WYKŁAD 15. EWOLUCJA ONTOGENEZY, ZWIĄZEK Z - I
Pytania wzmacniające materiał.
1. Czym jest specjacja?
2. Główne ścieżki i metody specjacji.
3. Zasada założyciela, z czego wynika jej działanie?
ROZDZIAŁ 4 PROBLEMY MAKROEWOLUCJI.
1 Pojęcie makroewolucji, podobieństwa i różnice między mikro- i makroewolucją.
2 Ogólne widoki o ontogenezie i ewolucji ontogenezy.
3 Prawo biogenetyczne, podsumowanie, doktryna filembryogenezy.
4 Zasady transformacji narządów i funkcji.
1 Pojęcie makroewolucji, podobieństwa i różnice między mikro- i makroewolucją. W czasach Karola Darwina i w późniejszej epoce rozkwitu jego nauczania ewolucyjnego prawie nic nie wiedziano o dwóch takich podstawowych zjawiskach życia i najbardziej ogólna charakterystyka organizmy żywe na Ziemi jako dziedziczność i zmienność. Zjawiska dziedziczności i zmienności organizmów żywych były ludziom znane, jednak nie istniały naukowe pomysły na temat natury i mechanizmów dziedziczenia cech i ich zmienności. Dopiero rozwój współczesnej genetyki z początku XX wieku umożliwił postawienie wystarczająco dokładnych informacji o podstawowych wzorach dziedziczenia oraz zmienności cech i właściwości organizmów jako podstawy nowego – mikroewolucyjnego etapu badań nad biologią. proces ewolucyjny. W epoce rozwoju klasycznego darwinizmu budowanie teorii ewolucji odbywało się na podstawie wyników uzyskanych w różnorodnych dziedzinach biologii przez badaczy, którzy pracowali wyłącznie przy użyciu opisów i metody porównawcze. Umożliwiło to stworzenie dość kompleksowego obrazu głównych etapów i zjawisk procesu ewolucyjnego, a także stworzenie, jako pierwszego przybliżenia, ogólnego schematu filogenezy organizmów żywych. Tym klasycznym kierunkiem rozwoju idei ewolucyjnych jest badanie procesu makroewolucji. Proces makroewolucyjny, w przeciwieństwie do mikroewolucyjnego, obejmuje duże okresy czasu, rozległe terytoria i wszystkie (w tym wyższe) taksony organizmów żywych, a także wszystkie główne ogólne i szczególne zjawiska ewolucji.
Dane z taksonomii, paleontologii, biogeografii, anatomii porównawczej, Biologia molekularna i inne dyscypliny biologiczne umożliwiają zrekonstruowanie z dużą dokładnością przebiegu procesu ewolucyjnego na dowolnym poziomie ponad gatunkiem. Całość tych danych stanowi podstawę filogenetyki – dyscypliny poświęconej wyjaśnianiu cech ewolucji dużych grup organiczny świat. Porównanie przebiegu procesu ewolucyjnego w różne grupy, w różnych warunkach środowiskowych, w różnych środowiskach biotycznych i abiotycznych itp. pozwala zidentyfikować wspólne cechy rozwoju historycznego charakterystyczne dla większości grup. Na poziomie makroewolucyjnym proces mikroewolucji trwa bez zakłóceń w nowo powstałych formach. Zakłócony zostaje jedynie charakter powiązań pomiędzy nowo powstającymi gatunkami. Teraz mogą nawiązywać relacje międzywioskowe. Relacje te mogą wpłynąć na zdarzenie ewolucyjne jedynie poprzez zmianę nacisku i kierunku działania elementarnych czynników ewolucyjnych, czyli poprzez poziom mikroewolucyjny. Zjawiska makroewolucyjne, posiadające ogromne skale czasowe, wykluczają możliwość ich bezpośredniego badania eksperymentalnego. Oznacza to, że ich rezultaty są zrozumiałe jedynie z punktu widzenia mechanizmu ewolucji – z punktu widzenia mikroewolucji. Na poziomie mikroewolucyjnym (wewnątrzgatunkowym) podczas badania ewolucji okazało się możliwe zastosowanie precyzyjnych podejść eksperymentalnych, które pomogły wyjaśnić rolę poszczególnych czynników ewolucyjnych, sformułować wyobrażenia o elementarnej jednostce ewolucyjnej, elementarnym materiale i zjawisku ewolucyjnym.
W latach 30. XX w. W wyniku intensywnego rozwoju genetyki populacyjnej pojawiła się obiektywna możliwość głębszego poznania mechanizmu powstawania nowych cech (adaptacji) i mechanizmu powstawania gatunków, niż było to dotychczas możliwe jedynie na podstawie obserwacji w Natura. Istotną kwestią w tym zakresie była możliwość bezpośredniego eksperymentu w badaniu mechanizmu ewolucji: dzięki zastosowaniu szybko rozmnażających się gatunków organizmów stała się możliwa symulacja sytuacji ewolucyjnych i obserwacja przebiegu procesu ewolucyjnego. W krótkim czasie możliwe stało się zaobserwowanie znaczących zmian ewolucyjnych w badanych populacjach, aż do pojawienia się izolacji reprodukcyjnej pierwotnej formy.
2 Ogólne poglądy na temat ontogenezy i ewolucji ontogenezy.Ontogeneza(gr. ontos - istnienie, geneza - pochodzenie) to indywidualny rozwój organizmów, podczas którego z zapłodnionego jaja rozwija się organizm dorosły (w partenogenezie z niezapłodnionego). U pierwotniaków ontogeneza zachodzi wewnątrz organizacja komórkowa. Termin ten wprowadził E. Haeckel w 1866 roku. Ontogeneza jest integralną właściwością życia, podobnie jak ewolucja i jej produkt. Proces ontogenezy jest realizacją Informacja genetyczna. Ontogeneza jest procesem z góry określonym i w odróżnieniu od ewolucji jest to rozwój według programu (służy za to genotyp danego osobnika), rozwój ukierunkowany na konkretny cel ostateczny, jakim jest osiągnięcie dojrzałości płciowej i reprodukcja. Jednocześnie komplikacja organizacji na przestrzeni pokoleń jest wynikiem procesu ewolucji. Im bardziej złożona organizacja dorosłego organizmu, co jest odzwierciedleniem ewolucji, tym bardziej złożony i długotrwały jest proces jego ontogenezy. W ten sposób indywidualny rozwój i ewolucja okazują się ściśle ze sobą powiązane (ryc. 4). Ontogeneza składa się z etapów (etapy są kolejną cechą ontogenezy): etap embrionalny, rozwój postembrionalny i życie dorosłego organizmu. Duże etapy (okresy) rozwoju można podzielić na mniejsze etapy, podobnie jak w rozwoju embrionalnym kręgowców - blastula, gastrula, neurula. Z kolei etap kruszenia może być
podzielony na stadia składające się z dwóch, czterech, ośmiu lub większej liczby blastomerów. W rezultacie zatraca się idea etapów ontogenezy i pojawia się całkowicie płynny proces indywidualnego rozwoju. Jak widać, ontogeneza jest uporządkowaną sekwencją procesów (A.S. Severtsov, 1987, 2005).
Zmiany ewolucyjne wiążą się nie tylko z powstawaniem i wymieraniem gatunków, transformacją narządów, ale także z restrukturyzacją rozwoju ontogenetycznego. Bez zmian na poszczególnych etapach ontogenezy filogeneza jest nie do pomyślenia. Filogeneza (gr. phyle - plemię, rodzaj, gatunek, geneza - pochodzenie) to historyczny rozwój świata organicznego, różnych grup systematycznych, poszczególnych narządów i ich układów. Wyróżnia się filogenezę grup zwierząt, roślin i filogenezę narządów.
W toku ewolucji obserwuje się integrację organizmu – ustanawianie coraz ściślejszych, dynamicznych powiązań pomiędzy jego strukturami. Zasada ta jest częściowo odzwierciedlona podczas embriogenezy. Ewolucji życia towarzyszy stopniowy wzrost zróżnicowania i integralności ontogenezy, wzrost stabilności ontogenezy podczas ewolucji życia. Organizm w ontogenezie na jakimkolwiek etapie rozwoju nie jest mozaiką części, narządów czy cech. Morfologiczna i funkcjonalna integralność organizmu w jego życiowych przejawach nie budzi żadnych wątpliwości. Już Arystoteles porównując różne organizmy ustalił jedność ich budowy i uzasadnił doktrynę podobieństwa morfologicznego,
wyrażona w położeniu i strukturze narządów u różnych zwierząt (współczesna homologia narządów), rozwinęła ideę relacji między narządami i współzależności w ich strukturze. Poglądy J. Cuviera miały ogromne znaczenie w historii zagadnienia współzależności części ciała. Według jego idei, jak zauważono wcześniej, organizm jest systemem integralnym, którego strukturę określa jego funkcja; poszczególne części i narządy są ze sobą powiązane, ich funkcje są skoordynowane i dostosowane do znanych warunków środowiskowych (zasada korelacji i zasada warunków życia). Charles Darwin jako najbardziej uderzającą cechę procesu ewolucyjnego wskazał adaptację organizmu do środowiska zewnętrznego i skomplikowanie jego struktury. Zauważył, że koordynacja części jest wynikiem historycznego procesu adaptacji organizmu do warunków życia. Następnie wielu naukowców podkreśliło fakt, że organizm zawsze rozwija się jako całość. Jest ich bardzo złożony system połączenia łączące wszystkie części rozwijającego się organizmu w jedną całość. Dzięki obecności tych połączeń, które pełnią rolę głównych, wewnętrznych czynników rozwoju jednostki, z jaja nie powstaje przypadkowy chaos narządów i tkanek, ale systematycznie skonstruowany organizm ze skoordynowanymi ze sobą funkcjonującymi częściami. Cała celowość reakcji organizmu podczas normalnego kontaktu jednej z jego rozwijających się części z drugą wynika z historycznego rozwoju tych relacji, tj. wynik ewolucji całego mechanizmu rozwoju indywidualnego.
Sposoby (sposoby) doskonalenia ontogenezy w procesie ewolucji: 1) pojawienie się nowych stadiów spowodowane tworzeniem się kompleksów adaptacyjnych zapewniających przetrwanie organizmu i osiągnięcie dojrzałości, co prowadzi do komplikacji ontogenezy; 2) wyłączenie niektórych etapów i zaprzestanie występującej na nich eliminacji, czemu towarzyszy wtórne uproszczenie.
Embrionizacja, autonomizacja, kanalizacja ontogenezy. mi Embrionizacja, autonomizacja, a także racjonalizacja są rezultatami ewolucji ontogenezy. Embrionizacja- jest to droga rozwojowa, w której ontogeneza zachodzi pod ochroną błon jajowych, jest dłużej izolowana od środowiska zewnętrznego i ma mniejszą złożoność w organizacji stadiów embrionalnych. Poprzez embrionizację nastąpiła ewolucja od roślin zarodnikowych do nagonasiennych i od nich do okrytozalążkowych. Przejście od rozwoju larwalnego (u bezkręgowców, ryb, płazów) do składania dużych jaj chronionych gęstymi błonami (u gadów, ptaków), do rozwoju wewnątrzmacicznego, żyworodności (u ssaków) jest wynikiem embrionizacji. Embrionizacja przejawia się w opiece nad potomstwem – wysiadywaniu jaj, wydawaniu potomstwa, budowaniu gniazd, przekazywaniu potomstwu indywidualnych doświadczeń, ochronie nasienia przez jajnik, owoc. Przejawia się to w uproszczeniu cykli rozwojowych – jest to przejście od rozwoju z metamorfozą do rozwoju bezpośredniego, do neotenii. Autonomia przejawia się w rosnącej niezależności ontogenezy od wpływów zewnętrznych i wewnętrznych; ta droga ewolucji tworzy ciągłość form w procesie ewolucyjnym. Autonomia rozwoju jednostki wynika z działania stabilizującej selekcji. Racjonalizacja jest usprawnienie procesu poprzez jego uproszczenie.
Jedna z tendencji ewolucji prowadzi do kanalizacji ontogenezy (I.I. Shmalgauzen, K. Waddington i in.). Głównym czynnikiem aktywnym jest w tym przypadku dobór naturalny, działający w formie doboru kanalizującego. Determinuje pojawienie się „standardowego” fenotypu w najbardziej zróżnicowanych, zmiennych warunkach środowiska wewnętrznego i zewnętrznego.
Ogólnie rzecz biorąc, ewolucja ontogenezy ma pewne cechy, przebiega określonymi ścieżkami, prowadzi do ważnych wyników i jest powiązana z filogenezą, co znajduje odzwierciedlenie w prawie biogenetycznym (o tym będzie mowa dalej).
Znaczenie korelacji i koordynacji. W procesie ontogenezy następuje różnicowanie organizmu (podział całości na części) i jego integracja (połączenie części w jedną całość). Odbywa się to za pomocą tego samego mechanizmu - interakcji rozwijających się podstaw. W ontogenezie nakładają się na siebie kolejno trzy fale zależności korelacyjnych: korelacja genomowa, morfogenetyczna i ergontyczna. Korelacje genomowe– korelacje oparte na interakcji genów, wyrażające się w zjawiskach łączenia genów i plejotropii (wpływ jednego genu na kształtowanie się różnych cech). Korelacje morfogenetyczne– interakcje zawiązków rozwojowych w oparciu o funkcjonowanie genów. Każde różnicowanie rozwijających się zawiązków poprzedzone jest różnicowaniem genetycznym, które wyraża się w zróżnicowanej represji i derepresji genów. Korelacje Ergontyczne– zmiany korelacyjne w narządach względem siebie. Przykładem jest zwiększony rozwój kości, tworzenie się na nich grzbietów w miejscach przyczepu mięśni.
Koordynacja oznaczają współzależności w procesach przemian filogenetycznych. Historycznie rzecz biorąc, rozwijają się na podstawie dziedzicznych zmian w częściach, połączone przez system korelacje, tj. nieunikniona zmiana tego ostatniego lub na innej podstawie - dziedziczna zmiana części niepowiązanych bezpośrednio korelacjami. Jeśli organizm stanowi spójną całość, to nawet przy zmianach swojej struktury w procesie ewolucji musi zachować znaczenie spójnej całości. Wiąże się to ze skoordynowaną zmianą części i narządów. Przykładów koordynacji jest wiele. Są to zależności w zmianach wielkości, kształtu czaszki oraz wielkości i kształtu mózgu – w procesie ewolucji wykształciła się bardzo precyzyjna zgodność kształtu i wielkości tych narządów. Koordynacja to związek między względną wielkością oczu a kształtem czaszki - wzrost wielkości oczu wiąże się ze wzrostem wielkości oczodołów. Koordynacja obejmuje związek pomiędzy stopniem rozwoju zmysłów (węchu, dotyku itp.) a stopniem rozwoju odpowiednich ośrodków i obszarów mózgu. Istnieją koordynacje między narządami wewnętrznymi jako związek między postępującym rozwojem mięśnia piersiowego, serca i płuc u ptaków. U zwierząt kopytnych zachodzi bardzo prosta koordynacja biologiczna pomiędzy długościami kończyn przednich i tylnych.
3 Prawo biogenetyczne, podsumowanie, doktryna filembryogenezy. Po raz pierwszy związek między ontogenezą a filogenezą ujawnił K. Baer w szeregu przepisów, którym Karol Darwin nadał ogólną nazwę „Prawo podobieństwa zarodkowego”. W zarodku naszych potomków – pisał Karol Darwin – widzimy „niejasny portret” naszych przodków. Świetne podobieństwo różne rodzaje w obrębie typu wykrywa się już we wczesnych stadiach embriogenezy. Dzięki temu historię danego gatunku można prześledzić poprzez rozwój indywidualny. W 1864 r. F. Muller sformułował stanowisko, że przemiany filogenetyczne wiążą się ze zmianami ontogenetycznymi i że związek ten objawia się dwojako. W pierwszym przypadku indywidualny rozwój potomków przebiega podobnie jak rozwój przodków tylko do czasu pojawienia się nowej cechy w ontogenezie. Zmiany w procesach morfogenezy powodują powtórzenie w rozwoju embrionalnym historii przodków dopiero w Ogólny zarys. W drugim przypadku potomkowie powtarzają cały rozwój swoich przodków, ale pod koniec embriogenezy dodawane są nowe etapy. F. Müller nazwał powtarzaniem cech dorosłych przodków w embriogenezie potomków rekapitulacją. Prace F. Müllera stały się podstawą do sformułowania przez E. Haeckela (1866) prawa biogenetycznego, zgodnie z którym „ontogeneza jest krótkim i szybkim powtórzeniem filogenezy”. Podstawą prawa biogenetycznego, a także rekapitulacji, jest empiryczna prawidłowość odzwierciedlona w prawie podobieństwa zarodkowego K. Baera. Jego istota jest następująca: najwcześniejszy etap zachowuje znaczne podobieństwo z odpowiednimi etapami rozwoju powiązanych form. Zatem proces ontogenezy polega na pewnym powtórzeniu (rekapitulacji) wielu cech strukturalnych form przodków, na wczesnych etapach rozwoju - bardziej odległych przodków, a na późniejszych etapach - form bardziej spokrewnionych.
Obecnie zjawisko rekapitulacji interpretowane jest szerzej jako sekwencja etapów embriogenezy, odzwierciedlająca historyczną sekwencję przemian ewolucyjnych danego gatunku. Rekapitulację tłumaczy się złożonością zależności, szczególnie we wczesnych fazach rozwoju, oraz trudnością w przebudowie systemu współzależności pomiędzy procesami formacyjnymi. Podstawowym zaburzeniom embriogenezy towarzyszą śmiertelne konsekwencje. Rekapitulacje okazują się najpełniejsze w tych organizmach i tych układach narządów, w których zależności morfogenetyczne osiągają szczególnie dużą złożoność. Dlatego najlepsze przykłady rekapitulacji można znaleźć w ontogenezie wyższych kręgowców.
Filembriogeneza to zmiany, które zachodzą w różne momenty ontogeneza, prowadząca do przemian filogenetycznych (filembryogeneza – ewolucyjne przemiany organizmów poprzez zmianę przebiegu rozwoju embrionalnego ich przodków, prowadzące do pojawienia się nowych cech u organizmów dorosłych). Twórcą teorii filembryogenezy jest A.N. Severtsov. Według jego poglądów ontogeneza ulega całkowitej restrukturyzacji w procesie ewolucji. Nowe zmiany często pojawiają się w ostatnich stadiach formacji. Powikłania ontogenezy spowodowane dodawaniem lub dodawaniem etapów nazywane są anabolią. Rozszerzenie dodaje nowe cechy strukturalne narządów i następuje ich dalszy rozwój. W tym przypadku istnieją wszystkie przesłanki do powtórzenia w ontogenezie historycznych etapów rozwoju tych części wśród odległych przodków. Dlatego właśnie podczas anabolizmu przestrzegane jest podstawowe prawo biogenetyczne. Na późniejszych etapach rozwoju zwykle zachodzą zmiany w budowie szkieletu kręgowców, zmiany w różnicowaniu mięśni i rozmieszczeniu naczyń krwionośnych. W wyniku anabolizmu u ptaków i ssaków powstaje czterokomorowe serce. Przegroda między komorami stanowi przedłużenie, powstaje w ostatnich stadiach rozwoju serca. W wyniku anabolizmu w roślinach pojawiły się rozcięte liście. Ontogeneza może jednak zmieniać się już na środkowych etapach rozwoju, odbiegając jednocześnie od dotychczasowej ścieżki wszystkie późniejsze etapy. Ten sposób zmiany ontogenezy nazywa się dewiacją. Odchylenie prowadzi do restrukturyzacji narządów, które istniały u przodków. Przykładem odchylenia jest tworzenie się rogowych łusek gadów, które początkowo tworzą się jak placoidalne łuski rekinów. Następnie u rekinów zaczynają intensywnie rozwijać się tkanki łącznej w brodawkach, a u gadów zaczyna rozwijać się część naskórkowa. W wyniku odchylenia powstają kolce, a pędy przekształcają się w bulwę lub cebulkę. Oprócz wskazanych sposobów (metod) zmian w ontogenezie możliwa jest także zmiana podstaw samych narządów lub ich części – droga ta nazywana jest archalaksją. Dobry przykład jest to rozwój włosów u ssaków. Poprzez archalaksję zmienia się liczba kręgów, liczba zębów u zwierząt itp. Archalaksja wystąpiła, gdy liczba pręcików podwoiła się, co jest początkiem jednoliścienności u roślin. Rozważane zmiany ewolucyjne w ontogenezie przedstawiono na rysunkach 4, 5.
