Lizosomy. Mitochondria. Plastydy
1. Jaka jest struktura i funkcje ATP?
2. Jakie znasz rodzaje plastydów?
Kiedy różne składniki odżywcze dostają się do komórki poprzez fagocytozę lub pinocytozę, muszą zostać strawione. W której wiewiórki muszą zostać rozbite na poszczególne aminokwasy, polisacharydy – na cząsteczki glukozy lub fruktozy, lipidy- na glicerol i kwasy tłuszczowe. Aby trawienie wewnątrzkomórkowe było możliwe, pęcherzyk fagocytarny lub pinocytarny musi połączyć się z lizosomem (ryc. 25). Lizosom to mały pęcherzyk o średnicy zaledwie 0,5–1,0 mikrona, zawierający duży zestaw enzymów zdolnych do niszczenia składników odżywczych. Jeden lizosom może zawierać 30-50 różnych enzymów.

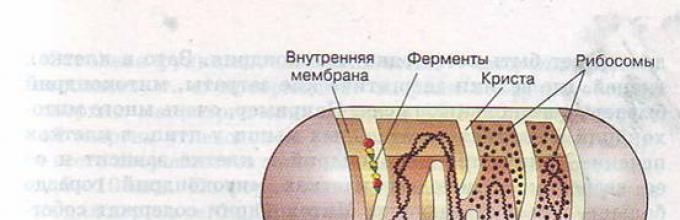
Mitochondria to trwałe organelle błonowe o kształcie okrągłym lub prętowym (często rozgałęzionym). Grubość – 0,5 mikrona, długość – 5–7 mikronów. Liczba mitochondriów w większości komórek zwierzęcych wynosi 150-1500; w jajach samic - do kilkuset tysięcy, w plemnikach - jedno spiralne mitochondrium owinięte wokół osiowej części wici.
Główne funkcje mitochondriów:
1) pełnią rolę stacji energetycznych komórek;
2) przechowują materiał dziedziczny w postaci mitochondrialnego DNA.
Funkcje uboczne - udział w syntezie hormonów steroidowych, niektórych aminokwasów (na przykład glutaminy).
Struktura mitochondriów
Mitochondria mają dwie błony: zewnętrzną (gładką) i wewnętrzną (tworząc narośla - w kształcie liścia (cristae) i rurkowatą (kanaliki)).
W mitochondriach wewnętrzną zawartością jest matrix - substancja koloidalna, w której za pomocą mikroskopu elektronowego odkryto ziarna o średnicy 20–30 nm (akumulują jony wapnia i magnezu, rezerwy składników odżywczych, np. Glikogenu).
W matrycy znajduje się aparat biosyntezy białek organelli: 2–6 kopii kolistego DNA pozbawionego białek histonowych, rybosomów, zestawu t-RNA, enzymów do reduplikacji, transkrypcji i translacji informacji dziedzicznej.
Mitochondria rozmnażają się przez ligację; mitochondria charakteryzują się względną autonomią w obrębie komórki.
Lizosomy to pęcherzyki o średnicy 200–400 µm. (zazwyczaj). Mają jednomembranową skorupę, która czasami jest pokryta na zewnątrz włóknistą warstwą białka. Główną funkcją jest wewnątrzkomórkowe trawienie różnych związków chemicznych i struktur komórkowych.
Wyróżnia się lizosomy pierwotne (nieaktywne) i wtórne (zachodzi w nich proces trawienia). Lizosomy wtórne powstają z lizosomów pierwotnych. Dzielą się na heterolizosomy i autolizosomy.
W heterolizosomach (lub fagolizosomach) proces trawienia materiału, który dostaje się do komórki z zewnątrz, odbywa się poprzez transport aktywny (pinocytoza i fagocytoza).
W autolizosomach (lub cytolizosomach) ich własne struktury komórkowe, które zakończyły swoje życie, ulegają zniszczeniu.
Lizosomy wtórne, które przestały trawić materiał, nazywane są ciałami resztkowymi. Nie zawierają hydrolaz i zawierają niestrawiony materiał.
Kiedy integralność błony lizosomów zostanie naruszona lub gdy komórka jest chora, hydrolazy dostają się do komórki z lizosomów i przeprowadzają jej samostrawienie (autolizę). Ten sam proces leży u podstaw procesu naturalnej śmierci wszystkich komórek (apoptozy).
Mikrociała
Mikrociała tworzą zbiorową grupę organelli. Są to pęcherzyki o średnicy 100–150 nm, ograniczone pojedynczą membraną. Zawierają drobnoziarnistą matrycę i często wtrącenia białkowe.
Trwałe struktury komórkowe, narządy komórkowe, które zapewniają wykonywanie określonych funkcji podczas życia komórki - przechowywanie i przekazywanie informacji genetycznej, przenoszenie substancji, synteza i transformacja substancji i energii, podział, ruch itp.
Do organoidów (organelli) komórek eukarionty odnieść się:
- chromosomy;
- Błona komórkowa;
- mitochondria;
- kompleks Golgiego;
- siateczka śródplazmatyczna;
- rybosomy;
- mikrotubule;
- mikrofilamenty;
- lizosomy.
Komórki zwierzęce zawierają również centriole i mikrofibryle, a komórki roślinne zawierają unikalne dla nich plastydy.
Czasami jądro jako całość klasyfikuje się jako organelle komórek eukariotycznych.
Prokarioty brakuje im większości organelli, mają jedynie błonę komórkową i rybosomy, które różnią się od rybosomów cytoplazmatycznych komórek eukariotycznych.
Wyspecjalizowane komórki eukariotyczne mogą mieć złożone struktury oparte na uniwersalnych organellach, takich jak mikrotubule i centriole – główne składniki wici i rzęsek. Mikrofibryle leżą u podstaw tono- i neurofibryli. Specjalne struktury organizmów jednokomórkowych, takie jak wici i rzęski (zbudowane w taki sam sposób jak w komórkach wielokomórkowych), pełnią funkcję narządów ruchu.
Coraz częściej we współczesnej literaturze używa się określenia „ organoidy " I " organelle „są używane jako synonimy.
Struktury wspólne dla komórek zwierzęcych i roślinnych
| Grafika ze schematem | Struktura | Funkcje |
|
| Błona plazmatyczna (plazmalemma, błona komórkowa) | Dwie warstwy lipidów (dwuwarstwy) pomiędzy dwiema warstwami białka | Selektywnie przepuszczalna bariera regulująca wymianę między komórką a środowiskiem |
|
| Rdzeń | Największa organella, zamknięta w skorupie dwóch membran, przeniknęła pory jądrowe. Zawiera chromatyna- w tej formie rozwinięte chromosomy znajdują się w interfazie. Zawiera również strukturę zwaną jąderko | Chromosomy zawierają DNA – substancję dziedziczności.DNA składa się z genów regulujących wszystkie rodzaje aktywności komórkowej. Podział jądrowy leży u podstaw reprodukcji komórek, a tym samym procesu reprodukcji. Rybosomy powstają w jąderku |
|
| Siateczka śródplazmatyczna (ER) | System spłaszczonych worków membranowych - czołgi- w postaci rur i płytek. Tworzy pojedynczą jednostkę z zewnętrzną błoną otoczki jądrowej | Jeśli powierzchnia ER jest pokryta rybosomami, nazywa się to surowy Białko syntetyzowane na rybosomach transportowane jest przez zbiorniki takiego ER. Gładki Ostry dyżur(bez rybosomów) służy jako miejsce syntezy lipidów i steroidów |
|
| Rybosomy | Bardzo małe organelle składające się z dwóch subcząstek - dużej i małej. Zawierają białko i RNA w mniej więcej równych proporcjach. Rybosomy znajdujące się w mitochondriach (a także w chloroplastach roślin) są jeszcze mniejsze | Miejsce syntezy białek, w którym różne oddziałujące cząsteczki utrzymywane są we właściwej pozycji. Rybosomy są związane z ER lub leżą swobodnie w cytoplazmie. Może powstać wiele rybosomów polisom (polirybosom), w którym są one nawleczone na pojedynczą nić informacyjnego RNA |
|
| Mitochondria | Mitochondrium otoczone jest powłoką złożoną z dwóch błon, błona wewnętrzna tworzy fałdy ( święta). Zawiera matrycę zawierającą niewielką liczbę rybosomów, jedną kolistą cząsteczkę DNA i granulki fosforanowe | Podczas oddychania tlenowego w cristae zachodzi fosforylacja oksydacyjna i transfer elektronów, a w matrixie działają enzymy biorące udział w cyklu Krebsa i utlenianiu kwasów tłuszczowych. |
|
| Aparat Golgiego | Stos spłaszczonych worków błonowych - czołgi. Z jednej strony tworzą się stosy woreczków, z drugiej zaś są one splecione w formie bąbelków. Stosy mogą istnieć jako oddzielne dictyosomy, jak w komórkach roślinnych, lub tworzyć sieć przestrzenną, jak w wielu komórkach zwierzęcych | Wiele materiałów komórkowych, takich jak enzymy z ER, ulega modyfikacji w cisternae i jest transportowanych w pęcherzykach. Aparat Golgiego bierze udział w procesie wydzielania i powstają w nim lizosomy |
|
| Lizosomy | Prosty kulisty worek błonowy (pojedyncza membrana) wypełniony enzymami trawiennymi (hydrolitycznymi). Treść wydaje się jednorodna | Pełnią wiele funkcji, zawsze związanych z rozpadem jakichkolwiek struktur lub cząsteczek |
|
| Mikrociała | Organelle nie mają całkiem regularnego, kulistego kształtu, otoczone pojedynczą błoną. Zawartość ma strukturę ziarnistą, czasem jednak znajduje się w niej krystaloid, czyli zbiór nitek | Wszystkie mikrociała zawierają katalazę, enzym katalizujący rozkład nadtlenku wodoru. Wszystkie są związane z reakcjami oksydacyjnymi |
|
| Ściana komórkowa, blaszka środkowa, plazmodesma |
