Anatomia
Kora nowa zawiera dwa główne typy neuronów: neurony piramidalne (~80% neuronów kory nowej) i neurony wewnętrzne (~20% neuronów kory nowej).
Struktura kory nowej jest stosunkowo jednorodna (stąd alternatywna nazwa: „izokora”). U człowieka posiada sześć poziomych warstw neuronów, różniących się rodzajem i charakterem połączeń. Pionowo neurony łączą się w tzw kolumny korowe. Delfiny kora nowa ma 3 poziome warstwy neuronów.
Zasada działania
Zasadniczo nowa teoria Algorytmy działania kory nowej zostały opracowane w Menlo Park w Kalifornii, USA (Dolina Krzemowa) przez Jeffa Hawkinsa. Teoria hierarchicznej pamięci tymczasowej została zaimplementowana w oprogramowaniu w postaci algorytmu komputerowego, który jest dostępny do wykorzystania na podstawie licencji w serwisie numenta.com.
- Ten sam algorytm przetwarza wszystkie zmysły.
- Funkcja neuronu obejmuje pamięć w czasie, coś w rodzaju związków przyczynowo-skutkowych, hierarchicznie rozwijających się w coraz większe obiekty z mniejszych.
Zobacz też
- Starożytna kora
Spinki do mankietów
- W. Mountcastle „Zasada organizacyjna funkcji mózgu: moduł elementarny i system rozproszony”
- Tłumaczenie na język rosyjski artykułu „Hierarchiczna pamięć tymczasowa” ze strony Numenta.com
Fundacja Wikimedia. 2010.
Zobacz, co „Nowa skorupa” znajduje się w innych słownikach:
NEOCORTEX (NOWA KORA)- Ewolucyjnie najnowsza i najbardziej złożona tkanka nerwowa. Płaty czołowy, ciemieniowy, skroniowy i potyliczny mózgu składają się z kory nowej... Słownik w psychologii
- (cortex hemispheria cerebri), paliusz lub płaszcz, warstwa istoty szarej (1–5 mm) pokrywająca półkule mózgu ssaków. Ta część mózgu, która rozwinęła się późno w ewolucji, odgrywa niezwykle ważną rolę w... ... Biologiczny słownik encyklopedyczny
Kora: W Wikisłowniku znajduje się hasło „kora”. W biologii: Kora to zewnętrzna część pnia drzewa. Kora dużych n... Wikipedia
kora- mózg: kora (kora mózgowa) górna warstwa półkul mózgowych, składająca się głównie z komórki nerwowe o orientacji pionowej (komórki piramidalne), a także z wiązek doprowadzających (dośrodkowych) i odprowadzających... ... Świetna encyklopedia psychologiczna
Warstwa istoty szarej o grubości 1–5 mm pokrywająca półkule mózgowe ssaków i ludzi. Ta część mózgu (patrz: mózg), która rozwinęła się na późniejszych etapach ewolucji królestwa zwierząt, odgrywa wyłącznie... ... Wielka encyklopedia radziecka
Mózg (cortex cerebri, PNA, LNH; istota korowa, BNA, JNA; synonim: K. cerebral hemispheres, K. cerebrum, mantle, cloak) warstwa powierzchniowa półkul mózgowych, utworzona przez jej istotę szarą; odgrywa ważną rolę w... Encyklopedia medyczna
- (kora mózgowa) istota szara zlokalizowana na powierzchni półkul mózgowych i składająca się z komórek nerwowych (neuronów), neurogleju, połączeń międzyneuronowych kory, a także naczyń krwionośnych. K.b. m. zawiera sekcje centralne (korowe)... ... Encyklopedia medyczna
NEOKORTEX NEOKORTEX
(od neo... i łac. kora - kora, muszla), nowa kora, neopallium, podstawowa. część kory mózgowej. N. zapewnia najwyższy poziom koordynacji funkcji i tworzenia mózgu złożone kształty zachowanie. W procesie ewolucji N. pojawia się najpierw u gadów, u których jest niewielkich rozmiarów i ma stosunkowo prostą budowę (tzw. kora boczna). N. ma typową budowę wielowarstwową tylko u ssaków, w której składa się z 6-7 warstw komórek (piramidalnej, gwiaździstej, wrzecionowatej) i dzieli się na płaty: czołowy, ciemieniowy, skroniowy, potyliczny i środkowopodstawny. Z kolei udziały podzielone są na obszary, podobszary i pola, które różnią się między sobą struktura komórkowa i połączenia z głębokimi częściami mózgu. Wraz z włóknami projekcyjnymi (pionowymi) neurony N. tworzą włókna asocjacyjne (poziome), które u ssaków, a zwłaszcza u ludzi, są zebrane w anatomicznie odrębne pęczki (na przykład wiązka potyliczno-czołowa), zapewniając jednoczesną skoordynowaną aktywność różnych typy. strefy N. N. składa się z najbardziej złożonej kory asocjacyjnej, krawędzie w procesie ewolucji doświadczają największego wzrostu, podczas gdy pierwotne pola czuciowe N. są stosunkowo zmniejszone. (patrz PÓŁKULE KOROWE MÓZGU).
.(Źródło: „Biological Encyclopedic Dictionary”. Redaktor naczelny M. S. Gilyarov; Redakcja: A. A. Babaev, G. G. Vinberg, G. A. Zavarzin i inni - wyd. 2, poprawione - M.: Sov. Encyclopedia, 1986.)
Zobacz, co „NEOCORTEX” znajduje się w innych słownikach:
Kora nowa...