Zasadnicze znaczenie teorii phylembryogenezy polega na tym, że wyjaśnia ona mechanizm ewolucji ontogenezy, mechanizm ewolucyjnych przemian narządów, powstawanie nowych cech w ontogenezie oraz wyjaśnia fakt rekapitulacji. Filembryogeneza jest wynikiem dziedzicznej restrukturyzacji aparatów formatywnych, zespołu dziedzicznie zdeterminowanych adaptacyjnych przekształceń ontogenezy.
Integralność ciała, wielofunkcyjność. Stanowisko w sprawie integralności ciała zostało szczegółowo omówione powyżej. Należy jednak zauważyć, że jednocześnie z tą cechą organizm charakteryzuje się autonomią poszczególnych narządów. Stanowisko to potwierdza zjawisko wielofunkcyjności oraz możliwości jakościowych i ilościowych zmian funkcji. Przekształcenia filogenetyczne narządów i ich funkcji mają dwa przesłanki: każdy narząd charakteryzuje się wielofunkcyjnością, a funkcje charakteryzują się zdolnością do zmian ilościowych. Kategorie te leżą u podstaw zasad ewolucyjnych zmian w narządach i ich funkcjach. Wielofunkcyjność narządów polega na tym, że każdy narząd oprócz swojej charakterystycznej funkcji głównej pełni także szereg drugorzędnych. Więc, główna funkcja liść - fotosynteza, ale dodatkowo pełni funkcje uwalniania i wchłaniania wody, narządu magazynującego, narządu rozrodczego itp. Przewód pokarmowy zwierząt to nie tylko narząd trawienny, ale także najważniejsze ogniwo w łańcuchu narządów wydzielania wewnętrznego, ważne ogniwo w układzie limfatycznym i krwionośnym. Ta sama funkcja może objawiać się w organizmach z większą lub mniejszą intensywnością, dlatego wszelkie formy aktywności życiowej mają nie tylko cechy jakościowe, ale także ilościowe. funkcja biegania,
na przykład wyraża się silniej u niektórych gatunków ssaków, a mniej u innych. W przypadku każdej właściwości zawsze istnieją różnice ilościowe pomiędzy osobnikami gatunku. Każda z funkcji organizmu zmienia się ilościowo w procesie indywidualnego rozwoju jednostki.
4 Zasady transformacji narządów i funkcji. Znanych jest ponad półtora tuzina metod ewolucji narządów i funkcji oraz zasad ich transformacji. Najważniejsze z nich zostały wymienione poniżej.
1) Zmiana funkcji: gdy zmieniają się warunki bytu, funkcja główna może stracić na znaczeniu, a jedna z drugorzędnych może zyskać znaczenie głównej (podział żołądka u ptaków na dwie - gruczołową i mięśniową).
2) Zasada rozszerzania funkcji: często towarzyszy postępującemu rozwojowi (trąba słonia, uszy słonia afrykańskiego).
3) Zasada funkcji zawężających (płetwy wielorybów).
4) Wzmocnienie, czyli intensyfikacja funkcji: związane z postępującym rozwojem narządu, jego większą koncentracją (postępujący rozwój mózgu ssaków).
5) Aktywacja funkcji - przekształcenie narządów biernych w aktywne (jadowity ząb u węży).
6) Unieruchomienie funkcji: przekształcenie narządu czynnego w bierny (utrata ruchomości górnej szczęki u niektórych kręgowców).
7) Rozdzielenie funkcji: towarzyszy podział narządu (na przykład mięśnia, części szkieletu) na niezależne sekcje. Przykładem jest podział niesparowanej płetwy ryby na sekcje i związane z tym zmiany funkcji poszczególnych części. Odcinki przednie - płetwy grzbietowa i odbytowa - stają się sterami kierującymi ruchem ryby, odcinek ogonowy jest głównym narządem motorycznym.
8) Utrwalenie faz: zwierzęta stepowe podczas chodzenia i biegania podnoszą się na palcach; w tej fazie ustala się chód cyfrowy u zwierząt kopytnych.
9) Substytucja narządów: w tym przypadku dochodzi do utraty narządu i jego funkcję pełni inny (zastąpienie cięciwy kręgosłupem).
10) Symulacja funkcji: narządy, które wcześniej różniły się formą i funkcją, upodobniają się do siebie (u węży podobne segmenty ciała powstały w wyniku symulacji ich funkcji).
11) Zasady oligomeryzacji i polimeryzacji. Podczas oligomeryzacji zmniejsza się liczba narządów homologicznych i funkcjonalnie podobnych, czemu towarzyszą zasadnicze zmiany w korelacyjnych połączeniach między narządami i układami. Tak, ciało pierścienice składa się z wielu powtarzających się segmentów, u owadów ich liczba jest znacznie zmniejszona, a u wyższych kręgowców nie ma wcale identycznych segmentów ciała. Polimeryzacji towarzyszy wzrost liczby organelli i narządów. Ona miała bardzo ważne w ewolucji pierwotniaków. Ta ścieżka rozwoju doprowadziła do pojawienia się kolonii, a następnie do pojawienia się wielokomórkowości. Zwiększenie liczby jednorodnych narządów wystąpiło także u zwierząt wielokomórkowych (np. węży). Podczas ewolucji oligomeryzacja została zastąpiona polimeryzacją i odwrotnie.
Należy zauważyć, że każdy organizm jest skoordynowaną całością, w której poszczególne części znajdują się w złożonym podporządkowaniu i współzależności. Jak zauważono powyżej, dobrze zbadana jest współzależność poszczególnych struktur (korelacja) w procesie ontogenezy, a także korelacje pojawiające się w procesie filogenezy i nazywane są koordynacjami. Złożoność powiązań ewolucyjnych narządów i układów jest widoczna, gdy analizujemy zasady transformacji narządów i funkcji. Zasady te pozwalają lepiej wyobrazić sobie ewolucyjne możliwości przekształcania danej organizacji w różnych kierunkach, pomimo ograniczeń narzucanych przez korelacje.
Tempo ewolucji poszczególnych cech i struktur, a także tempo ewolucji form (gatunków, rodzajów, rodzin, rzędów itp.) determinuje tempo ewolucji całości, co należy uwzględnić w praktyce działalność człowieka. Przykładowo stosując środki chemiczne należy wiedzieć jak szybko dany gatunek może uodpornić się na leki: leki u ludzi, środki owadobójcze u owadów itp. Tempo ewolucji poszczególnych cech w populacjach, a także tempo ewolucji całych gatunków struktury i narządy, zależy od wielu czynników: liczby populacji w obrębie gatunku, zagęszczenia osobników w populacjach, oczekiwanej długości życia pokoleń. Wszelkie czynniki będą miały przede wszystkim wpływ na tempo zmian populacji i gatunków poprzez zmiany nacisku elementarnych czynników ewolucyjnych.
Zasada antropiczna i globalny ewolucjonizm
I tak kilkadziesiąt lat temu B. Carter wysunął tzw. zasadę antropiczną (AP), stwierdzając istnienie związku pomiędzy parametrami Wszechświata a istnieniem w nim inteligencji. Formalny impuls do rozpoczęcia dyskusji o miejscu człowieka we Wszechświecie dało omówienie problemu koincydencji dużych liczb – dziwnej zależności liczbowej pomiędzy parametrami mikroświata (stała Plancka, ładunek elektronu, nukleon rozmiar) i globalną charakterystykę Metagalaktyki (jej masa, rozmiar, czas życia). Problem ten zrodził pytanie: jak losowe są parametry naszego świata, jak są ze sobą powiązane i co się stanie, jeśli nieznacznie się zmienią? Analiza możliwych zmian podstawowych parametrów fizycznych wykazała, że nawet niewielka zmiana w nich prowadzi do niemożności istnienia naszej Metagalaktyki w obserwowalnej formie i jest niezgodna z pojawieniem się w niej życia, a co za tym idzie, inteligencji.
Związek pomiędzy parametrami Wszechświata a pojawieniem się w nim inteligencji Carter wyraził w dwóch sformułowaniach – mocnym i słabym. Słabe AP stwierdza jedynie, że warunki istniejące we Wszechświecie nie zaprzeczają istnieniu człowieka: „Nasza pozycja we wszechświecie jest z konieczności uprzywilejowana w tym sensie, że musi być zgodna z naszym istnieniem jako obserwatorów”.(Carter B. Zbieżność dużych liczb i zasada antropologiczna w kosmologii // Kosmologia: teoria i obserwacje. M., 1978). Silne AP proponuje sztywniejszą zależność pomiędzy parametrami Wszechświata a możliwością i koniecznością pojawienia się w nim inteligencji: „Wszechświat (a zatem podstawowe parametry, od których zależy) musi być taki, aby na pewnym etapie jego ewolucji dozwolone było istnienie obserwatorów”..
Można sformułować dwa skrajne założenia uzasadniające AP:
1) inteligencja w naszej Metagalaktyce jest zjawiskiem absolutnie przypadkowym, które stało się możliwe jedynie dzięki nieprawdopodobnej, ale uświadomionej zbieżności wielu niezależnych parametrów fizycznych;
2) obecność biologicznych i społecznych form ruchu jest naturalną konsekwencją rozwoju Wszechświata i całości Charakterystyka fizyczna wzajemnie powiązane i współzależne w taki sposób, że z konieczności powodują pojawienie się rozumu.
Probabilistyczna hipoteza wyjaśniająca AP sprowadza ją z pozycji globalnej heurystycznej idei związku pomiędzy pojawieniem się inteligentnego życia a strukturą Wszechświata do całkowicie przypadkowego zbiegu okoliczności, co w istocie wykracza poza zakres rozważania naukowe. Hipotezę probabilistyczną można jednak traktować jedynie jako terminologiczną – zamiast pytania o możliwość pojawienia się obserwatora we Wszechświecie, stawiane jest pytanie o jego możliwe pojawienie się w całości Wszechświatów. I opierając się na koncepcji kategorii „Wszechświat” musimy przyznać, że jest ona absolutnie identyczna z terminem „całość wszechświatów”. Można jedynie rozróżnić warianty jedno- lub wielokrotno zdefiniowanego Wszechświata, co oczywiście nie jest istotne z punktu widzenia rozwiązania problemu możliwości i konieczności pojawienia się inteligencji we Wszechświecie.
Hipoteza oscylującego Wszechświata, jako możliwe wyjaśnienie AP, nie różni się od założenia o wielości Wszechświatów. Nieskończoność numeryczną zastępuje się nieskończonością czasową, ponownie redukując wyjaśnienie AP do absolutnie przypadkowego momentu.
Kolejną słomką, jaką teoria oscylującego Wszechświata oferuje dla formalnego wsparcia AP, jest stwierdzenie o możliwości przekazania informacji o parametrach Wszechświata w momencie osobliwości. Oznacza to, że zakłada się, że cechy fizyczne Metagalaktyki nie są przypadkowe, a ich wartości są zdeterminowane przez poprzedni cykl jej rozwoju (przed osobliwością). Całkiem logiczne i możliwe. Ale znowu ta hipoteza, spychająca rozwiązanie problemu na hipotetyczne poprzednie etapy historii Wszechświata, nie przybliża nas do naukowej interpretacji AP.
Rozważmy jedynie drugą, nieprobabilistyczną opcję uzasadnienia AP, która bezpośrednio wpisuje się w szeroko dyskutowany obecnie problem globalnego ewolucjonizmu. Ogólnie rzecz biorąc, ewolucyjne podejście do problemu AP można sformułować następująco: Wszechświat znajduje się w ciągłym procesie ewolucji, a pojawienie się życia i inteligencji jest naturalnym rezultatem tego procesu. Mając wystarczająco przekonującą konstrukcję teorii globalnego ewolucjonizmu, AP należy sprowadzić do jej szczególnego momentu - stwierdzenia, że rozum jest niezbędnym etapem ewolucji Wszechświata.
Ewolucja. NA ten moment terminu „ewolucja” używa się albo w wąskim znaczeniu – do opisania procesów powstawania i rozwoju systemy biologiczne lub w najszerszym znaczeniu, kiedy mówimy ogólnie o pojawieniu się nowych właściwości, definicji, formacji strukturalnych. Naturalnie, gdy będziemy dalej używać terminu „ewolucja” w dyskusji nad problemem globalnego ewolucjonizmu, nie będziemy mieli na myśli jedynie jego szczególnego znaczenia biologicznego. I oczywiście, aby uniknąć problemów terminologicznych, należy zwrócić szczególną uwagę na rozszerzoną interpretację terminu „ewolucja”, na przykład przy opisie procesów astronomicznych lub geologicznych. Często stajemy przed problemem terminologicznym użycia kategorii „ewolucja” zamiast kategorii „rozwój” i „zmiana”, choć rozróżnienie pomiędzy tymi trzema kategoriami dobrze wpisuje się w naturalne rozumienie znaczenia tych słów.
Mówiąc więc o ewolucji, mamy na myśli pojawienie się zasadniczo nowych, unikalnych definicji (parametrów, kategorii, systemów itp.), które wcześniej nie były dostępne. Co więcej, aby jednoznacznie odróżnić kategorię „ewolucja” od kategorii „rozwój”, nowość definicji musi być fundamentalna nie tylko dla rozpatrywanego systemu, ale dla całego Świata.
Rozwój zauważa pojawienie się nowych funkcji, które nie były wcześniej nieodłączne od określonego systemu, ale nie są unikalne dla świata jako całości. Zatem pojawienie się umysłu jest zjawiskiem ewolucyjnym, ale zmiany zachodzące podczas narodzin i wzrostu indywidualnej osoby należy oczywiście opisać terminem „rozwój”.
Zmiana kategorii opisuje procesy, które zachodzą bez pojawienia się nowych definicji. Właściwsze zatem jest opisywanie procesów zachodzących we współczesnych obiektach astronomicznych w kategoriach „zmiany” i „rozwoju”, a nie „ewolucji”. Ewolucyjne powstawanie obserwowanych dziś formacji kosmologicznych nastąpiło we wczesnych stadiach historii Wszechświata, a obecnie obserwujemy jedynie zmianę ich parametrów. To samo można powiedzieć o układach geologicznych - ich ewolucyjne powstawanie miało miejsce we wczesnych stadiach formowania się naszej planety, a obecnie obserwujemy jedynie zmiany w obiektach geologicznych, ale nie ich ewolucję.
Powyższe wyjaśnienia terminologiczne usuwają wiele problemów stojących na drodze do rozszerzenia teorii ewolucji na obiekty niebiologiczne. I następują proste wnioski.
Po pierwsze: różne systemy (rodzaje ruchu) można uznać za ewolucyjne dopiero na etapie ich powstawania, któremu towarzyszy pojawienie się zasadniczo nowych jakości i struktur, nowych nie tylko dla tego systemu, ale także dla Świata jako całości. Po zakończeniu tego etapu systemy nadal się zmieniają, ale nie ma to charakteru ewolucyjnego. I drugi wniosek: tylko Świat jest obiektem absolutnie ewolucyjnym.
W tym miejscu dyskusji zasadne jest rozważenie kolejnego pytania terminologicznego - czy powinniśmy mówić o ewolucji Metagalaktyki, czy Wszechświata? Właściwsze byłoby użycie określenia Wszechświat, co spycha na dalszy plan problem wielości metagalaktyk. Chociaż chyba bardziej poprawne jest użycie terminu Świat w ogóle, który nie ma konotacji kosmologicznej, ale zasadniczo pokrywa się z kategorią Wszechświata i wyraźniej obejmuje zarówno biologiczne, jak i społeczne formy ruchu. To właśnie zrobiliśmy powyżej, ale także nie unikając terminów Metagalaktyka i Wszechświat, uznając je wszystkie za synonimy, pomijając istniejące różnice jako nieistotne w kontekście tych notatek.
I jeszcze jedna uwaga czysto terminologiczna – celowe wykluczenie ze słownika kategorii „materia” i „duch”. Dokonano tego ze względu na brak możliwości podania ich prawidłowej definicji w ramach krótkich notatek, a także po to, aby nie powodować nieuniknionych – powtarzam czysto terminologicznie – problemów. Choć dla tych, którym jest łatwiej, termin „rodzaj ruchu” można odczytać jako „formę materii”, a przez „ewolucję Wszechświata” mamy na myśli „rozwój Ducha świata”. Warstwa omawianych tu problemów nie leży na poziomie rozróżnienia tych kategorii.
Oprócz terminologicznych istnieje szereg dość istotnych problemów, które wiążą się z trudnościami w opracowaniu podejścia ewolucyjnego. Tutaj wyrazimy je w formie tez, które zostaną doprecyzowane w dalszych dyskusjach. A więc problemy:
Strukturalny - zastosowanie terminu ewolucja do elementów systemów ewolucyjnych, chociaż zmiana właściwości tych elementów jest wtórna w stosunku do rzeczywistych procesów ewolucyjnych w całym systemie.
Ekstrapolacja jest próbą przeniesienia zasad zmian ewolucyjnych z jednego układu na inny (na przykład zasady zmienności i selekcji w układach biologicznych na ewolucję innych rodzajów ruchu).
Integracja – poszukiwanie wspólnych wzorców funkcjonowania systemów ewolucyjnych, próba formalnego połączenia teorii w jedną.
Globalny ewolucjonizm. Pojawienie się koncepcji globalnego ewolucjonizmu (GE) w dużej mierze wiąże się nie tylko z problemem uzasadnienia zasady antropicznej, ale także, oczywiście, z poszerzeniem granic podejścia ewolucyjnego przyjętego w biologii i biologii. nauki społeczne. Sam fakt historycznego pojawienia się i ewolucji tego rodzaju ruchu poddaje w wątpliwość absolutną statyczność i wieczność innych rodzajów ruchu. Tajemnicę skoków jakościowych do świata biologicznego i ze świata biologicznego do społecznego można z pewnością zrozumieć jedynie przy założeniu konieczności podobnych przejść pomiędzy innymi rodzajami ruchu. Czyli na podstawie faktu ewolucji Świata na ostatnich etapach jego historii możemy tak założyć Świat jako całość jest systemem ewolucyjnym, to znaczy wszystkie inne rodzaje ruchu (oprócz biologicznego i społecznego) powstały w wyniku ewolucji. To stwierdzenie jest najbardziej ogólnym sformułowaniem paradygmatu globalnego ewolucjonizmu.
Zauważ, że wyrażenie „globalna ewolucja” nie oznacza żadnego innego pojęcia niż po prostu „ewolucja” (o której mówiliśmy w poprzednim akapicie). Heurystyczny charakter wprowadzenia dodatkowego terminu polega na rozszerzeniu poglądu ewolucyjnego na wszystkie rodzaje ruchu, na cały Wszechświat, na wszystkie etapy jego historii, łącznie z momentem jego Początku. Co więcej, poszczególne procesy ewolucyjne (na przykład biologiczne czy społeczne) nie są elementami podrzędnymi ewolucji globalnej, lecz istotą ewolucji, jej konkretnymi realizacjami na określonych etapach historii Wszechświata.