|||
| Ściana komórkowa | Sztywna ściana komórkowa otaczająca komórkę składa się z mikrofibryli celulozowych osadzonych w matrycy zawierającej inne złożone polisacharydy, a mianowicie hemicelulozy i pektyny. W niektórych komórkach ściany komórkowe ulegają wtórnemu pogrubieniu | Zapewnia mechaniczne wsparcie i ochronę. Dzięki niemu powstaje ciśnienie turgorowe, które wzmacnia funkcję wspomagającą. Zapobiega pękaniu komórek osmotycznych. Wzdłuż ściany komórkowej następuje ruch wody i soli mineralnych. Różne modyfikacje, takie jak impregnacja ligniną, zapewniają wyspecjalizowane funkcje |
|
| płyta środkowa | Cienka warstwa substancji pektynowych (pektyniany wapnia i magnezu) | Utrzymuje komórki razem |
|
| plazmodesma | Cienki włókno cytoplazmatyczne, które łączy cytoplazmę dwóch sąsiadujących komórek poprzez cienki por w ścianie komórkowej. Por jest wyłożony błoną plazmatyczną, przez którą przechodzi desmotubula, często połączona na obu końcach z ER | Połącz protoplasty sąsiadujących komórek w jeden ciągły system - simplast, przez który substancje są transportowane pomiędzy tymi komórkami |
|
| Chloroplast | Duży plastyd zawierający chlorofil, w którym zachodzi fotosynteza. Chloroplast jest otoczony podwójną membraną i wypełniony żelatyną zrąb. Zrąb zawiera system połączonych membran półki na książki, Lub ziarna. Można w nim również osadzać skrobię. Ponadto zrąb zawiera rybosomy, kolistą cząsteczkę DNA i kropelki oleju | W tej organelli zachodzi fotosynteza, czyli synteza cukrów i innych substancji z CO 2 i wody pod wpływem energii świetlnej wychwytywanej przez chlorofil.Energia świetlna zamieniana jest na energię chemiczną |
|
| Duża centralna wakuola | Worek utworzony przez pojedynczą błonę, tzw tonoplast. Wakuola zawiera sok komórkowy – stężony roztwór różnych substancji, takich jak sole mineralne, cukry, pigmenty, kwasy organiczne i enzymy. W dojrzałych komórkach wakuole są zwykle duże | Magazynowane są tu różne substancje, w tym końcowe produkty przemiany materii. Właściwości osmotyczne komórki silnie zależą od zawartości wakuoli. Czasami wakuola pełni funkcję lizosomu |
|
Charakterystyka porównawcza RNA i DNA
| Oznaki | RNA | DNA |
| Lokalizacja w klatce | Jądro, rybosomy, cytoplazma, mitochondria, chloroplasty | Jądro, mitochondria, chloroplasty |
| Lokalizacja w jądrze | Jądro | Chromosomy |
| Struktura makrocząsteczki | Pojedynczy łańcuch polinukleotydowy | Podwójnie nierozgałęziony polimer liniowy, zwinięty w prawoskrętną spiralę |
| Monomery | Rybonukleotydy | Deoksyrybonukleotydy |
| Skład nukleotydów | Zasada azotowa (puryna – adenina, guanina, pirymidyna – uracyl, cytozyna); ryboza (węglowodany): reszta kwasu fosforowego | Zasada azotowa (puryna – adenina, guanina, pirymidyna – tymina, cytozyna); dezoksyryboza (węglowodany): reszta kwasu fosforowego |
| Rodzaje nukleotydów | Alenyl (A), guanyl (G), urydyl (U), cytydyl (C) | Alenyl (A), guanyl (G), tymidyl (T), cytydyl (C) |
| Nieruchomości | Niezdolny do samopodwojenia. Labilna | Zdolne do samopowielania zgodnie z zasadą komplementarności (reduplikacji): A-T, T-A, G-C, C-G Stabilny |
| Funkcje | Informacyjny (mRNA) - przekazuje kod dziedzicznej informacji o pierwotnej strukturze cząsteczki białka; rybosomalny (rRNA) - część rybosomów; transport (tRNA) - przenosi aminokwasy do rybosomów; mitochondrialny i plastydowy RNA - są częścią rybosomów tych organelli | Podstawa chemiczna chromosomalnego materiału genetycznego (genu); Synteza DNA, synteza RNA, informacja o strukturze białek |
Mitochondria to mikroskopijne organelle związane z błoną, które dostarczają komórce energię. Dlatego nazywane są stacjami energetycznymi (bateriami) ogniw.
Mitochondria nie występują w komórkach organizmów prostych, bakterii i entameb, które żyją bez użycia tlenu. Niektóre zielone algi, trypanosomy, zawierają jedno duże mitochondrium, a komórki mięśnia sercowego i mózgu mają od 100 do 1000 takich organelli.
Cechy konstrukcyjne
Mitochondria to organelle dwubłonowe, posiadają błonę zewnętrzną i wewnętrzną, przestrzeń międzybłonową pomiędzy nimi oraz macierz.
Zewnętrzna męmbrana. Jest gładka, nie ma fałd i oddziela zawartość wewnętrzną od cytoplazmy. Jego szerokość wynosi 7 nm i zawiera lipidy i białka. Ważną rolę odgrywa porina, białko tworzące kanały w błonie zewnętrznej. Zapewniają wymianę jonową i molekularną.
Przestrzeń międzybłonowa. Rozmiar przestrzeni międzybłonowej wynosi około 20 nm. Wypełniająca ją substancja ma podobny skład do cytoplazmy, z wyjątkiem dużych cząsteczek, które mogą tu przedostać się jedynie poprzez transport aktywny.

Wewnętrzna membrana. Zbudowany jest głównie z białka, tylko jedna trzecia przypada na substancje lipidowe. Duża liczba białek to białka transportowe, ponieważ w błonie wewnętrznej brakuje swobodnie przepuszczalnych porów. Tworzy wiele odrostów - cristae, które wyglądają jak spłaszczone grzbiety. Utlenianie związków organicznych do CO 2 w mitochondriach zachodzi na błonach cristae. Proces ten jest zależny od tlenu i zachodzi pod wpływem syntetazy ATP. Uwolniona energia jest magazynowana w postaci cząsteczek ATP i wykorzystywana w miarę potrzeb.
Matryca– środowisko wewnętrzne mitochondriów ma ziarnistą, jednorodną strukturę. W mikroskopie elektronowym można zobaczyć granulki i włókna w kulkach swobodnie leżących pomiędzy cristae. W matrixie znajduje się półautonomiczny układ syntezy białek - znajdują się tu DNA, wszystkie rodzaje RNA i rybosomy. Mimo to większość białek pochodzi z jądra, dlatego mitochondria nazywane są organellami półautonomicznymi.
Lokalizacja i podział komórki
Hondriom to grupa mitochondriów skupiona w jednej komórce. Są one różnie zlokalizowane w cytoplazmie, co zależy od specjalizacji komórek. Umieszczenie w cytoplazmie zależy również od otaczających organelli i inkluzji. W komórkach roślinnych zajmują obrzeża, ponieważ mitochondria są wypychane w kierunku błony przez centralną wakuolę. W komórkach nabłonka nerek błona tworzy wypustki, pomiędzy którymi znajdują się mitochondria.
W komórkach macierzystych, gdzie energia jest zużywana równomiernie przez wszystkie organelle, mitochondria są rozmieszczone chaotycznie. W wyspecjalizowanych ogniwach skupiają się one głównie w obszarach o największym zużyciu energii. Na przykład w mięśniach prążkowanych znajdują się one w pobliżu miofibryli. W plemnikach pokrywają one spiralnie oś wici, ponieważ do jej wprawienia i poruszenia plemnikiem potrzeba dużo energii. Pierwotniaki poruszające się za pomocą rzęsek również zawierają u podstawy dużą liczbę mitochondriów.
Dział. Mitochondria są zdolne do niezależnej reprodukcji, mając własny genom. Organelle są podzielone przewężeniami lub przegrodami. Tworzenie nowych mitochondriów w różnych komórkach różni się częstotliwością, np. w tkance wątroby są one wymieniane co 10 dni.
Funkcje w komórce
- Główną funkcją mitochondriów jest tworzenie cząsteczek ATP.
- Odkładanie jonów wapnia.
- Udział w wymianie wody.
- Synteza prekursorów hormonów steroidowych.
Biologia molekularna to nauka badająca rolę mitochondriów w metabolizmie. Przekształcają także pirogronian w acetylokoenzym A i beta-oksydację kwasów tłuszczowych.
| Tabela: budowa i funkcje mitochondriów (w skrócie) | ||
|---|---|---|
| Elementy konstrukcyjne | Struktura | Funkcje |
| Zewnętrzna męmbrana | Gładka skorupa zbudowana z lipidów i białek | Oddziela zawartość wewnętrzną od cytoplazmy |
| Przestrzeń międzybłonowa | Są jony wodoru, białka, mikrocząsteczki | Tworzy gradient protonowy |
| Wewnętrzna membrana | Tworzy wypustki - cristae, zawiera systemy transportu białek | Transfer makrocząsteczek, utrzymanie gradientu protonów |
| Matryca | Lokalizacja enzymów cyklu Krebsa, DNA, RNA, rybosomów | Utlenianie tlenowe z uwolnieniem energii, konwersja pirogronianu do acetylokoenzymu A. |
| Rybosomy | Połączone dwie podjednostki | Synteza białek |
Podobieństwa między mitochondriami i chloroplastami
Wspólne właściwości mitochondriów i chloroplastów wynikają przede wszystkim z obecności podwójnej błony.