Nowa kora (synonimy: neocortex, isocortex) (łac. neocortex) nowe obszary kory mózgowej, które u niższych ssaków są tylko zarysowane, ale u ludzi stanowią główną część kory. Nowa kora znajduje się w górnej warstwie półkul... ... Wikipedia
kora nowa- 3.1.15 kora nowa: Nowa kora mózgowa, która zapewnia realizację intelektualnej aktywności umysłowej poprzez ludzkie myślenie. 3.1.16 Źródło… Słownik-podręcznik terminów dokumentacji normatywnej i technicznej
- (kora nowa; neo + łac. kora korowa) patrz Nowa kora ... Duży słownik medyczny
kora nowa- y, h. Ewolucyjna innowacja i złożoność tkanek nerwowych tworzących czoło, a tym samym mosznę i inne części mózgu... Ukraiński słownik Tlumach
NEOCORTEX (NOWA KORA)- Ewolucyjnie najnowsza i najbardziej złożona tkanka nerwowa. Płaty czołowy, ciemieniowy, skroniowy i potyliczny mózgu składają się z kory nowej... Słownik objaśniający psychologii
Łuki, paleo, kora nowa... Słownik ortografii – podręcznik
kora- kora mózgowa: kora (kora mózgowa) górna warstwa półkul mózgowych, składająca się głównie z komórek nerwowych o orientacji pionowej (komórki piramidalne), a także wiązek doprowadzających (dośrodkowych) i odprowadzających... ... Świetna encyklopedia psychologiczna
Termin kora odnosi się do dowolnej zewnętrznej warstwy komórek mózgowych. W mózgu ssaków znajdują się trzy typy kory: kora gruszkowata, która pełni funkcje węchowe; stara kora (archicortex), która stanowi główną. Część… … Encyklopedia psychologiczna
Ze względu na pochodzenie kora mózgowa dzieli się na starożytną (pleocortex), starą (archecortex) i nową (neocortex). Starożytna kora obejmuje struktury związane z analizą bodźców węchowych i obejmuje opuszki węchowe, przewody i guzki. Stara kora obejmuje korę obręczy, korę hipokampa, zakręt zębaty i ciało migdałowate. Starożytna i stara kora tworzy mózg węchowy. Oprócz węchu mózg węchowy zapewnia reakcje czujności i uwagi, bierze udział w regulacji funkcji autonomicznych, odgrywa rolę w kształtowaniu zachowań seksualnych, jedzenia, instynktownych zachowań obronnych i dostarczaniu emocji.
Wszystkie pozostałe struktury korowe należą do kory nowej, która zajmuje około 96% Całkowita powierzchnia całą korę.
Położenie komórek nerwowych w korze określa się terminem „cytoarchitektura”. A włókna przewodzące nazywane są „mieloarchitekturą”.
Kora nowa składa się z 6 warstw komórek, które różnią się składem komórek, połączeniami nerwowymi i funkcjami. W obszarach starożytnej kory i starej kory wykrywa się tylko 2-3 warstwy komórek. Neurony w czterech górnych warstwach kory nowej przetwarzają przede wszystkim informacje z innych obszarów system nerwowy. Główną warstwą odśrodkową jest warstwa 5. Aksony jej komórek tworzą główne zstępujące ścieżki kory mózgowej, przewodzą sygnały kontrolujące funkcjonowanie struktur pnia i rdzeń kręgowy.
Warstwa 1 jest najbardziej zewnętrzną warstwą molekularną. Zawiera głównie włókna nerwowe z głębszych neuronów. Ponadto nie zawiera duża liczba małe komórki. Włókna warstwy molekularnej tworzą połączenia między różnymi obszarami kory
II warstwa – zewnętrzna granulowana. Zawiera dużą liczbę małych neuronów wielobiegunowych. Część wstępujących dendrytów z trzeciej warstwy kończy się w tej warstwie.
Warstwa 3 - zewnętrzna piramida. Jest najszerszy, zawiera głównie średnie, rzadziej małe i duże neurony piramidowe. Dendryty neuronów z tej warstwy kierowane są do warstwy drugiej.
4. warstwa - wewnętrzna granulowana. Zawiera duża liczba małe ziarniste, a także średnie i duże komórki gwiaździste. Podzielone są na dwie podwarstwy: 4a i 4b.
Warstwa 5 - zwojowa lub wewnętrzna piramida. Charakteryzuje się obecnością dużych neuronów piramidalnych. Ich skierowane ku górze dendryty docierają do warstwy molekularnej, a aksony podstawne i boczne są rozmieszczone w piątej warstwie.
Warstwa 6 - polimorficzna. Zawiera, wraz z komórkami innych form, neurony wrzecionowate. Kształty innych komórek są bardzo zróżnicowane: mają kształt trójkątny, piramidalny, owalny i wielokątny.
Kora - najwyższy wydział ośrodkowego układu nerwowego, zapewniający funkcjonowanie organizmu jako całości podczas jego interakcji z otoczeniem.
mózg (kora mózgowa, kora nowa) to warstwa istoty szarej, składająca się z 10-20 miliardów i pokrywająca półkule mózgowe (ryc. 1). Istota szara kory stanowi ponad połowę całkowitej istoty szarej ośrodkowego układu nerwowego. Całkowita powierzchnia istoty szarej kory wynosi około 0,2 m2, co osiąga się poprzez kręte pofałdowanie jej powierzchni i obecność rowków o różnej głębokości. Grubość kory w różnych jej częściach waha się od 1,3 do 4,5 mm (w przednim zakręcie środkowym). Neurony kory znajdują się w sześciu warstwach zorientowanych równolegle do jej powierzchni.
W obszarach kory należących do, znajdują się strefy o trójwarstwowym i pięciowarstwowym układzie neuronów w strukturze istoty szarej. Te obszary filogenetycznie starożytnej kory zajmują około 10% powierzchni półkul mózgowych, pozostałe 90% stanowi nową korę.

Ryż. 1. Kret powierzchni bocznej kory mózgowej (wg Brodmanna)
Struktura kory mózgowej
Kora mózgowa ma strukturę sześciowarstwową
Neurony różnych warstw różnią się cechami cytologicznymi i właściwościami funkcjonalnymi.
Warstwa molekularna- najbardziej powierzchowny. Jest reprezentowany przez niewielką liczbę neuronów i liczne rozgałęzione dendryty neuronów piramidalnych leżących w głębszych warstwach.
Zewnętrzna warstwa ziarnista utworzone przez gęsto rozmieszczone liczne małe neurony o różnych kształtach. Procesy komórek tej warstwy tworzą połączenia korowo-korowe.
Zewnętrzna warstwa piramidalna składa się ze średniej wielkości neuronów piramidalnych, których procesy biorą również udział w tworzeniu połączeń korowo-korowych między sąsiednimi obszarami kory.
Wewnętrzna warstwa ziarnista podobny do drugiej warstwy wyglądem komórek i układem włókien. Przez warstwę przechodzą wiązki włókien, łącząc różne obszary kory.
Neurony tej warstwy przenoszą sygnały z określonych jąder wzgórza. Warstwa jest bardzo dobrze reprezentowana w obszarach czuciowych kory.
Wewnętrzne warstwy piramidalne utworzone przez średnie i duże neurony piramidowe. W korze ruchowej neurony te są szczególnie duże (50–100 μm) i nazywane są gigantycznymi komórkami piramidalnymi Betza. Aksony tych komórek tworzą szybko przewodzące (do 120 m/s) włókna układu piramidowego.
Warstwa komórek polimorficznych reprezentowane głównie przez komórki, których aksony tworzą drogi korowo-wzgórzowe.