Ewolucyjne podejście do istniejących rodzajów ruchu nie oznacza, że wszystkie one znajdują się w ciągłym procesie ewolucji, lecz wręcz przeciwnie, stwierdza potrzebę ich konsekwentnego kształtowania się na określonych etapach historii Świata. Generalnie ewolucja, jako proces, który w każdym momencie dotyczy całego Wszechświata, realizuje się lokalnie tylko w jednym rodzaju ruchu. Oznacza to, że zawsze istnieje tylko jeden system lokalny (czyli nie identyczny z całym Światem), który można nazwać ewolucyjnym, w którym pojawiają się zasadniczo nowe, unikalne definicje Świata. Aby odróżnić ten system od innych, które przeszły już ewolucję, możemy wprowadzić termin „awangarda ewolucji”. Naturalnie awangardą ewolucji jest zawsze najnowsza forma ruchu, która pojawiła się na świecie (obecnie System społeczny). Wszystkie poprzednie rodzaje ruchów, które przeszły przez etap ewolucyjny i osiągnęły stan równowagi (nie statyczny, ale najprawdopodobniej stan powolnych zmian parametrów lub powtarzający się proces rozwoju poszczególnych elementów) służą jako podstawa do powstawania i ewolucja nowego ruchu. Możliwe, że w „poprzednich” ruchach pojawią się nowe cechy, ale z konieczności są one związane z ewolucją najnowszego typu ruchu (systemu) – awangardy ewolucji.
Zatem w każdym momencie historii Wszechświata można wyróżnić tylko dwa systemy ewolucyjne - cały Świat i rodzaj ruchu, który jest obecnie awangardą ewolucji. Sam proces ewolucyjny jest w nich absolutnie zbieżny, to znaczy pojawienie się nowych cech w danym systemie jest specyficzną realizacją ewolucji Świata. Globalny ewolucjonizm Świata różni się od ewolucji poszczególnych systemów jedynie swoją czasową ciągłością, przeniesieniem procesu ewolucji z jednego rodzaju ruchu na drugi. Natomiast proces ewolucyjny w oddzielnym układzie koniecznie kończy się po osiągnięciu pewnego stanu równowagi, a ewolucja trwa w kolejnej formie ruchu.
W każdym systemie ewolucyjnym (w każdym typie ruchu w momencie, gdy był on awangardą ewolucji) można wyróżnić parametr ewolucyjny - parametr, który oczywiście ulega ciągłym zmianom i wiąże się z pojawieniem się zasadniczo nowych definicji tego rodzaj ruchu. Zatem w ruchu społecznym jest to proces rozwoju naukowo-technicznego - ma charakter ciągły, nieodwracalny i determinuje zmiany innych parametrów ruchu społecznego. Również mówiąc o procesach ewolucyjnych w odrębnym systemie należy zwrócić uwagę na to, że dotyczą one całego systemu jako całości i niedopuszczalne jest mechaniczne przenoszenie ich na elementy systemu. A więc odkrywanie ewolucja społeczna należy mieć na uwadze system społeczny jako całość, a nie rozwój i upadek poszczególnych państw itp. (choć oczywiście wszystkie procesy w systemie społecznym mają znaczenie funkcjonalne, ich całość stanowi sam ruch społeczny, ale element ewolucyjny ruchu należy wyraźnie oddzielić od poszczególnych zmian i rozwoju poszczególnych elementów).
Jeden z punktów krytyki GE wiąże się z absolutną odmiennością procesów ewolucyjnych w systemach biologicznych i społecznych oraz występowaniem procesów kosmologicznych i geologicznych. Odnotowuje się różne ich kierunki i skutki – niektóre zmierzają w stronę komplikowania organizacji strukturalnej, czego nie można powiedzieć o tej drugiej. Na tę krytykę można najogólniej odpowiedzieć, opierając się na zrozumieniu czasowości ewolucyjnego rozwoju konkretnego systemu. System geologiczny był awangardą ewolucji na bardzo specyficznym etapie ewolucji Świata. To właśnie w tym okresie miały miejsce ewolucyjne procesy powstawania nowych dla świata obiektów i struktur geologicznych. A dalsze procesy rozkładu obiektów geologicznych nie mają nic wspólnego z procesem ewolucji. To samo można powiedzieć o obiektach kosmologicznych. Powstawanie astronomicznych elementów struktury Świata nastąpiło we wczesnych stadiach ewolucji Wszechświata, a współczesne procesy modyfikacji obiektów astronomicznych nie mają charakteru ewolucyjnego. I tylko w ostatnim historycznie rodzaju ruchu (awangardzie ewolucji) WE realizuje się w specyficznym procesie ewolucyjnym. (Mając na uwadze powyższe argumenty, możemy przyjąć założenie, że współczesne eksperymenty naukowe dotyczące syntezy układów żywych są z pewnością skazane na niepowodzenie. Pojawienie się pierwotnych oznak układów biologicznych było możliwe dopiero na określonym etapie ewolucji Wszechświata , ze specyficznymi parametrami fizycznymi, które są naturalnie nieosiągalne w tym momencie.)
Zanim przejdziemy do sformułowania tezy o naszym rozumieniu tego, czym powinna być ogólna teoria globalnego ewolucjonizmu, wróćmy do problemu AP, patrząc na niego ze stanowiska GE.
Zasada antropiczna ze stanowiska globalnego ewolucjonizmu. Silne AP z punktu widzenia GE jedynie stwierdza i postuluje ewolucję Wszechświata. Oznacza to, że mocne AP wysunięte przez Cartera w sformułowaniu: „Wszechświat (a w konsekwencji podstawowe parametry, od których zależy) musi być takie, aby na pewnym etapie ewolucji dopuszczono w nim istnienie obserwatorów” jest równoważne do stwierdzenia wyrażonego ze stanowiska GE: „Wszechświat znajduje się w ciągłym procesie ewolucji, koniecznie prowadzącym na pewnym etapie do wyłonienia się forma społeczna ruch."
AP wyrażone w kategoriach GE pozbawione jest istotnego punktu problematycznego - nie mówi nic o parametrach Wszechświata. Po pierwsze, rozciągnięcie procesu ewolucji Wszechświata w przeszłość przed jego początkiem całkowicie eliminuje kwestię parametrów początkowych - w początkowej chwili nie ma definicji Świata, a pojawiają się wszelkie rodzaje ruchu, a w konsekwencji ich parametry w procesie ewolucji. Po drugie, GE, stwierdzając pojawienie się ruchu społecznego jako niezbędny etap rozwoju Świata, w ogóle nie reguluje formy ucieleśnienia umysłu, co całkowicie eliminuje problem zgodności parametrów fizycznych Wszechświata do pewnej specyficznej formy realizacji „obserwatora”. Oznacza to, że postulując zasadę Wn, rozum we Wszechświecie powstaje koniecznie i niezależnie od określonych parametrów fizycznych „poprzednich” rodzajów ruchu, z wszelkimi odchyleniami (jeśli takie są możliwe). A ponadto z punktu widzenia warunkowości wdrożenia specyficzna forma ruch społeczny nie różni się od innych „poprzednich” rodzajów ruchu (biologicznych, chemicznych itp.).
W związku z tym AP należy sformułować nie jako zasadę zgodności warunków fizycznych i innych możliwości pojawienia się obserwatora, ale wręcz przeciwnie, jako zgodność określonej formy realizacji umysłu z istniejącymi warunkami. I w tym AP sprowadza się jedynie do najbardziej ogólnej formy wyrażenia zasady globalnego ewolucjonizmu.
Aby lepiej zrozumieć interpretację AP w ramach GE, rozważmy na przykład zgodność koloru określonego gatunku zwierzęcia z kolorem skał geologicznych na danym obszarze. Można omówić problem realizacji określonego umaszczenia rasy, bez którego istnienie danego zwierzęcia nie byłoby możliwe – i z jakim prawdopodobieństwem dokładnie takie umaszczenie mogłoby się pojawić, oraz postawić hipotezę o wielości miejscowości i że tylko jeden z nich, zupełnie przez przypadek, jest taki, że odpowiada umaszczeniu zwierzęcia.. Albo po prostu stwierdzić, że zgodność ta jest wynikiem rozwoju gatunku w warunkach danego obszaru.
Rozwijając Słabe i Silne AP, które głoszą, że istnienie inteligencji jest uzależnione od parametrów fizycznych Wszechświata, F. Tipler dodatkowo wysunął Finalistyczne AP w następującym sformułowaniu: „We Wszechświecie musi powstać inteligentne przetwarzanie informacji, a kiedy już się pojawi, nigdy nie ustanie” (Barrow J.D., Tipler F.J. The anthropic cosmological zasada (Oxfotd, 1986). Oznacza to, że finalistyczne AP stwierdza nie tylko związek między pojawieniem się inteligencji a historią Wszechświata, ale także jej bezwarunkowy wpływ na jego dalszą ewolucję.
Podobnie jak w przypadku słabego i mocnego AP, Finalistyczne AP rozpatrywane z punktu widzenia GE jest jedynie stwierdzeniem konsekwentnej ewolucji poszczególnych typów ruchu. Zwraca jedynie uwagę nie na warunkowość pojawienia się określonego systemu ewolucyjnego (w konkretnym przypadku społecznego) przez poprzednie typy ruchu, ale na niezbędny wpływ tego systemu na dalszy proces ewolucji Świata. Z tego punktu widzenia finalistyczna zasada ma oczywiście zastosowanie jednakowo do wszystkich rodzajów ruchu.
Ale możemy też wysunąć mocne finalistyczne AP: wraz z pojawieniem się rozumu, wraz z rozwojem ruchu społecznego, świat wszedł w nowy etap ewolucji, charakteryzujący się aktywnym wpływem jednej z form ruchu na cały system (w ramach tych notatek nie będę definiował etapów ewolucji świata). Koniec Świata z punktu widzenia Wn można rozumieć jako pewien akt synkretycznej jedności wszystkich rodzajów ruchu i powinien on być realizowany właśnie w wyniku rozwoju umysłu, jako akt absolutnej samowiedzy świat. (To stwierdzenie nie ma konotacji teologicznych ani katastroficznych.)
I tak, podsumowując, różne AP ze stanowiska GE można wyrazić następująco:
- Słabe AP: Rozum (system społeczny) jest jednym z rodzajów ruchu Świata;
- Silne AP: Rozum (system społeczny) jest obowiązkowym etapem ewolucji Świata, uwarunkowanym wszystkimi dotychczasowymi rodzajami ruchu;
- Finalistyczne AP: Rozsądna forma poruszania się Świata jest integralnym etapem rozwoju Świata, determinującym jego dalszą ewolucję;
- Mocne Finalistyczne AP: Koniec Świata jest koniecznie związany z rozwojem umysłu i może być interpretowany jako moment absolutnej samowiedzy przez Świat sam w sobie.
DO ogólna teoria globalny ewolucjonizm (OTGE). Ogólna teoria ewolucjonizmu globalnego jest teorią rozciągającą zasadę ewolucji na cały Świat, na wszystkie etapy historii Wszechświata. Głównym początkowym stwierdzeniem OTGE jest postulat ewolucyjnego, sekwencyjnego pojawiania się i dalszej zmiany wszystkich istniejących rodzajów ruchu, od grawitacyjnego do społecznego. A różnica między OTGE a innymi teoriami opisu świata polega właśnie na przyjęciu lub braku akceptacji samego faktu możliwości ewolucji. Skupiamy się na tym zapisie, aby jasno określić ramy teorii i przestrzec przed zastrzeżeniami w poszczególnych kwestiach, jeśli główny postulat nie zostanie przyjęty.
Przede wszystkim mówiąc o ogólnych zasadach konstruowania OTGE, należy wyjść od założenia, że sama teoria, jej istota, jej struktura muszą być adekwatne do badanego, opisywanego obiektu - Świata, który jest w procesie ewolucji . Jakie wnioski można wyciągnąć z tego bardzo ogólnego poglądu?
Po pierwsze, OTGE przede wszystkim musi być systemem ewolucyjnym. Oznacza to, że aby poprawnie opisać wzorce przejść ewolucyjnych, sam musi być systemem samorozwijającym się, wytwarzającym kategorie, pojęcia i prawa bezpośrednio w procesie swojego rozwoju (czyli zapewne musi to być system filozoficzny podobny do Logika dialektyczna Hegla.)
Po drugie, skoro zauważamy, że wszelkie rodzaje ruchu w procesie ewolucji, koniecznie warunkujące powstanie kolejnych układów, nie są do nich redukowane, nie rozpuszczają się w nich, ale w dalszym ciągu funkcjonują jako układy równowagi, to nauki specjalne, które opisują ruchów tych (fizyka, chemia, biologia itp.) nie należy do siebie redukować. To znaczy, tak jak każdy rodzaj ruchu zachowuje swoją specyfikę, swoją niezależność w procesie ewolucji Świata, tak też prawa naukowego opisu poszczególnych rodzajów ruchu nie mogą rozciągać się na poprzednie rodzaje ruchu (a już na pewno na kolejne) ). I dlatego opis naukowyżaden rodzaj ruchu (w szczególności interakcja fizyczna) nie może być uważany za szczególny przypadek(rozwiązanie prywatne) systemu naukowego opisującego kolejny typ (mianowicie takie podejście mające na celu reprezentowanie pewnych teorie naukowe ponieważ szczególne rozwiązania pewnego uogólnionego systemu są praktykowane we współczesnej nauce).
Zatem wnioski:
OTGE w swojej pełnej postaci powinno reprezentować system filozoficzny (logiczny w sensie heglowskim), w którym śledzona jest ewolucja Świata, w postaci spójnego wyprowadzenia współzależnych kategorii (definicji). Różne części (etapy) tego logicznego układu muszą jednoznacznie odpowiadać różnym etapom ewolucji Świata (poszczególnym typom ruchu). System musi zakończyć się wnioskiem o konieczności Końca Świata, jako absolutnego wyniku ewolucji, jako powrotu do synkretycznej jedności wszystkich jego definicji (rodzajów ruchu), powrotu do stanu niepewnego.
OTGE nie zalicza reszty nauki do „określonych rozwiązań”, a jedynie określa ich logiczną relację, wskazując, że w ramach każdego z tych systemów naukowych musi istnieć mechanizm rozwoju nieuchronnie prowadzący do wewnętrznych sprzeczności tego systemu, którymi są rozwiązany dopiero podczas przejścia do następnego etapu, do następnego systemu.
Główny kierunek ewolucji Świata w OTGE rozumiany jest nie jako przejście od chaosu do porządku, lecz wręcz przeciwnie – za punkt wyjścia przyjmuje się stan bezpośredniego, absolutnie nieokreślonego (bez definicji) porządku, od którego zaczyna się ewolucja upadek tego porządku w wyniku pierwszego określenia Świata przez sam fakt Początku. Pojawienie się pierwszej zróżnicowanej definicji Świata powoduje powstanie nowych definicji, wyznaczających nowy porządek, który rozpada się w wyniku korelacji nowych i starych definicji. Ten proces ruchu Świata, poprzez samostanowienie nowego porządku i jego rozkład „pod naciskiem” poprzednich definicji, można nazwać ewolucją.
Tę filozoficzną tyradę można wyrazić innymi słowami. Początek świata można przedstawić jako początek jakiegoś elementarnego ruchu. Zmiana świata w trakcie tego ruchu wyprowadza go z aktualnego stanu równowagi (pierwszym z nich był nieokreślony początek samego Świata). Ciągłość ruchu elementarnego decyduje o nieodwracalności stanów Świata, dlatego też nowy stan równowagi można osiągnąć jedynie poprzez powstanie nowych układów, o nowych parametrach i właściwościach. Zatem ewolucję Świata można przedstawić jako stopniowe pojawianie się nowych stanów równowagi na tle ogólnego ruchu Świata.
Każdy z tych stanów ma charakter dynamiczny (a nie statyczny), a zmiany parametrów poprzednich układów (ruchów) nieuchronnie prowadzą do rozpadu nowo powstałego stanu równowagi i pojawienia się nowego, który nie jest jedynie nowym etapem w procesie rozwoju Świata, zachowując jednak stabilność poprzednich systemów i kontynuując ich rozwój pod względem jakości swoich elementów.
Pierwsze definicje Świata, wywiedzione bezpośrednio z samego faktu Początku, to przestrzeń i czas. Przestrzeń, jako pewne jeszcze nieokreślone, bezpośrednie istnienie Świata (Początek „czego”) i Czas, jako pewne przejście od Nieistnienia do Bycia itp. (czyli w ten sposób należy nadal budować i rozwijać OTGE).
Hipoteza o sekwencyjnym pojawianiu się głównych typów oddziaływań w czasie, a także założenie o zasadniczej nieredukowalności teorii naukowych opisujących poszczególne rodzaje ruchu, mogą stymulować postęp w kierunku stworzenia jednolitego fizycznego obrazu Świata. Głównym wyjściowym paradygmatem powinno być rozpatrywanie różnych interakcji fizycznych nie jako częściowych realizacji pewnej „pojedynczej interakcji”, ale jako łańcucha (sekwencji) samowystarczalnych systemów, zawierającego element tymczasowej modyfikacji i koniecznie generującej nowe systemów, gdy w poprzednim systemie pojawiają się sprzeczne decyzje.
Konsekwentnie, tworząc jednolitą teorię fizyczną, nie należy szukać jednego układu równań, którego częściowe rozwiązania opisywałyby poszczególne rodzaje oddziaływań, ale dążyć do stworzenia aparatu matematycznego, który koniecznie zawiera element rozwoju. Schemat może z grubsza wyglądać tak: pewien układ równań opisuje pewne procesy, w tym układzie występuje parametr, po zmianie układ traci swoją wyjątkowość (pojawiają się nieskończone lub inne sprzeczne rozwiązania). Aby zachować „równowagę” systemu, konieczne jest wprowadzenie nowego parametru, który kompensuje „sprzeczne decyzje”. Ale wynikiem jest inny układ równań, którego nie można sprowadzić do poprzedniego za pomocą zwykłych przekształceń matematycznych i opisuje nowy typ procesów. Nowy układ równań musi uwzględniać poprzedni jako element konieczny, a nie konkretne rozwiązanie.
Założenie o ewolucyjnym (sekwencyjnym) pojawianiu się rodzajów ruchu (oddziaływania fizyczne itp.) usuwa jeden z istotnych problemów kosmologii. Współczesna fizyka bez żadnych zastrzeżeń zakłada, że prawa opisujące różnego rodzaju oddziaływania mają charakter stacjonarny i istnieją niezależnie od ewolucji Świata. Oznacza to, że przyjmuje się, że chociaż w pierwszych chwilach po powstaniu Wszechświata, ze względu na szczególne warunki fizyczne (duża gęstość energii itp.), istnienie cząstek elementarnych, atomów itp. było niemożliwe, same prawa ( interakcje) już istniały. Pojawia się kilka pytań. Po pierwsze, czy legalne jest przypisywanie określonych właściwości, parametrów, praw interakcji itp. przedmioty, które nie tylko nie mogą jeszcze istnieć, ale w zasadzie nigdy nie istniały? Po drugie, gdzie przechowywana była informacja o tych prawach w chwili T = 0? Pytanie trzecie brzmi – jeśli uznamy, że pewne prawa (a w konsekwencji rodzaje ruchu) Świata są wieczne i niezmienne, to gdzie przebiega granica między nimi a rodzajami ruchu, które wyraźnie powstały dopiero na pewnym etapie rozwoju świata? rozwój Wszechświata (biologiczny, społeczny)? Przyjmując podstawowe zapisy OTGE, możemy udzielić jednoznacznych odpowiedzi na te pytania – w chwili T = 0 oddziaływania fizyczne nie istniały, to znaczy nie tylko nie działały z powodu braku obiektów, ale także kategorycznie nie dało się sformułować. A potem... Następnie przychodzi zadanie opracowania zarówno systemu filozoficznego, który logicznie wytwarza ciąg kategorii i definicji Świata, jak i stworzenia nowego dialektyczno-ewolucyjnego aparatu fizycznego i matematycznego, zdolnego do prawidłowego opisu wzorców przejść pomiędzy rodzaje interakcji. (Jako zastosowanie do tych myśli możemy zaproponować zapoznanie się z pracą A. Boldacheva „Kosmologia akademika V.A. Ambartsumyana i hipoteza zmienności stałych światowych”).