Oznaki podobieństwa obejmują także zdolność do samodzielnej syntezy białka. Organelle te mają własne DNA, RNA i rybosomy.
Zarówno mitochondria, jak i chloroplasty mogą dzielić się przez zwężenie.
Łączy je również zdolność do wytwarzania energii; mitochondria są bardziej wyspecjalizowane w tej funkcji, ale chloroplasty wytwarzają również cząsteczki ATP podczas procesów fotosyntezy. Zatem komórki roślinne mają mniej mitochondriów niż komórki zwierzęce, ponieważ chloroplasty częściowo pełnią za nie funkcje.
Opiszmy pokrótce podobieństwa i różnice:
- Są to organelle dwubłonowe;
- wewnętrzna błona tworzy wypukłości: cristae są charakterystyczne dla mitochondriów, a thillacoidy są charakterystyczne dla chloroplastów;
- mają swój własny genom;
- zdolne do syntezy białek i energii.
Organelle te różnią się funkcjami: mitochondria służą do syntezy energii, zachodzi tu oddychanie komórkowe, chloroplasty są potrzebne komórkom roślinnym do fotosyntezy.
Treść artykułu
KOMÓRKA, podstawowa jednostka istot żywych. Komórka jest oddzielona od innych komórek lub od środowiska zewnętrznego specjalną membraną i posiada jądro lub jego odpowiednik, w którym koncentruje się większość informacji chemicznej kontrolującej dziedziczność. Cytologia bada strukturę komórek, a fizjologia zajmuje się ich funkcjonowaniem. Nauka badająca tkanki zbudowane z komórek nazywa się histologią.
Istnieją organizmy jednokomórkowe, których całe ciało składa się z jednej komórki. Do tej grupy zaliczają się bakterie i protisty (pierwotniaki i glony jednokomórkowe). Czasami nazywane są również bezkomórkowymi, ale częściej używa się terminu jednokomórkowe. Prawdziwe zwierzęta wielokomórkowe (Metazoa) i rośliny (Metaphyta) zawierają wiele komórek.
Zdecydowana większość tkanek składa się z komórek, ale są pewne wyjątki. Na przykład ciało śluzowców (myxomycetes) składa się z jednorodnej substancji, która nie jest podzielona na komórki z licznymi jądrami. Niektóre tkanki zwierzęce, w szczególności mięsień sercowy, są zorganizowane w podobny sposób. Ciało wegetatywne (talus) grzybów jest utworzone przez mikroskopijne nici - strzępki, często podzielone na segmenty; każdy taki wątek można uznać za odpowiednik komórki, choć o nietypowym kształcie.
Niektóre struktury organizmu, które nie biorą udziału w metabolizmie, w szczególności muszle, perły lub mineralna podstawa kości, nie są tworzone przez komórki, ale przez produkty ich wydzielania. Inne, takie jak drewno, kora, rogi, sierść i zewnętrzna warstwa skóry, nie są pochodzenia wydzielniczego, ale powstają z martwych komórek.
Małe organizmy, takie jak wrotki, składają się z zaledwie kilkuset komórek. Dla porównania: w organizmie człowieka znajduje się ok. 10 14 komórek, co sekundę 3 miliony czerwonych krwinek umiera i są zastępowane nowymi, a to tylko jedna dziesięciomilionowa całkowitej liczby komórek ciała.
Zazwyczaj rozmiary komórek roślinnych i zwierzęcych mieszczą się w zakresie od 5 do 20 mikronów średnicy. Typowa komórka bakteryjna jest znacznie mniejsza – ok. 2 mikrony, a najmniejsza znana to 0,2 mikrona.
Niektóre wolno żyjące komórki, takie jak pierwotniaki, takie jak otwornice, mogą osiągnąć kilka centymetrów; zawsze mają wiele rdzeni. Komórki cienkich włókien roślinnych osiągają długość jednego metra, a procesy komórek nerwowych u dużych zwierząt sięgają kilku metrów. Przy takiej długości objętość tych komórek jest niewielka, ale powierzchnia jest bardzo duża.
Największe komórki to niezapłodnione jaja ptasie wypełnione żółtkiem. Największe jajo (a zatem największa komórka) należało do wymarłego ogromnego ptaka - apyornis ( Aepyornis). Prawdopodobnie jego żółtko ważyło ok. 3,5 kg. Największe jajo wśród żyjących gatunków należy do strusia, którego żółtko waży ok. 0,5 kg.
Z reguły komórki dużych zwierząt i roślin są tylko nieznacznie większe niż komórki małych organizmów. Słoń jest większy od myszy nie dlatego, że jego komórki są większe, ale głównie dlatego, że samych komórek jest znacznie więcej. Istnieją grupy zwierząt, takie jak wrotki i nicienie, u których liczba komórek w organizmie pozostaje stała. Tak więc, chociaż duże gatunki nicieni mają większą liczbę komórek niż małe, główna różnica w wielkości wynika w tym przypadku z większych rozmiarów komórek.
W obrębie danego typu komórek ich rozmiary zależą zwykle od ploidii, tj. na liczbę zestawów chromosomów obecnych w jądrze. Komórki tetraploidalne (z czterema zestawami chromosomów) mają dwukrotnie większą objętość niż komórki diploidalne (z dwoma zestawami chromosomów). Ploidię rośliny można zwiększyć wprowadzając do niej ziołowy lek kolchicynę. Ponieważ rośliny narażone na ten efekt mają większe komórki, same są większe. Jednak zjawisko to można zaobserwować tylko w poliploidach niedawnego pochodzenia. W ewolucyjnie starożytnych roślinach poliploidalnych rozmiary komórek podlegają „odwrotnej regulacji” w kierunku normalnych wartości pomimo wzrostu liczby chromosomów.
STRUKTURA KOMÓRKOWA
Kiedyś komórkę uważano za mniej lub bardziej jednorodną kroplę materii organicznej, którą nazywano protoplazmą lub substancją żywą. Termin ten stracił aktualność po odkryciu, że komórka składa się z wielu wyraźnie odrębnych struktur zwanych organellami komórkowymi („małymi organami”).
Skład chemiczny.
Zwykle 70–80% masy komórek stanowi woda, w której rozpuszczone są różne sole i związki organiczne o niskiej masie cząsteczkowej. Najbardziej charakterystycznymi składnikami komórki są białka i kwasy nukleinowe. Niektóre białka są składnikami strukturalnymi komórki, inne są enzymami, tj. katalizatory decydujące o szybkości i kierunku reakcji chemicznych zachodzących w komórkach. Kwasy nukleinowe służą jako nośniki informacji dziedzicznej, która realizowana jest w procesie wewnątrzkomórkowej syntezy białek.
Często komórki zawierają pewną ilość substancji magazynujących, które służą jako rezerwa pożywienia. Komórki roślinne przechowują głównie skrobię, polimerową formę węglowodanów. Inny polimer węglowodanowy, glikogen, jest magazynowany w komórkach wątroby i mięśni. Często przechowywana żywność zawiera również tłuszcz, chociaż niektóre tłuszcze pełnią inną funkcję, a mianowicie pełnią funkcję niezbędnych składników strukturalnych. Białka w komórkach (z wyjątkiem komórek nasiennych) zwykle nie są magazynowane.
Nie da się opisać typowego składu komórki, przede wszystkim ze względu na duże różnice w ilości przechowywanego pożywienia i wody. Komórki wątroby zawierają na przykład 70% wody, 17% białek, 5% tłuszczów, 2% węglowodanów i 0,1% kwasów nukleinowych; pozostałe 6% pochodzi z soli i związków organicznych o niskiej masie cząsteczkowej, w szczególności aminokwasów. Komórki roślinne zazwyczaj zawierają mniej białka, znacznie więcej węglowodanów i nieco więcej wody; wyjątkiem są komórki będące w stanie spoczynku. Komórka spoczynkowa ziarna pszenicy, która jest źródłem składników odżywczych dla zarodka, zawiera ok. 12% białka (głównie białko zmagazynowane), 2% tłuszczu i 72% węglowodanów. Ilość wody osiąga normalny poziom (70–80%) dopiero na początku kiełkowania ziarna.
Główne części komórki.
Niektóre komórki, głównie roślinne i bakteryjne, mają zewnętrzną ścianę komórkową. U roślin wyższych składa się z celulozy. Ściana otacza samą komórkę, chroniąc ją przed naprężeniami mechanicznymi. Komórki, szczególnie komórki bakteryjne, mogą również wydzielać substancje śluzowe, tworząc w ten sposób wokół siebie otoczkę, która podobnie jak ściana komórkowa pełni funkcję ochronną.
To właśnie z niszczeniem ścian komórkowych wiąże się śmierć wielu bakterii pod wpływem penicyliny. Faktem jest, że wewnątrz komórki bakteryjnej stężenie soli i związków drobnocząsteczkowych jest bardzo duże, dlatego w przypadku braku ściany wzmacniającej napływ wody do komórki pod wpływem ciśnienia osmotycznego może doprowadzić do jej rozerwania. Penicylina, która zapobiega tworzeniu się jej ściany podczas wzrostu komórek, prowadzi do rozerwania komórek (lizy).