Neurony drugiej i czwartej warstwy kory biorą udział w percepcji i przetwarzaniu sygnałów odbieranych przez nie z neuronów w obszarach asocjacyjnych kory. Sygnały czuciowe z jąder przełączających wzgórza docierają głównie do neuronów czwartej warstwy, których ekspresja jest największa w pierwotnych obszarach czuciowych kory. Neurony pierwszej i innych warstw kory otrzymują sygnały z innych jąder wzgórza, zwojów podstawy i pnia mózgu. Neurony warstwy 3., 5. i 6. tworzą sygnały eferentne wysyłane do innych obszarów kory i wzdłuż zstępujących ścieżek do leżących poniżej części centralnego układu nerwowego. W szczególności neurony szóstej warstwy tworzą włókna, które docierają do wzgórza.
Istnieją znaczne różnice w składzie neuronów i cechach cytologicznych różnych obszarów kory. Na podstawie tych różnic Brodmann podzielił korę na 53 pola cytoarchitektoniczne (patrz ryc. 1).
Położenie wielu z tych zer, zidentyfikowane na podstawie danych histologicznych, pokrywa się w topografii z lokalizacją ośrodków korowych, identyfikowaną na podstawie pełnionych przez nie funkcji. Stosuje się także inne podejścia do podziału kory na regiony, na przykład w oparciu o zawartość określonych markerów w neuronach, zgodnie z naturą aktywności neuronowej i innymi kryteriami.
Istota biała półkul mózgowych zbudowana jest z włókien nerwowych. Atrakcja włókna asocjacyjne, podzielony na włókna łukowate, ale przez które sygnały są przekazywane między neuronami sąsiednich zakrętów i długimi podłużnymi wiązkami włókien, które dostarczają sygnały do neuronów w bardziej odległych częściach półkuli o tej samej nazwie.
Włókna spoidłowe - włókna poprzeczne, które przekazują sygnały między neuronami lewej i prawej półkuli.
Włókna projekcyjne - przewodzą sygnały pomiędzy neuronami kory mózgowej i innymi częściami mózgu.
Wymienione rodzaje włókien biorą udział w tworzeniu obwodów i sieci neuronowych, których neurony znajdują się w znacznych odległościach od siebie. Kora ma również specjalny typ lokalnych obwodów nerwowych utworzonych przez pobliskie neurony. Te struktury nerwowe nazywane są funkcjonalnymi kolumny korowe. Kolumny neuronalne tworzą grupy neuronów umieszczonych jeden nad drugim, prostopadle do powierzchni kory. Przynależność neuronów do tej samej kolumny można określić na podstawie wzrostu ich aktywności elektrycznej po stymulacji tego samego pola recepcyjnego. Aktywność taką rejestrujemy poprzez powolne przesuwanie elektrody rejestrującej w korze mózgowej w kierunku prostopadłym. Rejestrując aktywność elektryczną neuronów znajdujących się w płaszczyźnie poziomej kory, zauważamy wzrost ich aktywności po pobudzeniu różnych pól recepcyjnych.
Średnica kolumny funkcjonalnej wynosi do 1 mm. Neurony tej samej kolumny funkcjonalnej odbierają sygnały z tego samego doprowadzającego włókna wzgórzowo-korowego. Neurony sąsiednich kolumn są połączone ze sobą procesami, za pomocą których wymieniają informacje. Obecność takich wzajemnie połączonych kolumn funkcjonalnych w korze zwiększa niezawodność percepcji i analizy informacji docierających do kory.
Zapewniona jest także sprawność percepcji, przetwarzania i wykorzystania informacji przez korę mózgową do regulowania procesów fizjologicznych somatotopowa zasada organizacji pola czuciowe i motoryczne kory. Istotą tej organizacji jest to, że w pewnym (projekcyjnym) obszarze kory nie są byle jakie, ale topograficznie zarysowane obszary pola recepcyjnego powierzchni ciała, mięśni, stawów lub narządy wewnętrzne. Na przykład w korze somatosensorycznej powierzchnia ludzkiego ciała jest rzutowana w formie diagramu, gdy pola recepcyjne określonego obszaru powierzchni ciała są reprezentowane w pewnym punkcie kory. W sposób ściśle topograficzny pierwotna kora ruchowa zawiera neurony odprowadzające, których aktywacja powoduje skurcz niektórych mięśni ciała.
Scharakteryzowano także pola korowe zasada działania ekranu. W tym przypadku neuron receptorowy wysyła sygnał nie do pojedynczego neuronu lub do pojedynczego punktu ośrodka korowego, ale do sieci lub zera neuronów połączonych procesami. Komórki funkcjonalne tego pola (ekranu) to kolumny neuronów.
Kora mózgowa, powstająca na późniejszych etapach rozwoju ewolucyjnego organizmów wyższych, w pewnym stopniu podporządkowała sobie wszystkie podstawowe części centralnego układu nerwowego i jest w stanie skorygować ich funkcje. Jednocześnie aktywność funkcjonalna kory mózgowej jest determinowana przez napływ do niej sygnałów z neuronów tworzenia siatkowego pnia mózgu oraz sygnałów z pól recepcyjnych układów sensorycznych organizmu.
Obszary funkcjonalne kory mózgowej
W oparciu o ich cechy funkcjonalne, korę dzieli się na obszary czuciowe, skojarzeniowe i motoryczne.
Obszary czuciowe (wrażliwe, projekcyjne) kory
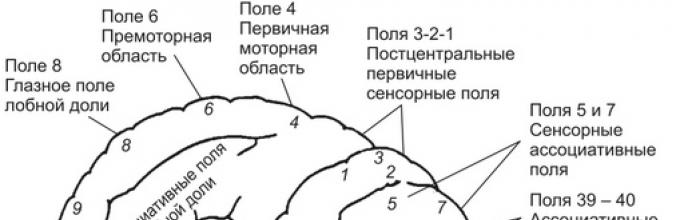
Składają się ze stref zawierających neurony, których aktywacja poprzez impulsy aferentne z receptorów czuciowych lub bezpośrednią ekspozycję na bodźce powoduje pojawienie się określonych wrażeń. Strefy te występują w obszarach kory potylicznej (pola 17-19), ciemieniowej (pola 1-3) i skroniowej (pola 21-22, 41-42).
W strefach czuciowych kory wyróżnia się centralne pola projekcyjne, zapewniające wyraźną, wyraźną percepcję wrażeń określonych modalności (światło, dźwięk, dotyk, ciepło, zimno) i wtórne pola projekcyjne. Zadaniem tego ostatniego jest zapewnienie zrozumienia związku między pierwotnym doznaniem a innymi obiektami i zjawiskami otaczającego świata.
