∙ niepłodność mieszana (połączenie form niepłodności żeńskiej i męskiej). PRZECIWWSKAZANIA
∙ choroby somatyczne i psychiczne stanowiące przeciwwskazanie do ciąży;
∙ wady wrodzone: powtarzające się porody dzieci z tym samym rodzajem wad rozwojowych; poprzednie narodziny dziecka z nieprawidłowościami chromosomowymi; choroby dziedziczne dominująco u jednego z rodziców wysoki stopień penetracja;
∙ choroby dziedziczne: heterozygotyczne nosicielstwo u małżonków wszelkich chorób monogenowych (zaburzenia metabolizmu aminokwasów, węglowodanów, glikolipidów, glikoprotein). wcześniejsze urodzenie dzieci z chorobami dziedzicznymi i związanymi z płcią (hemofilia, miopatia typu Duchenne’a itp.);
∙ stany przerostowe macicy i jajników;
∙ wady rozwojowe i anomalie macicy;
∙ nieuleczalne schorzenia kanału szyjki macicy.
Eksperci zalecają rozpoczęcie badania małżeństwa od mężczyzny, ponieważ badanie nasienia od razu wykaże przyczynę niepłodności męskiej, a zdiagnozowanie niepłodności kobiecej jest sprawą złożoną i długotrwałą. Aby spermogram miał charakter informacyjny, przed oddaniem nasienia do analizy należy powstrzymać się od aktywności seksualnej na 3-5 dni (najlepiej nie mniej, ale nie więcej). Nasienie do analizy najlepiej oddać w tym samym pomieszczeniu, w którym znajduje się laboratorium. Ochłodzenie plemnika prowadzi do zniekształcenia większości wskaźników jego jakości.
Kolejnym etapem egzaminu jest test zgodności. Niezgodność może mieć charakter immunologiczny i biologiczny. Określa szyjkowy czynnik niepłodności: śluz szyjkowy w przypadku niezgodności zmniejsza chemotaksję lub „zabija” plemniki. Następnie kobieta jest badana w celu zdiagnozowania niepłodności u kobiet. Po postawieniu diagnozy i wyjaśnieniu przyczyn niepłodności z reguły przystępują do samego procesu leczenia.
LECZENIE METODĄ IVF.
Po pierwsze, za pomocą hormonów konieczne jest osiągnięcie dojrzewania kilku jaj w jajnikach na raz (superowulacja). Głównymi lekami pierwszego etapu są agoniści hormonu uwalniającego gonadotropinę (α-HLH), leki zawierające ludzką gonadotropinę menopauzalną (HMG) i ludzką gonadotropinę kosmówkową (hCG). Podaje się je według opracowanych schematów leczenia, czyli „protokołów stymulacji superowulacji”. Proces dojrzewania monitoruje się za pomocą ultradźwięków i oznaczania poziomu hormonów (estradiolu).
Na krótko przed samoistnym procesem owulacji (uwolnieniem komórki jajowej z jajnika) wykonuje się nakłucie pęcherzyka i aspirację komórki jajowej. Bardzo ważne jest określenie momentu, w którym należy nakłuć (pobrać) pęcherzyki (jak najbliżej czasu naturalnej owulacji), co
wykonuje się za pomocą ultradźwięków i oznaczania stężenia hormonów w surowicy krwi.
Nakłucie przezpochwowe wykonuje się pod kontrolą USG po 36 godzinach od podania gonadotropiny kosmówkowej za pomocą specjalnych igieł do nakłuwania.
Nakłucie przezpochwowe wykonuje się na sali operacyjnej wyposażonej we wszystkie niezbędne instrumenty i sprzęt do leczenia chirurgicznego w nagłych przypadkach (respirator i inne). Środki przeciwbólowe stosuje się w zależności od stanu kobiety. Pobieranie, czyli aspiracja pęcherzyków, odbywa się z obu jajników.
Pobranie i przygotowanie plemników. Aby przygotować je do nawożenia przeprowadza się tzw. kapacytację, czyli tzw. przemywanie plemników z elementów osocza, a następnie specjalnymi metodami przygotowanie roztworu z żywymi plemnikami.
Po około 5-7 godzinach przebywania w pożywce, komórki jajowe i plemniki łączy się (inseminacja komórki jajowej) w „probówce” i umieszcza w inkubatorze na 24-42 godziny. Za dzień nakłucia uważa się dzień zerowy hodowli zarodków (0D); Za pierwszy dzień uprawy (1D) uważa się dzień następujący po nakłuciu. To właśnie tego dnia u większości zauważalne są pierwsze oznaki zapłodnienia. Pojawiają się, jak wspomniano powyżej, 16 - 18 godzin po zmieszaniu jaj z nasieniem (inseminacja). Zapłodnienie ocenia się ponownie po 24–26 godzinach od inseminacji. Kontrolę zapłodnienia oocytów przeprowadza asystent laboratoryjny-embriolog podczas oglądania naczyń z hodowanymi komórkami pod mikroskopem. Jednak ich obecność nie jest jeszcze wystarczająca, aby rozstrzygnąć kwestię możliwości przeniesienia zarodków do jamy macicy. Najpierw musisz upewnić się, że zarodki dzielą się i rozwijają prawidłowo. Można to ocenić jedynie na podstawie ilości i jakości dzielących się komórek zarodka i nie wcześniej niż dzień po zapłodnieniu, kiedy pojawią się pierwsze oznaki fragmentacji. Najwyraźniej pojawiają się dopiero w drugim dniu uprawy (2D). Można przenosić wyłącznie zarodki dobrej jakości. Transfer zarodków przeprowadza się najczęściej w 2. lub 3. dniu hodowli, w zależności od tempa ich rozwoju i jakości zarodków.
Przeniesienie zarodków do jamy macicy odbywa się za pomocą specjalnych cewników minimalna ilość pożywka(20-30 µl). Zaleca się przeniesienie do jamy macicy nie więcej niż 3–4 zarodków, gdyż przy przenoszeniu większej liczby zarodków możliwa jest implantacja dwóch lub większej liczby zarodków. Przeniesienie zarodków do macicy matki odbywa się zwykle bez znieczulenia. Następnie należy przepisać kobiecie leki wspomagające implantację i rozwój zarodków. Rozpoznanie ciąży przeprowadza się od dziesiątego dnia po transferze sztucznie zapłodnionych zarodków. Kobiety, które zajdą w ciążę po zastosowaniu metody IVF i ET, zaliczane są do grupy wysokiego ryzyka
musi znajdować się pod stałą kontrolą lekarza-położnika-ginekologa. Po transferze zarodka kobieta otrzymuje zwolnienie lekarskie z diagnozą „Wczesna ciąża, zagrożenie poronieniem”.
Skuteczność zapłodnienia in vitro wynosi obecnie średnio 20-30%,
jednak w niektórych ośrodkach przekracza 50%. To bardzo wysoki odsetek, zwłaszcza jeśli weźmiemy pod uwagę, że prawdopodobieństwo zapłodnienia w cyklu naturalnym u zupełnie zdrowego mężczyzny i kobiety w jednym cyklu kopulacyjnym nie przekracza 30%. Oto medyczne aspekty tego problemu. Z zapłodnieniem in vitro wiążą się także problemy moralne, etyczne i religijne. W szczególności wiele wyznań religijnych zabrania wierzącym uciekania się do zapłodnienia in vitro, uznając tę metodę porodu za grzeszną. W podstawach legislacji Federacja Rosyjska z dnia 22 lipca 1993 r. (ust. 7, art. 35 „Sztuczna inseminacja i implantacja zarodka”) stanowi, że każda pełnoletnia kobieta ma prawo do sztucznego zapłodnienia i implantacji zarodka.
IVF umożliwia przeprowadzenie diagnostyki przedimplantacyjnej dziedzicznych (genetycznych) chorób przyszłego płodu, zanim zarodek zostanie przeniesiony do macicy, czyli zanim zajdzie w ciążę. Możesz świadomie podejść do rozwiązania ważnego problemu, jeśli w Twojej rodzinie są dzieci cierpiące na choroby genetyczne. Diagnostykę przedimplantacyjną aberracji chromosomowych przeprowadza się metodą FISH, PCR lub cytogenetyką.
Zapłodnienie in vitro jest również szeroko stosowane do rozwiązywania problemów z zakresu podstawowej biologii i medycyny.
5.5 Klonowanie organizmów, narządów i tkanek. Problem klonowania zyskał w ostatnim czasie ostry oddźwięk społeczny, gdyż media często nieudolnie przedstawiają istotę zagadnienia.
Zgodnie z definicją przyjętą w genetyce, klonowanie to dokładne odtworzenie żywego obiektu. Głównym kryterium klonu jest tożsamość genetyczna. Klonowanie jest szeroko stosowane w uprawie roślin, przemyśle mikrobiologicznym i embriologii eksperymentalnej. Znane są przypadki naturalnego klonowania u ludzi – są to bliźnięta jednojajowe. Jednak obecnie mówimy o uzyskaniu dokładnych kopii dorosłego zwierzęcia lub osoby o szczególnie cennych cechach.
Teoria klonowania opiera się na doświadczeniach J. Gurdona, który przeszczepił jądra komórek nabłonka powłokowego do jądrzastych jaj żab i uzyskał z nich kijanki. W maju 1997 roku Ian Wilmut ze Szkocji opublikował wyniki klonowania owiec (słynnej Dolly). Pojawiły się także publikacje wyraźnie spekulacyjne na temat udanych prób klonowania ludzi.