Ściany komórkowe i kapsułki nie biorą udziału w metabolizmie i często można je oddzielić bez zabijania komórki. Można je zatem uznać za zewnętrzne pomocnicze części komórki. Komórkom zwierzęcym zwykle brakuje ścian komórkowych i torebek.
Sama komórka składa się z trzech głównych części. Poniżej ściany komórkowej, jeśli występuje, znajduje się błona komórkowa. Błona otacza heterogenny materiał zwany cytoplazmą. Okrągłe lub owalne jądro jest zanurzone w cytoplazmie. Poniżej przyjrzymy się bardziej szczegółowo strukturze i funkcjom tych części komórki.
BŁONA KOMÓRKOWA
Błona komórkowa jest bardzo ważną częścią komórki. Łączy wszystkie elementy komórkowe i wyznacza środowisko wewnętrzne i zewnętrzne. Ponadto zmodyfikowane fałdy błony komórkowej tworzą wiele organelli komórkowych.
Błona komórkowa to podwójna warstwa cząsteczek (warstwa dwucząsteczkowa lub dwuwarstwowa). Są to głównie cząsteczki fosfolipidów i innych substancji z nimi powiązanych. Cząsteczki lipidów mają dwoistą naturę, objawiającą się tym, jak zachowują się w stosunku do wody. Głowy cząsteczek są hydrofilowe, tj. mają powinowactwo do wody, a ich ogony węglowodorowe są hydrofobowe. Dlatego lipidy po zmieszaniu z wodą tworzą na swojej powierzchni film podobny do filmu olejowego; Co więcej, wszystkie ich cząsteczki są zorientowane w ten sam sposób: głowy cząsteczek znajdują się w wodzie, a ogony węglowodorowe znajdują się nad jej powierzchnią.
W błonie komórkowej występują dwie takie warstwy i w każdej z nich głowy cząsteczek są zwrócone na zewnątrz, a ogony skierowane do wnętrza błony, jedna w stronę drugiej, dzięki czemu nie mają kontaktu z wodą. Grubość takiej membrany wynosi ok. 7 nm. Oprócz głównych składników lipidowych zawiera duże cząsteczki białka, które potrafią „unosić się” w dwuwarstwie lipidowej i są ułożone tak, że jedna strona jest zwrócona do wnętrza komórki, a druga ma kontakt ze środowiskiem zewnętrznym. Niektóre białka znajdują się tylko na zewnętrznej lub tylko wewnętrznej powierzchni błony lub są tylko częściowo zanurzone w dwuwarstwie lipidowej.
Główną funkcją błony komórkowej jest regulacja transportu substancji do i z komórki. Ponieważ membrana fizycznie przypomina nieco olej, substancje rozpuszczalne w oleju lub rozpuszczalnikach organicznych, takich jak eter, łatwo przez nią przechodzą. To samo dotyczy gazów, takich jak tlen i dwutlenek węgla. Jednocześnie membrana jest praktycznie nieprzepuszczalna dla większości substancji rozpuszczalnych w wodzie, w szczególności cukrów i soli. Dzięki tym właściwościom jest w stanie utrzymać wewnątrz komórki środowisko chemiczne odmienne od tego na zewnątrz. Przykładowo we krwi stężenie jonów sodu jest wysokie, a jonów potasu niskie, natomiast w płynie wewnątrzkomórkowym jony te występują w odwrotnej proporcji. Podobna sytuacja jest typowa dla wielu innych związków chemicznych.
Wiadomo jednak, że komórki nie da się całkowicie odizolować od środowiska, gdyż musi ona otrzymać substancje niezbędne do metabolizmu i pozbyć się produktów końcowych. Ponadto dwuwarstwa lipidowa nie jest całkowicie nieprzepuszczalna nawet dla substancji rozpuszczalnych w wodzie i tzw. penetrujących ją. Białka „tworzące kanały” tworzą pory lub kanały, które mogą otwierać się i zamykać (w zależności od zmian w konformacji białka) oraz w stanie otwartym przewodzić pewne jony (Na +, K +, Ca 2+) wzdłuż gradientu stężeń . W konsekwencji różnica stężeń wewnątrz i na zewnątrz komórki nie może być utrzymana wyłącznie ze względu na niską przepuszczalność membrany. Tak naprawdę zawiera białka, które pełnią funkcję molekularnej „pompy”: transportują określone substancje zarówno do, jak i z komórki, działając wbrew gradientowi stężeń. W rezultacie, gdy stężenie np. aminokwasów wewnątrz komórki jest wysokie, a na zewnątrz niskie, aminokwasy mogą mimo to przepływać ze środowiska zewnętrznego do wewnętrznego. Transfer ten nazywa się transportem aktywnym i wykorzystuje energię dostarczaną przez metabolizm. Pompy membranowe są bardzo specyficzne: każda z nich jest w stanie transportować albo tylko jony określonego metalu, albo aminokwas, albo cukier. Kanały jonowe błony są również specyficzne.
Taka selektywna przepuszczalność jest fizjologicznie bardzo ważna, a jej brak jest pierwszym dowodem śmierci komórki. Łatwo to zilustrować na przykładzie buraków. Jeśli żywy korzeń buraka zanurzy się w zimnej wodzie, zachowuje swój pigment; jeśli buraki zostaną ugotowane, komórki obumierają, stają się łatwo przepuszczalne i tracą pigment, co powoduje, że woda staje się czerwona.
Komórka może „połykać” duże cząsteczki, takie jak białka. Pod wpływem niektórych białek, jeśli są one obecne w płynie otaczającym komórkę, następuje inwazja w błonie komórkowej, która następnie zamyka się, tworząc pęcherzyk – małą wakuolę zawierającą cząsteczki wody i białka; Następnie błona wokół wakuoli pęka, a zawartość dostaje się do komórki. Proces ten nazywany jest pinocytozą (dosłownie „piciem komórki”) lub endocytozą.
Większe cząstki, np. cząstki jedzenia, można w podobny sposób wchłonąć podczas tzw. fagocytoza. Zazwyczaj wakuola powstająca podczas fagocytozy jest większa, a żywność jest trawiona przez enzymy lizosomalne wewnątrz wakuoli, zanim otaczająca błona pęknie. Ten sposób odżywiania jest typowy dla pierwotniaków, takich jak ameby, które zjadają bakterie. Jednak zdolność do fagocytozy jest charakterystyczna zarówno dla komórek jelitowych zwierząt niższych, jak i fagocytów, jednego z rodzajów białych krwinek (leukocytów) kręgowców. W tym drugim przypadku znaczenie tego procesu nie polega na odżywianiu samych fagocytów, ale na ich niszczeniu bakterii, wirusów i innych obcych materiałów szkodliwych dla organizmu.
Funkcje wakuoli mogą być różne. Na przykład pierwotniaki żyjące w słodkiej wodzie doświadczają stałego osmotycznego dopływu wody, ponieważ stężenie soli wewnątrz komórki jest znacznie wyższe niż na zewnątrz. Są w stanie wydzielać wodę do specjalnej wakuoli wydalniczej (kurczliwej), która okresowo wypycha jej zawartość.
Komórki roślinne często mają jedną dużą centralną wakuolę zajmującą prawie całą komórkę; cytoplazma tworzy tylko bardzo cienką warstwę pomiędzy ścianą komórkową a wakuolą. Jedną z funkcji takiej wakuoli jest gromadzenie wody, co pozwala komórce szybko zwiększać rozmiar. Zdolność ta jest szczególnie potrzebna w okresie wzrostu tkanek roślinnych i tworzenia struktur włóknistych.
W tkankach, w miejscach ścisłego połączenia komórek, w ich błonach znajdują się liczne pory utworzone przez przenikające przez błonę białka – tzw. połączenia. Pory sąsiednich komórek znajdują się naprzeciw siebie, dzięki czemu substancje niskocząsteczkowe mogą przechodzić z komórki do komórki - ten system komunikacji chemicznej koordynuje ich aktywność życiową. Przykładem takiej koordynacji jest obserwowany w wielu tkankach mniej lub bardziej synchroniczny podział sąsiadujących komórek.
CYTOPLAZMA
Cytoplazma zawiera błony wewnętrzne, które są podobne do błony zewnętrznej i tworzą różnego rodzaju organelle. Błony te można traktować jako fałdy błony zewnętrznej; czasami membrany wewnętrzne stanowią integralną część z membraną zewnętrzną, ale często fałda wewnętrzna jest rozsznurowana i kontakt z membraną zewnętrzną zostaje przerwany. Jednakże, nawet jeśli kontakt zostanie utrzymany, membrana wewnętrzna i zewnętrzna nie zawsze są chemicznie identyczne. W szczególności skład białek błonowych różni się w różnych organellach komórkowych.
Siateczka endoplazmatyczna.
Sieć błon wewnętrznych składająca się z kanalików i pęcherzyków rozciąga się od powierzchni komórki do jądra. Sieć ta nazywa się retikulum endoplazmatycznym. Często zwracano uwagę, że kanaliki otwierają się na powierzchni komórki, a siateczka śródplazmatyczna pełni w ten sposób rolę aparatu mikrokrążenia, za pośrednictwem którego środowisko zewnętrzne może bezpośrednio oddziaływać z całą zawartością komórki. Interakcję tę stwierdzono w niektórych komórkach, zwłaszcza komórkach mięśniowych, lecz nie jest jeszcze jasne, czy jest ona powszechna. W każdym razie faktycznie zachodzi transport wielu substancji przez te kanaliki z jednej części komórki do drugiej.