Obszary reprezentacji pól recepcyjnych w obszarach czuciowych kory w dużym stopniu pokrywają się. Cechą ośrodków nerwowych w obszarze wtórnych pól projekcyjnych kory jest ich plastyczność, która objawia się możliwością restrukturyzacji specjalizacji i przywrócenia funkcji po uszkodzeniu któregokolwiek z ośrodków. Te zdolności kompensacyjne ośrodków nerwowych są szczególnie widoczne w dzieciństwo. Jednocześnie uszkodzeniu centralnych pól projekcyjnych po chorobie towarzyszy rażące upośledzenie funkcji sensorycznych i często niemożność ich przywrócenia.
Kora wzrokowa
Pierwotna kora wzrokowa (VI, obszar 17) znajduje się po obu stronach bruzdy kalkarynowej na przyśrodkowej powierzchni płata potylicznego mózgu. Zgodnie z identyfikacją naprzemiennych białych i ciemnych pasków w niezabarwionych obszarach kory wzrokowej, nazywana jest ona również korą prążkowaną. Neurony pierwotnej kory wzrokowej wysyłają sygnały wizualne z neuronów w bocznym ciele kolankowatym, które odbierają sygnały z komórek zwojowych siatkówki. Kora wzrokowa każdej półkuli odbiera sygnały wizualne z ipsilateralnej i przeciwnej połowy siatkówki obu oczu, a ich przybycie do neuronów korowych jest zorganizowane zgodnie z zasadą somatotopową. Neurony odbierające sygnały wzrokowe z fotoreceptorów są topograficznie zlokalizowane w korze wzrokowej, podobnie jak receptory w siatkówce. Co więcej, obszar plamki siatkówki ma stosunkowo większy obszar reprezentacji w korze niż inne obszary siatkówki.
Neurony pierwotnej kory wzrokowej odpowiadają za percepcję wzrokową, która na podstawie analizy sygnałów wejściowych objawia się ich zdolnością do wykrywania bodźca wzrokowego, określania jego specyficznego kształtu i orientacji w przestrzeni. W uproszczeniu możemy sobie wyobrazić funkcję sensoryczną kory wzrokowej w rozwiązywaniu problemu i odpowiadaniu na pytanie, czym jest obiekt wzrokowy.
W analizie innych cech sygnałów wzrokowych (np. lokalizacji w przestrzeni, ruchu, powiązań z innymi zdarzeniami itp.) biorą udział neurony pól 18 i 19 kory zewnątrzprążkowanej, zlokalizowane w sąsiedztwie zera 17. Informacje o sygnały otrzymane w czuciowych obszarach wzrokowych kory zostaną przekazane do dalszej analizy i wykorzystania wzroku do wykonywania innych funkcji mózgu w obszarach asocjacyjnych kory i innych części mózgu.
Kora słuchowa
Znajduje się w bruździe bocznej płat skroniowy w rejonie zakrętu Heschla (AI, pola 41-42). Neurony pierwotnej kory słuchowej odbierają sygnały z neuronów przyśrodkowych ciała kolankowate. Włókna przewodu słuchowego przenoszące sygnały dźwiękowe do kory słuchowej są zorganizowane tonotopowo, co umożliwia neuronom korowym odbieranie sygnałów od określonych komórek receptorów słuchowych w narządzie Cortiego. Kora słuchowa reguluje wrażliwość komórek słuchowych.
W pierwotnej korze słuchowej powstają wrażenia dźwiękowe i analizowane są indywidualne cechy dźwięków, aby odpowiedzieć na pytanie, czym jest postrzegany dźwięk. Pierwotna kora słuchowa odgrywa ważną rolę w analizie krótkich dźwięków i przerw pomiędzy nimi sygnały dźwiękowe, rytm, sekwencja dźwiękowa. Bardziej złożoną analizę dźwięków przeprowadza się w obszarach asocjacyjnych kory sąsiadujących z pierwotną korą słuchową. Na podstawie interakcji neuronów w tych obszarach kory przeprowadza się słyszenie obuuszne, określa się charakterystykę wysokości, barwy, głośności dźwięku i tożsamości dźwięku oraz określa się koncepcję trójwymiarowej przestrzeni dźwiękowej uformowany.
Kora przedsionkowa
Znajduje się w górnym i środkowym zakręcie skroniowym (obszary 21-22). Jego neurony odbierają sygnały od neuronów jąder przedsionkowych pnia mózgu, połączonych połączeniami doprowadzającymi z receptorami kanałów półkolistych aparatu przedsionkowego. Kora przedsionkowa tworzy poczucie położenia ciała w przestrzeni i przyspieszenia ruchów. Kora przedsionkowa oddziałuje z móżdżkiem (poprzez drogę skroniowo-mostową) i bierze udział w regulacji równowagi ciała i dostosowywaniu postawy do wykonywania celowych ruchów. Na podstawie interakcji tego obszaru z obszarami somatosensorczymi i asocjacyjnymi kory mózgowej następuje świadomość diagramu ciała.
Kora węchowa
Znajduje się w obszarze górnej części płata skroniowego (uncus, zero 34, 28). Kora zawiera wiele jąder i należy do struktur układu limbicznego. Jego neurony są rozmieszczone w trzech warstwach i odbierają sygnały doprowadzające z komórek mitralnych opuszki węchowej, połączonych połączeniami doprowadzającymi z neuronami receptorów węchowych. W korze węchowej przeprowadzana jest pierwotna analiza jakościowa zapachów i powstaje subiektywne odczucie zapachu, jego intensywności i przynależności. Uszkodzenie kory prowadzi do osłabienia węchu lub do rozwoju anosmii - utraty węchu. Przy sztucznej stymulacji tego obszaru powstają wrażenia różnych zapachów, podobne do halucynacji.
Kora smakowa
Znajduje się w dolnej części zakrętu somatosensorycznego, bezpośrednio przed obszarem projekcji twarzy (pole 43). Jego neurony otrzymują sygnały doprowadzające z neuronów przekaźnikowych wzgórza, które są połączone z neuronami jądra pasma samotnego rdzenia przedłużonego. Neurony tego jądra odbierają sygnały bezpośrednio z neuronów czuciowych, które tworzą synapsy na komórkach kubków smakowych. W korze smakowej przeprowadza się pierwotną analizę walorów smakowych gorzkiego, słonego, kwaśnego, słodkiego i na podstawie ich sumowania powstaje subiektywne odczucie smaku, jego intensywności i przynależności.