Naukowa analiza przedstawionych danych wykazała, że nie mówi się jeszcze o skutecznym klonowaniu zwierząt i ludzi.
Po pierwsze, praktyczna wydajność klonowania wynosi 1-2%, po drugie, nie udowodniono tożsamości genetycznej klonowanych organizmów, a po trzecie, żywotność i funkcjonalność „klonów” okazała się nieporównywalnie niższa niż ich naturalnych odpowiedników.
Są jeszcze inne powody, dla których na obecnym poziomie rozwoju nauki masowe klonowanie ssaków i ludzi nie jest możliwe. Istnieją także problemy społeczne i etyczne związane z klonowaniem, które prawdopodobnie nie zostaną rozwiązane w najbliższej przyszłości.
Na zupełnie innym planie leży problem klonowania narządów i tkanek zwierząt i ludzi w celu przeszczepienia. Jest to naprawdę obiecujące i praktycznie znaczące zadanie, które zostało pomyślnie rozwiązane. Udowodniono, że przeszczepienie klonu własnych komórek pacjenta lub wcześniej wyhodowanej tkanki (narządu) jest lepsze od materiału dawcy: znikają problemy związane z niezgodnością immunologiczną, zwiększa się dokładność dawki przeszczepu, możliwe jest tworzenie banków komórek, tkanek i narządach, pojawiają się wyjątkowe możliwości badań eksperymentalnych, znikają kwestie etyczne, problemy itp.
LITERATURA
1. Asanov A.Yu., Demikova N.S., Morozov S.A. Podstawy genetyki i dziedzicznych zaburzeń rozwoju u dzieci. M.: Centrum Wydawnicze
"Akademia". 2003. – 224 s.
2. Baranow V.S. Diagnostyka prenatalna chorób dziedzicznych i wrodzonych w Rosji. – Magazyn edukacyjny Sorosa. – 1998 r. – nr 10. -
3. Baranow V.S. Terapia genowa to medycyna XXI wieku. – Magazyn edukacyjny Sorosa. – 1999 r. – nr 3. - Z. 63-68.
4. Baranov V.S., Baranova E.V., Ivashchenko T.E., Aseev M.V. Genom człowieka i geny „podatności”. Wprowadzenie do medycyny predykcyjnej. Petersburg: „Intermedica”. 2000. – 271 s.
5. Barashnev Yu.I., Bakharev V.A., Novikov P.V. Diagnostyka i leczenie chorób wrodzonych i dziedzicznych u dzieci (przewodnik po genetyce klinicznej). M.:„Triada-X”. 2004. – 560 s.
6. Boczkow N.P. Genetyka kliniczna. - M.: GEOTAR-MED., 2001. – 448 s.
7. Wakharłowski V.G., Romanenko O.P., Gorbunova V.N. Genetyka w praktyce pediatrycznej. SPb.: „Feniks”. 2009. – 288 s.
8. Ginter E.K. Genetyka medyczna. – M.: Medycyna. – 2003. – 448 s.
9. Gorbunova V.N. Podstawa molekularna genetyka medyczna. – Petersburg: Intermedica. – 1999. – 212 s.
10.Gorbunova V.N., Baranov V.S. Wprowadzenie do diagnostyki molekularnej i terapii genowej chorób dziedzicznych. – Petersburg: literatura specjalna. – 1997. – 287 s.
11. Zayats R.G., Butvilovsky V.E., Rachkovskaya I.V., Davydov V.V. Genetyka ogólna i medyczna. Rostów nad Donem: „Feniks”. 2002. – 320 s.
12.Illarioshkin S.N., Ivanova-Smolenskaya I.A., Markova E.D. Diagnostyka DNA i medyczne poradnictwo genetyczne w neurologii. M.: Agencja informacji medycznej. 2002. – 591 s.
13. Kozlova S.I., Demikova N.S., Semanova E., Blinnikova O.E. Zespoły dziedziczne i medyczne poradnictwo genetyczne. - M.:
Ćwiczyć. – 1996. – 415 s.
Podręcznik edukacyjno-metodologiczny. – 1991. – 95 s.
16. Lilin E.T., Bogomazov E.A., Goffman-Kadoshnikov P.B. Genetyka dla lekarzy.
– M.: Medycyna. – 1990. – 312 s.
17. Lewin B. Geny. – M.: Mir. – 1987. – 647 s.
18.Mutovin G.R. Podstawy genetyki klinicznej. – Wyżej szkoła, 2001. – 234 s. 19.Murphy EA, Chase G.A. Podstawy medycznego poradnictwa genetycznego. M.:
Medycyna, 1979.
20. Prikhodchenko N.N., Shkurat T.P. Podstawy genetyki człowieka. – Rostów nad-
Don: Feniks. – 1997. – 368 s.
21.Prozorova M.V. Medyczne poradnictwo genetyczne w zakresie chorób chromosomowych i ich diagnostyki prenatalnej. – Petersburg: MAPO. – 1997. - 15.
22.Prozorova M.V. Choroby chromosomalne. – Petersburg: MAPO. – 1997. – 23 s. 23. Puzyrev V.P. Badania genomiczne i choroby człowieka. - Sorosowski
magazyn edukacyjny. – 1996 r. – nr 5. – s. 19-27.
24. Puzyrev V.P., Spepanov V.A. Anatomia patologiczna genomu człowieka. – Nowosybirsk: Nauka. – 1997. – 224 s.
25. Repin V.S., Sukhikh G.T. Biologia komórki medycznej. – M.: BEBiM. – 1998. – 200 s.
26. Singer M., Berg P. Geny i genomy. – M.: Mir. – 1998. – T.1. – 373 s.
27.Soifer V.N. Międzynarodowy projekt poznania ludzkiego genomu. – | Sorosowski |
|
magazyn edukacyjny. – 1996. - | Nr 12. – s. 4-11. | |
28.Teratologia człowieka. wyd. 2. – | wyd. G.I. Łazyuk. - M.: | Medycyna. – |
29. Fovorova O.O. Leczenie genami – | fantazja lub rzeczywistość. – | Sorosowski |
magazyn edukacyjny. – 1997. - | Nr 2. – s. 21-27. | |
30.Fogel F., Motulski A. Genetyka Człowieka, T.1. – M.: Mir. – 1989. – 312 s. 31.Shabalov N.P. Choroby dziecięce, T.2. – SPb: Piotr. 2004. – 736 s. 32. Szewczenko V.A., Topornina N.A., Stvolinskaya N.S. Genetyka człowieka. M.:
Humanitarny. wyd. Centrum VLADOS. 2002. – 240 s.
1.4 Metody badania genetyki i diagnostyki człowieka
choroby dziedziczne | ||
Choroby chromosomalne | ||
Zespoły spowodowane nieprawidłowościami w układzie autosomalnym | ||
Choroba Downa | ||
Zespół Edwardsa | ||
Zespół Pataua | ||
Syndrom płaczu kota | ||
Zespół Lejeune’a | ||
Przewlekła białaczka szpikowa | ||
Zespół trisomii 6q | ||
Syndrom Retora | ||
Zespół trisomii 11q |
2.2 Zespoły kliniczne spowodowane anomaliami
w układzie chromosomów płciowych | |||
Zespół Szereszewskiego-Turnera | |||
Zespół Klinefeltera | |||
Zespół Trisomii X | |||
Syndrom 47, XUU | |||
Choroby genowe | |||
Fenyloketonuria | |||
Galaktozemia | |||
Zespół nadnerczowo-płciowy | |||
Mukowiscydoza | |||
zespół Marfana | |||
Dystrofinopatie | |||
Zapobieganie dziedzicznej patologii | |||
Medyczne poradnictwo genetyczne | |||
Profilaktyka perikoncepcyjna | |||
Profilaktyka przedimplantacyjna | |||
Monitorowanie prenatalne | |||
Biopsja kosmówki kosmówki | |||
Amniocenteza | |||
Kordocenteza | |||
Perspektywy genetyki | |||
Diagnostyka DNA | |||
LEKCJA PRAKTYCZNA 1
TEMAT: Cytologiczne podstawy dziedziczności.
Biochemiczne podstawy dziedziczności.
Dziedziczna zmienność.
Czas trwania lekcji – 270 minut
CEL: Nauczyć się:
Analizuj mikrofotografie i diagramy: 1) fazy mitozy i mejozy, 2) etapy gametogenezy.
Modelowe procesy wdrażania Informacja genetyczna: transkrypcje, tłumaczenia.
Przeanalizuj: konsekwencje rozregulowania mitozy i mutacji genowych oraz przyczyny je powodujące.
W wyniku przestudiowania tematu student powinien:
móc:
przeprowadzać ankiety i prowadzić dokumentację pacjentów z dziedzicznymi patologiami
wiedzieć:
biochemiczne i cytologiczne podstawy dziedziczności.
główne typy zmienności, rodzaje mutacji u człowieka, czynniki mutagenezy;
Wygenerowane kompetencje ogólne:
OK1. Zrozum istotę i znaczenie społeczne jego przyszły zawód, okazuj jej stałe zainteresowanie.