Drobne ciałka zwane rybosomami pokrywają powierzchnię retikulum endoplazmatycznego, zwłaszcza w pobliżu jądra. Średnica rybosomu ok. 15 nm, składają się w połowie z białek i w połowie z kwasów rybonukleinowych. Ich główną funkcją jest synteza białek; informacyjny RNA i aminokwasy związane z transferowym RNA są przyłączone do ich powierzchni. Obszary siateczki pokryte rybosomami nazywane są szorstką siateczką śródplazmatyczną, a te bez nich nazywane są gładkimi. Oprócz rybosomów na retikulum endoplazmatycznym lub w inny sposób są do niego adsorbowane różne enzymy, w tym układy enzymatyczne, które zapewniają wykorzystanie tlenu do tworzenia steroli i neutralizacji niektórych trucizn. W niesprzyjających warunkach siateczka śródplazmatyczna szybko ulega degeneracji, dlatego jej stan służy jako czuły wskaźnik stanu zdrowia komórek.
Aparat Golgiego.
Aparat Golgiego (kompleks Golgiego) jest wyspecjalizowaną częścią retikulum endoplazmatycznego, składającą się z ułożonych w stos płaskich worków błonowych. Bierze udział w wydzielaniu białek przez komórkę (następuje w niej pakowanie wydzielanych białek w granulki) i dlatego jest szczególnie rozwinięty w komórkach pełniących funkcję wydzielniczą. Do ważnych funkcji aparatu Golgiego należy także przyłączanie grup węglowodanowych do białek i wykorzystanie tych białek do budowy błony komórkowej i błony lizosomów. U niektórych glonów włókna celulozowe są syntetyzowane w aparacie Golgiego.
Lizosomy
- Są to małe bąbelki otoczone pojedynczą membraną. Pączkują z aparatu Golgiego i prawdopodobnie z siateczki śródplazmatycznej. Lizosomy zawierają różnorodne enzymy rozkładające duże cząsteczki, w szczególności białka. Enzymy te, ze względu na swoje destrukcyjne działanie, są niejako „zamknięte” w lizosomach i uwalniane tylko w razie potrzeby. Zatem podczas trawienia wewnątrzkomórkowego enzymy są uwalniane z lizosomów do wakuoli trawiennych. Lizosomy są również niezbędne do niszczenia komórek; na przykład podczas przemiany kijanki w dorosłą żabę uwolnienie enzymów lizosomalnych zapewnia zniszczenie komórek ogona. W tym przypadku jest to normalne i korzystne dla organizmu, jednak czasami takie niszczenie komórek jest patologiczne. Na przykład wdychany pył azbestu może przedostać się do komórek płuc, w wyniku czego lizosomy pękają, następuje zniszczenie komórek i rozwija się choroba płuc.
Mitochondria i chloroplasty.
Mitochondria to stosunkowo duże struktury przypominające worki o dość złożonej strukturze. Składają się z matrycy otoczonej błoną wewnętrzną, przestrzeni międzybłonowej i błony zewnętrznej. Wewnętrzna membrana jest złożona w fałdy zwane cristae. Skupiska białek znajdują się na cristae. Wiele z nich to enzymy katalizujące utlenianie produktów rozkładu węglowodanów; inne katalizują reakcje syntezy i utleniania tłuszczów. Enzymy pomocnicze biorące udział w tych procesach są rozpuszczone w matrix mitochondrialnej.
W mitochondriach zachodzi utlenianie substancji organicznych połączone z syntezą adenozynotrójfosforanu (ATP). Rozkładowi ATP do difosforanu adenozyny (ADP) towarzyszy uwolnienie energii, która jest zużywana na różne procesy życiowe, na przykład syntezę białek i kwasów nukleinowych, transport substancji do i z komórki, transmisję impulsów nerwowych lub skurczów mięśni. Mitochondria są zatem stacjami energetycznymi, które przetwarzają „paliwo” – tłuszcze i węglowodany – w formę energii, która może zostać wykorzystana przez komórkę, a tym samym organizm jako całość.
Komórki roślinne również zawierają mitochondria, ale głównym źródłem energii dla ich komórek jest światło. Energia świetlna jest wykorzystywana przez te komórki do produkcji ATP i syntezy węglowodanów z dwutlenku węgla i wody. Chlorofil, pigment gromadzący energię świetlną, występuje w chloroplastach. Chloroplasty, podobnie jak mitochondria, mają błonę wewnętrzną i zewnętrzną. Z narośli błony wewnętrznej podczas rozwoju chloroplastów powstają tak zwane chloroplasty. błony tylakoidowe; te ostatnie tworzą spłaszczone worki, zebrane w stosy niczym kolumna monet; te stosy, zwane grana, zawierają chlorofil. Oprócz chlorofilu chloroplasty zawierają wszystkie inne składniki niezbędne do fotosyntezy.
Niektóre wyspecjalizowane chloroplasty nie przeprowadzają fotosyntezy, ale pełnią inne funkcje, takie jak przechowywanie skrobi lub pigmentów.
Względna autonomia.
Pod pewnymi względami mitochondria i chloroplasty zachowują się jak organizmy autonomiczne. Na przykład, podobnie jak same komórki, które powstają wyłącznie z komórek, mitochondria i chloroplasty powstają wyłącznie z wcześniej istniejących mitochondriów i chloroplastów. Wykazano to w doświadczeniach na komórkach roślinnych, w których tworzenie chloroplastów było hamowane przez antybiotyk streptomycynę, oraz na komórkach drożdży, gdzie tworzenie mitochondriów było hamowane przez inne leki. Po takich efektach komórki nigdy nie odtworzyły brakujących organelli. Powodem jest to, że mitochondria i chloroplasty zawierają pewną ilość własnego materiału genetycznego (DNA), który koduje część ich struktury. Jeśli DNA zostanie utracone, co ma miejsce, gdy tworzenie organelli zostanie zahamowane, struktura nie będzie mogła zostać odtworzona. Obydwa typy organelli mają swój własny system syntezy białek (rybosomy i przenoszące RNA), który różni się nieco od głównego układu syntezy białek w komórce; wiadomo na przykład, że układ organelli syntetyzujący białka można stłumić za pomocą antybiotyków, podczas gdy nie mają one wpływu na główny układ.
DNA organelli jest odpowiedzialne za większość dziedziczenia pozachromosomalnego, czyli cytoplazmatycznego. Dziedziczenie pozachromosomalne nie podlega prawom Mendla, ponieważ podczas podziału komórki DNA organelli jest przekazywane komórkom potomnym w inny sposób niż chromosomy. Badanie mutacji zachodzących w DNA organelli i DNA chromosomów wykazało, że DNA organelli jest odpowiedzialne tylko za niewielką część struktury organelli; większość ich białek jest kodowana w genach zlokalizowanych na chromosomach.
Częściowa autonomia genetyczna rozpatrywanych organelli oraz cechy ich systemów syntezy białek posłużyły za podstawę do założenia, że mitochondria i chloroplasty pochodzą od bakterii symbiotycznych, które zadomowiły się w komórkach 1–2 miliardy lat temu. Współczesnym przykładem takiej symbiozy są małe algi fotosyntetyzujące żyjące w komórkach niektórych koralowców i mięczaków. Glony dostarczają żywicielom tlen i pobierają od nich składniki odżywcze.
Struktury włókniste.
Cytoplazma komórki jest lepkim płynem, więc napięcie powierzchniowe powoduje, że można oczekiwać, że komórka będzie kulista, chyba że komórki będą ciasno upakowane. Jednak zwykle nie jest to przestrzegane. Wiele pierwotniaków ma gęste powłoki lub błony, które nadają komórce specyficzny, niekulisty kształt. Niemniej jednak, nawet bez błony, komórki mogą zachować kształt niekulisty ze względu na fakt, że cytoplazma jest zbudowana za pomocą licznych, raczej sztywnych, równoległych włókien. Te ostatnie są utworzone przez puste mikrotubule, które składają się z jednostek białkowych zorganizowanych w spiralę.
Niektóre pierwotniaki tworzą pseudopodia – długie, cienkie wypustki cytoplazmatyczne, którymi chwytają pożywienie. Pseudopodia zachowują swój kształt dzięki sztywności mikrotubul. Jeśli ciśnienie hydrostatyczne wzrośnie do około 100 atmosfer, mikrotubule rozpadają się, a komórka przyjmuje kształt kropli. Kiedy ciśnienie wraca do normy, mikrotubule ponownie się łączą, a komórka tworzy pseudopodia. Wiele innych komórek reaguje w podobny sposób na zmiany ciśnienia, co potwierdza udział mikrotubul w utrzymaniu kształtu komórki. Montaż i rozpad mikrotubul, niezbędny do szybkiej zmiany kształtu komórki, zachodzi nawet przy braku zmian ciśnienia.