Sygnały węchu i smaku docierają do neuronów przedniej części kory wyspowej, gdzie na podstawie ich integracji powstaje nowa, bardziej złożona jakość wrażeń, która determinuje nasz stosunek do źródeł węchu czy smaku (na przykład do jedzenia).
Kora somatosensoryczna
Zajmuje obszar zakrętu postcentralnego (SI, pola 1-3), w tym płatek paracentralny po przyśrodkowej stronie półkul (ryc. 9.14). Do obszaru somatosensorycznego docierają sygnały czuciowe z neuronów wzgórzowych, połączonych szlakami rdzeniowo-wzgórzowymi z receptorami skóry (dotyk, temperatura, wrażliwość na ból), proprioceptorami (wrzeciona mięśniowe, torebki stawowe, ścięgna) i interoreceptorami (narządy wewnętrzne).

Ryż. 9.14. Najważniejsze ośrodki i obszary kory mózgowej
Ze względu na przecięcie dróg doprowadzających sygnał z prawej strony ciała dociera do strefy somatosensorycznej lewej półkuli i odpowiednio do prawej półkuli - z lewej strony ciała. W tym czuciowym obszarze kory wszystkie części ciała są reprezentowane somatotopowo, ale najważniejsze strefy recepcyjne palców, warg, skóry twarzy, języka i krtani zajmują stosunkowo większe obszary niż występy takich powierzchni ciała jak plecy, przód tułowia i nogi.
Lokalizacja reprezentacji wrażliwości części ciała wzdłuż zakrętu pośrodkowego jest często nazywana „odwróconym homunkulusem”, ponieważ rzut głowy i szyi znajduje się w dolnej części zakrętu pośrodkowego, a rzut części ogonowej tułów i nogi znajdują się w górnej części. W tym przypadku wrażliwość nóg i stóp rzutowana jest na korę płatka przyśrodkowego środkowej powierzchni półkul. W obrębie pierwotnej kory somatosensorycznej występuje pewna specjalizacja neuronów. Przykładowo neurony pola 3 odbierają głównie sygnały z wrzecion mięśniowych i mechanoreceptorów skóry, pole 2 - z receptorów stawowych.
Kora zakrętu postcentralnego jest klasyfikowana jako pierwotny obszar somatosensoryczny (SI). Jego neurony wysyłają przetworzone sygnały do neuronów we wtórnej korze somatosensorycznej (SII). Znajduje się za zakrętem postcentralnym w korze ciemieniowej (obszary 5 i 7) i należy do kory asocjacyjnej. Neurony SII nie otrzymują bezpośrednich sygnałów doprowadzających z neuronów wzgórzowych. Są połączone z neuronami SI i neuronami innych obszarów kory mózgowej. Pozwala to na integralną ocenę sygnałów docierających do kory drogą rdzeniowo-wzgórzową z sygnałami pochodzącymi z innych układów sensorycznych (wzrokowych, słuchowych, przedsionkowych itp.). Najważniejszą funkcją tych pól kory ciemieniowej jest percepcja przestrzeni i przekształcanie sygnałów czuciowych na współrzędne motoryczne. W korze ciemieniowej powstaje chęć (intencja, chęć) wykonania czynności motorycznej, która jest podstawą do rozpoczęcia planowania w niej nadchodzącej aktywności ruchowej.

Integracja różnych sygnałów zmysłowych wiąże się z powstawaniem różnych wrażeń kierowanych do różnych części ciała. Wrażenia te służą do generowania reakcji zarówno psychicznych, jak i innych, czego przykładem mogą być ruchy wymagające jednoczesnego udziału mięśni obu stron ciała (na przykład poruszanie się, czucie obiema rękami, chwytanie, ruch jednokierunkowy obiema rękami). Funkcjonowanie tego obszaru jest niezbędne do rozpoznawania obiektów poprzez dotyk i określania przestrzennego położenia tych obiektów.
Normalną funkcją somatosensorycznych obszarów korowych jest ważny warunek powstawanie wrażeń takich jak ciepło, zimno, ból i ich adresowanie do konkretnej części ciała.
Uszkodzenie neuronów w obszarze pierwotnej kory somatosensorycznej prowadzi do zmniejszenia różnego rodzaju wrażliwości po przeciwnej stronie ciała, a uszkodzenie miejscowe prowadzi do utraty wrażliwości w określonej części ciała. Szczególnie narażona na uszkodzenie neuronów pierwotnej kory somatosensorycznej jest wrażliwość rozróżniająca skóry, a najmniej wrażliwa jest ból. Uszkodzeniu neuronów wtórnej kory somatosensorycznej może towarzyszyć upośledzenie zdolności rozpoznawania obiektów za pomocą dotyku (agnozja dotykowa) i umiejętności posługiwania się przedmiotami (apraksja).
Obszary kory ruchowej
Około 130 lat temu badacze zastosowali punktową stymulację kory mózgowej wstrząs elektryczny odkryli, że ekspozycja na powierzchnię przedniego zakrętu centralnego powoduje skurcz mięśni po przeciwnej stronie ciała. W ten sposób odkryto obecność jednego z obszarów motorycznych kory mózgowej. Następnie okazało się, że kilka obszarów kory mózgowej i jej innych struktur jest związanych z organizacją ruchów, a w obszarach kory ruchowej znajdują się nie tylko neurony ruchowe, ale także neurony pełniące inne funkcje.
Pierwotna kora ruchowaPierwotna kora ruchowa zlokalizowany w przednim zakręcie centralnym (MI, pole 4). Jego neurony otrzymują główne sygnały doprowadzające z neuronów kory somatosensorycznej - obszarów 1, 2, 5, kory przedruchowej i wzgórza. Ponadto neurony móżdżku wysyłają sygnały do zawału mięśnia sercowego poprzez wzgórze brzuszno-boczne.
Włókna odprowadzające przewodu piramidowego zaczynają się od neuronów piramidalnych Ml. Część włókien tej ścieżki biegnie do neuronów ruchowych jąder nerwów czaszkowych pnia mózgu (droga korowo-opuszkowa), część do neuronów jąder motorycznych pnia (jądro czerwone, jądra formacji siatkowej, związane z jądrami pnia z móżdżkiem) i część do neuronów między- i ruchowych rdzenia kręgowego, mózg (droga korowo-rdzeniowa).