OK2. Organizuj własne działania, wybieraj standardowe metody i sposoby wykonywania zadań zawodowych, oceniaj ich skuteczność i jakość.
OK3. Podejmuj decyzje w sytuacjach standardowych i niestandardowych i bierz za nie odpowiedzialność.
OK4. Wyszukuje i wykorzystuje informacje niezbędne do skutecznego wykonywania powierzonych mu zadań zawodowych, a także zawodowych i zawodowych rozwój osobisty.
OK6. Pracuj w zespole i zespole, skutecznie komunikuj się
ze współpracownikami, zarządem i konsumentami.
OK7. Brać odpowiedzialność za pracę członków zespołu (podwładnych) i wyniki realizacji zadań.
OK8. Samodzielnie wyznaczają zadania rozwoju zawodowego i osobistego, angażują się w samokształcenie, świadomie planują i przeprowadzają zaawansowane szkolenia.
OK12. Zorganizować Miejsce pracy zgodnie z wymogami ochrony pracy, higieny przemysłowej, infekcji i bezpieczeństwa przeciwpożarowego.
OK13. Aktualności zdrowy wizerunekżycie, angażuj się Kultura fizyczna i sportu dla poprawy zdrowia, osiągnięcia celów życiowych i zawodowych.
Wyposażenie metodyczne lekcji :
TCO: Laptop do pokazów slajdów
Rozdawać:
Opracowanie metodologiczne lekcji praktycznej dla uczniów.
Literatura do przygotowania:
Główny:
Khandogina K.I. Genetyka człowieka z podstawami genetyki medycznej: podręcznik. – M.: GEOTAR-Media, 2013. – 176 s.: z ilustr.
Dodatkowy :
Bochkov N.A. itp. Genetyka medyczna: podręcznik4 - M.: ACADEMA, 2003
Atlas chromosomów ludzkich - Moskwa, 1982
E.K. Timolyanova Genetyka medyczna Rostów nad Donem: Phoenix, 2003.
N.N. Prichodczenko, T.P. Shkurat Podstawy genetyki człowieka - Rostów nad Donem: Phoenix, 1997.
VA Orekhova, T.A. Łaszkowska, M.P. Szkoła wyższa Sheibach Medical Genetics w Mińsku 1999.
NS Demidova, O.E. Blinnikova Zespoły dziedziczne i poradnictwo medyczno-genetyczne Medycyna Leningradzka 1987.
Źródła internetowe:
1. Doradca studencki – Biblioteka Cyfrowa studia medycznewww/ medkollegelib. ru
Plan lekcji
Część wprowadzająca– 26 minut
Organizacja czasu;
Motywacja do lekcji;
Kontrola początkowego poziomu wiedzy.
Część główna – 230 minut
Badanie etapów mitozy;
Badanie etapów mejozy;
Samodzielna praca studentów nad analizą mitozy;
Badanie etapów gametogenezy;
Samodzielna praca studentów nad analizą gametogenezy;
Badanie wzorców implementacji informacji genetycznej na poziomie biochemicznym.
Samodzielna praca mająca na celu rozwój umiejętności modelowania i analizy procesów replikacji, transkrypcji i biosyntezy białek
Badanie zmienności dziedzicznej
Samodzielna praca rozwijająca umiejętności analizy zmienności dziedzicznej.
Część końcowa – 14 minut
Zreasumowanie;
Praca domowa.
Postęp lekcji
Część wprowadzająca
Znaczenie
Wzrost i rozwój są związane z reprodukcją komórek (proliferacją) organizm wielokomórkowy, procesy regeneracyjne. Zaburzenia mitozy leżą u podstaw występowania mutacji somatycznych – przyczyny nowotworów.
Zaburzenia mejozy (tworzenie się komórek rozrodczych) determinują występowanie mutacji generatywnych, które klinicznie objawiają się w postaci chorób dziedzicznych. Niedysjunkcja chromosomów jest przyczyną mutacji genomowych
Wśród szerokiej gamy składników molekularnych komórek zapewniających ich funkcjonowanie, główną rolę w zachowaniu i przekazywaniu informacji genetycznej należy do kwasów nukleinowych. Naruszenie struktury kwasy nukleinowe może prowadzić do zmian patologicznych w komórce – mutacji genowych.
Kontrola przychodząca
Co to jest chromatyna, chromosom, chromatyda?
Jakie znasz rodzaje podziałów komórkowych?
Co to jest interfaza?
Definicja amitoza.
Zdefiniuj mitozę.
Nazwij fazy mitozy.
Na jakich etapach mitozy chromosomy są wyraźnie widoczne?
Jakie choroby powstają w wyniku zakłócenia procesu mitozy?
Zdefiniuj mejozę.
Jaki zestaw chromosomów zawierają komórki płciowe?
Co to jest spermatogeneza i oogeneza?
Wymień okresy gametogenezy.
W jakich okresach zachodzi mitoza, a w jakich okresach mejoza?
Jak klasyfikuje się zmienność dziedziczną?
Jakie są przyczyny mutacji genów?
Jakie są przyczyny mutacji genomowych?
Głównym elementem
Badanie i analiza mikrofotografii, rysunków: 1) rodzaje podziałów komórkowych, fazy mitozy i mejozy, 2) etapyrozwój gametogenezy człowieka.
Zadanie 1. Cykl komórkowy (CC)
Spójrz na rysunekCC i odpowiadaj na pytania
Co składa się na cykl życiowy komórki (ryc. 1)?
Wymień 4 okresyCC;
Omów i wyciągnij wnioski na temat znaczenia punktu krytycznego na diagramie
Jaką strukturę mają chromosomy w różnych okresach?CC(ryc. 2)?
Dlaczego interfaza zajmuje co najmniej 90% czasuCC?
Zapisz w swoim pamiętniku, co dzieje się w okresie presyntetycznym (G1), syntetyczne (S), postsyntetyczny (G2).
Przeczytaj tekst „Czas trwania CC w komórkach różnych tkanek”. Formułuj wnioski.
„Czas trwania CC w komórkach różnych tkanek”
„Cykle komórkowe (CC) w tym samym organizmie mają różny czas trwania, w zależności od tkanki. Na przykład u ludzi czas trwania CC wynosi: dla leukocytów 3-5 dni, nabłonka skóry -20-25 dni, nabłonka rogówki -2-3 dni, komórek szpiku kostnego 8-12 godzin i komórki nerwowe Z reguły żyją tak długo, jak osoba bez ukończenia CC (G1)”.
Rysunek1 . Cykl komórkowy (życia).
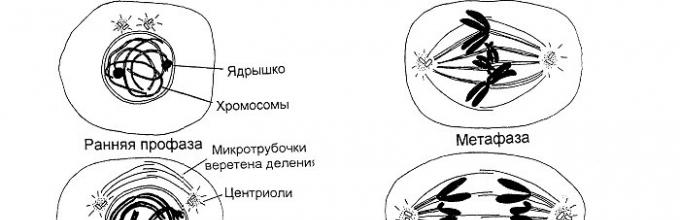
Zadanie 2. Mitoza. Badanie i analiza mikrofotografii, rysunków faz mitozy.
Rozważ schemat mitozy i odpowiedz na pytania (ryc. 2)
Dlaczego etapy profazy, metafazy, anafazy, telofazy dzieli się na wczesne i późne?
Jaki jest kluczowy proces profazy?
Jaki jest kluczowy proces metafazy?
Jaki jest kluczowy proces anafazy?

Rysunek2 . Fazy mitozy

Rysunek3 . Etapy mitozy (mitozę przedstawiono schematycznie na ryc. 4)
2 . Przyjrzyj się mikrofotografiom mitozy (ryc. 4) i odpowiedz na pytania:
Jaka jest różnica między schematycznym przedstawieniem mitozy a jej mikrofotografią?
Który okres mitozy można zidentyfikować jako najbardziej podatny na równomierne rozmieszczenie materiału genetycznego?

Rysunek4 . Mikrofotografia mitozy. Proces ten pokazano pod mikroskopem fluorescencyjnym. DNA świeci na niebiesko, a tubulina (a zatem i mikrotubule) świeci na zielono:
Zadanie 3. Mejoza. Badanie i analiza mikrofotografii, rysunki faz mejozy.
Rozważ schemat mejozy (ryc. 5) i odpowiedz na pytania:
Rysunek 5Diagram mejozy (mikrografie i rysunki)

Jakie komórki powstają w wyniku mejozy?
Ile podziałów występuje w mejozie?
Jakie cechy występują w profazie, metafazie, anafazie, telofazie I podziału?
Jakie cechy występują w profazie, metafazie, anafazie, telofazie 2 podziałów?
Jaka jest liczba chromosomów na początku mejozy i na końcu 1 podziału?
Jaka jest liczba chromosomów na początku mejozy i na końcu podziału 2?
Spójrz na ryc. 6 i schematycznie przedstaw etapy mejozy, których brakuje na ryc. 5. Odpowiedz na pytania:
Czym metafaza 1 różni się od metafazy mitozy?
Jaki proces pokazano na ryc. 7?
Co daje ten proces?
Co to są grupy sprzęgieł?
Co się stanie, gdy zostaną naruszone?


Rysunek 6. Schemat mitozy Rycina 7. Chromosomy w fazie profazy 1
Przeczytaj tekst „Znaczenie mejozy”. Sformułuj wnioski i zapisz je.
Znaczenie mejozy.
