażania płciowego.
Gamety zawsze zawierają o połowę mniej chromosomów niż komórki somatyczne (u człowieka - 23), dlatego nazywa się zestaw chromosomów komórek rozrodczych haploidalny(n) lub pojedynczy. Jego powstawanie jest związane z podziałem komórek mejotycznych.
Ilość DNA w komórkach somatycznych oznaczono jako 2c, a w komórkach płciowych - 1c. Wzór genetyczny komórek somatycznych zapisany jest jako 2n2c, a komórek płciowych - 1n1c.
W jądrach niektórych komórek somatycznych liczba chromosomów może różnić się od ich liczby w komórkach somatycznych. Jeśli ta różnica jest większa niż jeden, dwa, trzy itd. zbiory haploidalne, wówczas takie komórki nazywa się poliploidalny(odpowiednio tri-, tetra-, pentaploidalne). W takich komórkach procesy metaboliczne przebiegają zwykle bardzo intensywnie.
Liczba chromosomów sama w sobie nie jest cechą gatunkową, ponieważ różne organizmy mogą mieć tę samą liczbę chromosomów, ale spokrewnione mogą mieć inną liczbę. Na przykład plazmodium malarii i glista końska mają po dwa chromosomy, podczas gdy ludzie i szympansy mają odpowiednio 46 i 48.
Ludzkie chromosomy dzielą się na dwie grupy: autosomy i chromosomy płciowe (heterochromosomy). Autosom w ludzkich komórkach somatycznych są 22 pary, są one takie same dla mężczyzn i kobiet oraz chromosomy płciowe tylko jedna para, ale to ona określa płeć jednostki. Istnieją dwa rodzaje chromosomów płciowych – X i Y. Komórki ciała kobiet noszą dwa chromosomy X, a mężczyzn – X i Y.
Kariotyp- jest to zespół cech zestawu chromosomów organizmu (liczba chromosomów, ich kształt i wielkość).
Warunkowy zapis kariotypu obejmuje całkowitą liczbę chromosomów, chromosomów płciowych i możliwe odchylenia w zestawie chromosomów. Na przykład kariotyp normalnego mężczyzny zapisywany jest jako 46, XY, a kariotyp normalnej kobiety to 46, XX.
Cykl życia komórki: interfaza i mitoza
Komórki nie powstają za każdym razem na nowo, powstają jedynie w wyniku podziału komórek macierzystych. Komórki potomne po podziale potrzebują czasu, aby uformować organelle i uzyskać odpowiednią strukturę zapewniającą pełnienie określonej funkcji. Ten okres czasu nazywa się dojrzewanie.
Okres od pojawienia się komórki w wyniku podziału do jej podziału lub śmierci nazywa się cykl życiowy komórki.
W komórkach eukariotycznych cykl życiowy dzieli się na dwa główne etapy: interfazę i mitozę.
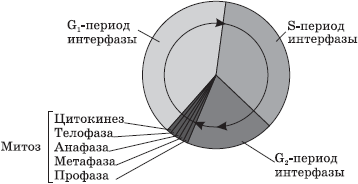
Interfaza- jest to okres w cyklu życiowym, podczas którego komórka nie dzieli się i funkcjonuje normalnie. Interfaza jest podzielona na trzy okresy: okresy G 1, S i G 2.
G 1 -okres(presyntetyczny, postmitotyczny) to okres wzrostu i rozwoju komórki, podczas którego następuje aktywna synteza RNA, białek i innych substancji niezbędnych do pełnego podtrzymania życia nowo powstałej komórki. Pod koniec tego okresu komórka może zacząć przygotowywać się do duplikacji swojego DNA.
W Okres S(syntetyczny) zachodzi sam proces replikacji DNA. Jedyną częścią chromosomu, która nie ulega replikacji, jest centromer, więc powstałe cząsteczki DNA nie rozchodzą się całkowicie, ale pozostają w nim połączone, a na początku podziału chromosom ma wygląd w kształcie litery X. Wzór genetyczny komórki po podwojeniu DNA to 2n4c. Również w okresie S centriole centrum komórki ulegają podwojeniu.
G 2 -okres(postsyntetyczny, premitotyczny) charakteryzuje się intensywną syntezą RNA, białek i ATP niezbędnych w procesie podziału komórki, a także separacji centrioli, mitochondriów i plastydów. Do końca interfazy chromatyna i jąderko pozostają wyraźnie rozróżnialne, integralność otoczki jądrowej nie zostaje naruszona, a organelle nie ulegają zmianie.