W zawale serca istnieje somatotopowa organizacja lokalizacji neuronów, które kontrolują skurcze różnych grup mięśni ciała. Neurony kontrolujące mięśnie nóg i tułowia znajdują się w górnych partiach zakrętu i zajmują stosunkowo niewielką powierzchnię, natomiast neurony kontrolujące mięśnie rąk, zwłaszcza palców, twarzy, języka i gardła znajdują się w dolnych partiach i zajmują dużą powierzchnię. Tak więc w pierwotnej korze ruchowej stosunkowo duży obszar zajmują grupy nerwowe, które kontrolują mięśnie wykonujące różne, precyzyjne, małe, precyzyjnie regulowane ruchy.
Ponieważ wiele neuronów Ml zwiększa aktywność elektryczną bezpośrednio przed wystąpieniem dobrowolnych skurczów, pierwotna kora ruchowa odgrywa wiodącą rolę w kontrolowaniu aktywności jąder ruchowych pnia mózgu i neuronów ruchowych rdzenia kręgowego oraz inicjowaniu dobrowolnych, ukierunkowanych na cel ruchów. Uszkodzenie pola Ml prowadzi do niedowładu mięśni i niemożności wykonywania drobnych, dowolnych ruchów.
Wtórna kora ruchowaObejmuje obszary kory przedruchowej i dodatkowej kory ruchowej (MII, pole 6). Kora przedruchowa znajduje się w obszarze 6, na bocznej powierzchni mózgu, przed pierwotną korą ruchową. Jego neurony odbierają sygnały doprowadzające przez wzgórze z obszarów potylicznych, somatosensorycznych, ciemieniowych, przedczołowych kory i móżdżku. Przetwarzane w nim neurony korowe wysyłają sygnały wzdłuż włókien odprowadzających do kory ruchowej MI, niewielka liczba do rdzenia kręgowego, a większa liczba do jąder czerwonych, jąder formacji siatkowej, zwojów podstawy i móżdżku. Kora przedruchowa odgrywa główną rolę w programowaniu i organizowaniu ruchów pod kontrolą wzrokową. Kora bierze udział w organizowaniu postawy i ruchach wspierających czynności wykonywane przez mięśnie dystalne kończyn. Uszkodzenie kory wzrokowej często powoduje tendencję do powtarzania rozpoczętego ruchu (persewerację), nawet jeśli ruch osiągnął cel.
W dolnej części kory przedruchowej lewego płata czołowego, bezpośrednio przed obszarem pierwotnej kory ruchowej, która zawiera neurony kontrolujące mięśnie twarzy, obszar mowy, Lub Ośrodek mowy ruchowej Broki. Naruszeniu jego funkcji towarzyszy upośledzenie artykulacji mowy lub afazja ruchowa.
Dodatkowa kora ruchowa znajduje się w górnej części obszaru 6. Jego neurony odbierają sygnały doprowadzające z obszarów somatosensorycznych, ciemieniowych i przedczołowych kory mózgowej. Sygnały przetwarzane przez neurony korowe są wysyłane włóknami eferentnymi do pierwotnej kory ruchowej, rdzenia kręgowego i jąder motorycznych pnia. Aktywność neuronów w dodatkowej korze ruchowej wzrasta wcześniej niż neuronów w korze zawału mięśnia sercowego, głównie w związku z realizacją ruchów złożonych. Jednocześnie wzrost aktywności nerwowej w dodatkowej korze ruchowej nie jest powiązany z ruchami jako takimi, w tym celu wystarczy mentalnie wyobrazić sobie model nadchodzących złożonych ruchów. Dodatkowa kora ruchowa bierze udział w tworzeniu programu nadchodzących złożonych ruchów oraz w organizacji reakcji motorycznych na specyfikę bodźców zmysłowych.
Ponieważ neurony wtórnej kory ruchowej wysyłają wiele aksonów do pola MI, uważa się, że jest to wyższa struktura w hierarchii ośrodków motorycznych organizujących ruchy, stojąca nad ośrodkami motorycznymi kory ruchowej MI. Ośrodki nerwowe wtórnej kory ruchowej mogą wpływać na aktywność neuronów ruchowych rdzenia kręgowego na dwa sposoby: bezpośrednio przez drogę korowo-rdzeniową i poprzez pole MI. Dlatego czasami nazywane są polami nadruchowymi, których funkcją jest instruowanie ośrodków pola MI.
Z obserwacji klinicznych wiadomo, że utrzymanie prawidłowej funkcji wtórnej kory ruchowej jest istotne dla wykonywania precyzyjnych ruchów ręki, a zwłaszcza wykonywania ruchów rytmicznych. Jeśli np. zostaną uszkodzone, pianista przestaje czuć rytm i utrzymywać interwał. Upośledzona jest zdolność wykonywania przeciwnych ruchów rękami (manipulacja obiema rękami).
Przy równoczesnym uszkodzeniu obszarów motorycznych MI i MII kory, utrata zdolności do wykonywania precyzyjnych skoordynowanych ruchów. Podrażnieniom punktowym w tych obszarach strefy ruchowej towarzyszy aktywacja nie poszczególnych mięśni, ale całej grupy mięśni, które powodują ukierunkowany ruch w stawach. Obserwacje te doprowadziły do wniosku, że kora ruchowa reprezentuje nie tyle mięśnie, ile ruchy.
Kora przedczołowaZnajduje się w obszarze pola 8. Jego neurony odbierają główne sygnały doprowadzające z potylicznej kory wzrokowej, ciemieniowej kory asocjacyjnej i wzgórków górnych. Przetworzone sygnały są przesyłane włóknami eferentnymi do kory przedruchowej, wzgórka górnego i ośrodków motorycznych pnia mózgu. Kora odgrywa decydującą rolę w organizowaniu ruchów pod kontrolą wzroku i jest bezpośrednio zaangażowana w inicjowanie i kontrolowanie ruchów oczu i głowy.
Mechanizmy realizujące przekształcenie planu ruchu w konkretny program motoryczny, w salwy impulsów wysyłanych do określonych grup mięśni, pozostają niedostatecznie poznane. Uważa się, że zamiar ruchu powstaje w wyniku funkcji asocjacyjnych i innych obszarów kory, oddziałując z wieloma strukturami mózgu.