„W organizmach rozmnażających się płciowo zapobiega się podwojeniu liczby chromosomów w każdym pokoleniu, ponieważ podczas tworzenia komórek rozrodczych w wyniku mejozy liczba chromosomów ulega zmniejszeniu.
Mejoza stwarza szansę na pojawienie się nowych kombinacji genów (zmienność kombinacyjna), w miarę powstawania genetycznie odmiennych gamet.
Zmniejszenie liczby (połowienie) chromosomów prowadzi do powstania „czystych gamet” zawierających tylko jeden allel odpowiedniego locus.
Położenie biwalentów płytki równikowej wrzeciona w metafazie 1 i chromosomów w metafazie 2 jest ustalane losowo. Późniejsza rozbieżność chromosomów w anafazie prowadzi do powstania nowych kombinacji alleli w gametach.
U podstaw leży niezależna segregacja chromosomówTrzecie prawo Mendla.
Zadanie 4. Badanie etapów gametogenezy człowieka.
Zarodkowe komórki nabłonkowe gonad męskich i żeńskich przechodzą serię kolejnych podziałów mitotycznych i mejotycznych zwanych gametogenezą.


Rycina 8. Schemat gametogenezy
Rozważ schemat głównych etapów spermatogenezy i oogenezy na ryc. 7 i odpowiedz na pytania.
Jakie są etapy gametogenezy?
Jaki podział zachodzi na etapie reprodukcji?
Jaki podział zachodzi w fazie wzrostu? Jakie procesy zachodzą na tym etapie?
Jak nazywają się powstałe komórki? Określ zestaw chromosomów.
Jakie podziały zachodzą w fazie dojrzewania? Jaki jest zestaw chromosomów oocytów i spermatocytówII-ta kolejność?
Przeczytaj tekst „Oogeneza”. Krótko opisz cechy oogenezy. Odpowiedz na pytania:
Ile razy ciało kobiety przechodzi fazę rozrodczą?
Kiedy kończy się tworzenie oocytów?I-ta kolejność?
Jakie cechy ma okres dojrzewania oocytu?II-ta kolejność?
Znajdź nieścisłość w stwierdzeniu ostatniego akapitu tekstu „Oogeneza”. Zapisz to stwierdzenie.
Wyjaśnij, dlaczego wiek matki jest uważany za jedną z głównych przyczyn występowania mutacji w komórkach rozrodczych, a zatem pojawiających się dziedzicznych patologii u dzieci?
„Oogeneza”
„W przeciwieństwie do produkcji nasienia, która u mężczyzn rozpoczyna się dopiero w okresie dojrzewania, tworzenie komórek jajowych u kobiet rozpoczyna się jeszcze przed ich urodzeniem. Okres lęgowy odbywa się całkowicie w fazie rozwoju embrionalnego, około 12 tygodni, i kończy się w momencie urodzenia.
W wieku 12-13 lat co miesiąc jeden z oocytów I rzędu kontynuuje mejozę. W wyniku pierwszego podziału mejotycznego powstają dwie komórki potomne. Jedno z nich, stosunkowo małe, nazywane jest pierwszym ciałkiem polarnym, a drugie, większe, jest oocytem II rzędu.
Drugi podział mejozy następuje przed etapem metafazy II i będzie kontynuowany dopiero po interakcji oocytu drugiego rzędu z plemnikiem i nastąpi zapłodnienie.
Ściśle rzecz ujmując, z jajnika nie wychodzi jajo, lecz oocyt drugiego rzędu.
Dopiero po zapłodnieniu następuje podział, w wyniku czegojajko (Lubjajko ) Idrugie ciało polarne . Jednak tradycyjnie, dla wygody, jajo nazywane jest oocytem drugiego rzędu, gotowym do interakcji z plemnikiem.
Dlatego bardzo ważne jest, aby przyszła mama prowadziła zdrowy tryb życia, gdyż wpływa to na zdrowie nie tylko nienarodzonego dziecka, ale także przyszłych wnuków.”
2.2. Badanie wzorców implementacji informacji genetycznej na poziomie biochemicznym.

Rysunek 7Schemat rodzajów kwasów nukleinowych
1. Spójrz na rysunki 7, 8 i odpowiedz na pytania:
Jakie znasz kwasy nukleinowe i czym się różnią?
Co to jest cząsteczka DNA?
Co to jest monomer DNA?
Co to jest nukleotyd, jakie są rodzaje?
Wymień rodzaje RNA i ich funkcje.

Rysunek 8Struktury kwasów nukleinowych
6. Jak rozszyfrowana jest informacja genetyczna? Narysuj uproszczony schemat realizacji informacji dziedzicznej.
7. Czym jest transkrypcja i translacja (Rysunek 8)?
8. Co to jest kodon?
9. Zdefiniuj kod genetyczny.
10. Wymień właściwości kodu genetycznego.


Rysunek 9Rysunek transmisji 10. Opcja nagrywania sektorowego,
koło wewnętrzne - pierwsza zasada kodonu
(od końca 5")
2. Rozwiązywać problemy:
Jakie zmiany zajdą w strukturze białka, jeżeli guanina znajdzie się w regionie genu TAACAAAAGAACAAAA pomiędzy 10 a 11 nukleotydami, a pomiędzy13 i 14 cytozyna, a na końcu pojawia się adenina? Jak nazywają się wydarzenia?mutacje?
Na fragmencie jednej nici DNA nukleotydy ułożone są w sekwencji AATAGTCATGTGTGATCAG. a) Narysuj schemat dwuniciowej cząsteczki DNA, wyjaśnij, jaką właściwością DNA się kierowałeś? b) Zapisz mRNA na dolnej nici. Jak nazywa się ten proces? c) Jaka jest struktura białkowa kodowanego genu.
Polipeptyd składa się z następujących aminokwasów: walina – alanina – glicyna – lizyna – tryptofan – walina – seryna – asparagina – kwas glutaminowy. Określ strukturę odcinka DNA kodującego określony polipeptyd.
2.3. Badanie zmienności dziedzicznej
1. Rozważmy schemat klasyfikacji mutacji na ryc. 11. Zdefiniuj każdy typ.
( Tautomeryzm (od greckiego ταύτίς – to samo i μέρος – miara) to zjawisko odwracalnej izomerii, w której dwa lub więcej izomerów łatwo przekształca się w siebie.

Rysunek 11Klasyfikacja mutacji.
2. Przeczytaj tekst. „Chromosom Filadelfii”, zapisz wnioski
Chromosom Filadelfii
„Pierwszą opisaną strukturalną rearanżacją genomu w komórkach somatycznych powodującą raka jest tak zwany chromosom Filadelfia, który zgodnie z Międzynarodową Nomenklaturą Cytogenetyczną Człowieka ma swoje własne oznaczenie – Ph.
Chromosom ten został nazwany na cześć miasta w USA, w którym pracowali jego odkrywcy P. Nowell i D. Hungerford, którzy w 1960 roku opisali niezwykły mały chromosom u dwóch pacjentów z przewlekłą białaczką szpikową. Obecnie wiadomo, że chromosom Filadelfia powstaje w wyniku wzajemnej translokacji między chromosomami 9 i 22, a mutacja tapowoduje 95% przypadków przewlekłej białaczki szpikowej. Mutacja ta jest również jedną z najczęstszych w ostrej białaczce limfoblastycznej z komórek B u dorosłych.
Nie jest jasne, dlaczego tak się dzieje, ale zidentyfikowano czynnik, który to powoduje – promieniowanie jonizujące”.
3. Rozważ rysunek 13 i zapisz odpowiednio czynniki mutagenne (MF) i zmiany mutacyjne (MI) w 2 kolumnach. Uzupełnij kolumnę MF przykładami
Pirymidyny:C(C),T(T),U(U), puryny: A(A),G(G).
Przykład:
Promieniowanie UV.Niekontrolowane narażenie na aktywne promieniowanie słoneczne.
Solarium
Lampy bakteriobójcze
1. Tworzenie dimerów T-T. Nieprawidłowa rekombinacja: delecje. wstawki

Rysunek 12Czynniki mutagenne - konsekwencje ich wpływu na DNA
3 . Rozwiązywać problemy:
U osoby chorej na cystynurię (większa niż zwykle liczba aminokwasów w moczu) z moczem wydalane są aminokwasy, które odpowiadają trójkom mRNA: UCU, UGU, HCU, GGU, CAG, CGU, AAA. U zdrowego człowieka w moczu stwierdza się alaninę, serynę, kwas glutaminowy i glicynę. Wydalanie jakich aminokwasów z moczem jest typowe dla pacjentów z cystynurią? Zapisz trojaczki odpowiadające aminokwasom występującym w moczu osoby zdrowej
Czwarty peptyd w normalnej hemoglobinie (hemoglobina A) składa się z następujących aminokwasów: walina – histydyna – leucyna – treonina – prolina – kwas glutaminowy – kwas glutaminowy – lizyna. U pacjenta z objawami powiększenia śledziony i umiarkowanej niedokrwistości stwierdzono następujący skład czwartego peptydu: walina – histydyna – leucyna – treonina – prolina – lizyna – kwas glutaminowy – lizyna. Określ zmiany, jakie zaszły w DNA kodującym czwarty peptyd hemoglobiny po mutacji.
