Wszystkie glony różnią się znacznie zestawem pigmentów fotosyntetycznych. Takie grupy w taksonomii roślin mają status działów.
Głównym pigmentem wszystkich alg jest zielony pigment chlorofil. Znane są cztery rodzaje chlorofilu, różniące się budową: chlorofil a– występuje we wszystkich algach i roślinach wyższych; chlorofil b– występuje w algach zielonych, charofitach, euglenoidach i roślinach wyższych: rośliny zawierające ten chlorofil zawsze mają jasnozieloną barwę; chlorofil c– występuje w algach heterokontowych; chlorofil d– rzadka forma, występująca u alg czerwonych i niebieskozielonych. Większość roślin fotosyntetyzujących zawiera dwa różne chlorofile, z których jeden jest zawsze chlorofilem A. W niektórych przypadkach zamiast drugiego chlorofilu są biliproteiny. Istnieją dwa rodzaje biliprotein występujących w niebiesko-zielonych i czerwonych algach: fikocyjanina– niebieski pigment, fikoerytryna- czerwony pigment.
Obowiązkowymi pigmentami zawartymi w błonach fotosyntetycznych są pigmenty żółte - karotenoidy. Różnią się od chlorofilów widmem pochłanianego światła i uważa się, że pełnią funkcję ochronną, chroniąc cząsteczki chlorofilu przed destrukcyjnym działaniem tlenu cząsteczkowego.
Oprócz wymienionych pigmentów algi zawierają również: fukoksantyna– złoty pigment; ksantofil- brązowy pigment.
Koniec pracy -
Ten temat należy do działu:
Wodorost
Uniwersytet Rybacki.. Instytut Biologii Morza im. A. w Żyrmuńskim Dworze Ran.. l l Arbuzov..
Jeśli potrzebujesz dodatkowy materiał na ten temat lub nie znalazłeś tego, czego szukałeś, polecamy skorzystać z wyszukiwarki w naszej bazie dzieł:
Co zrobimy z otrzymanym materiałem:
Jeśli ten materiał był dla Ciebie przydatny, możesz zapisać go na swojej stronie w sieciach społecznościowych:
| Ćwierkać |
Wszystkie tematy w tym dziale:
Osłony komórek
Osłony komórek zapewniają odporność wewnętrznej zawartości komórek na wpływy zewnętrzne i nadają komórkom określony kształt. Osłony są przepuszczalne dla wody i małych cząsteczek w niej rozpuszczonych
Wici
Monadyczne komórki wegetatywne i monadyczne etapy cyklu życiowego (zoospory i gamety) glonów są wyposażone w wici - długie i dość grube wyrostki komórkowe, pokryte zewnętrznie plazmalemmą. I
Mitochondria
Mitochondria występują w komórkach alg eukariotycznych. Kształt i struktura mitochondriów w komórkach glonów jest bardziej zróżnicowana w porównaniu do mitochondriów roślin wyższych. Mogą być okrągłe
Plastydy
Pigmenty w komórkach alg eukariotycznych znajdują się w plastydach, jak we wszystkich roślinach. W algach występują dwa rodzaje plastydów: kolorowe chloroplasty (chromatofory) i bezbarwne leukoplasty (ami
Jądro i aparat mitotyczny
Jądro glonów ma strukturę typową dla eukariontów. Liczba jąder w komórce może wahać się od jednego do kilku. Na zewnątrz rdzeń pokryty jest powłoką składającą się z dwóch membran, czyli membrany zewnętrznej
Monadyczny (wiciowy) typ struktury plechy
Bardzo cecha charakterystyczna, który determinuje ten typ struktury, to obecność wici, za pomocą których organizmy monadyczne aktywnie poruszają się środowisko wodne(ryc. 9, A). Ruchome z
Struktura typu ryzopodialnego (ameboidalnego).
Najważniejszymi cechami struktury typu ameboidalnego jest brak silnych osłon komórkowych i zdolność do poruszania się ameboidalnego za pomocą qi tymczasowo utworzonego na powierzchni komórki
Struktura typu palmelloidowego (hemimonadalnego).
Charakterystyczne dla tego typu struktury jest połączenie stacjonarnego trybu życia roślin z obecnością organelli komórkowych charakterystycznych dla organizmów monadycznych: wakuoli kurczliwych, piętna, opaski uciskowej
Struktura typu kokosowego
Ten typ łączy glony jednokomórkowe i kolonialne, nieruchome w stanie wegetatywnym. Komórki typu kokosowego są pokryte błoną i mają protoplast typu roślinnego (tonoplast bez sokratesa
Struktura typu trichal (nitkowatego).