Informacja o zamiarze ruchu przekazywana jest do obszarów motorycznych kory czołowej. Kora ruchowa poprzez zstępujące ścieżki aktywuje układy zapewniające rozwój i wykorzystanie nowych programów motorycznych lub wykorzystanie starych, już wyćwiczonych i zapisanych w pamięci. Integralna część Układy te to zwoje podstawy i móżdżek (patrz ich funkcje powyżej). Programy ruchowe opracowane przy udziale móżdżku i zwojów podstawy mózgu przekazywane są poprzez wzgórze do obszarów motorycznych, a przede wszystkim do pierwotnego obszaru motorycznego kory. Obszar ten bezpośrednio inicjuje wykonanie ruchów, łącząc z nim określone mięśnie i zapewniając sekwencję ich skurczu i rozluźnienia. Polecenia z kory mózgowej przekazywane są do ośrodków motorycznych pnia mózgu, neuronów ruchowych rdzenia kręgowego i neuronów ruchowych jąder nerwów czaszkowych. Podczas wykonywania ruchów neurony ruchowe pełnią rolę ostatniej ścieżki, przez którą polecenia motoryczne przekazywane są bezpośrednio do mięśni. Cechy przekazywania sygnału z kory mózgowej do ośrodków motorycznych pnia mózgu i rdzenia kręgowego opisano w rozdziale poświęconym ośrodkowemu układowi nerwowemu (pień mózgu, rdzeń kręgowy).
Stowarzyszenie obszary korowe
U ludzi obszary asocjacyjne kory zajmują około 50% powierzchni całej kory mózgowej. Znajdują się one w obszarach pomiędzy obszarami czuciowymi i motorycznymi kory. Obszary asocjacyjne nie mają wyraźnych granic z wtórnymi obszarami sensorycznymi, zarówno pod względem morfologicznym, jak i cechy funkcjonalne. W korze mózgowej znajdują się ciemieniowe, skroniowe i czołowe obszary asocjacji.
Kora skojarzeniowa ciemieniowa. Znajduje się w polach 5 i 7 górnego i dolnego płata ciemieniowego mózgu. Region ten jest ograniczony z przodu przez korę somatosensoryczną, a z tyłu przez korę wzrokową i słuchową. Sygnały wizualne, dźwiękowe, dotykowe, proprioceptywne, bólowe z aparatu pamięci i inne sygnały mogą dotrzeć i aktywować neurony ciemieniowego obszaru skojarzeniowego. Niektóre neurony są wielozmysłowe i mogą zwiększyć swoją aktywność, gdy dotrą do nich sygnały somatosensoryczne i wizualne. Jednakże stopień wzrostu aktywności neuronów w korze skojarzeniowej na odbiór sygnałów aferentnych zależy od aktualnej motywacji, uwagi osoby badanej oraz informacji odzyskanych z pamięci. Pozostaje nieistotne, jeśli sygnał dochodzący z obszarów sensorycznych mózgu jest dla podmiotu obojętny i znacznie wzrasta, jeśli pokrywa się z istniejącą motywacją i przyciąga jego uwagę. Na przykład, gdy małpie podano banana, aktywność neuronów w skojarzeniowej korze ciemieniowej pozostaje niska, jeśli zwierzę jest pełne, i odwrotnie, aktywność gwałtownie wzrasta u głodnych zwierząt, które lubią banany.
Neurony kory skojarzeniowej ciemieniowej są połączone połączeniami odprowadzającymi z neuronami obszarów przedczołowych, przedmotorycznych, motorycznych płata czołowego i zakrętu obręczy. Na podstawie obserwacji eksperymentalnych i klinicznych powszechnie przyjmuje się, że jedną z funkcji obszaru 5 kory jest wykorzystanie informacji somatosensorycznych do wykonywania celowych, dobrowolnych ruchów i manipulowania przedmiotami. Funkcją kory obszaru 7 jest integracja sygnałów wzrokowych i somatosensorycznych w celu koordynowania ruchów oczu i ruchów rąk sterowanych wzrokowo.
Naruszenie tych funkcji kory skojarzeniowej ciemieniowej w przypadku uszkodzenia jej połączeń z korą płata czołowego lub choroby samego płata czołowego wyjaśnia objawy następstw chorób zlokalizowanych w obszarze kory skojarzeniowej ciemieniowej. Mogą objawiać się trudnościami w zrozumieniu treści semantycznej sygnałów (agnozja), czego przykładem może być utrata umiejętności rozpoznawania kształtu i położenia przestrzennego obiektu. Procesy przekształcania sygnałów zmysłowych w odpowiednie działania motoryczne mogą zostać zakłócone. W tym drugim przypadku pacjent traci umiejętności praktyczne użycie znanych narzędzi i przedmiotów (apraksja) i może rozwinąć się niezdolność do wykonywania ruchów sterowanych wzrokowo (na przykład przesuwania ręki w stronę obiektu).
Przednia kora asocjacyjna. Znajduje się w korze przedczołowej, która jest częścią kory płata czołowego, położonej przed polami 6 i 8. Neurony czołowej kory skojarzeniowej odbierają przetworzone sygnały czuciowe poprzez połączenia aferentne z neuronów korowych w płatach potylicznym, ciemieniowym i skroniowym mózgu i neuronów w zakręcie obręczy. Przednia kora asocjacyjna odbiera sygnały o aktualnych stanach motywacyjnych i emocjonalnych z jąder wzgórza, struktur limbicznych i innych struktur mózgowych. Ponadto kora czołowa może operować abstrakcyjnymi, wirtualnymi sygnałami. Asocjacyjna kora czołowa wysyła sygnały eferentne z powrotem do struktur mózgu, z których zostały odebrane, do obszarów motorycznych kory czołowej, jądra ogoniastego zwojów podstawy i podwzgórza.
Ten obszar kory odgrywa podstawową rolę w kształtowaniu wyższych funkcji umysłowych człowieka. Zapewnia kształtowanie docelowych ustawień i programów świadomych reakcji behawioralnych, rozpoznawania i oceny semantycznej obiektów i zjawisk, rozumienia mowy, logiczne myślenie. Po rozległym uszkodzeniu kory czołowej u pacjentów może rozwinąć się apatia, obniżone tło emocjonalne, krytyczne podejście do działań własnych i innych, samozadowolenie i upośledzona zdolność wykorzystania przeszłych doświadczeń do zmiany zachowania. Zachowanie pacjentów może stać się nieprzewidywalne i niewłaściwe.
Kora skojarzeń czasowych. Znajduje się w polach 20, 21, 22. Neurony korowe odbierają sygnały czuciowe z neuronów kory słuchowej, pozaprążkowanej kory wzrokowej i przedczołowej, hipokampa i ciała migdałowatego.