3. Część końcowa – 14 minut
3.1 . Zreasumowanie;
Ostateczna rozmowa
Cechowanie
3.2 . Praca domowa.
Powtórz tematy: „Dziedziczenie cech w skrzyżowaniach monohybrydowych, dihybrydowych i polihybrydowych. Dziedziczne właściwości krwi.”
Przygotuj odpowiedzi na pytania:
Co to jest gen, geny alleliczne?
Jak oznaczane są znaki?
Jakie cechy nazywamy dominującymi i recesywnymi?
Co to jest genotyp, fenotyp?
W jaki sposób dziedziczone są cechy w krzyżówce monohybrydowej?
W jaki sposób dziedziczone są cechy w przypadku niepełnej dominacji?
Jakie są zasady klasyfikacji chorób dziedzicznych?
Z czym wiąże się patologia chorób chromosomalnych?
Jakie są zasady klasyfikacji chorób genowych?
Wymień rodzaje klasyfikacji chorób jednogenowych.
Co oznacza skrzyżowanie mono-, di- i polihybrydowe?
Co oznacza penetracja i ekspresja?

Strona 15
- (aspekty demograficzne), dział genetyki zajmujący się badaniem zjawisk dziedziczności i zmienności u człowieka. Materialną podstawą dziedziczności u człowieka, podobnie jak u innych organizmów, są geny zlokalizowane na chromosomach i przekazywane z pokolenia na pokolenie... ... Demograficzny słownik encyklopedyczny
Dział genetyki ściśle powiązany z antropologią i medycyną. G. h. są tradycyjnie podzielone na antropogenetykę, która bada dziedziczność i zmienność normalnych cech organizmu ludzkiego oraz genetykę medyczną (patrz Genetyka ... ...
Federalny stan organizacja finansowana przez państwo Instytut Naukowy Genetyki Molekularnej Akademia Rosyjska Nauki (IMG RAS) Założona w 1978 r. Członek dyrektora. kor. Pracownicy S. V. Kostrov 134 (2 ... Wikipedia
Spis treści 1 Początki genetyki 2 Era genetyki klasycznej 3 Era DNA… Wikipedia
POWSTANIE ROSYJSKIEJ AKADEMII NAUK INSTYTUT CYTOLOGII I GENETYKI SYBERYJSKI ODDZIAŁ RAS (ICIG SB RAS) Nazwa międzynarodowa Instytut Cytologii i Genetyki Syberyjskiego Oddziału Rosyjskiej Akademii Nauk Założona 1957 Dyrektor... ... Wikipedia
Spis treści 1 Początki genetyki 2 Era genetyki klasycznej 3 Era DNA 4 Era genomu… Wikipedia
Federalna Państwowa Instytucja Budżetowa Naukowego Instytutu Cytologii i Genetyki Syberyjskiego Oddziału Rosyjskiej Akademii Nauk (ICiG SB RAS) ... Wikipedia
Etologia to dziedzina zoologii zajmująca się badaniem zachowań zwierząt. Termin wprowadził w 1859 roku francuski zoolog I. Geoffroy Saint-Hilaire. Ściśle związany z zoologią, teorią ewolucji, fizjologią, genetyką, psychologią porównawczą.... ... Wikipedia
- (z greckiego eugenēs dobrego rodzaju) doktryna o dziedzicznym zdrowiu człowieka i sposobach poprawy jego dziedzicznych właściwości, o możliwych metodach aktywnego wpływania na ewolucję ludzkości w celu dalszego doskonalenia jej natury, o ... ... Wielka encyklopedia radziecka
Liceum Technologie informacyjne Nr 1537 Dyrektor: TRUNOVA Tatiana Władimirowna. Typ: Licealiści: 400 Adres: 129347, Moskwa, ul. Prokhodchikov, 9 (dzielnica północno-wschodnia, stacja metra „Babushkinskaya”) Telefon ... Wikipedia
© Wydawnictwo „SpetsLit” LLC, 2005
Przedmowa
Podstawą jest genetyka jako nauka o prawach dziedziczności i zmienności współczesna biologia, ponieważ determinuje rozwój wszystkich pozostałych dyscyplin biologicznych. Rola genetyki nie ogranicza się jednak do dziedziny biologii. Zachowanie człowieka, ekologia, socjologia, psychologia, medycyna - są dalekie od tego pełna lista kierunki naukowe, którego postęp zależy od poziomu wiedzy z zakresu genetyki. Biorąc pod uwagę „strefę wpływu” genetyki, jej metodologiczna rola jest jasna.
Jeden z charakterystyczne cechy nowoczesna nauka to stale pogłębiające się zróżnicowanie i specjalizacja. Proces ten osiągnął poziom, powyżej którego istnieje już realne zagrożenie utratą wzajemnego zrozumienia nawet pomiędzy przedstawicielami tej samej nauki. W biologii, ze względu na bogactwo dyscyplin specjalnych, tendencje odśrodkowe są szczególnie ostre. Obecnie to genetyka wyznacza jedność nauk biologicznych, dzięki powszechności praw dziedziczności i podstawowych informacji usystematyzowanych w przepisach genetyki ogólnej. Metodologiczna rola genetyki w pełni odnosi się do wszystkich nauk humanistycznych.
W związku z tym chciałbym poczynić krytyczne uwagi na temat prowadzenia zajęć z psychogenetyki na wydziałach psychologii uniwersytetów. Psychogenetyka jest jedną z najbardziej złożonych i najsłabiej rozwiniętych gałęzi genetyki. Jego badania powinny opierać się na podstawowym ogólnym treningu biologicznym i ogólnym genetycznym. W przeciwnym razie przebieg psychogenetyki stanie się czysto dekoracyjny, reprezentujący bardziej odmianę psychologii różnicowej niż genetykę, co możemy obecnie zaobserwować. Znajomość praw dziedziczności odgrywa ogromną rolę w edukacji psychologicznej. Każde ludzkie zachowanie jest w takim czy innym stopniu powiązane z dziedzictwem filogenetycznym. Aby zrozumieć subtelne mechanizmy tej zależności, wymagana jest nie powierzchowna, ale głęboka wiedza.
Metodologiczna rola genetyki w edukacji z góry określa specjalne wymagania dotyczące jej nauczania, które musi łączyć w sobie szeroki zakres, głębię naukową i dostępność prezentacji. Podręcznik ten odpowiednio analizuje wszystkie sekcje współczesnej nauki genetyki niezbędne do zrozumienia genetyki i zachowania człowieka, dlatego mamy nadzieję, że będzie przydatny dla wszystkich studentów i studentów pracownicy naukowi którzy studiują te dziedziny. Krótkie, ale całościowe prezentacje podstawowych zasad genetyki są szczególnie potrzebne na wydziałach psychologii.
W naszym kraju opublikowano wiele dobre podręczniki I pomoc naukowa na temat genetyki autorstwa autorów rosyjskich i zagranicznych (Gershenzon S.M., 1983; Ayala F., Kaiger J., 1988; Alikhanyan S.S., Akifev A.P., 1988; Inge-Vechtomov S.G., 1989). Wiele podręczników koncentruje się na genetyce człowieka (Fogel F., Motulski A., 1989–1990; Bochkov N. P., 2004). Niedawno, po krótkiej przerwie, na półkach naszych sklepów ponownie pojawiają się książki o genetyce (Żimulew I.
F., 2003; Tarantula V.Z., 2003; Grinev V.V., 2006). Tak różnorodna literatura na ten temat może tylko zadowolić każdego, kto jest pasjonatem tak wspaniałej nauki, jak genetyka.
Rozdział 1. Historia i znaczenie genetyki
Genetyka jest podstawą nauk biologicznych. Różnorodność form i procesów życia można ująć w jedną całość jedynie w ramach genetyki.
Genetyka bada dwie nierozłączne właściwości organizmów żywych - dziedziczność i zmienność. Obecnie stanowi podstawę współczesnej biologii.
1.1. Historia genetyki
Chociaż genetyka jako nauka ma nieco ponad 100 lat, historia jej początków sięga wieków wstecz. Historia genetyki to nie tylko historia konkretnej nauki, ale raczej samodzielny dział biologii, w którym splatają się problemy biologiczne, psychologiczne i filozoficzne (Gaisinovich A. E., 1988; Zakharov I. P., 1999). W tej historii są momenty pełne dramatyzmu. A obecnie genetyka pozostaje w czołówce dyskursu społecznego, wywołując gorące dyskusje wokół problemów determinacji zachowania, klonowania ludzi, Inżynieria genetyczna. Historia genetyki w naszym kraju jest zupełnie wyjątkowa, znająca czasy globalnej interwencji ideologii w nauce (Soifer V.N., 1989; Dubinin N.P., 1990).
Jaka jest przyczyna tak wyjątkowej roli genetyki w życiu społeczeństwa? Genetyka jest rdzeniem współczesnej biologii, podstawą zrozumienia takich zjawisk jak życie, ewolucja, rozwój, a także natura samego człowieka. W historii nauk przyrodniczych rozważa się problem dziedziczności, zaczynając od dzieł starożytnych myślicieli. We współczesnej nauce jest to szczegółowo omawiane w pracach takich luminarzy, jak C. Linneusz (1707–1778), J. Buffon (1707–1788), K. F. Wolf (1734–1794), J.-B. Lamarck (1744–1829), C. Darwin (1809–1882), T. Huxley (1825–1895), A. Weissman (1834–1914) i wielu innych. W tamtych czasach problemy genetyki rozpatrywano w kategoriach zagadnień hybrydyzacji, rozwoju, transformacji (lub odwrotnie, stałości) gatunków.