Niektóre komórki organizmu są w stanie pełnić swoje funkcje przez całe życie organizmu (neurony naszego mózgu, komórki mięśniowe serca), inne natomiast istnieją przez krótki czas, po czym obumierają (komórki nabłonka jelit, komórki naskórka Skóra). W związku z tym organizm musi stale podlegać procesom podziału komórek i powstawania nowych, które zastąpią te martwe. Nazywa się komórki zdolne do podziału trzon. W organizmie człowieka znajdują się w czerwonym szpiku kostnym, w głębokich warstwach naskórka skóry i innych miejscach. Za pomocą tych komórek możesz wyhodować nowy narząd, osiągnąć odmłodzenie, a także sklonować ciało. Perspektywy wykorzystania komórek macierzystych są całkowicie jasne, jednak moralne i etyczne aspekty tego problemu są nadal przedmiotem dyskusji, gdyż w większości przypadków wykorzystuje się embrionalne komórki macierzyste uzyskane z ludzkich embrionów zabitych podczas aborcji.
Czas trwania interfazy w komórkach roślinnych i zwierzęcych wynosi średnio 10-20 godzin, natomiast mitoza trwa około 1-2 godzin.
Podczas kolejnych podziałów organizmów wielokomórkowych komórki potomne stają się coraz bardziej zróżnicowane w miarę odczytywania informacji z coraz większej liczby genów.
Niektóre komórki z czasem przestają się dzielić i obumierają, co może wynikać z dopełnienia określonych funkcji, jak ma to miejsce w przypadku komórek naskórka i komórek krwi, lub z powodu uszkodzenia tych komórek przez czynniki środowiskowe, w szczególności patogeny. Genetycznie zaprogramowana śmierć komórki nazywa się apoptoza, zaś przypadkowa śmierć - martwica.
Mitoza to podział komórek somatycznych. Fazy mitozy
Mitoza- metoda pośredniego podziału komórek somatycznych.
Podczas mitozy komórka przechodzi szereg kolejnych faz, w wyniku których każda komórka potomna otrzymuje taki sam zestaw chromosomów jak komórka macierzysta.
Mitoza dzieli się na cztery główne fazy: profazę, metafazę, anafazę i telofazę. Profaza- najdłuższy etap mitozy, podczas którego chromatyna ulega kondensacji, w wyniku czego widoczne stają się chromosomy w kształcie litery X składające się z dwóch chromatyd (chromosomy potomne). W tym przypadku jąderko zanika, centriole rozchodzą się w stronę biegunów komórki i zaczyna tworzyć się wrzeciono achromatyny (wrzeciono podziału) z mikrotubul. Pod koniec profazy błona jądrowa rozpada się na oddzielne pęcherzyki.

W metafaza Chromosomy ułożone są wzdłuż równika komórki wraz z centromerami, do których przyczepione są mikrotubule w pełni uformowanego wrzeciona. Na tym etapie podziału chromosomy są najbardziej zwarte i mają charakterystyczny kształt, co umożliwia badanie kariotypu.
W anafaza W centromerach zachodzi szybka replikacja DNA, w wyniku czego chromosomy ulegają rozszczepieniu, a chromatydy rozchodzą się w stronę biegunów komórki, rozciągnięte przez mikrotubule. Rozkład chromatyd musi być absolutnie równy, ponieważ to właśnie ten proces zapewnia utrzymanie stałej liczby chromosomów w komórkach organizmu.
Na scenie telofazy Chromosomy potomne gromadzą się na biegunach, despirują, wokół nich tworzą się błony jądrowe z pęcherzyków, a w nowo powstałych jądrach pojawiają się jąderka.
Po podziale jądrowym następuje podział cytoplazmatyczny - cytokineza, podczas którego następuje mniej więcej równomierne rozmieszczenie wszystkich organelli komórki macierzystej.
Zatem w wyniku mitozy z jednej komórki macierzystej powstają dwie komórki potomne, z których każda jest genetyczną kopią komórki macierzystej (2n2c).
W chorych, uszkodzonych, starzejących się komórkach i wyspecjalizowanych tkankach organizmu może zachodzić nieco inny proces podziału – amitoza. Amitoza zwany bezpośrednim podziałem komórek eukariotycznych, w którym nie dochodzi do tworzenia genetycznie równoważnych komórek, ponieważ składniki komórkowe są rozmieszczone nierównomiernie. Występuje w roślinach w bielmie, au zwierząt - w wątrobie, chrząstce i rogówce oka.