Charakterystyczną cechą struktury typu nitkowatego jest nitkowaty układ nieruchomych komórek, które powstają wegetatywnie w wyniku podziału komórek, który zachodzi głównie
Struktura typu heterotrychalnego (niewłóknistego).
Typ heterofilamentowy powstał na bazie typu nitkowatego. Plecha heterofilamentowa składa się głównie z nici poziomych pełzających po podłożu, pełniących funkcję mocowania, oraz nitek pionowych, wzdłuż
Struktura miąższowa (tkankowa).
Jeden z kierunków ewolucji plechy heterofilamentowej wiązał się z pojawieniem się plechy miąższowej. Zdolność do nieograniczonego wzrostu i podziału komórek w różnych kierunkach doprowadziła do ewolucji
Konstrukcja typu syfonowego
Struktura typu syfonalnego (niekomórkowego) charakteryzuje się brakiem komórek wewnątrz wzgórza, które osiąga stosunkowo duże, zwykle makroskopowe rozmiary i pewien stopień zróżnicowania
Konstrukcja typu syfonokladalnego
Główną cechą struktury typu syfonokladalnego jest zdolność do tworzenia kompleksowo ułożonych wzgórz, składających się głównie z segmentów wielojądrzastych, z pierwotnego wzgórza niekomórkowego. W
Rozmnażanie bezpłciowe
Rozmnażanie bezpłciowe glonów odbywa się za pomocą wyspecjalizowanych komórek - zarodników. Sporulacji zwykle towarzyszy podział protoplastu na części i uwolnienie z nich produktów rozszczepienia
Prosty podział
Ta metoda rozmnażania występuje tylko w jednokomórkowych formach glonów. Podział zachodzi najłatwiej w komórkach o budowie ciała typu ameboidalnego. Podział form ameboidalnych
Podział
Fragmentacja jest nieodłączna dla wszystkich grup glonów wielokomórkowych i objawia się w różnych postaciach: tworzeniu hormogoniów, regeneracji oderwanych części plechy, spontanicznej utraty gałęzi, odrastania
Rozmnażanie przez pędy, rozłogi, pąki czerwowe, guzki, akinety
W postaciach tkankowych zielonych, brunatnych i czerwonych alg rozmnażanie wegetatywne przybiera pełną formę, która niewiele różni się od rozmnażania wegetatywnego roślin wyższych. Trzymanie drogi
Rozmnażanie płciowe
Rozmnażanie płciowe u alg jest to związane z procesem płciowym, który polega na połączeniu dwóch komórek, w wyniku czego powstaje zygota, z której wyrasta nowy osobnik lub wytwarza zoospory.
Zmiana faz jądrowych
Podczas procesu seksualnego, w wyniku połączenia gamet i ich jąder, liczba chromosomów w jądrze podwaja się. Na pewnym etapie cyklu rozwojowego, podczas mejozy, następuje zmniejszenie liczby chromosomów
Endofity/endozoity lub endosymbionty
Endosymbionty, czyli symbionty wewnątrzkomórkowe, to glony żyjące w tkankach lub komórkach innych organizmów (bezkręgowców lub glonów). Tworzą rodzaj grupy ekologicznej
Dział sinic (cyjanobakterii) – cyanophyta
Nazwa działu (od greckiego cyanos – niebieski) odzwierciedla charakterystyczną cechę tych alg – kolor plechy, związany ze stosunkowo dużą zawartością niebieskiego barwnika – fikocyjaniny. Cyjan
Zamówienie – Chroococcales
Występują jako jednokomórkowe „proste” osobniki lub częściej tworzą kolonie śluzowe. Kiedy komórki dzielą się na dwie płaszczyzny, pojawiają się jednowarstwowe kolonie blaszkowate. Podział w trzech punktach
Dział krasnorostów – rhodophyta
Nazwa działu pochodzi od greckiego słowa rhodon („rodon”) – różowy. Kolor czerwonych alg wynika z różnych kombinacji pigmentów. Występuje w kolorze szarym i fioletowym
Zamawiaj Banguiaceae – Bangiales
Rodzaj Porphyra ma plechę w postaci cienkiej błyszczącej płytki o gładkich lub złożonych krawędziach, składającej się z jednej lub dwóch warstw ściśle połączonych komórek. Podstawa płyty zwykle wchodzi
Zamów Rhodymeniales
Rodzaj Sparlingia (Rodimenia) - płaskie płytki do 45 cm wysokości, w kształcie liścia i klina, rozszerzone i palmatelnie rozcięte u góry, od jasnoróżowego lub jasnopomarańczowego do
Zamów Coralline - Corallinales
Rodzaj Coralline to podzielony na segmenty, wachlarzowaty, rozgałęziony krzew do 10 cm wysokości, rozgałęziony, wapienny, od różowo-liliowego do prawie białego. Rozmnaża się bezpłciowo i płciowo. Spo