Za twórcę genetyki uważa się G. Mendla (1822–1884), który uzasadnił podstawowe prawa dziedziczności. Odkrycie to nie zostało docenione przez jego współczesnych, w tym przez czołowego biologa tamtych czasów K. Nägeli (1817–1891), któremu G. Mendel przesyłał swoje prace do recenzji.
Ponowne odkrycie praw Mendla przez G. de Vriesa (1848–1935), K. Corrensa (1864–1933), E. Cermaka (1871–1962) w 1900 uważa się za datę narodzin genetyki jako samodzielnej nauki. W tym czasie społeczność naukowa biologów była gotowa to zaakceptować nowy koncept. Odkryto już zjawiska mitozy i mejozy, opisano chromosomy i proces zapłodnienia, stworzono nuklearną teorię dziedziczności. Pomysły inspirowane „odkrytymi na nowo” wzorami rozprzestrzeniły się po całym świecie z niesamowitą szybkością. świat naukowy, posłużył jako potężny impuls do rozwoju wszystkich dziedzin biologii.
Najciekawsza historia genetyki, chronologia najważniejsze odkrycia, biografie G. Mendla i innych wybitnych naukowców opisano w setkach książek. Opisane szczegółowo i tragiczna historia genetyka w ZSRR. Wiele książek czyta się z niesłabnącym zainteresowaniem i dostarczają niezastąpionego materiału do zrozumienia tej nauki, związku praw genetyki z problemami społeczeństwa ludzkiego.
Przyjrzyjmy się niektórym kamieniom milowym w historii genetyki
1901 - G. de Vries zaproponował pierwszą teorię mutacji.
1903 – W. Sutton (1876–1916) i T. Boveri (1862–1915) wysunęli hipotezę chromosomową, „łącząc” mendlowskie czynniki dziedziczności z chromosomami.
1906 – W. Bateson (1861–1926) ukuł termin „genetyka”.
1907 - W. Bateson opisał warianty interakcji między genami („czynniki dziedziczne”) i wprowadził pojęcia „komplementarności”, „epistazy” i „niepełnej dominacji”. Wcześniej (1902) wprowadził terminy „homozygota” i „heterozygota”.
1908 – G. Nilsson-Ehle (1873–1949) wyjaśnił i wprowadził pojęcie „polimeryzmu”, które oznacza najważniejsze zjawisko w genetyce cech ilościowych.
G. Hardy (1877–1947) i V. Weinberg (1862–1937) zaproponowali wzór na rozmieszczenie genów w populacji, znany później jako prawo Hardy'ego-Weinberga, kluczowe prawo genetyki populacyjnej.
1909 – V. Johannsen (1857–1927) sformułował szereg podstawowych zasad genetyki i wprowadził podstawowe pojęcia: „gen”, „genotyp”, „fenotyp”, „allel”. V. Volterek wprowadził pojęcie „normy reakcji”, która charakteryzuje możliwe spektrum manifestacji genu.
1910 – L. Plate (1862–1937) opracował koncepcję działania wielogenowego i wprowadził pojęcie „plejotropii”.
1912 – T. Morgan (1866–1945) zaproponował teorię lokalizacji genów na chromosomach. Już w połowie lat dwudziestych XX w. T. Morgan i przedstawiciele jego szkoły – A. Sturtevant (1891–1970), K. Bridges (1889–1938), G. Meller (1890–1967) sformułowali własną wersję teorii genu. Problem genów stał się głównym problemem genetyki.
1920 – G. Winkler wprowadził termin „genom”. Następnie rozwój tej koncepcji stał się nowym etapem rozwoju genetyki.
N.I. Wawiłow (1887–1943) sformułował prawo szeregów homologicznych o dziedzicznej zmienności.
1921 - L. N. Delaunay (1891–1969) zaproponował termin „kariotyp” na określenie całości chromosomów organizmu. Termin „idiogram”, zaproponowany wcześniej przez S. G. Navashina (1857–1930), później zaczęto stosować w odniesieniu do standaryzowanych kariotypów.
1926 - N.V. Timofeev-Resovsky (1900–1981) rozwinął problem wpływu genotypu na manifestację cechy i sformułował pojęcia „penetracji” i „ekspresywności”.
1927 - G. Möller otrzymuje mutacje sztucznie pod wpływem promieniowania radioaktywnego. W 1946 roku otrzymał Nagrodę Nobla za dowody mutacyjnego wpływu promieniowania.
1929 – A. S. Serebrovsky (1892–1948) jako pierwszy wykazał złożoną naturę genu i wykazał, że gen nie jest jednostką mutacji. Sformułował także koncepcję „puli genowej”.
1930–1931 – D. D. Romashov (1899–1963), N. P. Dubinin (1907–1998), S. Wright (1889–1988), R. Fisher (1890–1962), J. Haldane (1860–1936) opracowali teoretyczne kierunki genetyki populacyjnej i przedstawił stanowisko dryfu genetycznego.
1941 – J. Beadle (1903–1989) i E. Tatum (1909–1975) formułują zasadnicze stanowisko: „jeden gen, jeden enzym” (Nagroda Nobla 1958).
1944 – O. Avery (1877–1955), K. McLeod (1909–1972), M. McCarthy udowodnili genetyczną rolę DNA w doświadczeniach nad transformacją mikroorganizmów. Odkrycie to symbolizowało początek nowego etapu - narodziny genetyki molekularnej.
1946 – J. Lederberg, E. Tatum, M. Delbrück (1906–1981) opisali rekombinację genetyczną u bakterii i wirusów.
1947 - B. McClintock (1902–1992) po raz pierwszy opisał migrujące elementy genetyczne (to wybitne odkrycie Zaobserwowano nagroda Nobla dopiero w 1983 r.).
1950 – E. Chargaff wykazał zgodność nukleotydów purynowych i pirymidynowych w cząsteczce DNA (reguła Chargaffa) i jej specyficzność gatunkową.
1951 – J. Lederberg wraz ze współpracownikami odkrył zjawisko transdukcji, które później odegrało kluczową rolę w rozwoju inżynierii genetycznej.
1952 – A. Hershey (1908–1997) i M. Chase wykazali decydującą rolę kwasu deoksyrybonukleinowego w infekcji wirusowej, co było ostatecznym potwierdzeniem znaczenie genetyczne DNA.
1953 - J. Watson i F. Crick zaproponowali strukturalny model DNA. Ta data jest brana pod uwagę początek ery współczesnej biologii.
1955 – S. Ochoa (1905–1993) wyizolował enzym Polimeraza RNA i jako pierwszy zsyntetyzował RNA in vitro.
1956 – A. Kornberg wyizolował enzym Polimeraza DNA i przeprowadził proces replikacji DNA w warunkach laboratoryjnych.
1957 – M. Meselson i F. Stahl udowodnili semikonserwatywny mechanizm replikacji DNA. t-RNA odkryto w laboratorium M. Hoaglanda.
1958 - F. Crick sformułował „centralny dogmat Biologia molekularna».
1960 - M. Nirenberg, J. Mattei, G. Korana rozpoczęli badania nad rozszyfrowaniem kodu genetycznego. Prace (obejmujące kilka grup badawczych) zakończono w 1966 roku. Opracowanie słownika kodowego było jednym z największych osiągnięć naukowych w historii ludzkości.
1961 – F. Jacob i J. Monod (1910–1976) sformułowali teorię operonu – teorię genetycznej regulacji syntezy białek u bakterii.
1962 - J. Gurdon po raz pierwszy uzyskał sklonowane kręgowce.
1965 – R. Holley (1922–1993) odkrył strukturę t-RNA.
1969 - G. Korana po raz pierwszy zsyntetyzował gen w laboratorium.
1970 – G. Temin (1934–1994) i D. Baltimore odkryli zjawisko odwrotnej transkrypcji.
1972 – P. Berg uzyskał pierwszą zrekombinowaną cząsteczkę DNA. Datę tę uważa się za datę narodzin inżynierii genetycznej.
1974 – R. Kornberg, A. Olins, D. Olins sformułowali teorię organizacji nukleosomów chromatyny.
1975 – Z inicjatywy grupy naukowców pod przewodnictwem P. Berga („Komitet Berga”) w Asilomar (USA) odbyła się Międzynarodowa Konferencja na temat Zagadnień Etycznych Inżynierii Genetycznej, na której uchwalono tymczasowe moratorium na szereg badań zdeklarowany.
Moratorium nie wstrzymało prac nad inżynierią genetyczną, a w kolejnych latach dziedzina ta aktywnie się rozwijała i narodził się nowy kierunek - biotechnologia.
1976 - D. Bishop i G. Varmus odkryli naturę onkogenu (Nagroda Nobla 1989).
1977 – W. Gilbert, A. Maxam, F. Senger opracowali metody sekwencjonowania (określania sekwencji nukleotydowej kwasów nukleinowych).
R. Roberts i F. Sharp pokazali strukturę mozaikową (intron-ekson) genu eukariotycznego (Nagroda Nobla 1993).
1978 – Przeprowadzono transfer genów eukariotycznych (insulina) do komórki bakteryjnej, gdzie syntetyzowane jest w niej białko.