Mejoza. Fazy mejozy
Mejoza to metoda pośredniego podziału pierwotnych komórek rozrodczych (2n2c), w wyniku której powstają komórki haploidalne (1n1c), najczęściej komórki rozrodcze.
W przeciwieństwie do mitozy, mejoza składa się z dwóch kolejnych podziałów komórkowych, z których każdy jest poprzedzony interfazą. Nazywa się pierwszy podział mejozy (mejoza I). redukcjonista, ponieważ w tym przypadku liczba chromosomów jest zmniejszona o połowę, a drugi podział (mejoza II) - równa, ponieważ w jego procesie zostaje zachowana liczba chromosomów.

Interfaza I przebiega jak interfaza mitozy. Mejoza I dzieli się na cztery fazy: profazę I, metafazę I, anafazę I i telofazę I. B profaza I Zachodzą dwa ważne procesy: koniugacja i krzyżowanie. Koniugacja- Jest to proces fuzji homologicznych (sparowanych) chromosomów na całej długości. Pary chromosomów powstałe podczas koniugacji zachowują się do końca metafazy I.
Przechodzić przez- wzajemna wymiana homologicznych regionów homologicznych chromosomów. W wyniku krzyżowania chromosomy otrzymane przez organizm od obojga rodziców nabywają nowe kombinacje genów, co powoduje pojawienie się potomstwa zróżnicowanego genetycznie. Pod koniec profazy I, podobnie jak w profazie mitozy, jąderko zanika, centriole rozchodzą się w stronę biegunów komórki, a błona jądrowa ulega rozpadowi.

W metafaza I pary chromosomów są ułożone wzdłuż równika komórki, a mikrotubule wrzeciona są przyczepione do ich centromerów.
W anafaza I Całe chromosomy homologiczne, składające się z dwóch chromatyd, rozchodzą się w stronę biegunów.
W telofaza I Błony jądrowe tworzą się wokół skupisk chromosomów na biegunach komórki i tworzą się jąderka.
Cytokineza I zapewnia separację cytoplazmy komórek potomnych.
Komórki potomne (1n2c) powstałe w wyniku mejozy I są genetycznie niejednorodne, gdyż ich chromosomy, losowo rozproszone na biegunach komórek, zawierają różne geny.
Charakterystyka porównawcza mitozy i mejozy
| Podpisać | Mitoza | Mejoza | |
| Które komórki zaczynają się dzielić? | Somatyczny (2n) | Pierwotne komórki rozrodcze (2n) | |
| Liczba podziałów | 1 | 2 | |
| Ile i jakiego rodzaju komórek powstaje podczas podziału? | 2 somatyczne (2n) | 4 seksualny (n) | |
| Interfaza | Przygotowanie komórki do podziału, podwojenie DNA | Bardzo krótko, podwojenie DNA nie występuje | |
| Fazy | Mejoza I | Mejoza II | |
| Profaza | Może nastąpić kondensacja chromosomów, zanik jąderka, rozpad błony jądrowej, koniugacja i krzyżowanie | Kondensacja chromosomów, zanik jąderka, rozpad błony jądrowej | |
| Metafaza | Pary chromosomów znajdują się wzdłuż równika, powstaje wrzeciono | Chromosomy ustawiają się wzdłuż równika, powstaje wrzeciono | |
| Anafaza | Chromosomy homologiczne z dwóch chromatyd przemieszczają się w kierunku biegunów | Chromatydy przesuwają się w stronę biegunów | |
| Telofaza | Tworzą się chromosomy despiralne, powstają nowe błony jądrowe i jąderka | Tworzą się chromosomy despiralne, powstają nowe błony jądrowe i jąderka | |
Interfaza II bardzo krótki, ponieważ nie zachodzi w nim podwojenie DNA, to znaczy nie ma okresu S.
Mejoza II dzieli się również na cztery fazy: profazę II, metafazę II, anafazę II i telofazę II. W profaza II zachodzą te same procesy, co w profazie I, z wyjątkiem koniugacji i krzyżowania.
W metafaza II chromosomy znajdują się wzdłuż równika komórki.
W anafaza II chromosomy są rozdzielane w centromerach, a chromatydy rozciągają się w kierunku biegunów.
W telofaza II Błony jądrowe i jąderka powstają wokół skupisk chromosomów potomnych.