Po obustronnej chorobie skroniowych obszarów skojarzonych obejmujących hipokamp lub połączenia z nim w procesie patologicznym, u pacjentów może rozwinąć się poważne upośledzenie pamięci, zachowania emocjonalne i niezdolność do koncentracji uwagi (roztargnienie). U niektórych osób, jeśli uszkodzony zostanie obszar dolno-skroniowy, w którym rzekomo znajduje się ośrodek rozpoznawania twarzy, może rozwinąć się agnozja wzrokowa – niemożność rozpoznania twarzy znajomych osób lub obiektów przy jednoczesnym zachowaniu wzroku.
Na granicy obszarów skroniowych, wzrokowych i ciemieniowych kory w dolnej ciemieniowej i tylnej części płata skroniowego znajduje się obszar asocjacyjny kory, zwany zmysłowy ośrodek mowy, czyli ośrodek Wernickego. Po jego uszkodzeniu rozwija się dysfunkcja rozumienia mowy, przy zachowaniu funkcji motorycznych mowy.
Kora nowa - ewolucyjnie najmłodsza część kory, zajmująca większość powierzchni półkul. Jego grubość u ludzi wynosi około 3 mm.
Skład komórkowy kory nowej jest bardzo zróżnicowany, ale około trzy czwarte neuronów korowych to neurony piramidalne (piramidy), dlatego jedna z głównych klasyfikacji neuronów korowych dzieli je na piramidalne i niepiramidalne (wrzecionowate, gwiaździste, ziarniste , ogniwa żyrandolowe, ogniwa Martinottiego itp.). Inna klasyfikacja jest związana z długością aksonu (patrz paragraf 2.4). Komórki Golgiego I o długich aksonach to głównie piramidy i wrzeciona, ich aksony mogą wychodzić z kory, pozostałe komórki to komórki Golgiego II o krótkich aksonach.
Neurony korowe różnią się także wielkością ciała komórki: rozmiar ultramałych neuronów wynosi 6x5 mikronów, rozmiar gigantycznych ponad 40 x 18. Największe neurony to piramidy Betza, ich rozmiar to 120 x 30-60 mikrony.
Neurony piramidalne (patrz ryc. 2.6, G) mają kształt ciała w kształcie piramidy, której wierzchołek jest skierowany do góry. Od tego wierzchołka rozciąga się dendryt wierzchołkowy i wznosi się do leżących nad nim warstw korowych. Dendryty podstawne wystają z pozostałych części somy. Wszystkie dendryty mają kolce. Od podstawy komórki odchodzi długi akson, tworząc liczne zabezpieczenia, w tym nawracające, które wyginają się i wznoszą do góry. Komórki gwiaździste nie mają wierzchołkowego dendrytu i w większości przypadków na dendrytach nie ma kolców. W komórkach wrzecionowatych dwa duże dendryty wystają z przeciwległych biegunów ciała, są też małe dendryty wystające z reszty ciała. Dendryty mają kolce. Akson jest długi i ma niewiele odgałęzień.
Podczas rozwój zarodkowy Nowa kora koniecznie przechodzi przez etap struktury sześciowarstwowej, podczas dojrzewania w niektórych obszarach liczba warstw może się zmniejszyć. Warstwy głębokie są filogenetycznie starsze, warstwy zewnętrzne są młodsze. Każda warstwa kory charakteryzuje się składem neuronalnym i grubością, które mogą różnić się od siebie w różnych obszarach kory.
Zróbmy listę warstwy neokory(ryc. 9.8).
Nakładam - molekularny- najbardziej zewnętrzny, zawiera niewielką liczbę neuronów i składa się głównie z włókien biegnących równolegle do powierzchni. Tutaj również wznoszą się dendryty neuronów znajdujących się w niższych warstwach.
II warstwa - zewnętrzny granulat, Lub zewnętrzny granulat, - składa się głównie z małych neuronów piramidalnych i niewielkiej liczby średnich komórek gwiaździstych.
III warstwa - zewnętrzna piramida - najszersza i najgrubsza warstwa, zawiera głównie małe i średnie neurony piramidalne i gwiaździste. W głębi warstwy znajdują się duże i gigantyczne piramidy.
Warstwa IV - wewnętrzny ziarnisty, Lub wewnętrzny ziarnisty, - składa się głównie z małych neuronów wszelkich odmian, jest też kilka dużych piramid.
Warstwa V - wewnętrzna piramida, Lub ganglionowy, cecha charakterystyczna czyli obecność dużych, a w niektórych obszarach (głównie w polach 4 i 6; ryc. 9.9; podrozdział 9.3.4) - gigantycznych neuronów piramidalnych (piramid Betza). Szczytowe dendryty piramid z reguły sięgają warstwy I.
VI warstwa - polimorficzny, Lub wielopostaciowy, - zawiera głównie neurony wrzecionowate, a także komórki wszystkich innych form. Warstwa ta jest podzielona na dwie podwarstwy, które wielu badaczy uważa za warstwy niezależne, mówiąc w tym przypadku o siedmiowarstwowej korze.
Ryż. 9,8.
A- neurony są całkowicie zabarwione; B- kolorowe są tylko ciała neuronów; V- malowane
tylko procesy neuronowe
Główne funkcje Każda warstwa jest również inna. Warstwy I i II realizują połączenia pomiędzy neuronami różnych warstw kory mózgowej. Włókna modzelowe i asocjacyjne pochodzą głównie z piramid warstwy III i docierają do warstwy II. Główne włókna doprowadzające wchodzące do kory ze wzgórza kończą się na neuronach warstwy IV. Warstwa V kojarzona jest głównie z układem zstępujących włókien projekcyjnych. Aksony piramid tej warstwy tworzą główne drogi odprowadzające kory mózgowej.
W większości pól korowych wszystkie sześć warstw jest równie dobrze wyrażonych. Ta kora nazywa się homotypowy. Jednakże w niektórych dziedzinach ekspresja warstw może ulec zmianie w trakcie wywoływania. Ten rodzaj kory nazywa się heterotypowy. Występuje w dwóch rodzajach:
ziarnisty (zera 3, 17, 41; ryc. 9.9), w którym znacznie zwiększa się liczba neuronów w warstwie ziarnistej zewnętrznej (II), a zwłaszcza wewnętrznej (IV), w wyniku czego warstwa IV dzieli się na trzy podwarstwy. Taka kora jest charakterystyczna dla pierwotnych obszarów sensorycznych (patrz poniżej);
Agranularny (pola 4 i 6, czyli kora ruchowa i przedruchowa; ryc. 9.9), w którym przeciwnie, występuje bardzo wąska warstwa II i praktycznie nie ma IV, ale bardzo szerokie warstwy piramidalne, zwłaszcza wewnętrzna (V) .