Zamów Gigartinales – Gigartinales
Rodzaj Hondrus - gęste, skórzaste krzewy chrzęstne do 20 cm wysokości, 3-4 razy rozgałęzione, jasnożółte, jasnoróżowe, fioletowo-ciemnoczerwone. Rośnie w dolnej części strefy przybrzeżnej i
Zamów Ceramiaceae – Ceramiales
Rodzaj Ceramium to delikatny, puszysty, segmentowany krzew dorastający do 10 cm wysokości, rozgałęziony dychotomicznie lub naprzemiennie, ciemnożółty z różowawym odcieniem. Oczywiście rozgałęzienie od dwóch do czterech rzędów
Podział okrzemek - Bacillariophyta
Dział nazywa się Okrzemki (od greckiego di - dwa, tom - cięcie, sekcja) lub Bacillaria (bacillum - kij). Obejmuje jednokomórkową organizację samotną lub kolonialną.
Podział glonów heterokontowych (heterokontophyta)
Wszystkie heterokonty mają podobną strukturę aparatu wiciowego. Są 2 wici, a jedna z nich ma bardzo charakterystyczne rurkowate, trójczłonowe pierzaste narośla, czyli włosy - mastigonemy. Dokładnie gotówką
Taksonomia
Kokolity kopalne znane są z osadów mezozoiku i występowały obficie przez większość okresów jurajskiego i kredowego. Prymnesiofity osiągnęły największą różnorodność w późnej kredzie,
Dział glonów kryptofitowych (kryptomonad) – cryptophyta
Nazwa działu pochodzi od rodzaju Cryptomonas (od greckiego kryptos – ukryty, monas – indywidualny). Obejmuje organizmy jednokomórkowe, ruchliwe i monadyczne. Komórki kryptofitów
A B C D E
Ryż. 53. Wygląd glony kryptofitowe (wg: G.A. Belyakova i in., 2006): A – Rodomonas, B – Chroomonas, C – Cryptomonas, D – Chilomonas, D – Goniomonas puszka
Dział zielonych alg – chlorophyta
Zielonki są najbardziej rozległym ze wszystkich działów numerycznych glonów różne szacunki od 4 do 13 - 20 tysięcy gatunków. Wszystkie mają zielony kolor thalli, co wynika z przewagi chloru
Zamów Ulothrixales – Ulotrichales
Rodzaj Ulothrix (ryc. 54). Gatunki Ulotrix częściej żyją w wodach słodkich, rzadziej w morzach, zbiornikach słonawych i w glebie. Przywiązują się do podwodnych obiektów, tworząc jasnozielone krzewy.
Zamów Bryopsidae – Bryopsidales
Większość gatunków występuje w wodach słodkich i słonawych. Część z nich rośnie na ziemi, na kamieniach, piasku, czasem na słonych bagnach. Rodzaj Bryopsis - krzewy nitkowate do 6-8 s
Zamów Volvocales - Volvocales
Rodzaj Chlamydomonas (ryc. 57) obejmuje ponad 500 gatunków jednokomórkowych glonów żyjących w świeżych, małych, dobrze ogrzanych i zanieczyszczonych zbiornikach wodnych: stawach, kałużach, rowach itp. Itp
Oddział Charophyta (Characeae) – Charophyta
Charofity to linia słodkowodnych zielonych alg, z których wywodzą się rośliny wyższe. Są to formy przeważnie z plechą nitkowatą. Często plecha jest pionowa, rozcięta i niesiona
Podział dinofitów (dinofagellates) – dinophyta
1. Nazwa katedry pochodzi z języka greckiego. dineo – obracać się. Łączy przeważnie jednokomórkowe monadyczne, rzadziej kokosowe, ameboidalne lub palmelloidowe, czasem kolonialne
Podział euglenozoa - euglenovae
Nazwa działu pochodzi od rodzaju typu - Euglena (od greckiego eu - dobrze rozwinięty, glene - źrenica, oko). Łączy pojedynczych przedstawicieli monadycznych lub ameboidalnych. Czasami się spotykamy
Słownik terminów
Autogamia to rozmnażanie płciowe, w którym dwa siostrzane jądra haploidalne łączą się we wspólnej cytoplazmie. Autospora to struktura rozmnażania bezpłciowego, która jest
Stan Techniczny Dalekiego Wschodu
uniwersytet rybacki
Instytut Biologii Morza im. AV Żyrmuński Dalekowschodni Oddział Rosyjskiej Akademii Nauk
LL. Arbuzowa
I.R. Levenets
Wodorost

Recenzenci:
– V.G. Chavtur, doktor nauk biologicznych, profesor Katedry Biologii Morza i Akwakultury Państwowego Uniwersytetu Dalekiego Wschodu
– dr S.V. Nesterova, starszy pracownik naukowy w Laboratorium Flory Dalekiego Wschodu, Instytut Ogrodu Botanicznego Dalekowschodniego Oddziału Rosyjskiej Akademii Nauk
Arbuzova L.L., Levenets I.R. Algi: badania. wieś Władywostok: Dalrybvtuz, IBM FEB RAS, 2010. 177 s.