Filogeneza
W współczesna biologia Znaczenie idei ewolucyjnej jest ogromne, jak w żadnym innym dziale nauk przyrodniczych. Dzieje się tak dlatego, że materiał dotyczący różnorodności organizmów żywych dostarcza najwięcej do myślenia o przemianie jednych rzeczy w inne. I nie jest przypadkiem, że kształtowanie się współczesnego globalnego ewolucjonizmu rozpoczęło się właśnie od teorii ewolucji Darwina, która wyjaśnia pochodzenie gatunków biologicznych.
Ponieważ różnorodność biologiczna jest wynikiem długiego rozwoju historycznego, nie da się zrozumieć budowy i funkcjonowania istot żywych bez znajomości ich historii. Okoliczność ta sprawia, że rekonstrukcja historyczna jest jednym z priorytetowych zadań współczesnej biologii.
Specjalna dyscyplina zajmuje się znajomością ścieżek i wzorców historycznego rozwoju organizmów - filogenetyka.
Filogenetyka powstała w latach 60. XIX wieku. wkrótce po opublikowaniu w 1859 roku książki Karola Darwina „O powstawaniu gatunków wg naturalna selekcja" Termin „filogeneza” pojawił się po raz pierwszy w 1866 r. w podstawowym dziele niemieckiego biologa ewolucyjnego E. Haeckela „Ogólna morfologia zwierząt”. Za pomocą tej koncepcji autor określił zarówno proces historycznego rozwoju organizmów, jak i strukturę powiązanych (filogenetycznych) zależności między nimi. Termin „ewolucja”, wprowadzony do obiegu naukowego mniej więcej w tych samych latach przez angielskiego filozofa G. Spencera, w jego współczesnym historycznym rozumieniu (wcześniej oznaczał rozwój indywidualny), również szybko zyskał popularność.
W rezultacie pojęcia filogenezy i ewolucji zaczęto postrzegać jako bardzo bliskie znaczeniu. Ta klasyczna interpretacja istnieje do dziś: filogenezę definiuje się jako ścieżki, wzorce i przyczyny historycznego rozwoju organizmów. Wybitny rosyjski ewolucjonista II Szmalgauzen uważał filogenezę za łańcuch zastępujących się nawzajem ontogenii.
Obecnie kształtuje się biotocentryczna koncepcja filogenezy, zgodnie z którą ewolucja biologiczna to samorozwój fauny i flory jako integralnego układu, a filogeneza jest jednym z aspektów tego rozwoju. Takie rozumienie ewolucji biologicznej w ogóle, a filogenezy w szczególności jest najbardziej spójne ze współczesnymi koncepcjami ogólnych praw rozwoju rozwijanych przez synergetykę.
Jednym z najważniejszych rezultatów ewolucji jest globalna struktura fauny i flory Ziemi, która przejawia się w wielopoziomowej hierarchii grup, odmiennie zintegrowanych i zorganizowanych. W pewnym przybliżeniu można uznać, że struktura ta składa się z dwóch hierarchii. Jeden z nich związany jest z różnorodnością biocenoz (naturalnych ekosystemów), połączonych relacjami ekologicznymi. Historyczne ukształtowanie się tej hierarchii określa się mianem filocenogeneza. Drugie związane jest z różnorodnością taksonów – grup filogenetycznych, których członkowie są ze sobą spokrewnieni. Tworzenie się właśnie tej hierarchii jest filogeneza.
Filogeneza, czyli filogeneza (starożytny grecki phylon - plemię, rasa i inne greckie genetikos - związane z narodzinami) to historyczny rozwój organizmów. W biologii filogeneza bada rozwój gatunku biologicznego w czasie. Klasyfikacja biologiczna opiera się na filogenezie, ale może różnić się metodologicznie od filogenetycznej reprezentacji organizmów.
Filogeneza uważa ewolucję za proces, w którym linia genetyczna – organizmy od przodka do potomka – rozgałęzia się w czasie, a jej poszczególne gałęzie mogą ulegać pewnym zmianom lub zanikać w wyniku wyginięcia.
Obecną wiedzę na temat rozgałęzień drzewa filogenetycznego uzyskano poprzez skonstruowanie klasyfikacji organizmów żywych, która została pierwotnie pomyślana przez Carla Linneusza jako odzwierciedlenie „Systemu Naturalnego” wszelkiej przyrody (w tym nieożywionej). Następnie ustalono, że taki „system naturalny” nie istnieje, a to, co K. Linneusz uznał za przejaw tego systemu u zwierząt i roślin, to filogeneza, czyli wynik ewolucji biologicznej. Dla efektywniejszej analizy filogenezy opracowywane są obecnie zasady, w których udoskonala się sposób zapisu klasyfikacji w porównaniu do metody Linneusza, co pozwala na dokładniejsze rejestrowanie filogenezy w formie klasyfikacji i kontynuację jej analizy.
Ryż. Filogeneza głównych grup organizmów
W filogenezie wyróżnia się kilka elementów, które umożliwiają badanie rozwoju historycznego pod różnymi kątami. Na początku XX wieku. austriacki paleontolog O. Abel wyróżnił je w następujący sposób: seria przodków - „prawdziwe filogenezy”; rzędy urządzeń odnoszących się do jednego narządu; szereg kroków mających na celu poprawę organizacji. Obecnie „prawdziwa filogeneza” – rozwój (pojawienie się i/lub zmiana składu) grup filogenetycznych organizmów jako takich, niezależnie od ich właściwości – określana jest jako kladogeneza, czyli historia kladystyczna. Termin ten zaproponował angielski biolog J. Huxley w latach 40. XX wieku, przeciwstawiając go anagenezie – zmianie poziomu organizacji istot żywych w procesie ewolucji. W latach 60. niemiecki taksonomista i ewolucjonista W. Zimmerman zaproponował nazwanie zmian historycznych zachodzących w poszczególnych narządach i właściwościach organizmów semofilogenezą. Bardziej odpowiednim terminem jest semogeneza, która oznacza historyczny rozwój (pojawienie się, zmianę, zanik) poszczególnych struktur morfologicznych i innych, rozpatrywanych bez związku z określonymi grupami organizmów, do których są one nieodłączne.
Grupy powstałe w wyniku kladogenezy nazywane są kladami: na przykład strunami, a w ich obrębie - kręgowcami. Grupy powstałe w wyniku anagenezy to stopnie, etapy rozwoju ewolucyjnego: są to zwierzęta wielokomórkowe w stosunku do zwierząt jednokomórkowych, a wśród kręgowców - zwierzęta homeotermiczne (ptaki i ssaki) w stosunku do zwierząt poikilotermicznych (niższe kręgowce). Podstawową różnicą między tymi dwiema kategoriami jest sposób ich nabywania właściwości ogólne. Klady dziedziczą je od wspólnego przodka, a w miastach pojawiają się w wyniku równoległej lub zbieżnej ewolucji. Współczesna filogenetyka bada przede wszystkim powstawanie hierarchii grup filogenetycznych i ich specyficzne właściwości, a jej głównym zadaniem jest rekonstrukcja kladogenezy. Z tego jasno wynika, że na obecnym etapie rozwoju filogenetyka to przede wszystkim kladogenetyka.
Od systemu naturalnego do idei ewolucyjnej. W biologii klasycznej w XVII-XIX wieku. jedną z kluczowych była idea systemu naturalnego jako uniwersalnego prawa, któremu podlega wszystko, co istnieje. System ten jednoczy naturalne grupy organizmów, które istnieją w samej naturze. Systematyka była i jest zaangażowana w jej rekonstrukcję, której zadaniem jest rozpoznanie tych grup po ich „naturze”.
Ponieważ ostatecznym celem wiedzy naukowej jest poznanie przyczyn tego czy innego zjawiska, sam system naturalny potrzebował wyjaśnienia: jaka była jego pierwotna przyczyna. W średniowieczu uważano to za boski plan stworzenia. Wraz z pojawieniem się współczesnej nauki zaczęto szukać przyczyny w samej Naturze: w ten sposób idea rozwoju historycznego weszła do światopoglądu nauk przyrodniczych, który dał najbardziej wiarygodną interpretację systemu naturalnego z materialistycznego punktu widzenia .
Niemiecka filozofia przyrody odegrała ważną rolę w tworzeniu systematyki i filogenetyki. Wiąże się z tym z jednej strony idea „Wielkich Schodów Natury” (G. Leibniz), sięgająca czasów Arystotelesa, a z drugiej strony przyrównanie Natury do superorganizmu (F. Schelling, L. Oken). Podali dwa sposoby reprezentacji układu naturalnego – liniowy i hierarchiczny. W pierwszym przypadku chodziło o ciągły ciąg organizmów („natura nie robi skoków”), skierowany od niższych do wyższych form życia. W drugim przypadku sugerowano, że fauna i flora, podobnie jak organizm, jest podzielona na części o różnych poziomach zbiorowości - ugrupowania naturalne.
Przyrównanie fauny i flory do superorganizmu miało szczególne znaczenie dla ukształtowania się ewolucyjnego światopoglądu: wszak nie do pomyślenia jest żywa istota bez rozwoju ukierunkowanego na większą doskonałość i różnicowanie. Na tej podstawie, w połączeniu z ideą „drabiny doskonalenia” (inna nazwa „drabiny natury”), ukształtowała się kluczowa idea klasycznego ewolucjonizmu, a wraz z nią klasycznej filogenetyki – utożsamienie historyczny rozwój fauny i flory po indywidualny rozwój organizmu.
Ewolucyjna interpretacja ciągłego ciągu form dała początek nauczaniu J.-B. Lamarcka, zgodnie z którym głównym zadaniem teorii ewolucji było wyjaśnienie, w jaki sposób powstają nowe właściwości organizmów, które zapewniają wzrost poziomu ich organizacji. W ten sposób w filogenetyce narodził się paradygmat adaptacjonistyczny.
System hierarchiczny najściślej odpowiada rozbieżnej koncepcji ewolucji, kojarzonej z imieniem Karola Darwina. W tym przypadku historia sprowadza się właściwie do wielokrotnie rozgałęziających się „łańcuchów” transformacji jednego gatunku w drugi. Ta wizja rozwoju ewolucyjnego utworzyła paradygmat genealogiczny w filogenetyce.
Biolodzy o nastawieniu przyrodniczo-filozoficznym postrzegali rozwój historyczny jako proces w skali makro. Dla nich najważniejszym zadaniem była (i pozostaje do dziś) rekonstrukcja formowania się głównych grup organizmów żywych – królestw, typów, etapów rozwoju. Dla zwolenników coraz bardziej popularnej darwinowskiej interpretacji ewolucji kluczowymi wydarzeniami były te, które miały miejsce na poziomie gatunku. W związku z tym w rekonstrukcji historii główny nacisk kładziony jest na wydarzenia „lokalne” - przemianę jednego gatunku w drugi.
Na początku XX wieku. cały naturalny filozoficzny obraz świata uznano za „nienaukowy”, gdyż jego twierdzenia nie zostały zweryfikowane empirycznie. Darwinowski selekcjonizm, uzupełniony teorią mutacji, doskonale wpisuje się w nowy światopogląd. Stopniowo wyłoniła się syntetyczna teoria ewolucji, której podstawą ideologiczną było myślenie populacyjne. Terminologicznie ustalił podział rozwoju historycznego na mikroewolucję i makroewolucję (autorem terminów jest Yu.A. Filipchenko); jednocześnie absolutyzowano pierwszą, a drugą faktycznie odrzucono.
Sprowadzenie mechanizmów ewolucji do procesów populacyjnych pozwoliło uznać filogenezę za ich „mechaniczną” konsekwencję, wynikającą po prostu z faktu, że życie istnieje i rozwija się przez bardzo długi czas. To rozwój idei ewolucyjnej w pierwszej połowie - połowie XX wieku. uczyniła filogenetykę w pewnym stopniu „rzeczą drugiej kategorii” i pozbawiła ją dawnego statusu priorytetowego. Jednak w drugiej połowie tego samego stulecia znaczącą rolę w kształtowaniu odegrały idee populacyjne nowoczesna scena rozwój tej dyscypliny.
Filogenetyka klasyczna. Rozumienie filogenezy jako naturalnego procesu historycznego nie przeszkadza nam w odmiennej interpretacji znaczenia jedności filogenetycznej, która wyróżnia grupy naturalne. Dało to początek kilku kierunkom rozwoju filogenetyki klasycznej, które odpowiadają trzem wspomnianym już kluczowym składnikom filogenezy – kladogenezie, semogenezie i częściowo anagenezie.
Uznanie jedności filogenetycznej przede wszystkim jako wspólnoty pochodzenia, tj. monofilia, pobudziła rozwój filogenetyki w kierunku kladogenetycznym. Metafora rozgałęzionego drzewa filogenetycznego z pojedynczym korzeniem stała się dla kladogenetyki rodzajem symbolu.
W interpretacji Haeckela ta koncepcja filogenezy łączy w sobie filozofię przyrody i darwinowskie rozumienie ewolucji. Rozwój historyczny rozpatrywany jest na „poziomie makro”, ale gatunek uważa się za przodka dowolnej grupy monofiletycznej. Jako „dziedzictwo” filozofii przyrody, idea postępu ewolucyjnego obecna jest także w filogenetyce Haeckela: wyraźnie widać to po kształcie drzewa filogenetycznego, w którym dominuje skierowany ku górze pień centralny, z którego odchodzą gałęzie boczne .
Badając semogenetyczny komponent filogenezy, głównym zadaniem jest wyjaśnienie wzorców i przyczyn transformacji poszczególnych struktur. W tym przypadku o jedności filogenetycznej organizmów decyduje głównie wspólność trendów ewolucyjnych, która wyraża się w jedności kierunków i etapów rozwoju historycznego. Na takie rozumienie filogenezy duży wpływ miała koncepcja ewolucji jako procesu jednokierunkowego, rozwinięta przez niektórych paleontologów (E. Kop, V.O. Kovalevsky, G. Osborne). Bardziej spójne jest to z metaforą nie drzewa, ale krzaka, a nawet pola trawy, na którym każdy „łodyga” – linia filetyczna – wyrasta z własnego korzenia (L.S. Berg).
Metodologia filogenetyki klasycznej opiera się na metodzie potrójnego równoległości, tj. w sprawie maksymalnej harmonizacji danych z anatomii porównawczej, embriologii i paleontologii. Kluczowe znaczenie ma koncepcja homologii, pierwotnie opracowana przez angielskiego anatoma platońskiego R. Owena: rozumie się, że dla rekonstrukcji filogenezy ważne są tylko struktury homologiczne, podczas gdy struktury analogiczne w tym przeszkadzają. Wiele uwagi poświęca się określeniu prawdopodobnych ścieżek ewolucji tych struktur w oparciu o kryteria opracowane w ramach modeli perfekcjonistycznych (od prostych do złożonych, od mniej do bardziej ustrukturyzowanych) i adaptacjonistycznych (od organizmów mniej do bardziej przystosowanych). Częścią klasycznej metodologii jest analiza form przodków: za ich pomocą odmienne grupy są „łączone” w formacje monofiletyczne. Szczególną uwagę zwraca się na materiały kopalne, wśród których poszukuje się prawdziwych przodków; w przypadku ich braku rekonstruowane są postacie przodków.
Ważną rolę odgrywa ważenie cech. W filogenetyce klasycznej jest to zawsze interpretowane jako różniczkowe: podawane są różne znaki inne znaczenie(inna „waga”). Jednak w różnych szkołach filogenetyki to ogólne pojęcie jest interpretowane w specyficzny sposób, w zależności od rozumienia istoty filogenezy.
Interpretacja genealogiczna polega na podziale cech na „filogenetyczne”, niezwiązane z adaptacjami prywatnymi, oraz „adaptacyjne”. Rozumie się, że to pierwsze należy wykorzystać do ustalenia pokrewnych relacji między gatunkami, ponieważ są one dziedziczone od przodków (bliskich lub dalszych), co wskazuje na wspólnotę filogenetyczną ich potomków.
Interpretacja adaptacjonistyczna natomiast przypisuje największą wagę tym cechom, które odzwierciedlają adaptacyjną specyfikę danej grupy. W istocie koncepcja ta jest echem poprzedniego rozumienia „głównej istoty” taksonów naturalnych. W tym przypadku znaczenie struktur morfologicznych wiąże się z ich znaczeniem adaptacyjnym (najczęściej funkcjonalnym).
Obecnie tradycja filogenetyki klasycznej rozwija się w kierunku podejścia zwanego filistyką. W nim rozwój filogenetyczny jest nadal uważany za uporządkowaną adaptogenezę, w której regularne trendy przeważają nad przypadkowymi. Dlatego też, charakteryzując filogenetyczną jedność grup, dużą wagę przywiązuje się nie tylko do jedności pochodzenia, ale także do jedności tendencji ewolucyjnych ich przedstawicieli jako dodatkowego dowodu monofilii.
Nowa filogenetyka. Większość współczesnych rekonstrukcji przeprowadzana jest w ramach nowej filogenetyki. Powstała w wyniku połączenia trzech podejść, które początkowo (w latach 60.) rozwijały się niezależnie - kladystyki, filetyki numerycznej i genofiletyki. Można ją zdefiniować jako gałąź filogenetyki, która rozwija hipotezy filogenetyczne w oparciu o metodologię kladystyczną z wykorzystaniem filetyki numerycznej z wykorzystaniem molekularnych danych genetycznych.
Ideologicznym rdzeniem nowej filogenetyki jest kladystyka, której powstanie wiąże się z nazwiskiem niemieckiego entomologa W. Genniga. Jego przełomowa książka ukazała się w języku niemieckim w 1950 r., jednak kierunek ten zyskał szczególną popularność po opublikowaniu wydania anglojęzycznego (Hennig W. Systematyka filogenetyczna. Urbana, 1966.). W tym samym czasie pojawił się sam termin „kladystyka”.
Podstawą metodologiczną podejścia kladystycznego jest zasada ekonomii, która w miarę możliwości wyklucza wstępne spekulacje na temat badanego zjawiska. W związku z tym rekonstrukcje historyczne przeprowadza się przy minimalnych założeniach apriorycznych dotyczących właściwości procesu ewolucyjnego. Ogólna koncepcja ewolucji jako adaptogenezy, która zobowiązuje do szczegółowego opisu scenariuszy ewolucji, ważenia cech według ich znaczenia adaptacyjnego itp., okazuje się zbędna. Uzasadnia to redukcję rozwoju historycznego do kladogenezy, co jest kluczowe dla nowej filogenetyki, w której nie ma równoległych linii ani etapów rozwoju ewolucyjnego.

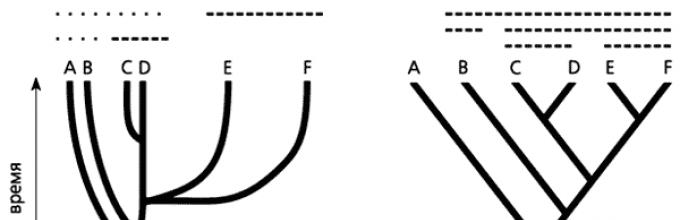
Ryż. Stylizowane reprezentacje schematów filogenetycznych we współczesnej filogenetyce: a – filogram, b – kladogram. Linie ciągłe pokazują grupy parafiletyczne, linie przerywane pokazują grupy holofiletyczne.