1981 – Wyhodowano pierwsze zwierzęta transgeniczne (myszy). Określono pełną sekwencję nukleotydową ludzkiego genomu mitochondrialnego.
1982 – Wykazano, że RNA może mieć właściwości katalityczne, podobnie jak białko. Fakt ten dodatkowo promował RNA do roli „pierwszej cząsteczki” w teoriach pochodzenia życia.
1985 – Sklonowano i zsekwencjonowano DNA wyizolowane ze starożytnej egipskiej mumii.
1988 – Z inicjatywy amerykańskich genetyków powstał międzynarodowy projekt poznania genomu ludzkiego.
1990 – V. Andersen po raz pierwszy wprowadził do organizmu człowieka nowy gen.
1995 – Odszyfrowano pierwszy genom bakterii. Kształtowanie się genomiki jako samodzielnej gałęzi genetyki.
1997 - J. Wilmut przeprowadził pierwszy udany eksperyment z klonowaniem ssaków ( Dolly, owca).
1998 – Zsekwencjonowano genom pierwszego przedstawiciela eukariontów – nicienia Caenorhabditis eleganckie.
2000 – Zakończenie sekwencjonowania ludzkiego genomu.
Genetyka staje się coraz bardziej częścią życie codzienne ludzi, w dużej mierze determinując przyszłość ludzkości. Badania nad ludzkim genomem prowadzone są coraz intensywniej.
Nie ma wątpliwości, że eksperymenty z „inżynierią ludzką” będą kontynuowane pomimo wszelkich zakazów. Tematyka klonowania człowieka, wpływ na jego genotyp, zagrożenia związane z modyfikowanymi produktami są coraz częściej poruszane w prasie... Nie sposób przewidzieć, jak to wszystko wpłynie na losy ludzkości.
1.2. Zagadnienia kluczowe w historii genetyki
W historii genetyki (i jej prehistorii) można wyróżnić szereg kluczowych tematów ze względu na ich znaczenie dla światopoglądu naukowego i intensywność dyskusji. W XVII–XVIII w. - był to problem „preformacjonizmu - epigenezy”, a obóz preformacjonistów dzielił się na „ovistów” i „animalkulistów”, w zależności od tego, czy nosicielem „zarodka” była płeć żeńska czy męska. Aktywnie dyskutowano także nad problemem „stałości – transformizmu”.
Problem dziedziczenia cech nabytych, wielokrotnie „ostatecznie” pogrzebany w historii genetyki, powracał równie wielokrotnie. W Związku Radzieckim dyskusje wokół tej pozornie prywatnej kwestii naukowej zyskały na pewnym etapie historii ogromny oddźwięk społeczny, co zaowocowało licznymi ludzkimi tragediami. Nie ma to odpowiednika w innych naukach. W 1958 r. F. Crick sformułował „centralny dogmat biologii molekularnej”, zgodnie z którym przekazywanie informacji dziedzicznej przebiega w kierunku od DNA do RNA i od RNA do białek. Głównym punktem tego schematu jest niemożność kodowania białek do kwasów nukleinowych (chociaż dopuszczona jest możliwość przeniesienia informacji z RNA do DNA). Dlatego wszelkie próby wskrzeszenia hipotezy o dziedziczeniu cech nabytych w oparciu o nowe odkrycia (a są takie próby) zostały przez genetykę odrzucone. Obecnie kwestia ta jest ponownie aktywnie omawiana w związku z ostatnimi odkryciami.
Szczególnie interesujący w historii genetyki był problem nośnika informacji dziedzicznej. Chromosomy nie zostały od razu rozpoznane jako struktury odpowiedzialne za dziedziczność. Po tym rozpoznaniu rolę molekularnego nośnika informacji genetycznej bardziej skłaniano się do przypisania białek. DNA wydawało się cząsteczką zbyt prostą do pełnienia tak ważnej funkcji. Przełom w rozumieniu roli DNA nastąpił w 1944 r. po doświadczeniach O. Avery'ego, K. McLeoda, M. McCarthy'ego nad transformacją cech u pneumokoków i identyfikacją czynnika transformującego, jakim jest DNA. Choć odkrycie to symbolizuje narodziny genetyki molekularnej, należy stwierdzić, że ostateczne potwierdzenie roli DNA uzyskano dopiero w 1952 roku po pracach A. Hersheya i M. Chase'a nad badaniem transdukcji przez bakteriofagi.
Znajomość historii pokazuje, że rozwój genetyki nie był ściśle postępowy, że genialne odkrycia przeplatały się z długimi złudzeniami, że najwięksi uczeni często byli niewolnikami fałszywych przekonań. Założyciel teoria chromosomów dziedziczność Sam T. Morgan przez długi czas wątpił w rolę chromosomów. Przeciwnikami teorii chromosomów byli W. Batson i V. Johannsen. A. Hershey, któremu przypisuje się definitywne udowodnienie genetycznej roli DNA, wyraził wątpliwości co do tej hipotezy.
Można podać wiele takich przykładów. Natura nie chciała zdradzać swoich tajemnic. Myśl teoretyczna często nie nadążała szybki rozwój badania eksperymentalne, ciągłe komplikacje obserwowanych wzorców. Nie było także jednomyślności w interpretacji tych wzorców.
Nowa era współczesnej genetyki (i całej biologii) rozpoczyna się w 1953 roku, kiedy J. Watson i F. Crick opublikowali strukturalny model DNA. Ale nawet teraz, ponad pół wieku później, pomimo wybitnych odkryć i osiągnięć, genetyka jest pełna tajemnic. To czyni ją intrygująco interesującą.
1.3. Struktura genetyki i jej ogólne znaczenie biologiczne
Współczesna genetyka to obszerne drzewo dyscyplin pochodnych. Jej wyspecjalizowane sekcje zaczęto uważać za duże niezależne nauki - genetykę człowieka, cytogenetykę, genetyka molekularna, genetyka populacyjna, immunogenetyka, genetyka środowiskowa, genetyka rozwojowa, genomika itp.
Tendencja do różnicowania nauk ujawniła się także w kierunku badania genetyczne człowieka: powstały takie działy, jak genetyka kliniczna, genetyka biochemiczna człowieka, cytogenetyka człowieka, neurogenetyka itp. Jednocześnie problem „wąskiej specjalizacji” w genetyce nie jest tak dotkliwy jak w innych naukach. Wszystkie specjalistyczne dyscypliny genetyczne łączą podstawowe informacje usystematyzowane w ramach genetyki ogólnej. Co więcej, pod wieloma względami to genetyka stanowi obecnie o jedności współczesnej biologii, dlatego XVI Światowy Kongres Genetyczny w 1988 roku odbył się pod hasłem „Genetyka i jedność biologii”.
Bez przesady można powiedzieć, że genetyka w takim czy innym stopniu warunkuje rozwój wszystkich dziedzin biologii i stanowi jej podstawę metodologiczną. Przedmiotem badań genetyki jest dziedziczność i zmienność – właściwości uniwersalne dla wszystkich istot żywych. Dlatego prawa genetyki są również uniwersalne.
Rozdział 2. Molekularne podstawy dziedziczności
Wyobraź sobie, że gdybyś powiększył osobę do rozmiarów Wielkiej Brytanii, wówczas komórka miałaby wielkość budynku fabrycznego. Wewnątrz komórki znajdują się cząsteczki zawierające tysiące atomów, w tym cząsteczki kwasu nukleinowego. Zatem nawet przy tak ogromnym wzroście cząsteczki kwasu nukleinowego będą cieńsze niż przewody elektryczne.
J. Kendrew, angielski biochemik, laureat Nagrody Nobla w 1962 r
Eksperymenty 1940–1950 przekonująco udowodnił, że to kwasy nukleinowe (a nie, jak wielu przypuszczało, białka) są nośnikami informacji dziedzicznej we wszystkich organizmach.
2.1. Struktura kwasu nukleinowego
Kwasy nukleinowe zapewniają różne procesy przechowywania, wdrażania i odtwarzania informacji genetycznej.
Kwasy nukleinowe to polimery, których monomery są nukleotydy. Nukleotyd obejmuje zasada azotowa węglowodan pentoza i reszta Kwas fosforowy(ryc. 2.1).
Zasady azotowe nukleotydów dzielą się na dwa typy: pirymidynę (składającą się z jednego pierścienia 6-członowego) i purynę (składającą się z dwóch skondensowanych pierścieni 5- i 6-członowych). Każdy atom węgla pierścieni podstawowych ma swoją własną, określoną liczbę. Każdy atom węgla pentozy ma również swój własny numer, ale z indeksem pierwszym ("). W nukleotydzie zasada azotowa jest zawsze przyłączona do pierwszego atomu węgla pentozy.
To właśnie zasady azotowe decydują o unikalnej strukturze cząsteczek DNA i RNA. Istnieje 5 głównych rodzajów zasad azotowych występujących w kwasach nukleinowych (purynowe - adenina i guanina, pirymidyna – tymina, cytozyna, uracyl) i ponad 50 rzadkich (nietypowych) baz. Główne zasady azotowe są oznaczone początkowymi literami: A, G, T, C, U. Większość nietypowych zasad jest specyficzna dla określonego typu komórek.