Po cytokineza II Wzór genetyczny wszystkich czterech komórek potomnych to 1n1c, ale wszystkie mają inny zestaw genów, co jest wynikiem krzyżowania i losowej kombinacji chromosomów organizmu matki i ojca w komórkach potomnych.
Rozwój komórek rozrodczych u roślin i zwierząt
Gametogeneza(z greckiego gameta- żona, gamety- mąż i geneza- pochodzenie, pojawienie się) to proces powstawania dojrzałych komórek rozrodczych.
Ponieważ rozmnażanie płciowe najczęściej wymaga dwóch osobników - kobiety i mężczyzny, wytwarzających różne komórki płciowe - komórki jajowe i plemniki, wówczas procesy powstawania tych gamet muszą być różne.
Charakter procesu zależy w dużej mierze od tego, czy zachodzi on w komórce roślinnej, czy zwierzęcej, gdyż u roślin podczas tworzenia gamet zachodzi tylko mitoza, a u zwierząt zarówno mitoza, jak i mejoza.
Rozwój komórek rozrodczych w roślinach. U okrytonasiennych tworzenie męskich i żeńskich komórek rozrodczych zachodzi w różnych częściach kwiatu - odpowiednio pręcikach i słupkach.
Przed powstaniem męskich komórek rozrodczych - mikrogametogeneza(z greckiego mikro- mały) - zdarza się mikrosporogeneza, czyli powstawanie mikrospor w pylnikach pręcików. Proces ten związany jest z podziałem mejotycznym komórki macierzystej, w wyniku którego powstają cztery haploidalne mikrospory. Mikrogametogeneza związana jest z mitotycznym podziałem mikrospory, w wyniku którego z dwóch komórek powstaje męski gametofit – duża wegetatywny(syfonogenny) i płytki generatywny. Po podziale gametofit męski pokrywa się gęstymi błonami i tworzy ziarno pyłku. W niektórych przypadkach, już w trakcie dojrzewania pyłku, a czasami dopiero po przeniesieniu na piętno słupka, komórka generatywna dzieli się mitotycznie, tworząc dwie nieruchome męskie komórki płciowe - sperma. Po zapyleniu z komórki wegetatywnej tworzy się łagiewka pyłkowa, przez którą plemniki przedostają się do jajnika słupka w celu zapłodnienia.

Nazywa się rozwój żeńskich komórek rozrodczych w roślinach megagametogeneza(z greckiego mega- duży). Występuje w jajniku słupka, który jest poprzedzony megasporogeneza, w wyniku czego z komórki macierzystej megaspory leżącej w jądrze w wyniku podziału mejotycznego powstają cztery megaspory. Jedna z megaspor dzieli się mitotycznie trzykrotnie, dając żeński gametofit - woreczek zarodkowy z ośmioma jądrami. Wraz z późniejszym oddzieleniem cytoplazmy komórek potomnych, jedna z powstałych komórek staje się jajem, po bokach którego leżą tzw. synergidy, na przeciwległym końcu worka zarodkowego powstają trzy antypody, a pośrodku w wyniku fuzji dwóch jąder haploidalnych powstaje diploidalna komórka centralna.

Rozwój komórek rozrodczych u zwierząt. U zwierząt zachodzą dwa procesy powstawania komórek rozrodczych – spermatogeneza i oogeneza.
Spermatogeneza(z greckiego plemniki, plemniki- nasiona i geneza- pochodzenie, występowanie) to proces powstawania dojrzałych męskich komórek rozrodczych - plemników. U ludzi występuje w jądrach lub jądrach i dzieli się na cztery okresy: rozmnażanie, wzrost, dojrzewanie i tworzenie.

W gody pierwotne komórki rozrodcze dzielą się mitotycznie, w wyniku czego powstają diploidy spermatogonia. W okres wzrostu spermatogonie gromadzą składniki odżywcze w cytoplazmie, powiększają się i przekształcają pierwotne spermatocyty, Lub Spermatocyty I rzędu. Dopiero potem wchodzą w mejozę ( okres dojrzewania), w wyniku czego powstają pierwsze dwa wtórny spermatocyt, Lub Spermatocyt II rzędu, a następnie - cztery komórki haploidalne z wciąż dość dużą ilością cytoplazmy - spermatydy. W okres formacji tracą prawie całą cytoplazmę i tworzą wici, zamieniając się w plemniki.
Sperma, Lub żywe istoty, - bardzo małe mobilne męskie komórki rozrodcze z głową, szyją i ogonem.