To uproszczone zrozumienie filogenezy znacząco zmieniło jej graficzną reprezentację. Kladogram – stylizowane drzewo filogenetyczne – stał się swoistym symbolem kladystyki, a wraz z nią całej nowej filogenetyki (ryc.). Na nim, w przeciwieństwie do filogramu pokazującego stopień rozbieżności i poziom zaawansowania grup, znajdują się jedynie punkty rozgałęzień odzwierciedlające kolejność powstawania grup monofiletycznych. Nie ma anagenetycznego składnika rozwoju ewolucyjnego; dlatego patrząc na kladogram nie można ocenić poziomu rozbieżności i zaawansowania grup organizmów.
Jedną z przyczyn sukcesu kladystyki jest skuteczna metodologia rekonstrukcji filogenetycznych, która zasadniczo umożliwiła ich powtarzalność. Jej ważną częścią jest bardziej „wąska” interpretacja pojęć monofilii i grupy monofiletycznej (por. ryc. 4). W klasycznej filogenetyce grupę uważa się za monofiletyczną, jeśli obejmuje jakichkolwiek potomków danej formy przodków. Oczywiście przy identyfikacji grup monofiletycznych na takim drzewie filogenetycznym możliwe są bardzo duże rozbieżności. Kladystyka rozpoznaje jedynie grupy holofiletyczne, do których zaliczają się wszyscy potomkowie danej formy przodków. Grupy, w których nie uwzględnia się tylko części takich potomków (parafiletycznych).

Ryc. Kategorie podobieństwa związane z pojęciem synapomorfii. Wielkie litery oznaczają apomorfie, małe litery oznaczają plezjomorfie; Apomorfie definiujące grupy holofiletyczne są wyróżnione kolorem.
Częścią tej metodologii była paradoksalna z punktu widzenia filogenetyki klasycznej odmowa uwzględnienia relacji przodek-potomek przy omawianiu pokrewieństwa i monofilii. Podstawą tego było założenie, że dla zdecydowanej większości grup, zwłaszcza tych wysokiej rangi, nie da się ustalić konkretnego gatunku przodków. W tym przypadku bardziej poprawne jest odniesienie się do grupy siostrzanej, która powstała w wyniku jednego zdarzenia kladystycznego razem z badaną. Pozwala to zignorować geochronologiczny składnik ewolucji i „zrównać” dane paleontologiczne ze współczesnymi. Stanowisko to stanowi teoretyczną podstawę dla rekonstrukcji genofiletycznych, w przypadku których początkowo nie trzeba badać form przodków.
Podstawą większości algorytmów nowej filogenetyki jest zasada synapomorfii, zgodnie z którą monofilię grupy można ujawnić jedynie poprzez podobieństwo stanu pochodnego cechy (apomorfia, ryc.). Natomiast symplezjomorfia – podobieństwo w stanie początkowym charakteru (plezjomorfia) – nie pozwala na określenie monofilii grupy. W tym przypadku wyróżnia się podobieństwo synapomorficzne odziedziczone od najbliższego przodka tej grupy (prawdziwa synapomorfia). Wręcz przeciwnie, podobieństwa wynikające z ewolucji równoległej (fałszywa synapomorfia) uważa się za nieistotne: to zasadniczo odróżnia kladystykę od klasycznej filogenetyki.
To ostatnie stanowi jedno z najważniejszych kryteriów – znaczenie cech: ich „waga” jest odwrotnie proporcjonalna do prawdopodobieństwa wystąpienia równoległości i odwrotów w ewolucji. Im wyższe to prawdopodobieństwo, tym mniej istotne są cechy, ponieważ pozwalają nam oceniać monofilię z mniejszą wiarygodnością. Zamiast selekcji cech najbardziej znaczących wprowadza się kryterium ilościowe całościowej oceny istotności podobieństwa: jakie duża liczba dwa taksony mają wspólne apomorfie, tym większe jest prawdopodobieństwo ich przynależności do grupy monofiletycznej. Aby zatem dowiedzieć się, czy określone grupy taksonów są monofiletyczne, wystarczy zsumować synapomorfie specyficzne dla każdej z nich i porównać wyniki. Umożliwia to po prostu rozwiązywanie konfliktów pojawiających się między różnymi postaciami, co może prowadzić do różnych schematów filogenetycznych.
Kryterium to stymulowało aktywny rozwój współczesnej filogenetyki metod ilościowych rozwijanych przez filetykę numeryczną. Obecnie istnieje swego rodzaju „przemysł” wytwarzania metod ilościowej oceny podobieństwa organizmów oraz konstrukcji drzew filogenetycznych i ich wdrażania w postaci programy komputerowe.
Obecnie dwie najpopularniejsze metody to oszczędność i maksymalne prawdopodobieństwo. Zasadnicze różnice między nimi polegają na różnej interpretacji warunków początkowych rekonstrukcji. Metody pierwszej grupy, wyodrębnione ze względu na omówioną powyżej zasadę ekonomii (stąd ich nazwa), polegają na całkowitym odrzuceniu apriorycznego określania ewolucji cech interpretowanych fenetycznie. W drugiej grupie wprowadza się postacie jako elementarne hipotezy dotyczące semogenezy, w których określa się prawdopodobne kierunki przemian ewolucyjnych odpowiednich struktur. W pierwszym przypadku ostateczna hipoteza jest optymalna, jeśli ma minimalną liczbę równoległości; w drugim przypadku, jeśli w najbardziej przekonujący sposób podsumowuje wszystkie ewolucyjne scenariusze postaci.
Niewątpliwą zaletą filetyki numerycznej jest to, że pozwala ona na manipulację dużą ilością danych. Morfolog najczęściej jest w stanie intuicyjnie uchwycić jedynie ograniczoną liczbę struktur, które można opisać za pomocą kilkudziesięciu sformalizowanych cech. Jednak bez zastosowania technik ilościowych porównanie kilkudziesięciu taksonów na podstawie sekwencji nukleotydowych składających się z kilku tysięcy par zasad jest prawie niemożliwe. Ale i tutaj są pułapki. Zatem sumowanie kilkudziesięciu cech morfologicznych i kilkuset cech molekularnych prowadzi do tego, że te pierwsze są „wchłaniane” przez te drugie. W rezultacie schemat filogenetyczny jest zdeterminowany prawie całkowicie przez molekularne dane genetyczne.
Aktywna „numeracja” nowej filogenetyki w coraz większym stopniu przenosi nacisk z teorii i metodologii rekonstrukcji na ich „technologię”. Omówiono szczegóły obliczania przedziałów ufności przy zastosowaniu kryteriów probabilistycznych, szybkość programów komputerowych, ich ograniczenia w manipulowaniu dużymi ilościami danych itp. Jednocześnie coraz mniej uwagi poświęca się biologicznemu rozumieniu wyników, co w dużej mierze zaciemnia ewolucyjny sens takich rekonstrukcji.
Specjalnym działem filetyki numerycznej jest komputerowe modelowanie procesów filogenetycznych. Pozwala na badanie niektórych właściwości historycznego rozwoju organizmów w oparciu o modele symulacyjne.
Genofiletyka (filogenomika) to dział nowej filogenetyki, w którym wnioski dotyczące historii kladystycznej wyciąga się na podstawie analizy wyłącznie molekularnych danych genetycznych. Ten najnowszy etap rozwój filogenetyki molekularnej, związanej z badaniem sekwencji nukleotydowych. Teoretyczne przesłanki wykorzystania takich danych w filogenetyce zostały sformułowane w ramach syntetycznej teorii ewolucji (STE), która sprowadzała ewolucję do zmian zachodzących na poziomie genetycznym. Na tej podstawie wysunięto koncepcję: im „bliżej” genotypu znajdują się postacie, tym większe jest ich znaczenie dla rekonstrukcji filogenetycznych. Nie trzeba dodawać, że uzyskanie technicznego dostępu do pierwotnej struktury informacyjnych makrocząsteczek uznano za „rewolucję molekularną” w filogenetyce.
W genofiletyce przy rekonstrukcji relacji rodzinnych szczególne znaczenie (w nawiązaniu do Darwina) przywiązuje się do zmian w strukturze pierwszorzędowej makrocząsteczek, które nie są związane z rozwojem poszczególnych adaptacji. Taka interpretacja ewolucji jest w pełni zgodna z zasadami kladystyki, która w naturalny sposób łączy tę drugą i genofiletykę w ramach nowej filogenetyki. Tę okoliczność należy szczególnie podkreślić: gdyby we współczesnej filogenetyce dominowała koncepcja adaptacjonistyczna, nie byłoby w niej miejsca na rekonstrukcje genofiletyczne, gdyż nie są one interpretowalne z punktu widzenia genezy adaptacji.
W genofiletyce organizm sprowadza się do pierwotnej struktury biopolimerów, których każdy element (nukleotyd) jest uważany za odrębną postać, a może być ich kilka tysięcy. Sądy dotyczące pokrewieństwa opierają się na sumarycznej ocenie podobieństwa na podstawie tych elementarnych cech, a im więcej ich (im dłuższa sekwencja), tym bardziej wiarygodne są te sądy. Dlatego jedyną akceptowalną metodą oceny podobieństwa jest metoda ilościowa.
Aby przejść od podobieństwa struktur molekularnych do pokrewieństwa, opracowano koncepcję zegara molekularnego. Opiera się ona na tzw. modelu „neutralnej ewolucji”, który zakłada stochastyczny charakter akumulacji neutralnych mutacji w liniach filetycznych. Kluczem do koncepcji było założenie, że postęp tych „zegarów” jest w przybliżeniu jednolity i taki sam dla różnych struktur molekularnych i różnych grup organizmów. W konsekwencji, jeśli znane jest tempo akumulacji mutacji, można określić czas separacji grup i na tej podstawie ustalić pokrewieństwo genetyczne między nimi.
Ilościowe określenie różnic w strukturach molekularnych (odległość genetyczna) pozwala uzyskać wewnętrzne czasy rozbieżności, których nie można bezpośrednio porównywać dla różnych grup organizmów. Jego przeliczenie na jeden dla nich czas absolutny (w milionach lat) odbywa się poprzez kalibrację „zegara molekularnego”. W tym celu obliczone odległości porównuje się z czasem pojawienia się w zapisie kopalnym pierwszych przedstawicieli grup monofiletycznych zidentyfikowanych metodami molekularnymi. Następnie odległości są przeliczane w jednostkach szybkości zmian struktura pierwotna makrocząsteczek i wyznacza się bezwzględny czas dywergencji.
Pokazuje to, że problem kalibracji „zegara molekularnego” nie jest rozwiązany w ramach samej biologii molekularnej: wynik zależy od danych paleontologicznych. Bardzo ważna jest tutaj dokładność określenia czasu pochówku szczątków kopalnych oraz poprawność ich przypisania do grup identyfikowanych za pomocą markerów molekularnych. Ponadto różne metody szacowania odległości genetycznych, a także kalibrowania samego „zegara”, mogą dawać różne wyniki. Wszystko to stanowi poważną podstawę do krytyki pierwotnej, uproszczonej interpretacji koncepcji „zegara molekularnego”. Zgromadzone dane pokazują, że tempo zmian w strukturze pierwszorzędowej jest różne dla różnych makrocząsteczek, a dla tych samych makrocząsteczek – dla różnych ich przekrojów i w różnych grupach organizmów. Oznacza to, że uniwersalny „zegar molekularny” nie istnieje ze względu na nierównomierność jego biegu.
Możliwości genofiletyki znacznie ogranicza fakt, że molekularne dane genetyczne nadające się do badań filogenetycznych zgromadzono dotychczas jedynie od kilkudziesięciu tysięcy do prawie 2 milionów. gatunki współczesne, znane nauce. Istnieje jednak także wiele milionów wymarłych gatunków, w przypadku których tego rodzaju dane nigdy nie będą dostępne. Wszystko to czyni go wyjątkowym faktyczny problem niekompletność próby: im mniej należących do niej taksonów jest reprezentowanych w badanej grupie, tym mniej trafna jest ocena powiązań filogenetycznych między nimi.
Zastosowania nowej filogenetyki. Wiele dyscyplin biologicznych (systematyka, biogeografia itp.) ostatnie lata także ekologia i etologia) w takiej czy innej formie korzystają z wyników badań filogenetycznych. I w tym sensie można je uznać za obszary zastosowania filogenetyki jako dyscypliny naukowej.
Obecnie istnieje cała klasa problemów, których rozwiązanie w taki czy inny sposób wiąże się z analizą sygnału filogenetycznego (dokładniej kladogenetycznego) - informacji o pochodzeniu organizmów zawartej w strukturze ich podobnych powiązań. Ocena tego sygnału jest równoznaczna ze wskazaniem m.in. przyczyn historycznych, które determinowały obserwowane zróżnicowanie. Oczywiście, aby zidentyfikować ten sygnał, konieczne jest w miarę pełne zrekonstruowanie historii kladystycznej organizmów.
Rzeczywiście, każdy rozwijający się system, mówiąc w przenośni, jest „ofiarą swojej historii”. Oznacza to, że o strukturze lub stylu życia organizmów można decydować nie tylko na podstawie cech ich siedliska, ale także cech ich prehistorii. Na przykład, dlaczego różne zwierzęta lądowe mają różną liczbę kończyn – cztery u kręgowców, sześć u owadów, osiem u pajęczaków? Nie ma sensu szukać odpowiedzi w ich warunkach życia. Leżą raczej w morfologicznej organizacji form przodków każdej z tych grup, od których potomkowie odziedziczyli swoje cechy. Zrozumienie tego doprowadziło do powstania programu badawczego - filogenetyki porównawczej. Bada przejawy przyczyn historycznych w charakterystyce stylu życia, morfologii, zachowaniu itp. w różnych grupach organizmów.
Niemal od pierwszych etapów swojego rozwoju filogenetyka jest ściśle związana z biogeografią historyczną, dostarczając jej informacji o historii powstawania taksonów tworzących tę czy inną lokalną faunę i florę. Obecnie powiązanie to ukształtowało się w postaci biogeografii kladystycznej, która w całości opiera się na analizie kladystycznego składnika filogenezy.
Jeśli początkowo filogenetyka faktycznie „wyrosła” z systematyki biologicznej, dziś wręcz przeciwnie, nowa filogenetyka ma znaczący wpływ na systematykę, jej zasady i metody. Filogenetyka klasyczna definiuje zadania systematyki jako odzwierciedlenie wyników filogenezy w systemie taksonów charakteryzującym się jednością filogenetyczną. Wyjaśnienie tej jedności dało początek tzw. systematyce kladystycznej, której celem jest „przełożenie” kladogramu na system taksonomiczny.
Paradoks sytuacji polega na tym, że ta różnica, bardzo nieznaczna w formie, bardzo silnie wpłynęła na państwo współczesna taksonomia. Wiele taksonów, uznanych przez pokolenia taksonomistów, okazało się parafiletycznych i dlatego podlegało „zniesieniu”. Spowodowało to falę znaczącej restrukturyzacji naturalnego systemu organizmów żywych i w związku z tym ostre kontrowersje dotyczące ich kompetencji i ważności. Typowym przykładem takich nieporozumień jest sytuacja, która rozwinęła się w systemie współczesnych przedstawicieli gadów i ptaków (ryc.). W klasycznej taksonomii za grupy naturalne uznawane są Linneuszowskie klasy Reptilia (żółwie, hatterie, lepidozaury i krokodyle) oraz Aves (ptaki). Powód jest dość oczywisty: ewolucja ptaków wyprowadziła je poza granice gadziej normy adaptacyjnej, przez co utraciły z nimi jedność.

Ryż. Klasyczna (po lewej) i kladystyczna (po prawej) klasyfikacja gadów i ptaków.
Ale w schemacie genealogicznym wśród wymienionych grup najbliższymi krewnymi krokodyli są ptaki. Z punktu widzenia kladystyki klasyczny takson Reptilia jest parafiletyczny: w jego składzie powinny pozostać jedynie hatteria i lepidozaury (jaszczurki i węże). Żółwie często zalicza się do klasy Parareptilia, natomiast ptaki i krokodyle należy klasyfikować do klasy Archosauria (która wśród form kopalnych obejmuje także dinozaury i pterozaury).
Sekcja genofiletyczna nowej filogenetyki rozwija tzw. systematykę genów, w której taksony tworzone są wyłącznie na podstawie danych genetycznych. Jego zasługa polega na wyjątkowym merytorycznym wsparciu starej idei „Drzewa Życia” - globalnej klasyfikacji świata organizmów żywych. W szczególności bardzo szczegółowo opracowywany jest system taksonomiczny prokariotów, prowadzący do identyfikacji nowych królestw organizmów żywych.
Systematyka kladystyczna nie ignorowała nomenklatury taksonomicznej. Obecnie opracowywany jest tzw. „Filokod”, którego ideologiczną podstawą jest zasada monofilii w jej „wąskiej” interpretacji (Cantino P.D., Queiroz K. The PhyloCode: a phylogenetic code of biologi nomenclature. 2003).
Rozwój nauki przypomina w pewnym sensie ewolucję biologiczną. Różnorodność pomysłów, podejść i metod wpisana jest także w dynamicznie rozwijającą się firmę dyscyplina naukowa, jak różnorodność gatunków - ewoluująca fauna i flora. Ciągła rywalizacja idei, powstawanie i śmierć szkoły naukowe, rozwój nowych kierunków – wszystko to jest zarówno konsekwencją, jak i przyczyną postępu naukowego.
Filogenetyka nie jest pod tym względem wyjątkiem. Jego główny wektor rozwoju przez cały czas wyznacza oczywisty „supercel” - coraz pełniejsza rekonstrukcja historii powstawania fauny i flory. Jak to osiągnąć, to główny problem tej dyscypliny. Metody jego rozwiązania dają podstawę do różnych podejść do rekonstrukcji filogenetycznych. Tworzenie nowej filogenetyki wiąże się z wyjaśnieniem tego wspólne zadanie i próbuje rozwiązać stare problemy w nowy sposób. Jednak każde podejście działa w granicach swoich możliwości, a jego nadmiernie optymistyczna interpretacja często prowadzi w złym kierunku. Błędem byłoby zatem zakładać, że zaproponowane dziś rekonstrukcje historyczne są ostateczne.
Jest oczywiste, że nowa filogenetyka zostanie zastąpiona innym podejściem. Można przypuszczać, że pod warunkiem tendencji rozwoju teorii ewolucji, uwolni się on od nadmiaru redukcjonizmu nieodłącznie związanego ze współczesnymi badaniami.
Przejdźmy teraz do rozważenia przyczyn i czynników napędzających ewolucję biologiczną.
(1) Czym jest filogeneza
Nazywa się proces nieodwracalnego historycznego rozwoju świata organizmów żywych, zarówno w ogóle, jak i poszczególnych grup taksonomicznych filogeneza (z greckiego filon- klan, plemię i geneza– pochodzenie, powstanie).
Rozwój, zarówno indywidualny, jak i historyczny, jest najważniejszą właściwością życia. Jednak rozwój i samoorganizacja są nieodłącznie związane z materią w ogóle. W szerokim znaczeniu proces rozwoju to ruch, zmiana w przestrzeni i czasie. Ruch jako sposób istnienia materii nie powstaje i nie znika, istnieje wiecznie – bez początku i końca. Jednocześnie ruch jest procesem wektorowym, ukierunkowanym, nieodwracalnym, co oznacza, że na odrębnych odcinkach ma początek i koniec: ... à ... à ... à ...