Ryż. 2.1. Struktura nukleotydów
Tworzenie liniowego łańcucha polinukleotydowego następuje poprzez utworzenie wiązania fosfodiestrowego pomiędzy pentozą jednego nukleotydu i fosforanem innego. Szkielet pentozofosforanowy składa się z (5 " – 3" ) – połączenia. Końcowy nukleotyd na jednym końcu łańcucha zawsze ma wolny 5" -grupa, z drugiej - 3 " -Grupa.
W przyrodzie występują dwa rodzaje kwasów nukleinowych: DNA i RNA. W organizmach prokariotycznych i eukariotycznych funkcje genetyczne pełnią oba typy kwasów nukleinowych. Wirusy zawsze zawierają tylko jeden rodzaj kwasu nukleinowego.
Genetyka jako nauka o prawach dziedziczności i zmienności stanowi podstawę współczesnej biologii, gdyż determinuje rozwój wszystkich pozostałych dyscyplin biologicznych. Rola genetyki nie ogranicza się jednak do dziedziny biologii. Zachowanie człowieka, ekologia, socjologia, psychologia, medycyna – to nie jest pełna lista dziedzin naukowych, których postęp zależy od poziomu wiedzy genetycznej.
Biorąc pod uwagę „strefę wpływów” genetyki, jej metodologiczna rola jest jasna. Jedną z charakterystycznych cech współczesnej nauki jest stale pogłębiające się zróżnicowanie i specjalizacja. Proces ten osiągnął punkt, po przekroczeniu którego istnieje już realne zagrożenie utratą wzajemnego zrozumienia nawet pomiędzy przedstawicielami tej samej nauki. W biologii, ze względu na bogactwo dyscyplin specjalnych, tendencje odśrodkowe są szczególnie ostre. Obecnie to genetyka wyznacza jedność nauk biologicznych, dzięki powszechności praw dziedziczności i podstawowych informacji usystematyzowanych w przepisach genetyki ogólnej. Ta metodologiczna rola genetyki w pełni odnosi się do wszystkich nauk humanistycznych.
Podręcznik do samodzielnej nauki analizuje zagadnienia i podstawowe zasady dziedziczności i zmienności, strukturalną i funkcjonalną organizację materiału genetycznego, genetyczne podstawy ewolucji, zachowania i rozwoju. Odrębnie rozpatrywane są zagadnienia genetyki człowieka, genetyki medycznej i psychogenetyki.
Podręcznik przedstawia różne, często alternatywne, punkty widzenia na nierozwiązane problemy, które powinny pokazać studentom brak utartych ścieżek w nauce i potrzebę analizowania dodatkowej literatury.
Każdy temat zawiera opis jego treści, podstawowe pojęcia, diagramy i tabele. W zadaniach dla niezależna praca nacisk kładzie się na złożone i kontrowersyjne zagadnienia naukowe. W celu samodzielnego sprawdzenia każdy rozdział się kończy pytania kontrolne. Aby uzyskać więcej dogłębne studium podano wykaz dodatkowej literatury. Lista terminów znajdująca się na końcu książki pozwoli studentom sprawdzić swoją wiedzę z studiowanego materiału.
Temat 1. Historia i znaczenie genetyki
Genetyka jest podstawą nauk biologicznych. Różnorodność form i procesów życia można ująć w jedną całość jedynie w ramach genetyki.
F. Ayala, amerykański genetyk
Genetyka bada dwie nierozłączne właściwości organizmów żywych - dziedziczność i zmienność. Obecnie stanowi podstawę współczesnej biologii.
Genetyka jako nauka o dziedziczności i zmienności. Historia genetyki. Główne etapy i kluczowe zagadnienia w historii genetyki. Problem molekularnego nośnika dziedziczności. Działy współczesnej genetyki. Powiązania genetyki z innymi naukami. Uniwersalność praw genetyki.
Za twórcę genetyki uważa się G. Mendla (1822–1884), który uzasadnił podstawowe prawa dziedziczności. Ponowne odkrycie praw Mendla przez G. de Vriesa (1848–1935), K. Corrensa (1864–1933), E. Cermaka (1871–1962) w 1900 uważa się za datę narodzin genetyki jako samodzielnej nauki.
Przyjrzyjmy się kamieniom milowym w rozwoju genetyki w XX wieku.
1901 - G. de Vries zaproponował pierwszą teorię mutacji.
1903 – W. Sutton (1876–1916) i T. Boveri (1862–1915) wysunęli hipotezę chromosomową, „łącząc” mendlowskie czynniki dziedziczności z chromosomami.
1905 – W. Bateson (1861–1926) ukuł termin „genetyka”.
1907 - W. Bateson opisał warianty interakcji między genami („czynniki dziedziczne”) i wprowadził pojęcia „komplementarności”, „epistazy” i „niepełnej dominacji”. On także wcześniej (1902) wprowadził terminy „homozygota” i „heterozygota”.
1908 – G. Nilsson-Ehle (1873–1949) wyjaśnił i wprowadził pojęcie „polimeryzmu”, najważniejszego zjawiska w genetyce cech ilościowych.
G. Hardy (1877–1947) i V. Weinberg (1862–1937) zaproponowali wzór na rozmieszczenie genów w populacji, znany później jako prawo Hardy'ego-Weinberga, kluczowe prawo genetyki populacyjnej.
1909 – V. Johannsen (1857–1927) sformułował szereg podstawowych zasad genetyki i wprowadził podstawowe pojęcia terminologii genetycznej: „gen”, „genotyp”, „fenotyp”, „allel”.
V. Volterek wprowadził pojęcie „normy reakcji”, która charakteryzuje możliwe spektrum manifestacji genu.
1910 - L. Plate opracował koncepcję wielokrotnego działania genów i wprowadził pojęcie „plejotropii”.
1912 – T. Morgan (1866–1945) zaproponował teorię lokalizacji genów na chromosomach. Do połowy lat 20. T. Morgan i przedstawiciele jego szkoły – A. Sturtevant (1891–1970), K. Bridges (1889–1938), G. Meller (1890–1967) sformułowali własną wersję teorii genu. Problem genów stał się głównym problemem genetyki.
1920 – G. Winkler wprowadził termin „genom”. Następnie rozwój tej koncepcji stał się nowym etapem rozwoju genetyki.
N.I. Wawiłow (1887–1943) sformułował prawo szeregów homologicznych o dziedzicznej zmienności.
1921 - L. N. Delaunay (1891–1969) zaproponował termin „kariotyp” na określenie całości chromosomów organizmu. Termin „idiogram”, zaproponowany wcześniej przez S. G. Navashina (1857–1930), później zaczęto stosować w odniesieniu do standaryzowanych kariotypów.
1926 - N.V. Timofeev-Resovsky (1900–1981) rozwinął problem wpływu genotypu na manifestację cechy i sformułował pojęcia „penetracji” i „ekspresywności”.
1927 - G. Möller otrzymał mutacje sztucznie pod wpływem promieniowania radioaktywnego. W 1946 roku otrzymał Nagrodę Nobla za dowody mutacyjnego wpływu promieniowania.
1929 – A. S. Serebrovsky (1892–1948) jako pierwszy wykazał złożoną naturę genu i wykazał, że gen nie jest jednostką mutacji. Sformułował także koncepcję „puli genowej”.
1930–1931 – D. D. Romashov (1899–1963), N. P. Dubinin (1907–1998), S. Wright (1889–1988), R. Fisher (1890–1962), J. Haldane (1860–1936) opracowali teoretyczne kierunki genetyki populacyjnej i przedstawił stanowisko dryfu genetycznego.
1941 – J. Beadle (1903–1989) i E. Tatum (1909–1975) sformułowali zasadnicze stanowisko: „jeden gen, jeden enzym” (Nagroda Nobla 1958).
1944 – O. Avery (1877–1955), C. McLeod (1909–1972), M. McCarthy udowodnili genetyczną rolę DNA w doświadczeniach nad transformacją mikroorganizmów. Odkrycie to symbolizowało początek nowego etapu - narodziny genetyki molekularnej.
1946 – J. Ledenberg, E. Tatum, M. Delbrück (1906–1981) opisują rekombinację genetyczną u bakterii i wirusów.
1947 - B. Mack - Clintock (1902–1992) jako pierwszy opisał migrujące elementy genetyczne (to wybitne odkrycie zostało nagrodzone Nagrodą Nobla dopiero w 1983 r.).
1950 – E. Chargaff wykazał zgodność nukleotydów purynowych i pirymidynowych w cząsteczce DNA (reguła Chargaffa) i jej specyficzność gatunkową.
1951 – J. Lederberg wraz z kolegami odkrył zjawisko transdukcji, które później odegrało kluczową rolę w rozwoju inżynierii genetycznej.
1952 – A. Hershey (1908–1997) i M. Chase wykazali decydującą rolę DNA w infekcji wirusowej, co ostatecznie potwierdziło jego znaczenie genetyczne.
1953 – D. Watson i F. Crick zaproponowali strukturalny model DNA. Ta data jest brana pod uwagę początek ery współczesnej biologii.
1955 – wyróżnienie S. Ochoa (1905–1993). Polimeraza RNA i jako pierwszy zsyntetyzował RNA in vitro.
1956 – A. Kornberg wyizolował enzym Polimeraza DNA i przeprowadził proces replikacji DNA w warunkach laboratoryjnych.