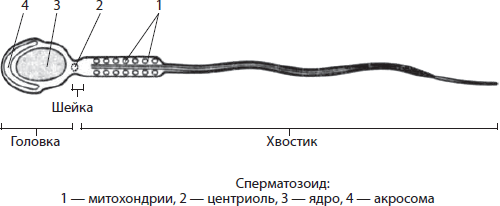
W głowa, oprócz rdzenia, jest akrosom- zmodyfikowany kompleks Golgiego zapewniający rozpuszczenie błon jajowych podczas zapłodnienia. W szyjka macicy to centriole centrum komórki i podstawa koński ogon tworzą mikrotubule, które bezpośrednio wspomagają ruch plemników. Zawiera także mitochondria, które dostarczają plemnikom energię ATP niezbędną do poruszania się.
Oogeneza(z greckiego ONZ- jajko i geneza- pochodzenie, występowanie) to proces powstawania dojrzałych żeńskich komórek rozrodczych - jaj. U ludzi zachodzi w jajnikach i składa się z trzech okresów: reprodukcji, wzrostu i dojrzewania. Okresy reprodukcji i wzrostu, podobne do tych w spermatogenezie, występują podczas rozwoju wewnątrzmacicznego. W tym przypadku komórki diploidalne powstają z pierwotnych komórek rozrodczych w wyniku mitozy. oogonia, które następnie przekształcają się w diploidalny pierwotny oocyty, Lub Oocyty I rzędu. Mejoza i następująca po niej cytokineza zachodząca w okres dojrzewania, charakteryzują się nierównym podziałem cytoplazmy komórki macierzystej, w wyniku czego w pierwszej kolejności powstaje jedna wtórny oocyt, Lub Oocyt II rzędu, I pierwsze ciało polarne, a następnie z wtórnego oocytu - jaja, które zachowuje cały zapas składników odżywczych, i drugiego ciała polarnego, podczas gdy pierwsze ciało polarne dzieli się na dwa. Ciała polarne pochłaniają nadmiar materiału genetycznego.
U ludzi jaja powstają w odstępie 28–29 dni. Cykl związany z dojrzewaniem i uwalnianiem komórek jajowych nazywa się menstruacją.
jajko- duża żeńska komórka rozrodcza, która niesie nie tylko haploidalny zestaw chromosomów, ale także znaczną podaż składników odżywczych dla późniejszego rozwoju zarodka.

Jajo u ssaków jest pokryte czterema błonami, które zmniejszają prawdopodobieństwo uszkodzenia przez różne czynniki. Średnica jaja u człowieka sięga 150-200 mikronów, natomiast u strusia może wynosić kilka centymetrów.
Podział komórek jest podstawą wzrostu, rozwoju i rozmnażania organizmów. Rola mitozy i mejozy
Jeśli w organizmach jednokomórkowych podział komórek prowadzi do wzrostu liczby osobników, czyli rozmnażania, to w organizmach wielokomórkowych proces ten może mieć inne znaczenie. Zatem podział komórek embrionalnych, począwszy od zygoty, jest biologiczną podstawą wzajemnie powiązanych procesów wzrostu i rozwoju. Podobne zmiany obserwuje się u ludzi w okresie dojrzewania, kiedy nie tylko wzrasta liczba komórek, ale także następuje zmiana jakościowa w organizmie. Rozmnażanie organizmów wielokomórkowych również opiera się na podziale komórek, np. W rozmnażaniu bezpłciowym, dzięki temu procesowi przywracana jest cała część organizmu, a w rozmnażaniu płciowym, w procesie gametogenezy powstają komórki płciowe, które następnie dają początek nowemu organizmowi. Należy zauważyć, że główne metody podziału komórki eukariotycznej – mitoza i mejoza – mają różne znaczenie w cyklach życiowych organizmów.
W wyniku mitozy następuje równomierne rozmieszczenie materiału dziedzicznego pomiędzy komórkami potomnymi – dokładnymi kopiami matki. Bez mitozy istnienie i rozwój organizmów wielokomórkowych rozwijających się z pojedynczej komórki, zygoty, byłoby niemożliwe, ponieważ wszystkie komórki takich organizmów muszą zawierać tę samą informację genetyczną.
W procesie podziału komórki potomne stają się coraz bardziej zróżnicowane pod względem struktury i funkcji, co wiąże się z aktywacją w nich coraz większej liczby nowych grup genów w wyniku interakcji międzykomórkowych. Zatem mitoza jest niezbędna do rozwoju organizmu.