To znaczy, że Rozwój historyczny jako ruch jest nieskończony i jednocześnie istnieje za każdym razem jako odrębny, skończony proces. Te pozorne sprzeczności można rozwiązać za pomocą filozoficznego spojrzenia na proces rozwoju. Historyczny rozwój każdego systemu: fizycznego, biologicznego, społecznego – podlega ogólnym prawom dialektyki i synergetyki. Rozwój ma miejsce w spirali (to z dialektyki) i towarzyszy mu okresowość rozwidlenia (zgodnie z prawami synergetyki) (ryc. 6.5).
Tak naprawdę jeden obrót spirali to pewien proces skończony. W schemacie rozwoju życia na Ziemi taka rewolucja stanowi odrębność ontogeneza – indywidualny rozwój jednostki. Bifurkacje, czyli rozbieżności, to rozbieżność ścieżek rozwojowych, powstanie wielu nowych ontogeniów, tj. nowe gatunki. Zatem spirala z licznymi rozgałęzieniami reprezentuje niekończący się proces historyczny filogeneza – nieodwracalna ewolucja organizmów prowadząca do wzrostu i złożoności różnorodności biologicznej.
Ryż. 6.5. Zasada rozwoju spiralno-bifurkacyjnego
Każdy obrót spirali jest osobną ontogenezą; cała rozgałęziona spirala – filogeneza
Zatem, Transformacje filogenetyczne zachodzą poprzez restrukturyzację ontogenezy jednostek; Co więcej, zmiany na każdym etapie rozwoju jednostki mogą mieć wartość adaptacyjną. Można tak powiedzieć filogeneza to kolejny ciąg (a biorąc pod uwagę rozbieżności - drzewo) ontogenezy kolejnych pokoleń.
(2) Od Lamarcka i Darwina do syntetycznej teorii ewolucji
Jakie czynniki zapewniają rozwój filogenetyczny i w jaki sposób? Dlaczego zachodzą zmiany w osobniczej ontogenezie? Co dzieje się w punkcie rozwidlenia (dywergencji), w którym jeden gatunek biologiczny zamienia się w dwa lub więcej gatunków o nowych cechach? Na te pytania odpowiada współczesność syntetyczna teoria ewolucji . Jego powstanie ma swoją złożoną historię i trudno je uznać za zakończone.
Pierwszym prawdziwym ewolucjonistą był francuski biolog Jean Baptiste Lamarck (1744–1829). W jego słynna książka„Filozofia zoologii” (1809) zaproponował pierwszą holistyczną teorię ewolucji świata żywego, odrzucając metafizyczną ideę stałości i niezmienności gatunków. Nauczanie Lamarcka opiera się na dużej ilości porównawczego materiału anatomicznego i nowej wizji systemu - od organizmów prostych po złożone. Potwierdzał postępujący rozwój życia na Ziemi. Jednocześnie przeważająca większość teoretyków biologii uważa, że Lamarck mylił się w ocenie sił napędowych (czynników) ewolucji. Całkiem słusznie uznając wiodącą rolę w zmienianiu gatunków pod wpływem warunków zewnętrznych (klimat, gleba, pożywienie, światło, ciepło itp.), uważał, że same organizmy charakteryzuje „wewnętrzna chęć doskonalenia”. Lamarck w to wierzył ćwiczenie narządów jednostki prowadzi do ich specyficznej, celowej zmiany i ulepszenia, a te pożyteczne zmiany są automatycznie utrwalane w potomstwie i przekazywane w drodze dziedziczenia (dziedziczenie cech nabytych). Przesadny przykład w tej kwestii jest powszechnie znany: żyrafy w wyniku ciągłych „ćwiczeń” w zrywaniu liści z drzew, przez wiele pokoleń nabyły długą szyję.
Przyczyny i siły napędowe procesów filogenetycznych zostały ponownie przemyślane przez Darwina wkrótce po Lamarcku. Charles Darwin (1809–1882) – wielki angielski przyrodnik, twórca doktryny o powstawaniu gatunków poprzez dobór naturalny. W latach 1831–1836 młody Darwin jako przyrodnik odbył podróż dookoła świata na statku Beagle, podczas którego zebrał dużą ilość materiału biologicznego, geograficznego, geologicznego i paleontologicznego, który stanowił podstawę jego nauk. Ponadto Darwin dokładnie przestudiował historię hodowli i selekcji zwierząt domowych i roślin i doszedł do wniosku, że podobnie sztuczna selekcja, stosowany w praktyce rolniczej, „działa” na wolności naturalna selekcja, który odgrywa główną rolę twórczą w tworzeniu najbardziej przystosowanych gatunków.
Syntetyczna teoria ewolucji opracowana w połowie XX wieku na podstawie Klasyczna teoria ewolucji Darwina, genetyka i ekologia populacji. Obecnie stara się wnieść swój wkład immunologia. W istocie teoria syntetyczna rozwija na nowym poziomie wiedzy ideę doboru naturalnego, uzasadnioną przez Darwina 100 lat przed tą syntezą. Jakie podstawowe postanowienia wprowadzono do teorii syntetycznej z poprzednich źródeł?
(3) Darwinizm jest podstawą syntetycznej teorii ewolucji
darwinizm pozostał fundamentem nowa teoria. W klasyczna praca Podpisano książkę Darwina „O powstawaniu gatunków drogą doboru naturalnego…” główne czynniki lub siły napędowe procesu ewolucyjnego. Teoria Darwina zawiera kilka przepisów.
1. Osobniki wydają na świat więcej potomstwa, niż jest to konieczne do utrzymania liczebności, dlatego organizmy mają zdolność do nieograniczonego rozmnażania się; jednocześnie charakteryzują się niepewna odziedziczalność(bezkierunkowa, losowa – w przeciwieństwie do zdecydowanej, ukierunkowanej zmienności Lamarcka).
2. Ze względu na ograniczone zasoby naturalne (żywność, przestrzeń itp.) pomiędzy jednostkami istnieje walka o byt, podczas którego większość umiera i nie rodzi potomstwa. Zauważmy, że idea walki o byt „wisiała w powietrzu”, Darwin ją podchwycił i rozwinął za T. Malthusem, który wykorzystał „walkę o byt” w swojej socjologicznej teorii regulacji populacji.
3. Organizmy najlepiej przystosowane do danych warunków środowiskowych przeżywają i pozostawiają potomstwo, co się objawia naturalna selekcja.
4. W wyniku selekcji osobniki tego samego gatunku kumulują się inaczej cechy adaptacyjne(twórcza rola selekcji), co prowadzi do ich rozbieżność(łac. odchodzić- odbiegać, odbiegać) i powstawanie nowych gatunków.
Zatem zdaniem Darwina zmienność dziedziczna reprezentuje przesłankę ewolucji , walka o byt i dobór naturalny- jego proces i specjacja- wynik.
(4) Wkład genetyki w teorię ewolucji
Genetyka już w nowym XX wieku dał zrozumienie natura dziedziczności (chromosomy, DNA) i mechanizmy zmienności (mutacje i rekombinacja genów). Odkryte przez Mendla w 1865 r. i ponownie odkryte w 1900 r., prawa niezależne dziedziczenie indywidualnych cech i ich zachowanie w stanie utajonym pozwoliło wyjaśnić to, czego Darwin nie rozumiał i nie mógł zrozumieć, ponieważ natura dziedziczności nie była w jego czasach znana. Genetyka i cytologia wykazały, że każda cecha organizmu jest kodowana przez dwa geny alleliczne zlokalizowane na chromosomach homologicznych – od dwojga rodziców (diploidalny zestaw chromosomów). Pozwala to organizmowi, bez ryzyka natychmiastowej śmierci, „wypróbować” różne warianty zmienności mutacyjnej w jednym z alleli.
Dziś wiemy, że zmienność mutacyjna osobników powstaje w wyniku spontanicznych lub indukowanych mutacji w komórkach rozrodczych rodziców i niesie potencjalnie nowe cechy. Spontaniczne (spontaniczne) mutacje występują jako błędy w doborze nukleotydów podczas replikacji DNA. Indukowane lub indukowane mutacje są spowodowane ekspozycją na różne czynniki mutagenne. Naturalne mutageny to: promieniowanie jonizujące przestrzeń kosmiczna i głębina ziemi, światło ultrafioletowe, substancje chemiczne, w tym żywność, substancje (zwłaszcza wolne, aktywne rodniki tlenowe), wirusy itp.
Opracowując te konstrukcje genetyczne, rosyjski biolog S.S. Czetwerikow w latach 20. XX wieku wypracował ważne stanowisko ws puli genowej gatunków i populacji, przez który rozumie się ogół wszystkich dzikich i zmutowanych wariantów genów (u wszystkich osobników) występujących w danym gatunku lub populacji i uczestniczących w swobodnym krzyżowaniu. Podczas rozmnażania płciowego ukryte wcześniej postacie ujawniają się w nowych kombinacjach i objawiają się nowym kolorem, nowym kształtem, nową aktywnością enzymatyczną itp., a następnie testowane przez dobór naturalny na jego użyteczność lub szkodliwość. Zatem, Im bardziej populacja jest nasycona ukrytymi mutacjami, tym częściej pojawiają się nowe kombinacje mutantów jako nowe propozycje selekcji. Pula genowa stanowi rezerwę dziedzicznej zmienności populacji i jej potencjału ewolucyjnego.
Nowoczesny immunogenetyka w poszukiwaniu mechanizmów rozwoju i dziedziczenia tzw. odporności nabytej dochodzi do nieoczekiwanego, alternatywnego wniosku w duchu neolamarkizm. Wielu naukowców uważa, że mutacje w produkcji użytecznych immunoglobulin, które występują w limfocytach (i to somatyczny komórki) mogą zostać wyselekcjonowane i przeniesione do komórek rozrodczych i w ten sposób utrwalone u potomstwa. Okazuje się, że organizm macierzysty tworzy w komórkach somatycznych (limfocytach) skierowany, znana korzystna zmienność odporności i konsoliduje przez komórki rozrodcze u potomstwa (Steele i in., 2002). Te nowe dane, jeśli zostaną powszechnie zaakceptowane, mogą zmienić lub znacząco uzupełnić już ustalone poglądy na temat mechanizmów dziedzicznej zmienności.
(5) Wzorce ekologiczne w teorii ewolucji
Wreszcie, ekologia populacji pozwoliło to zrozumieć syntetycznej teorii ewolucji elementarne procesy zmienności, krzyżowania i dziedziczenia zachodzą w obrębie całej populacji organizmów. Populacja jest jednostką strukturalną procesu ewolucyjnego. Pomysł o fale demograficzne– wahania liczebności populacji, które regularnie występują podczas samoregulacji w łańcuchach pokarmowych, w związku z cyklami księżycowymi (miesięcznymi), sezonowymi (rocznymi), słonecznymi (11-letnimi) i innymi, w wyniku klęsk żywiołowych lub katastrof spowodowanych przez człowieka - pożary, susze, powodzie, zlodowacenia i tak dalej. Badanie fal populacyjnych przyczyniło się do zrozumienia tych zjawisk dryf genetyczny, tj. gwałtowna zmiana częstotliwości występowania niektórych genów w krytycznie małych populacjach. Z kolei koncepcję dryfu genetycznego zaczęto postrzegać jako ważny mechanizm szybkie lokalne przemiany ewolucyjne.
Ponadto z ekologii teoria ewolucji wykorzystuje wiedzę o przyczynach i mechanizmach izolacja geograficzna i środowiskowa. Izolacja to pojawienie się różnych przeszkód utrudniających swobodne przemieszczanie się osobników w obrębie populacji, a także pomiędzy bliskimi sobie populacjami, które nadal zachowują wymiana genetyczna na krańcach swoich zasięgów. Kiedy następuje izolacja, rozmnażanie następuje w obrębie izolatów – oddzielnych grup, tak że ustaje wymiana genów między nimi. Zachodzące wcześniej zmiany w genotypie utrwalają się u potomstwa i nasilają, co prowadzi do rozbieżności cech osobników (dywergencji) w obrębie dawnej jednorodnej populacji.
Generalnie możemy tak stwierdzić ewolucja biologiczna, podobnie jak pochodzenie życia, jest naturalna proces materialny rozwój Ziemi, kontynuacja ewolucji geologicznej. Byłoby całkiem logiczne znaleźć się w tych globalnych przemianach natury wspólne powody i siły napędowe.
Spróbujmy zastosować zasadę trójskładnikową, którą już przetestowaliśmy, do definicji istoty ewolucji biologicznej, aby znaleźć „trzy filary”, które wspierają ewolucję organizmów żywych. Po pierwsze, to genetyka – dziedziczność gatunku założycielskiego, na podstawie której zachodzą procesy ewolucyjne. Dane z zakresu genetyki molekularnej pokazują, że u blisko spokrewnionych gatunków podobieństwo (homologia) DNA wynosi ponad 99%. Po drugie, to epigenetyka – cały zespół różnych czynników środowiskowych, które kierują zarówno procesem mutacji (zmienność dziedziczna), jak i doborem naturalnym. Wreszcie, po trzecie, synergia – procesy samoorganizujące się. Aby znaleźć ten trzeci składnik w mechanizmach ewolucji, pamiętajmy, że w naukach Darwina najważniejsze miejsce zajmuje pojęcie rozbieżność, co oznacza rozbieżność spokrewnionych organizmów według nowo powstałych cech. Jest to bezpośredni wynik dziedzicznej zmienności i selekcji, która ostatecznie prowadzi do powstania nowych gatunków z gatunków poprzedników. Dywergencja w populacjach następuje w wyniku przypadkowych mutacji i rekombinacji genetycznych (te ostatnie zachodzą podczas rozmnażania płciowego). To całkiem oczywiste mutacje i rekombinacje geny reprezentują określoną manifestację biologiczną wahania w synergetyce i darwinizmie rozbieżność- to typowe rozwidlenie, po czym następuje elementarna „katastrofa” w postaci aktu samoorganizacji jakościowo nowy system. Jak widać, Darwin antycypował współczesne koncepcje synergiczne mechanizmów rozwoju już od samego procesu specjacja Opisywany przez niego zawiera elementy typowe samoorganizacja, A nowy rodzaj i istnieje jakościowo nowy system.
Podkreślamy, że podejście to nie oznacza sprowadzania ewolucji biologicznej wyłącznie do procesów samoorganizacji, jak próbowali to robić niektórzy autorzy. Co więcej, nie zaprzecza teorii doboru naturalnego Darwina. Wręcz przeciwnie, trójskładnikowa zasada określania istoty ewolucji wskazuje jej miejsce różnym czynnikom napędowym - czynnikom dziedzicznym, samoorganizującym się i środowiskowym.
Po raz kolejny jesteśmy przekonani, że rozwój jako ogólna forma ruchu materii ma również zasadniczo wspólne siły napędowe. Triadę „genetyka – synergetyka – epigenetyka” widzieliśmy w rozwój indywidualny organizmów (patrz 4.7), w rzekomych mechanizmach powstania życia na Ziemi (patrz 6.2) i znajdują się w tej samej kombinacji semantycznej w procesach globalnej ewolucji biologicznej.
Rozwiązanie:
Doświadczenia z przemianą substancji niskocząsteczkowych (cyjanki, acetylen, formaldehyd i fosforany) we fragment nukleotydowy potwierdzają hipotezę o spontanicznej syntezie monomerów kwasy nukleinowe z dość prostych substancji wyjściowych, które mogą znajdować się w odpowiednich warunkach wczesna ziemia.
Eksperyment, w którym otrzymano kwasy nukleinowe poprzez przepuszczenie wyładowania elektrycznego przez mieszaninę nukleotydów, dowodzi możliwości syntezy biopolimerów ze związków niskocząsteczkowych w warunkach wczesnej Ziemi.
Eksperyment, w którym poprzez zmieszanie biopolimerów w środowisku wodnym otrzymano ich kompleksy posiadające podstawy właściwości współczesnych komórek, potwierdza tezę o możliwości samoistnego tworzenia się koacerwatów.
6. Ustal zgodność między koncepcją pochodzenia życia a jego treścią:
2) stan ustalony
3) kreacjonizm
początek życia wiąże się z abiogennym powstawaniem substancji organicznych z nieorganicznych
gatunki żywej materii, takie jak Ziemia, nigdy nie powstały, ale istniały wiecznie
życie zostało stworzone przez Stwórcę w odległej przeszłości
życie przyniesione z kosmosu w postaci zarodników mikroorganizmów
Rozwiązanie:
Zgodnie z koncepcją ewolucja biochemiczna początek życia wiąże się z abiogenicznym powstawaniem substancji organicznych z nieorganicznych. Zgodnie z koncepcją stan stabilny, rodzaje żywej materii, takie jak Ziemia, nigdy nie powstały, ale istniały wiecznie. Zwolennicy kreacjonizm(od łac. creatio – stworzenie) wierzą, że życie zostało stworzone przez Stwórcę w odległej przeszłości.
7. Ustal zgodność między koncepcją pochodzenia życia a jego treścią:
1) teoria ewolucji biochemicznej
2) stan ustalony
3) kreacjonizm
pojawienie się życia jest wynikiem długotrwałych procesów samoorganizacji materii nieożywionej
problem pochodzenia życia nie istnieje, życie istniało zawsze
życie jest wynikiem boskiego stworzenia
życie ziemskie ma kosmiczne pochodzenie
Rozwiązanie:
Zgodnie z koncepcją ewolucja biochemicznażycie powstało w wyniku procesów samoorganizacji materii nieożywionej w warunkach wczesnej Ziemi. Zgodnie z koncepcją stan stabilny, problem pochodzenia życia nie istnieje, życie istniało zawsze. Zwolennicy kreacjonizm(od łac. creatio - stworzenie) wierzą, że życie jest wynikiem boskiego stworzenia.
Temat 25: Ewolucja systemów żywych
1. Historyczna ewolucja systemów żywych (filogeneza) to ...
spontaniczny
bezkierunkowe
odwracalny
ściśle przewidywalne
Rozwiązanie:
Historyczna ewolucja systemów żywych ma charakter spontaniczny, jest wynikiem wewnętrznych możliwości systemów żywych i działania sił doboru naturalnego.
2. Syntetyczna teoria ewolucji składa się strukturalnie z teorii mikro- i makroewolucji. Teoria badań mikroewolucji...
ukierunkowane zmiany w pulach genowych populacji
podstawowe wzorce rozwoju życia na Ziemi jako całości
przemiany ewolucyjne prowadzące do powstania nowych rodzajów
rozwój poszczególnych organizmów od urodzenia aż do śmierci
Rozwiązanie:
Teoria badań mikroewolucyjnych ukierunkowała zmiany w pulach genowych populacji znajdujących się pod wpływem różne czynniki. Mikroewolucja kończy się wraz z powstaniem nowych gatunków organizmów, bada zatem proces specjacji, a nie proces powstawania większych taksonów.
3. Według syntetycznej teorii ewolucji elementarnym zjawiskiem ewolucyjnym jest zmiana...
pula genów populacji
genotyp organizmu
pojedynczy gen
zestaw chromosomów organizmu
Rozwiązanie:
Elementarnym zjawiskiem ewolucyjnym jest zmiana puli genowej populacji. Jednostka od urodzenia do śmierci podlega jedynie rozwojowi ontogenetycznemu i nie ma możliwości ewolucji, dlatego zmiany w poszczególnych genach, zestawie genów (genotypach) lub zestawie chromosomów pojedynczego organizmu nie mogą być elementarnym zjawiskiem ewolucyjnym.
4. Historyczna ewolucja systemów żywych (filogeneza) to...
nieodwracalny
bezkierunkowe
nie spontaniczne
ściśle przewidywalne
Rozwiązanie:
Historyczna ewolucja systemów żywych jest nieodwracalna. Ewolucja organizmów opiera się na procesach probabilistycznych, w szczególności na występowaniu przypadkowych mutacji, dlatego jest nieodwracalna.