Mikrotubule tworzą również struktury włókniste, które służą jako narządy ruchu komórek. Niektóre komórki mają biczowate wypustki zwane wiciami lub rzęskami – ich bicie zapewnia ruch komórki w wodzie. Jeśli komórka jest nieruchoma, struktury te wypychają wodę, cząsteczki jedzenia i inne cząstki w kierunku komórki lub od niej. Wici są stosunkowo duże i zwykle komórka ma tylko jedną, czasem kilka wici. Rzęski są znacznie mniejsze i pokrywają całą powierzchnię komórki. Chociaż struktury te są charakterystyczne głównie dla najprostszych, mogą występować również w formach wysoce zorganizowanych. W organizmie człowieka wszystkie drogi oddechowe są wyścielone rzęskami. Małe cząsteczki, które dostają się do nich, są zwykle wychwytywane przez śluz na powierzchni komórek, a rzęski wypychają je wraz ze śluzem, chroniąc w ten sposób płuca. Męskie komórki rozrodcze większości zwierząt i niektórych niższych roślin poruszają się za pomocą wici.
Istnieją inne rodzaje ruchu komórkowego. Jednym z nich jest ruch ameboidalny. Ameba, podobnie jak niektóre komórki organizmów wielokomórkowych, „przepływa” z miejsca na miejsce, tj. poruszać się pod wpływem przepływu zawartości komórki. Wewnątrz komórek roślinnych istnieje także stały prąd materii, jednak nie powoduje on ruchu komórki jako całości. Najbardziej zbadanym rodzajem ruchu komórkowego jest skurcz komórek mięśniowych; odbywa się to poprzez przesuwanie się względem siebie włókienek (nici białkowych), co prowadzi do skrócenia komórki.
RDZEŃ
Jądro otoczone jest podwójną błoną. Bardzo wąska (około 40 nm) przestrzeń pomiędzy dwiema błonami nazywana jest okołojądrową. Błony jądrowe przechodzą do błon retikulum endoplazmatycznego, a przestrzeń okołojądrowa otwiera się na przestrzeń siatkową. Zazwyczaj błona jądrowa ma bardzo wąskie pory. Najwyraźniej transportowane są przez nie duże cząsteczki, takie jak informacyjny RNA, który jest syntetyzowany na DNA, a następnie przedostaje się do cytoplazmy.
Większość materiału genetycznego zlokalizowana jest w chromosomach jądra komórkowego. Chromosomy składają się z długich łańcuchów dwuniciowego DNA, do którego przyłączone są zasadowe (tj. zasadowe) białka. Czasami chromosomy mają kilka identycznych nici DNA leżących obok siebie - takie chromosomy nazywane są polietylenem (wieloniciowym). Liczba chromosomów jest różna u różnych gatunków. Komórki diploidalne ludzkiego ciała zawierają 46 chromosomów, czyli 23 pary.
W komórce niedzielącej się chromosomy są przyczepione w jednym lub większej liczbie punktów do błony jądrowej. W normalnym stanie rozwiniętym chromosomy są tak cienkie, że nie są widoczne pod mikroskopem świetlnym. W określonych loci (odcinkach) jednego lub większej liczby chromosomów powstaje gęste ciało, które występuje w jądrach większości komórek – tzw. jąderko. W jąderkach zachodzi synteza i akumulacja RNA wykorzystywanego do budowy rybosomów, a także niektórych innych typów RNA.
PODZIAŁ KOMÓREK
Chociaż wszystkie komórki powstają w wyniku podziału poprzedniej komórki, nie wszystkie dzielą się dalej. Na przykład komórki nerwowe w mózgu po utworzeniu nie dzielą się. Ich liczba stopniowo maleje; Uszkodzona tkanka mózgowa nie jest w stanie zregenerować się poprzez regenerację. Jeśli komórki nadal się dzielą, charakteryzują się cyklem komórkowym składającym się z dwóch głównych etapów: interfazy i mitozy.
Sama interfaza składa się z trzech faz: G 1, S i G 2. Poniżej przedstawiono czas ich trwania, typowy dla komórek roślinnych i zwierzęcych.
G 1 (4–8 godzin). Faza ta rozpoczyna się natychmiast po narodzinach komórki. W fazie G 1 komórka, z wyjątkiem chromosomów (które się nie zmieniają), zwiększa swoją masę. Jeśli komórka nie dzieli się dalej, pozostaje w tej fazie.
S (6–9 godzin). Masa komórek stale rośnie i następuje podwojenie (duplikacja) chromosomalnego DNA. Jednakże chromosomy pozostają pojedyncze w strukturze, chociaż ich masa jest podwojona, ponieważ dwie kopie każdego chromosomu (chromatydy) są nadal połączone ze sobą na całej swojej długości.
G2. Masa komórki nadal rośnie, aż osiągnie w przybliżeniu dwukrotność masy pierwotnej, po czym następuje mitoza.
Po zduplikowaniu chromosomów każda z komórek potomnych powinna otrzymać pełny zestaw chromosomów. Prosty podział komórki nie jest w stanie tego osiągnąć – wynik ten osiąga się w procesie zwanym mitozą. Nie wchodząc w szczegóły, za początek tego procesu należy uznać ułożenie chromosomów w płaszczyźnie równikowej komórki. Następnie każdy chromosom dzieli się wzdłużnie na dwie chromatydy, które zaczynają się rozchodzić w przeciwnych kierunkach, stając się niezależnymi chromosomami. W rezultacie pełny zestaw chromosomów znajduje się na obu końcach komórki. Następnie komórka dzieli się na dwie części, a każda komórka potomna otrzymuje pełny zestaw chromosomów.
Poniżej znajduje się opis mitozy w typowej komórce zwierzęcej. Zwykle dzieli się go na cztery etapy.
I. Profaza. Specjalna struktura komórkowa - centriola - podwaja się (czasami to podwojenie następuje w okresie S interfazy), a dwie centriole zaczynają się rozchodzić w kierunku przeciwnych biegunów jądra. Błona jądrowa ulega zniszczeniu; jednocześnie specjalne białka łączą się (agregują), tworząc mikrotubule w postaci nici. Centriole, obecnie zlokalizowane na przeciwległych biegunach komórki, organizują mikrotubule, które w rezultacie ustawiają się promieniście, tworząc strukturę przypominającą wyglądem kwiat asteru („gwiazdę”). Pozostałe nici mikrotubul rozciągają się od jednej centrioli do drugiej, tworząc tzw. wrzeciono rozszczepialne. W tym czasie chromosomy są w stanie spiralnym, przypominającym sprężynę. Są wyraźnie widoczne w mikroskopie świetlnym, szczególnie po zabarwieniu. W profazie chromosomy ulegają podziałowi, ale chromatydy nadal pozostają połączone parami w strefie centromeru - organelli chromosomalnej podobnej pod względem funkcji do centrioli. Centromery mają również wpływ organizujący na włókna wrzeciona, które teraz rozciągają się od centrioli do centromerów i od nich do innej centrioli.
II. Metafaza. Chromosomy, dotychczas ułożone losowo, zaczynają się poruszać, jakby ciągnięte przez nitki wrzeciona przyczepione do ich centromerów, i stopniowo ustawiają się w tej samej płaszczyźnie, w określonym położeniu i w równej odległości od obu biegunów. Centromery leżące w tej samej płaszczyźnie wraz z chromosomami tworzą tzw. płyta równikowa. Centromery łączące pary chromatyd dzielą się, po czym chromosomy siostrzane są całkowicie rozdzielane.
III. Anafaza. Chromosomy każdej pary poruszają się w przeciwnych kierunkach w kierunku biegunów, jakby były ciągnięte przez nitki wrzeciona. W tym przypadku nici powstają również pomiędzy centromerami sparowanych chromosomów.
IV. Telofaza. Gdy tylko chromosomy zbliżą się do przeciwnych biegunów, sama komórka zaczyna się dzielić wzdłuż płaszczyzny, w której znajdowała się płyta równikowa. W rezultacie powstają dwie komórki. Nici wrzeciona ulegają zniszczeniu, chromosomy rozwijają się i stają się niewidoczne, a wokół nich tworzy się błona jądrowa. Komórki powracają do fazy interfazy G1. Cały proces mitozy trwa około godziny.
Szczegóły mitozy różnią się nieco w zależności od typu komórek. Typowa komórka roślinna tworzy wrzeciono, ale brakuje mu centrioli. U grzybów mitoza zachodzi wewnątrz jądra, bez wcześniejszego rozpadu błony jądrowej.
Sam podział komórki, zwany cytokinezą, nie ma ścisłego związku z mitozą. Czasami zachodzi jedna lub więcej mitoz bez podziału komórki; W rezultacie powstają komórki wielojądrowe, często spotykane w algach. Jeśli jądro zostanie usunięte z jaja jeżowca poprzez mikromanipulację, wrzeciono nadal się tworzy, a jajo nadal się dzieli. To pokazuje, że obecność chromosomów nie jest warunkiem koniecznym podziału komórki.
Rozmnażanie przez mitozę nazywa się rozmnażaniem bezpłciowym, rozmnażaniem wegetatywnym lub klonowaniem. Jego najważniejszym aspektem jest genetyka: przy takiej reprodukcji nie ma rozbieżności czynników dziedzicznych u potomstwa. Powstałe komórki potomne są genetycznie dokładnie takie same jak komórka macierzysta. Mitoza to jedyny sposób samoreprodukcji u gatunków, które nie rozmnażają się płciowo, takich jak wiele organizmów jednokomórkowych. Jednak nawet u gatunków rozmnażających się płciowo komórki organizmu dzielą się w drodze mitozy i pochodzą z pojedynczej komórki, zapłodnionego jaja, a zatem wszystkie są genetycznie identyczne. Rośliny wyższe mogą rozmnażać się bezpłciowo (poprzez mitozę) przez sadzonki i wąsy (dobrze znanym przykładem są truskawki).