Ta metoda podziału komórek jest niezbędna w procesach rozmnażania bezpłciowego i regeneracji (odbudowy) uszkodzonych tkanek i narządów.
Mejoza z kolei zapewnia stałość kariotypu podczas rozmnażania płciowego, ponieważ przed rozmnażaniem płciowym zmniejsza o połowę zestaw chromosomów, który następnie jest przywracany w wyniku zapłodnienia. Ponadto mejoza prowadzi do pojawienia się nowych kombinacji genów rodzicielskich w wyniku krzyżowania i losowego łączenia chromosomów w komórkach potomnych. Dzięki temu potomstwo okazuje się zróżnicowane genetycznie, co dostarcza materiału do doboru naturalnego i stanowi materialną podstawę ewolucji. Zmiana liczby, kształtu i wielkości chromosomów z jednej strony może prowadzić do pojawienia się różnych odchyleń w rozwoju organizmu, a nawet jego śmierci, z drugiej strony może prowadzić do pojawienia się osobników bardziej przystosowane do otoczenia.
Zatem komórka jest jednostką wzrostu, rozwoju i rozmnażania organizmów.
podsumowanie innych prezentacji„Metody nauczania biologii” - Zoologia szkolna. Zapoznanie studentów z wykorzystaniem naukowych danych zoologicznych. Edukacja moralna. Dodatkowe błogosławieństwo kurnika. Wybór metod. Procesy życiowe. Ryby akwariowe. Odżywianie. Edukacja ekologiczna. Materialność procesów życiowych. Wyniki negatywne. Uwaga studenci. Formularz obowiązkowy. Patrząc na małe zwierzęta. Cele i zadania biologii. Fabuła.
„Nauka problemowa na lekcjach biologii” – Wiedza. Nowe podręczniki. Droga do rozwiązania. Problem. Seminaria. Co to jest zadanie? Albrechta Durera. Nauczanie problemowe na lekcjach biologii. Lekcje niestandardowe. Co oznacza uczenie się oparte na problemach? Jakość życia. Biologia jako przedmiot akademicki. Pytanie. Lekcja rozwiązywania problemów. Zmniejszone zainteresowanie tematem. Zajęcia laboratoryjne problemowe.
„Krytyczne myślenie na lekcjach biologii” - Technologia „krytycznego myślenia”. Stosowanie technologii „rozwoju krytycznego myślenia”. Stół do lekcji. Motywacja do nauki. Ekosystemy. Znaczenie „rozwijania krytycznego myślenia”. Znaki technologii. Technologia RKM. Struktura lekcji. Główne kierunki. Historia technologii. Technologie pedagogiczne. Zasady technologii. Zadania z biologii. Fotosynteza. Techniki stosowane na różnych etapach lekcji.
„Lekcje biologii z tablicą interaktywną” - Podręczniki elektroniczne. Korzyści dla studentów. Tablica interaktywna pomaga przekazać informacje każdemu uczniowi. Zadania dydaktyczne. Rozwiązywanie problemów biologicznych. Zalety pracy z tablicami interaktywnymi. Praca z prezentacjami. Pracuj nad porównywaniem obiektów. Poruszające się obiekty. Korzystanie z arkuszy kalkulacyjnych. Wykorzystanie tablicy interaktywnej w procesie nauczania dzieci w wieku szkolnym. Korzyści dla nauczycieli.
„Podejście systemowo-aktywne w biologii” – pytania seminaryjne. Metoda działania. Dryopitek. Pozaziemska ścieżka ludzkiego pochodzenia. Lizosomy. Organizacja chemiczna. Nagonasienne. Metabolizm. Analizatory. Podejście systemowo-aktywne do nauczania biologii. Chromosomy. Cytoplazma. Ślepota. Długość ucha. Klasyfikacja osoby. Szkielet ssaka. Drogi ewolucji człowieka. Mitoza. Powierzchowny kompleks. Problematyczne pytanie. Jądro. Koperta nuklearna.
„Komputer w biologii” – Wspólne działanie studentów. Rodziny okrytozalążkowe. Szkolenie interaktywne. Modele uczenia się. Przykład systemu oceniania. Pytania dotyczące karty instrukcji. Przykład karty instrukcji. Naukowcy. Mikrogrupy. Interaktywne technologie nauczania. Karuzela. Interaktywne technologie nauczania. Interaktywne podejście do lekcji biologii. Grupowa forma pracy. Zadania dla grup „badaczy”.