5. Czynnikiem ewolucyjnym, dzięki któremu ewolucja nabiera charakteru kierunkowego, jest (są)…
naturalna selekcja
proces mutacji
izolacja
fale demograficzne
Rozwiązanie:
Czynnikiem ewolucyjnym, dzięki któremu ewolucja nabiera charakteru kierunkowego, jest dobór naturalny.
Temat 26: Historia życia na Ziemi i metody badania ewolucji (ewolucja i rozwój układów żywych)
1. Morfologiczne metody badania ewolucji przyrody żywej obejmują badanie...
prymitywne narządy, które są słabo rozwinięte i utraciły swoje podstawowe znaczenie, co może wskazywać na formy przodków
formy reliktowe, czyli małe grupy organizmów o zespole cech charakterystycznych dla gatunków dawno wymarłych
wczesne etapy ontogenezy, w których stwierdza się więcej podobieństw między różnymi grupami organizmów
wzajemne przystosowanie się gatunków w zbiorowiskach naturalnych
Rozwiązanie:
Morfologiczne metody badania ewolucji są związane z badaniem cech strukturalnych narządów i organizmów o porównywanych formach, dlatego badanie słabo rozwiniętych i prymitywnych narządów, które utraciły swoje główne znaczenie, które może wskazywać na formy przodków, odnosi się do metod morfologia.
2. Biogeograficzne metody badania ewolucji przyrody żywej obejmują...
porównanie składu fauny i flory wysp z historią ich powstania
badanie narządów szczątkowych wskazujące na formy przodków organizmów żywych
porównanie wczesnych etapów ontogenezy organizmów różnych grup
badanie wzajemnej zdolności przystosowania się gatunków w zbiorowiskach naturalnych
Rozwiązanie:
Biogeograficzne metody badania ewolucji są związane z badaniem rozmieszczenia roślin i zwierząt na powierzchni naszej planety, dlatego porównywanie składu fauny i flory wysp z historią ich pochodzenia należy do metod biogeografii.
3. Konsekwencją pojawienia się eukariontów w historii życia na Ziemi jest...
uporządkowanie i lokalizacja aparatu dziedziczności w komórce
występowanie oddychania tlenowego
Rozwiązanie:
Konsekwencją pojawienia się eukariontów w historii życia na Ziemi jest uporządkowanie i lokalizacja aparatu dziedziczności w komórce. Protoplazma komórki eukariotycznej jest kompleksowo zróżnicowana, zawiera oddzielne jądro i inne organelle. Aparat chromosomalny zlokalizowany jest w jądrze, w którym koncentruje się główna część informacji dziedzicznej.
4. Ekologiczne metody badania ewolucji przyrody żywej obejmują badanie...
rola specyficznych adaptacji w populacjach modelowych
powiązania pomiędzy wyjątkowością flory, fauny i historią geologiczną terytoriów
prymitywne narządy, które są słabo rozwinięte i utraciły swoje podstawowe znaczenie
proces ontogenezy organizmów danego gatunku we wczesnych stadiach
Rozwiązanie:
Proces ewolucyjny to proces powstawania i rozwoju adaptacji. Ekologia, badając warunki bytowania i relacje pomiędzy organizmami żywymi w układach naturalnych czy populacjach modelowych, odkrywa znaczenie specyficznych adaptacji.
5. Konsekwencją fotosyntezy, najważniejszej aromorfozy w historii życia na Ziemi, jest...
tworzenie się osłony ozonowej
lokalizacja aparatu dziedzicznego w komórce
różnicowanie tkanek, narządów i ich funkcji
poprawa oddychania beztlenowego
Rozwiązanie:
Konsekwencją fotosyntezy, najważniejszej aromorfozy w historii życia na Ziemi, jest utworzenie się ekranu ozonowego, który powstał w wyniku gromadzenia się tlenu w atmosferze ziemskiej.
6. Poszerzenie areny życia w historii rozwoju świata organicznego ułatwiło...
gromadzenie się tlenu w atmosferze
pojawienie się eukariontów
gwałtowny spadek średniej temperatury powierzchni Ziemi
zalanie większości kontynentów wodami morskimi
Rozwiązanie:
Ekspansję areny życia w historii rozwoju świata organicznego ułatwiło gromadzenie się tlenu w atmosferze, a następnie utworzenie warstwy ozonowej. Ekran ozonowy chronił przed ostrym promieniowaniem ultrafioletowym, w wyniku czego organizmy rozwinęły bogatsze w energię górne warstwy zbiorników, następnie obszary przybrzeżne, a następnie dotarły do lądu. W przypadku braku osłony ozonowej życie było możliwe jedynie pod ochroną warstwy wody o grubości około 10 metrów.
7. Aromorfoza, która powstała podczas ewolucji świata organicznego, jest ...
pojawienie się fotosyntezy
pojawienie się przystosowań do zapylania
zmiana koloru kwiatów
pojawienie się ochronnych igieł i kolców
Rozwiązanie:
Aromorfozy to zmiany w strukturze i funkcjach narządów, które mają Ogólne znaczenie dla organizmu jako całości i podnieść poziom jego organizacji. Najważniejszą aromorfozą, która powstała podczas ewolucji świata organicznego, jest fotosynteza. Pojawienie się fotosyntezy doprowadziło do szeregu przemian ewolucyjnych, zarówno w organizmach żywych, jak i w środowisku: pojawienie się oddychania tlenowego, ekspansja odżywiania autotroficznego, nasycenie atmosfery ziemskiej tlenem, pojawienie się warstwy ozonowej, kolonizacja ziemi i powietrza przez organizmy.
Temat 27: Genetyka i ewolucja
1. Ustal zgodność między rodzajem zmienności a jej przykładem:
1) zmienność mutacyjna
wady rozwojowe układu nerwowego wynikające z naruszenia struktury obszaru chromosomu
zmiana koloru kwiatów w zależności od temperatury i wilgotności
kolor oczu dziecka różni się od koloru oczu rodziców, wynikający z kombinacji genów podczas rozmnażania płciowego
Rozwiązanie:
Wady rozwojowe układu nerwowego, które są wynikiem naruszenia struktury regionu chromosomowego, są zmiennością mutacyjną. Zmiany koloru kwiatów w zależności od temperatury i wilgotności powietrza stanowią zmienność modyfikacji.
2. Ustal zgodność między genotypami i ich przejawem w fenotypie:
dwa genotypy tej samej cechy, identycznie manifestujące się w fenotypie
dwa genotypy tej samej cechy, manifestujące się odmiennie fenotypem
dwa genotypy dla dwóch różnych cech, manifestujących się odmiennie w fenotypie
Rozwiązanie:
Geny alleliczne determinują rozwój różne opcje o tej samej charakterystyce są oznaczone tą samą literą alfabetu łacińskiego - Wielka litera, jeśli gen jest dominujący, i małą literą, jeśli gen jest recesywny. W fenotypie w równym stopniu manifestują się dwa genotypy - AA, Aa, ponieważ heterozygota Aa wykazuje cechę genu dominującego. Dwa genotypy tej samej cechy - AA, aa - manifestują się inaczej w fenotypie, ponieważ gen recesywny objawia się w stanie homozygotycznym aa.
3. Ustal zgodność między właściwością materiału genetycznego a przejawem tej właściwości:
1) dyskrecja
2) ciągłość
istnieją elementarne jednostki materiału dziedzicznego - geny
życie charakteryzuje się trwaniem istnienia w czasie, które zapewnia zdolność systemów żywych do reprodukcji
jednostki dziedziczności - geny - są zlokalizowane na chromosomach w określonej kolejności
Rozwiązanie:
Dyskrecja materiał genetyczny objawia się tym, że istnieją elementarne jednostki materiału dziedzicznego - geny. Życie jako zjawisko szczególne charakteryzuje się trwaniem istnienia w czasie, pewnym ciągłość, co zapewnia zdolność systemów żywych do samoreprodukcji - następuje zmiana pokoleń komórek, organizmów w populacjach, zmiana gatunków w systemie biocenoz, zmiana biocenoz tworzących biosferę
4. Ustal zgodność między typem cechy a jej zdolnością do ujawnienia się w pokoleniu:
1) niebieski kolor oczu jest cechą recesywną
2) brązowy kolor oczu jest cechą dominującą
nie występuje w stanie heterozygotycznym
pojawia się w stanie heterozygotycznym
nie występuje w stanie homozygotycznym
Rozwiązanie:
Cecha recesywna pojawia się tylko w stanie homozygotycznym, a w stanie heterozygotycznym cecha recesywna jest tłumiona przez dominującą i nie pojawia się. Cecha dominująca z całkowitą dominacją objawia się zarówno w stanach homozygotycznych, jak i heterozygotycznych.
5. Ustal zgodność między właściwością materiału genetycznego a przejawem tej właściwości:
1) liniowość
2) dyskrecja
geny są zlokalizowane na chromosomach w określonej kolejności
gen decyduje o możliwości rozwinięcia określonej cechy danego organizmu
materiał dziedziczny ma zdolność do samoreprodukcji
Rozwiązanie:
Liniowość materiał genetyczny objawia się tym, że geny znajdują się na chromosomach w określonej kolejności, czyli w porządku liniowym. Gen determinuje możliwość rozwinięcia określonej cechy danego organizmu, która go charakteryzuje dyskrecja jego akcje.
6. Ustal zgodność pomiędzy pojęciem a jego definicją:
1) genotyp
2) fenotyp
ogół wszystkich genów diploidalnego zestawu chromosomów organizmu
ogół wszystkich właściwości i cech konkretnego organizmu
zestaw genów haploidalnego zestawu chromosomów organizmu
Rozwiązanie:
Genotyp– całość wszystkich genów diploidalnego zestawu chromosomów organizmu. Fenotyp- całość wszystkich właściwości i cech konkretnego organizmu.
7. Ustal zgodność między rodzajem zmienności a jej przykładem:
1) zmienność mutacyjna
2) zmienność modyfikacji
zmiany w strukturze chromosomów podczas podziału komórki
zmiana koloru kwiatów podczas przenoszenia rośliny z warunków wewnętrznych do ciepłej, wilgotnej szklarni
zmiany związane z różnymi kombinacjami genów podczas rozmnażania płciowego
Rozwiązanie:
Zmiany w strukturze chromosomów podczas podziału komórki to zmienność mutacyjna. Zmiana koloru kwiatów po przeniesieniu rośliny z warunków wewnętrznych do ciepłej, wilgotnej szklarni oznacza zmienność modyfikacji.
Temat 28: Ekosystemy (różnorodność organizmów żywych jest podstawą organizacji i trwałości systemów żywych)
1. Ustal zgodność między grupą funkcjonalną organizmów w ekosystemie a przykładami organizmów:
1) konsumenci
2) producenci
3) rozkładacze
zające i wilki
rośliny zielone i bakterie fotosyntetyzujące
heterotroficzne bakterie i grzyby
glony i mikroorganizmy glebowe
Rozwiązanie:
Konsumenci są organizmy heterotroficzne spożywanie materii organicznej producentów lub innych konsumentów. Zające i wilki są konsumentami. Producenci to organizmy autotroficzne, zdolne do syntezy związków organicznych i budowania z nich swoich ciał. Do producentów należą rośliny zielone, glony i bakterie fotosyntetyzujące. Organizmy rozkładające to organizmy, które żyją z martwej materii organicznej i przekształcają ją z powrotem w związki nieorganiczne. Reduktory to bakterie i grzyby.
Główny element świata żywego, stanowiący podstawę strukturalną wszelkiej różnorodności związki organiczne Jest Tlen
Hipoteza jest... Teoretyczne założenie o istocie bezpośrednio obserwowanych zjawisk.
Głównym elementem świata ożywionego, stanowiącym podstawę strukturalną całej różnorodności związków organicznych, jest... Węgiel.
Głównym elementem świata żywego, który stanowi podstawę strukturalną całej różnorodności związków organicznych, jest -węgiel
Homeostaza jest jednym z charakterystyczne właściwości systemów - objawia się tym, że...
ISTNIEJĄ MOLEKULARNE MECHANIZMY UTRZYMANIA ZAWARTOŚCI WEWNĘTRZNEGO UKŁADU SEKSUALNEGO -
Do pomiarów galaktycznych i międzygalaktycznych. odległości są używane jako jednostki długości :
lata świetlne
Nauki przyrodnicze charakteryzują się... ^ Obowiązkowe eksperymentalne testowanie zdobytej wiedzy.
Ziemia różni się od innych planet grupa naziemna(Merkury, Wenus i Mars) ...
dużo płynu na powierzchni
Ustal podobieństwo organizmów na etapie rozwoju embrionalnego
Hierarchia systemów naturalnych przejawia się w tym, że
Systemy niższego poziomu organizacji są częścią systemów wyższego poziomu
Badanie procesów ewolucji w biologii molekularnej Analizować stopień podobieństwa kwasów nukleinowych różnych organizmów
Historyczna ewolucja systemów żywych (filogeneza) jest...Nieodwracalny
Badanie procesów ewolucji w biologii molekularnej... ^ Przeanalizuj stopień podobieństwa kwasów nukleinowych różnych organizmów.
Uczciwość- jeden z… Wszystkie właściwości systemu są ze sobą powiązane i wzajemnie się warunkują
Historyczna ewolucja systemów żywych (filogeneza) jest... spontaniczny
Zarówno w mechanicznym, jak i elektromagnetycznym obrazie świata istniały idee przypadkowości jako odzwierciedlenia niekompletności istniejącej wiedzy; jedyny możliwy rozwój zdarzeń dla danego stanu początkowego.
Integralność – jedna z charakterystycznych właściwości systemów naturalnych – przejawia się w tym, że... System zyskuje nowe właściwości, których nie mają komponenty, a powstają w wyniku interakcji komponentów
^ W stronę niszczycielskiego zanieczyszczenia środowisko wskazówki...Budowa autostrad.
Do zjawisk samoorganizacji zalicza się:
- przemiana chemiczna Reakcje oscylacyjne przy wysokich stężeniach odczynników
Substancja bioinertna obejmuje woda rzeczna
Zasada uniwersalnego ewolucjonizmu obejmuje stanowisko, że...
Wszechświat jako całość i wszystkie jego podsystemy podlegają ciągłemu rozwojowi
Komety, które czasami pojawiają się na ziemskim niebie, ... krążą wokół Słońca
Kosmogonia bada pochodzenie… ciał niebieskich i ich układów
Do zasad uniwersalnego ewolucjonizmu zalicza się stanowisko, że Wszechświat jako całość i wszystkie jego podsystemy podlegają ciągłemu rozwojowi
Kosmologia bada pochodzenie i rozwój WSZECHŚWIAT
Statystyczne teorie naukowe obejmują... ^ Mechanika kwantowa.
Warunki niezbędne do samoorganizacji w systemie obejmują...
SILNA NIERÓWNOWAGA UKŁADÓW
Koncepcja zrównoważonego rozwoju ludzkości oznacza... Kompromis pomiędzy dążeniem ludzkości do zaspokojenia swoich potrzeb a koniecznością ochrony biosfery.
Program badań kontinuum Arystotelesa opierał się na ideach, które...! ^ Pusta przestrzeń nie istnieje, materia wypełnia wszystko bez pustek; Materia jest ciągła i nieskończenie podzielna;
Kosmologia bada pochodzenie i rozwój Wszechświat
Jakiekolwiek formy życia zostaną odkryte(lub nieodkryte) przez wyprawę na inne światy, nie może to być zaprzeczeniem koncepcji pochodzenia życia na Ziemi, znanej jako… koncepcja kreacjonizmu;teoria ewolucji biochemicznej
Warunki niezbędne do samoorganizacji w systemie obejmują... ^ Silna nierównowaga systemu
Liniowość materiału genetycznego przejawia się w tym, że…
JEDNOSTKI DZIEDZICTWA - GENY - LOKALIZACJA W CHROMASOMACH W OKREŚLONEJ KOLEJNOŚCI -; JEDYNY MOŻLIWY ROZWÓJ ZDARZEŃ W DANYM STANIE POCZĄTKOWYM -
Różne poziomy hierarchiczne jednego systemu-Wszechświata
Cząsteczka jest -strukturalna jednostka materii
Megaświat, makroświat, mikroświat to... RÓŻNE POZIOMY HIERARCHICZNE JEDNEGO SYSTEMU - WSZECHŚWIAT
Megaświat, makroświat, mikroświat - to jest
Formuła molekularna glukoza (C6H12O6) i każda inna substancja oznacza...substancje.
SKŁAD JAKOŚCIOWY I ILOŚCIOWY
Megaświat, makroświat, mikroświat - to jest...Różne poziomy hierarchiczne jednego systemu – Wszechświata
Pomiędzy obiektami megaświata dominuje interakcja Grawitacyjny
Masa zwykłej materii, dostępnej do bezpośredniej obserwacji przez teleskopy i skupionej głównie w gwiazdach, stanowi ____________ całkowitej materii wszechświatamniej niż 5%
Nie jest to jedna z form energii znanych w fizyce i w ogóle w naukach przyrodniczych...Pozytywna i negatywna psychoenergia.
Nasza galaktyka jest...Spirala
Nasze Słońce jest obecnie na etapie... przewaga termojądrowych reakcji syntezy jąder helu z jądrami wodoru
Newton wprowadził pojęcie „czasu absolutnego”. Czas jest absolutny, ponieważ...
Jego przepływ jest całkowicie niezależny od ciał materialnych i tego, co się z nimi dzieje.
Nie jest to jedna z form energii znanych w fizyce i w ogóle w naukach przyrodniczych... ^ Pozytywna i negatywna psychoenergia.
Nasza galaktyka jest -Spirala
Newton wprowadził pojęcie „przestrzeni absolutnej”. Przestrzeń jest Absolutna, ponieważ jego właściwości nie zależą od ciał materialnych i tego, co się z nimi dzieje
Newton wprowadził pojęcie „czasu absolutnego”. Czas jest absolutny, ponieważ...
jego przepływ jest całkowicie niezależny od ciał materialnych i tego, co się z nimi dzieje
Żadne dane naukowe uzyskane w wyniku poszukiwań życia na innych światach nie mogą obalić koncepcji pochodzenia życia, zwanej kreacjonizmem, ponieważ... w zasadzie niezaprzeczalny; nie może być przedmiotem rozważań naukowych
Kosmologia naukowa zaczęła się rozwijać w...Renesans oparty na systemie heliocentrycznym Kopernika
Żadne dane naukowe uzyskane w wyniku poszukiwań życia na innych światach nie mogą potwierdzić koncepcji pochodzenia życia na Ziemi, znanej jako...
Koncepcja stanu ustalonego
Hipoteza stałego, spontanicznego generowania
Newton przypisywany wprowadzonym do koncepcji własności „czasu absolutnego”. Niezależność od ciał materialnych
Newton wprowadził pojęcie „czasu absolutnego”. Czas jest absolutny, ponieważ...
JEGO PRZEPŁYW JEST CAŁKOWICIE NIEZALEŻNY OD CIAŁ MATERIAŁOWYCH I TEGO, CO SIĘ Z NIMI DZIEJE -
Obserwacja– uzyskanie informacji nt stan skupienia, kolor, zapach substancji uwalnianych w reakcji chemicznej.
Niuton nie atrybutował właściwość, którą wprowadził do pojęcia „czasu absolutnego”... zależą od ruchu obserwatora
Kosmologia naukowa zaczęła się rozwijać ^ XX wiek w oparciu o ogólną teorię względności.
Newton przypisał pojęciu „Czasu Absolutnego”, że wprowadził własność…
Niezależnie od ciał materialnych
Żadne dane naukowe uzyskane podczas poszukiwań życia na innych światach nie mogą potwierdzić koncepcji pochodzenia życia na Ziemi. Znany jako koncepcja stanu stacjonarnego, hipoteza ciągłego generowania spontanicznego.