Rozmnażanie płciowe organizmów odbywa się za pomocą wyspecjalizowanych komórek, tzw. gamety - oocyty (jaja) i plemniki (plemniki). Gamety łączą się, tworząc jedną komórkę – zygotę. Każda gameta jest haploidalna, tj. ma jeden zestaw chromosomów. W zestawie wszystkie chromosomy są różne, ale każdy chromosom komórki jajowej odpowiada jednemu z chromosomów plemnika. Dlatego zygota zawiera już parę odpowiadających sobie chromosomów, które nazywa się homologicznymi. Chromosomy homologiczne są podobne, ponieważ mają te same geny lub ich warianty (allele), które determinują specyficzne cechy. Na przykład jeden ze sparowanych chromosomów może mieć gen kodujący grupę krwi A, a drugi może mieć wariant kodujący grupę krwi B. Chromosomy zygoty pochodzące z komórki jajowej są matczyne, a te pochodzące z plemnika są ojcowskie.
W wyniku powtarzających się podziałów mitotycznych z powstałej zygoty powstaje organizm wielokomórkowy lub liczne wolno żyjące komórki, jak to ma miejsce w przypadku pierwotniaków rozmnażających się płciowo i w glonach jednokomórkowych.
Podczas tworzenia gamet diploidalny zestaw chromosomów obecny w zygocie musi zostać zmniejszony o połowę. Gdyby tak się nie stało, w każdym pokoleniu fuzja gamet doprowadziłaby do podwojenia zestawu chromosomów. Redukcja do haploidalnej liczby chromosomów następuje w wyniku podziału redukcyjnego – tzw. mejoza, która jest odmianą mitozy.
Rozszczepienie i rekombinacja.
Osobliwością mejozy jest to, że podczas podziału komórki płytka równikowa jest tworzona przez pary homologicznych chromosomów, a nie przez zduplikowane pojedyncze chromosomy, jak w przypadku mitozy. Sparowane chromosomy, z których każdy pozostaje pojedynczy, rozchodzą się do przeciwnych biegunów komórki, komórka dzieli się, w wyniku czego komórki potomne otrzymują połowę zestawu chromosomów w porównaniu z zygotą.
Załóżmy na przykład, że zestaw haploidalny składa się z dwóch chromosomów. W zygocie (a zatem we wszystkich komórkach organizmu wytwarzających gamety) obecne są chromosomy matczyne A i B oraz chromosomy ojcowskie A” i B”. Podczas mejozy dzielą się w następujący sposób:
Najważniejszą rzeczą w tym przykładzie jest fakt, że gdy chromosomy się rozchodzą, niekoniecznie powstaje pierwotny zestaw matczyny i ojcowski, ale możliwa jest rekombinacja genów, jak w przypadku gamet AB” i A”B na powyższym schemacie.
Załóżmy teraz, że para chromosomów AA" zawiera dwa allele - A I B– gen określający grupy krwi A i B. Podobnie para chromosomów „BB” zawiera allele M I N inny gen determinujący grupy krwi M i N. Rozdzielenie tych alleli może przebiegać w następujący sposób:
Oczywiście powstałe gamety mogą zawierać dowolną z następujących kombinacji alleli dwóch genów: jestem, miliard, bm Lub jakiś.
Jeśli chromosomów jest więcej, pary alleli będą segregować niezależnie według tej samej zasady. Oznacza to, że te same zygoty mogą wytwarzać gamety z różnymi kombinacjami alleli genów i dawać początek różnym genotypom u potomstwa.
Podział mejotyczny.
Obydwa przykłady ilustrują zasadę mejozy. W rzeczywistości mejoza jest procesem znacznie bardziej złożonym, ponieważ obejmuje dwa kolejne podziały. Najważniejsze w mejozie jest to, że chromosomy podwajają się tylko raz, podczas gdy komórka dzieli się dwukrotnie, w wyniku czego liczba chromosomów zmniejsza się, a zestaw diploidalny zamienia się w haploidalny.
Podczas profazy pierwszego podziału chromosomy homologiczne ulegają koniugacji, czyli łączą się w pary. W wyniku tego bardzo precyzyjnego procesu każdy gen kończy się naprzeciw swojego homologu na innym chromosomie. Następnie oba chromosomy podwajają się, ale chromatydy pozostają połączone ze sobą wspólnym centromerem.
W metafazie cztery połączone chromatydy ustawiają się w jednej linii, tworząc płytkę równikową, jakby były jednym zduplikowanym chromosomem. W przeciwieństwie do tego, co dzieje się podczas mitozy, centromery nie dzielą się. W rezultacie każda komórka potomna otrzymuje parę chromatyd nadal połączonych centromerem. Podczas drugiego podziału chromosomy, już pojedyncze, ponownie ustawiają się w szeregu, tworząc niczym w przypadku mitozy płytkę równikową, ale podczas tego podziału nie następuje ich podwojenie. Centromery następnie dzielą się i każda komórka potomna otrzymuje jedną chromatydę.
Podział cytoplazmatyczny.
W wyniku dwóch podziałów mejotycznych komórki diploidalnej powstają cztery komórki. Kiedy tworzą się męskie komórki rozrodcze, otrzymuje się cztery plemniki mniej więcej tej samej wielkości. Kiedy powstają jaja, podział cytoplazmy zachodzi bardzo nierównomiernie: jedna komórka pozostaje duża, a pozostałe trzy są tak małe, że prawie w całości są zajęte przez jądro. Te małe komórki, tzw. ciała polarne służą jedynie do przyjmowania nadmiaru chromosomów powstałych w wyniku mejozy. Większość cytoplazmy niezbędnej dla zygoty pozostaje w jednej komórce - jaju.
Koniugacja i krzyżowanie.
Podczas koniugacji chromatydy homologicznych chromosomów mogą się rozbić, a następnie połączyć w nowym porządku, wymieniając sekcje w następujący sposób:

Ta wymiana odcinków homologicznych chromosomów nazywana jest krzyżowaniem. Jak pokazano powyżej, krzyżowanie prowadzi do pojawienia się nowych kombinacji alleli połączonych genów. Tak więc, jeśli oryginalne chromosomy miały kombinacje AB I ok, to po przekroczeniu będą zawierać Ab I aB. Ten mechanizm powstawania nowych kombinacji genów uzupełnia efekt niezależnego sortowania chromosomów, który zachodzi podczas mejozy. Różnica polega na tym, że krzyżowanie oddziela geny na tym samym chromosomie, podczas gdy sortowanie niezależne oddziela tylko geny na różnych chromosomach.
PRZEMIENNE POKOLENIA
PRYMITYWNE KOMÓRKI: PROKARYOTY
Wszystko to dotyczy komórek roślin, zwierząt, pierwotniaków i jednokomórkowych glonów, zwanych łącznie eukariontami. Eukarionty wyewoluowały z prostszej formy, prokariotów, które są obecnie reprezentowane przez bakterie, w tym archebakterie i cyjanobakterie (te ostatnie zwane wcześniej niebiesko-zielonymi algami). W porównaniu z komórkami eukariotycznymi komórki prokariotyczne są mniejsze i mają mniej organelli komórkowych. Mają błonę komórkową, ale brakuje im retikulum endoplazmatycznego, a rybosomy swobodnie unoszą się w cytoplazmie. Mitochondria są nieobecne, ale enzymy utleniające są zwykle przyłączone do błony komórkowej, która w ten sposób staje się odpowiednikiem mitochondriów. Prokariotom brakuje również chloroplastów, a chlorofil, jeśli jest obecny, występuje w postaci bardzo małych granulek.
Prokarioty nie mają jądra otoczonego błoną, chociaż lokalizację DNA można zidentyfikować na podstawie jego gęstości optycznej. Odpowiednikiem chromosomu jest nić DNA, zwykle okrągła, do której przyłączonych jest znacznie mniej białek. Łańcuch DNA jest przyłączony do błony komórkowej w jednym punkcie. U prokariotów nie ma mitozy. Zastępuje go następujący proces: DNA podwaja się, po czym błona komórkowa zaczyna rosnąć pomiędzy sąsiadującymi punktami połączenia dwóch kopii cząsteczki DNA, które w rezultacie stopniowo się rozchodzą. Komórka ostatecznie dzieli się pomiędzy punktami przyłączenia cząsteczek DNA, tworząc dwie komórki, każda z własną kopią DNA.
RÓŻNICOWANIE KOMÓREK
Rośliny i zwierzęta wielokomórkowe wyewoluowały z organizmów jednokomórkowych, których komórki po podziale pozostały razem, tworząc kolonię. Początkowo wszystkie komórki były identyczne, jednak dalsza ewolucja doprowadziła do zróżnicowania. Przede wszystkim różnicowały się komórki somatyczne (tj. komórki organizmu) i komórki rozrodcze. Dalsze różnicowanie stało się bardziej skomplikowane – powstawało coraz więcej różnych typów komórek. Ontogeneza – indywidualny rozwój organizmu wielokomórkowego – powtarza w ogólnym ujęciu ten proces ewolucyjny (filogeneza).
Fizjologicznie komórki różnicują się częściowo poprzez wzmocnienie jednej lub drugiej cechy wspólnej dla wszystkich komórek. Na przykład funkcja skurczowa ulega wzmocnieniu w komórkach mięśniowych, co może być wynikiem ulepszenia mechanizmu wykonującego ruch ameboidalny lub inne rodzaje ruchu w mniej wyspecjalizowanych komórkach. Podobnym przykładem są cienkościenne komórki korzeniowe wraz z ich wyrostkami, tzw. włośniki, które służą do wchłaniania soli i wody; w takim czy innym stopniu funkcja ta jest nieodłączna dla wszystkich komórek. Czasem specjalizacja wiąże się z nabyciem nowych struktur i funkcji – przykładem jest rozwój narządu ruchu (wici) w plemniku.