Newton wprowadził pojęcie przestrzeni absolutnej. Przestrzeń jest Absolutem, ponieważ...
Jego właściwości nie zależą od ciał materialnych i tego, co się z nimi dzieje��
Ogólna teoria względności, w przeciwieństwie do szczególnej teorii względności, ma zastosowanie... w bardzo mocnym pola grawitacyjne
Jednym z empirycznych dowodów słuszności ogólnej teorii względności jest-
Jednym ze wskaźników globalnego kryzysu ekologicznego naszych czasów jest
antropogeniczne zmiany składu atmosfery
Jeśli chodzi o symetrie przestrzeni, prawdziwe jest stwierdzenie, że przestrzeń...
izotropowo
masywne ciała materialne swoją obecnością i ruchem zaginają geometrię czasoprzestrzeni
Anomalne przemieszczenie peryhelium orbity Merkurego
Jeśli chodzi o symetrie przestrzeni, prawdziwe jest stwierdzenie, że przestrzeń...Izotropowo
Jednym z empirycznych dowodów słuszności ogólnej teorii względności jest...Anomalne przemieszczenie peryhelium orbity Merkurego
Jednym ze wskaźników globalnego kryzysu ekologicznego naszych czasów jest...
degradacja zasobów wodnych, gruntowych i leśnych
Jeden z powodów aktywność chemiczna organiczne molekuły jest w nich obecność grupy funkcyjne, które wynikają ze zdolności węgla …
formularz aktywne połączenia z atomami tlenu, azotu, siarki
tworzą cztery wiązania z innymi atomami
Jednym z empirycznych dowodów słuszności ogólnej teorii względności jest...Anomalne przemieszczenie peryhelium orbity Merkurego
Jednym z kryteriów klasyfikacji cząstek elementarnych jest...Dożywotni
Jedną z konsekwencji zasady równoważności wywodzącej się z ogólnej teorii względności jest to, że... Masa i energia są równoważne, powiązane są zależnością Einsteina��
Jedną z konsekwencji zasady równoważności wywodzącej się z ogólnej teorii względności jest to, że... w silnych polach grawitacyjnych tempo czasu zwalnia
Podstawowe informacje o składzie i dynamice głębokie wnętrzności Przynosi ziemię... analiza propagacji fal sejsmicznych
Różnica między popularną w XIX wieku koncepcją świata eteru a newtonowską koncepcją przestrzeni absolutnej polegała na tym, że zakładano, że eter... interakcji z ciałami materialnymi
Eksperyment Michelsona-Morleya polegający na pomiarze prędkości światła względem poruszającego się obserwatora, przeprowadzony wielokrotnie pod koniec XIX wieku, wykazał...pełna zgodność koncepcji eteru świata z rzeczywistością naszego świata
Jedną z konsekwencji środowiskowych rewolucji neolitycznej jest technogenne zanieczyszczenie środowiska
Fakt obserwacyjny mówi o ekspansji wszechświata...
-Przesunięcie ku czerwieni w widmach wszystkich odległych galaktyk-
Jedną z konsekwencji środowiskowych rewolucji neolitycznej (10-8 p.n.e.) jest
DESERTYFIKACJA TERYTORIÓW-
Jednym z empirycznych dowodów słuszności ogólnej teorii względności jest
-odchylenie promieni świetlnych przechodzących w pobliżu słońca
Obecność i funkcjonowanie organizmów żywych sygnalizowane jest obecnością... Gęsta atmosfera z rozwiniętą cyrkulacją
- Wystarczająca ilość dużych otwartych zbiorników
Obiektem zajmującym większą część pola na tym zdjęciu nie może być nasza Galaktyka (Droga Mleczna), ponieważ znajdujemy się wewnątrz naszej Galaktyki, a zdjęcie wyraźnie zostało zrobione z zewnątrz
Obiektem zajmującym większą część pola widzenia na tym zdjęciu nie może być nasza Galaktyka (Droga Mleczna) ponieważ nasza Galaktyka jest eliptyczna, a zdjęcie przedstawia spiralę
Jednym z kryteriów klasyfikacji cząstek elementarnych jest kręcić się
Jednym z empirycznych dowodów słuszności ogólnej teorii względności jest...Anomalne przemieszczenie peryhelium orbity Merkurego
Fakt obserwacyjny mówi o rozszerzaniu się Wszechświata... ^ Przesunięcie ku czerwieni w widmach wszystkich odległych galaktyk
^ Jedną z konsekwencji zasady równoważności, wywodzącej się z ogólnej teorii względności, jest to, że... masywne ciała materialne swoją obecnością i ruchem zaginają geometrię czasoprzestrzeni
Para genów recesywnych determinujących rozwój jednej cechy będzie zbiorem...aha
Koncepcji pochodzenia życia, znanej jako hipoteza panspermii, można sprzeciwić się odkryciem… faktu, że nigdzie w kosmosie nie ma życia poza Ziemią; żywe organizmy typu ziemskiego w układzie planetarnym Syriusza i życie o zupełnie innej bazie chemicznej na Marsie.
Jako para geny alleliczne to geny, które decydują o... brązowym i niebieskim kolorze oczu
Pseudonauka różni się od prawdziwej nauki Odporny na krytykę i chwalenie swoich wyników
Postulat, który został wprowadzony do ogólnej teorii względności, to stanowisko Niezmienność praw natury w różnych układy inercyjne odliczanie
Przykładem zmienności mutacyjnej jest zmiana Struktury chromosomów podczas podziału komórki
Pseudonauka różni się od prawdziwej nauki... Odporność na krytykę i pochwałę swoich wyników��
Zgodnie z zasadą szeroko pojętej komplementarności są one ze sobą powiązane... metody dedukcji i indukcji w wiedzy naukowej
Zgodnie z zasadą szeroko pojętej komplementarności są one ze sobą powiązane... Mechanika kwantowa i klasyczna��
Pomysł charakterystyczne jest to, że Ziemia zajmuje zwyczajną, niczym nie wyróżniającą się pozycję we Wszechświecie.
Pomysł charakterystyczne jest to, że każdą zmianę w świecie można sprowadzić do ruchów niektórych ciałek ...atomistyczne nauki Leukipposa i Demokryta
Idea czasoprzestrzeni jako pełnoprawnego, aktywnego, złożonego składnika świata materialnego jest charakterystyczna dla:nowoczesny obraz naukowy pokój
Przeprowadzanie wielokrotnych eksperymentów w celu ustalenia prawdziwości hipotezy jest przykładem zastosowania zasady w nauce...Weryfikacja
Przewidywania szczególnej teorii względności i mechaniki klasycznej są zbieżne, pod warunkiem, że...Prędkości podróżowania są znacznie mniejsze niż prędkość światła
Pod względem wielkości Ziemia plasuje się_______ wśród ośmiu planet Układu Słonecznego.piąty
Odkrycie... (1 z 2) może zaprzeczyć koncepcji pochodzenia życia, znanej jako hipoteza panspermii.
Organizmy żywe typu lądowego na Marsie i życie o zupełnie innej podstawie chemicznej w układzie planetarnym Syriusza
^ Fakt, że nigdzie poza Ziemią nie ma życia w kosmosie
Charakterystyczne jest rozumienie przestrzeni i czasu jako pewnych substancji, które istnieją niezależnie i nie są związane z ciałami materialnymi … Naturalny filozoficzny obraz świata Arystotelesa
Przykładem zmienności modyfikacji jest zmiana-kolory skrzydeł motyli pod wpływem warunków środowiskowych
Przykładem zmienności mutacyjnej jest zmiana...Struktury chromosomów podczas podziału komórki.
Przykładem zmienności modyfikacji jest zmiana...
Barwy skrzydeł motyli pod wpływem warunków środowiskowych
Idea czasoprzestrzeni jako pełnoprawnego, aktywnego, złożonego składnika świata materialnego jest charakterystyczna dla...
Współczesny naukowy obraz świata
Specjalne prognozy Teorie względności i mechaniki klasycznej są zbieżne, pod warunkiem, że... ^Prędkość ruchu jest znacznie mniejsza niż prędkość światła.
Postulat, który został wprowadzony do ogólnej teorii względności, to stanowisko o (o)... Równoważność ruchu przyspieszonego i spoczynku w polu grawitacyjnym.
Pojmowanie przestrzeni i czasu jako jedynie układu relacji pomiędzy ciałami materialnymi jest charakterystyczne dla...nauki starożytnych greckich atomistów
Producenci w ekosystemie lądowym to... rośliny
Para genów recesywnych determinujących rozwój jednej cechy będzie zbiorem... aha
Według współczesnych wyobrażeń, za około 5 miliardów lat Słońce wyczerpie główne rezerwy swojego paliwa termojądrowego i... zamieni się w białego karła
Idea czasoprzestrzeni jako pełnoprawnego, aktywnego, złożonego składnika świata materialnego jest charakterystyczna dla... współczesny naukowy obraz świata
Przeprowadzanie powtarzanych eksperymentów w celu ustalenia prawdziwości hipotezy jest przykładem zastosowania tej zasady w nauce
Weryfikacja
Para genów recesywnych determinujących rozwój jednej cechy będzie zbiorem... aha.
Producenci w ekosystemie lądowym to... Rośliny.
Charakterystyczne jest rozumienie przestrzeni i czasu jako pewnych substancji, które istnieją niezależnie i nie są związane z ciałami materialnymi. Mechaniczny naukowy obraz świata.
Założenie, że światło ma właściwości kwantowe(korpuskularne) okazały się konieczne do wyjaśnienia ustalonych eksperymentalnie praw... EFEKT FOTOGRAFICZNY
Rozpad wolnego neutronu na elektron i antyproton jest prawnie zabroniony... ZACHOWANIE ŁADUNKU ELEKTRYCZNEGO
Ekspansja Wszechświata trwa...Jednorodny, bez specjalnego lub wyróżnionego centrum ekspansji.
Poszerzeniu areny życia w historii rozwoju świata organicznego sprzyjały... gromadzenie się tlenu w atmosferze
Rola dla żywej natury anomalnej gęstości lodu – stanu stałego wody – objawia się tym, że możliwe jest... ochrona życia w zamarzniętych zbiornikach wodnych
Wynikiem procesu różnicowania wiedzy naukowej jest... Biochemia
Efektem różnicowania wiedzy naukowej jest...Embriologia
Wynik procesu różnicowania wiedza naukowa jest … mechanika kwantowa
Z
Z termodynamicznego punktu widzenia, przedsiębiorstwa elektroenergetyczne... przekształcać niewygodne formy energii częściowo w energię elektryczną, a częściowo w formy energii niskiej jakości
Badania nad synergią...Wzorce samoorganizacji w systemach naturalnych i społecznych.
Syntetyczna teoria ewolucji składa się strukturalnie z teorii mikro- i makroewolucji. Teoria badań mikroewolucji… ukierunkowała zmiany w pulach genowych populacji
Według syntetycznej teorii ewolucji elementarnym zjawiskiem ewolucyjnym jest zmiana... pula genów populacji
Konsekwencją pojawienia się eukariontów w historii życia na Ziemi jest.... uporządkowanie i lokalizacja aparatu dziedziczności w komórce
Konsekwencją fotosyntezy, najważniejszej aromorfozy w historii życia na Ziemi, jest... tworzenie się osłony ozonowej
Istotą odkrycia dokonanego przez amerykańskiego astronoma-obserwatora E. Hubble'a w latach 20. XX wieku, które stało się empiryczną podstawą rozwoju naukowej kosmologii, było to, że galaktyki... oddalają się od siebie im szybciej, tym bardziej odległość między nimi
Według współczesnych koncepcji kosmologicznych ekspansja Wszechświata... przyspiesza i nastąpi, im dalej, tym szybciej
^ Według jednego z koncepcje historyczne pochodzenie życia – hipoteza panspermii, życie...Przywieziony na Ziemię z kosmosu.
Zgodnie z ideami mechanicznego obrazu świata, gdyby można było usunąć z Wszechświata wszystkie ciała materialne, to...Właściwości przestrzeni i czasu pozostałyby niezmienione
Według współczesnych koncepcji ewolucji biologicznej, pierwsze organizmy żywe były beztlenowe, czyli...
Według teorii względności, przechodząc z jednego układu odniesienia do drugiego, fakt, że… światła w połowie Kalifornii zgasły w momencie, gdy Maria Iwanowna Pietrowa w Moskwie przekręciła włącznik w swoim mieszkaniu
Według szczególnej teorii względności, przy przejściu z jednego systemu raportowania do drugiego, może nastąpić zmiana...Odległość między dwoma zdarzeniami
Zgodnie z zależnością niepewności W. Heisenberga...
Nie da się dokładnie i jednocześnie zmierzyć współrzędnych i pędu obiektu materialnego
Zgodnie z jednym ze sformułowań drugiej zasady termodynamiki ...
Niemożliwe jest całkowite przekształcenie energii cieplnej w użyteczną pracę
Badania nad synergią...
Struktury makrokosmosu są... Cząsteczka wody
Zgodnie z relacją niepewności Heisenberga dokładna wartość pędu konkretnego obiektu... Znajomość jego współrzędnych czyni go niedokładnym��
Przykładem wykorzystania metody jest wykonanie makiety elektrowni gazowej... modelowanie
Zgodnie z relacją niepewności W. Heisenberga dokładna znajomość pędu konkretnego obiektu... sprawia, że znajomość jego współrzędnych jest niedokładna
przywieziony na Ziemię z kosmosu
Zdolność atomów węgla do łączenia się z atomami azotu i fosforu w celu utworzenia... istnienie grup funkcyjnych. spowodowanie...
Układ słoneczny jest zorganizowany więc... OGROMNE PLANETY ZNAJDUJĄ SIĘ ŚREDNIO DZIESIĄTKI RAZY DALSZE OD SŁOŃCA NIŻ PLANETA ZIEMSKA
Zdolność atomów węgla do wiązania się między sobą i innymi atomami... tworzenie konstrukcji podstawowej...
Zdolność atomu węgla do tworzenia związków organicznych... możliwość tworzenia wysokocząsteczkowych...
Podobieństwo pomiędzy Wielkim Wybuchem (procesem, podczas którego nasz Wszechświat powstał i nabył swoje właściwości) a zwykłą eksplozją pocisku artyleryjskiego polega na tym, że... różne strony fragmenty eksplodowanego pocisku
Wśród uczestników dyskusji na temat ruchu panował pogląd, że ruch trwa tak długo, jak działa siła, która go spowodowała.... Arystoteles; Heraklit
Zgodnie z jednym ze sformułowań drugiej zasady termodynamiki ... niemożliwe jest całkowite przekształcenie energii cieplnej w użyteczną pracę
Zgodnie z wyobrażeniami o przestrzeni i czasie, tkwiącymi w mechanicznym obrazie świata, gdyby z Wszechświata zniknęły wszystkie ciała materialne, wówczas przestrzeń i czas... nadal istniałyby, a ich właściwości nie uległyby żadnej zmianie
Zgodnie z wyobrażeniami o przestrzeni i czasie, tkwiącymi w mechanicznym obrazie świata, gdyby z Wszechświata zniknęły wszystkie ciała materialne, wówczas przestrzeń i czas... Nadal istniałyby, a ich właściwości w ogóle by się nie zmieniły
Według współczesnych koncepcji kosmologicznych ekspansja Wszechświata... przyspiesza i będzie to kontynuować, tym szybciej
Zgodnie z koncepcją dualizmu korpuskularno-falowego każdy obiekt materialny... ma właściwości zarówno fali, jak i cząstki, ale w konkretnym eksperymencie wykazuje albo jedno, albo drugie
Zgodność metody poznania naukowego z przykładem jej zastosowania:
Modelowanie– zapisywanie wzorów strukturalnych związków bioorganicznych;
Eksperyment– wielokrotna synteza cząsteczek organicznych charakterystycznych dla organizmów żywych z gazów nieorganicznych;
Pomiary grubości warstwy skał osadowych nagromadzonych na przestrzeni dziejów Ziemi
Według współczesnych koncepcji ewolucji biologicznej pierwszymi organizmami żywymi były beztlenowce, czyli... Mogli żyć bez tlenu
Syntetyczna teoria ewolucji składa się strukturalnie z teorii mikroewolucji i makroewolucji. Teoria badań makroewolucji... Transformacje ewolucyjne w długim okresie historycznym na poziomach powyżej poziomu gatunku
Mieszanina prosta substancja wyświetla się formuła...O3
Struktury makrokosmosu są... Bakteria.
Badania nad synergią... Wzorce samoorganizacji w systemach naturalnych i społecznych.
Najdokładniejsze oszacowanie wieku Ziemi pochodzi z... Pomiary stężenia izotopów promieniotwórczych i produktów ich rozpadu w skałach ziemnych i meteorytach.
Według współczesnych koncepcji ewolucji biologicznej pierwszymi organizmami żywymi były heterotrofy, czyli... ^ Jedli gotowe substancje organiczne.
Skład prostej substancji przedstawia wzór...O 3
Przykładem wykorzystania metody jest wykonanie makiety elektrowni gazowej...
Symulacja
Istotą odkrycia dokonanego przez amerykańskiego astronoma obserwacyjnego E. Hubble'a w latach 20. XX wieku, które stało się empiryczną podstawą rozwoju kosmologii naukowej, było to, że galaktyki...oddalają się od siebie, im szybciej, tym większa jest między nimi odległość
Systematyczny charakter wiedzy naukowej znajduje odzwierciedlenie w Przejrzysta struktura wiedzy i sposobów poznania
Złożony cząstka elementarna Jest -proton
Rola dla żywej natury anomalnej gęstości lodu – stanu stałego wody – objawia się tym, że możliwe jest... Ochrona życia w zamarzniętych zbiornikach wodnych
Zgodnie z koncepcją determinizmu mechanicznego-każde przyszłe wydarzenie można dokładnie przewidzieć, znając stan początkowy i korzystając z praw mechaniki
Najniższa entropia dla równych ilości ma(-y) Lodowata woda
Stan układu zbliżający się do punktu bifurkacji charakteryzuje sięniestabilność
Najdokładniejsze oszacowanie wieku Ziemi uzyskuje się metodą-pomiary stężenia izotopów promieniotwórczych i produktów ich rozpadu w skałach ziemnych i meteorytach
Według jednej z historycznych koncepcji powstania życia - koncepcji kreacjonizmu-życie jest wynikiem boskiego stworzenia
Syntetyczna teoria ewolucji składa się strukturalnie z teorii mikro- i makroewolucji.
Właściwość odzwierciedlająca proces zdobywania wiedzy naukowej poprzez dowody logiczne i weryfikację eksperymentalną nazywa się... niezawodność
Struktury mikroświata są... cząsteczki wody
Skład prostej substancji odzwierciedla formuła... O 3
Zgodnie z jednym ze sformułowań drugiej zasady termodynamiki, podczas procesu zachodzącego w układzie izolowanym... entropia wzrasta
Stan układu zbliżający się do punktu bifurkacji charakteryzuje się...
niestabilność
Według jednej z historycznych koncepcji powstania życia - hipotezy panspermii, życie... przywieziony na Ziemię z kosmosu
Według współczesnych koncepcji ewolucji biologicznej pierwszymi organizmami żywymi były beztlenowce, tj.?
- BYLI W STANIE ŻYĆ BEZ TLENU-
Według syntetycznej teorii ewolucji i teorii Karola Darwina głównym czynnikiem napędzającym ewolucję jest...
NATURALNA SELEKCJA-
Według współczesnej kosmologii wiek wszechświata wynosi kilka razy wiek słońca i ziemi
Systematyczny charakter wiedzy naukowej znajduje odzwierciedlenie w Przejrzysta struktura wiedzy o sposobach poznania