Szczegółowo zbadano różnicowanie na poziomie komórkowym lub tkankowym. Wiemy na przykład, że czasami zachodzi to autonomicznie, tj. jeden typ komórki może przekształcić się w inny, niezależnie od rodzaju komórek sąsiednich komórek. Jednak tzw Indukcja embrionalna to zjawisko, w którym jeden rodzaj tkanki stymuluje komórki innego typu do różnicowania się w określonym kierunku.
W ogólnym przypadku różnicowanie jest nieodwracalne, tj. wysoce zróżnicowane komórki nie mogą przekształcić się w inny typ komórek. Jednak nie zawsze tak jest, szczególnie w komórkach roślinnych.
Różnice w strukturze i funkcji ostatecznie zależą od rodzaju białek syntetyzowanych w komórce. Ponieważ synteza białek jest kontrolowana przez geny, a zestaw genów jest taki sam we wszystkich komórkach organizmu, różnicowanie musi zależeć od aktywacji lub inaktywacji pewnych genów w różnych typach komórek. Regulacja aktywności genów zachodzi na poziomie transkrypcji, tj. tworzenie informacyjnego RNA przy użyciu DNA jako matrycy. Tylko transkrybowane geny wytwarzają białka. Syntetyzowane białka mogą blokować transkrypcję, ale czasami także ją aktywują. Ponadto, ponieważ białka są produktami genów, niektóre geny mogą kontrolować transkrypcję innych genów. W regulacji transkrypcji biorą także udział hormony, w szczególności steroidy. Bardzo aktywne geny można powielać (podwajać) wiele razy, aby wytworzyć więcej informacyjnego RNA.
Rozwój nowotworów złośliwych często uważa się za szczególny przypadek różnicowania komórkowego. Jednak pojawienie się komórek złośliwych jest wynikiem zmian w strukturze DNA (mutacji), a nie procesów transkrypcji i translacji na białko normalnego DNA.
METODY BADANIA KOMÓREK
Mikroskop świetlny.
Pierwszym narzędziem do badania formy i struktury komórek był mikroskop świetlny. Jego zdolność rozdzielcza jest ograniczona wymiarami porównywalnymi z długością fali światła (0,4–0,7 μm dla światła widzialnego). Jednak wiele elementów struktury komórkowej ma znacznie mniejsze rozmiary.
Inną trudnością jest to, że większość składników komórkowych jest przezroczysta i ma współczynnik załamania światła prawie taki sam jak woda. Aby poprawić widoczność, często stosuje się barwniki, które mają różne powinowactwo do różnych składników komórkowych. Barwienie służy również do badania chemii komórki. Na przykład niektóre barwniki wiążą się preferencyjnie z kwasami nukleinowymi i w ten sposób ujawniają ich lokalizację w komórce. Niewielką część barwników — zwanych barwnikami przyżyciowymi — można zastosować do barwienia żywych komórek, ale zazwyczaj komórki należy najpierw utrwalić (przy użyciu substancji koagulujących białka), zanim będzie można je zabarwić. Cm. HISTOLOGIA.
Przed badaniem komórki lub kawałki tkanki są zwykle zatapiane w parafinie lub plastiku, a następnie cięte na bardzo cienkie skrawki za pomocą mikrotomu. Metoda ta jest szeroko stosowana w laboratoriach klinicznych do identyfikacji komórek nowotworowych. Oprócz konwencjonalnej mikroskopii świetlnej opracowano inne optyczne metody badania komórek: mikroskopię fluorescencyjną, mikroskopię z kontrastem fazowym, spektroskopię i analizę dyfrakcji promieni rentgenowskich.
Mikroskop elektronowy.
Mikroskop elektronowy ma rozdzielczość ok. 1–2 nm. To wystarczy do badania dużych cząsteczek białka. Zwykle konieczne jest pokolorowanie i kontrastowanie obiektu solami metali lub metalami. Z tego powodu, a także dlatego, że obiekty bada się w próżni, za pomocą mikroskopu elektronowego można badać jedynie martwe komórki.
Autoriografia.
Jeśli do pożywki zostanie dodany izotop promieniotwórczy, który jest absorbowany przez komórki podczas metabolizmu, można następnie wykryć jego wewnątrzkomórkową lokalizację za pomocą autoradiografii. Dzięki tej metodzie cienkie skrawki komórek umieszcza się na folii. Film ciemnieje pod miejscami, w których znajdują się izotopy promieniotwórcze.
Wirowanie.
Do badań biochemicznych składników komórkowych komórki muszą zostać zniszczone – mechanicznie, chemicznie lub ultradźwiękowo. Uwolnione składniki zawieszone są w cieczy i można je wyizolować i oczyścić poprzez wirowanie (najczęściej w gradiencie gęstości). Zazwyczaj takie oczyszczone składniki zachowują wysoką aktywność biochemiczną.
Hodowle komórkowe.
Niektóre tkanki można podzielić na pojedyncze komórki, dzięki czemu komórki pozostają żywe i często są w stanie się rozmnażać. Fakt ten definitywnie potwierdza ideę komórki jako żywej jednostki. Gąbkę, prymitywny organizm wielokomórkowy, można rozdzielić na komórki poprzez przetarcie jej przez sito. Po pewnym czasie komórki te łączą się ponownie i tworzą gąbkę. Tkanki embrionalne zwierząt można doprowadzić do dysocjacji za pomocą enzymów lub innych środków osłabiających wiązania między komórkami.
Amerykański embriolog R. Harrison (1879–1959) jako pierwszy wykazał, że komórki embrionalne, a nawet niektóre dojrzałe, w odpowiednim środowisku, mogą rosnąć i rozmnażać się poza organizmem. Technikę tę, zwaną hodowlą komórkową, udoskonalił francuski biolog A. Carrel (1873–1959). Komórki roślinne można również hodować w hodowli, ale w porównaniu z komórkami zwierzęcymi tworzą one większe skupiska i są mocniej ze sobą połączone, dlatego w miarę wzrostu kultury tworzą się tkanki, a nie pojedyncze komórki. W hodowli komórkowej z pojedynczej komórki można wyhodować całą dorosłą roślinę, np. marchewkę.
Mikrochirurgia.
Za pomocą mikromanipulatora można usunąć, dodać lub w jakiś sposób zmodyfikować poszczególne części komórki. Dużą komórkę ameby można podzielić na trzy główne elementy – błonę komórkową, cytoplazmę i jądro, a następnie elementy te można ponownie złożyć, tworząc żywą komórkę. W ten sposób można uzyskać sztuczne komórki składające się ze składników różnych typów ameb.
Jeśli weźmiemy pod uwagę, że sztuczna synteza niektórych składników komórkowych wydaje się możliwa, to eksperymenty ze składaniem sztucznych komórek mogą być pierwszym krokiem w kierunku stworzenia nowych form życia w laboratorium. Ponieważ każdy organizm rozwija się z pojedynczej komórki, metoda wytwarzania sztucznych komórek w zasadzie pozwala na konstruowanie organizmów danego typu, jeśli jednocześnie użyje się komponentów nieco różniących się od tych, które znajdują się w istniejących komórkach. W rzeczywistości jednak nie jest wymagana pełna synteza wszystkich składników komórkowych. Strukturę większości, jeśli nie wszystkich, składników komórki określają kwasy nukleinowe. Zatem problem powstawania nowych organizmów sprowadza się do syntezy nowych typów kwasów nukleinowych i zastąpienia ich naturalnymi kwasami nukleinowymi w określonych komórkach.
Fuzja komórkowa.
Inny rodzaj sztucznych komórek można otrzymać poprzez fuzję komórek tego samego lub różnych gatunków. Aby osiągnąć fuzję, komórki poddaje się działaniu enzymów wirusowych; w tym przypadku zewnętrzne powierzchnie dwóch komórek są sklejane ze sobą, a błona między nimi ulega zniszczeniu i powstaje komórka, w której dwa zestawy chromosomów są zamknięte w jednym jądrze. Możliwe jest łączenie komórek różnych typów lub znajdujących się na różnych etapach podziału. Metodą tą udało się uzyskać komórki hybrydowe myszy i kurczaka, człowieka i myszy oraz człowieka i ropuchy. Komórki takie tylko początkowo są hybrydowe, a po licznych podziałach komórkowych tracą większość chromosomów jednego lub drugiego typu. Produktem końcowym staje się na przykład zasadniczo komórka myszy, która nie zawiera żadnych ludzkich genów lub zawiera je tylko w śladowych ilościach. Szczególnie interesująca jest fuzja komórek normalnych i złośliwych. W niektórych przypadkach hybrydy stają się złośliwe, w innych nie, tj. obie właściwości mogą objawiać się zarówno jako dominujące, jak i recesywne. Wynik ten nie jest nieoczekiwany, ponieważ nowotwór złośliwy może być spowodowany różnymi czynnikami i ma złożony mechanizm.
Literatura:
Ham A., Cormack D. Histologia, t. 1. M., 1982
Alberts B., Bray D., Lewis J., Raff M., Roberts K., Watson J. Biologia molekularna komórki, t. 1. M., 1994