Podręcznik dostarcza nowoczesnych informacji na temat anatomii, morfologii, taksonomii, stylu życia i praktycznego znaczenia glonów.
Podręcznik przeznaczony jest dla studentów studiów licencjackich na kierunkach „Biozasoby wodne i akwakultura” oraz „Ekologia i zarządzanie środowiskiem” stacjonarnych i niestacjonarnych, magistrów ekologii, biologii, ichtiologii i hodowli ryb.
©Państwo Dalekiego Wschodu
rybołówstwo techniczne
uniwersytet, 2010
©Instytut Biologii Morskiej im. AV Żyrmuński Dalekowschodni Oddział Rosyjskiej Akademii Nauk, 2010
ISBN ……………………..
Wstęp…………………………………………………………………………………
1. Struktura komórek glonów……………………………………
2. Ogólna charakterystyka alg……………………………
2.1. Rodzaje pożywienia………………………………………………………
2.2. Rodzaje thalli……………………………………………..
2.3. Rozmnażanie się glonów…………………………………..
2.4. Cykle życiowe glonów…………………………….
3. Grupy ekologiczne glonów…………………………….
3.1. Glony siedlisk wodnych…………………………..
3.1.1. Fitoplankton………………………………………………….
3.1.2. Fitobentos ………………………………………………..
3.1.3. Glony ekstremalnych ekosystemów wodnych............
3.2. Glony siedlisk niewodnych………………………
3.2.1. Algi aerofilne............................................................
3.2.2. Algi edafilne………………………………….
3.2.3. Algi litofilne............................................................
4. Rola glonów w przyrodzie i znaczenie praktyczne………
5. Współczesna taksonomia alg………………………..
5.1. Algi prokariotyczne……………………………..
5.1.1. Zakład Błękitnice………………………
5.2. Algi eukariotyczne............................................................
5.2.1. Dział Algi czerwone…………………………….
5.2.2. Podział okrzemek………………………..
5.2.3. Zakład Heterokont Algi…………………….
Klasa Algi brunatne ………………………………
Klasa Złociste algi …………………………
Klasa Algi Sinura …………………………..
Klasa Algi Pheotamnia………………………
Klasa Raphid algi ………………………….
Klasa glonów Eustigma …………………………
Klasa Algi żółto-zielone………………………
5.2.4. Zakład Glony Prymnesiophyte…………….
5.2.5. Zakład Algi kryptofitowe……………………
5.2.6. Dział Zielone algi………………….…………
5.2.7. Podział Characeae…………………………….
5.2.8. Dział Algi Dinophyte………………………
5.2.9. Sekcja Algi Euglena …………………………
Literatura………………………………………………………………………………
Słownik terminów……………………………………………………….
Aplikacja …………………………………………………………….
WSTĘP
Glony tradycyjnie obejmują zróżnicowaną grupę organizmów plechowych, fotosyntetycznych, zarodnikowych i jałowych. Podobnie jak wszystkie niższe rośliny, narządy rozrodcze glonów są pozbawione powłoki, ciało nie jest podzielone na narządy i nie ma tkanek. Wśród glonów występują zarówno formy eukariotyczne, jak i prokariotyczne. Te ostatnie w odróżnieniu od chlorobakterii podczas fotosyntezy uwalniają do środowiska wolny tlen.
Glony zajmują dominującą pozycję zarówno w zbiornikach wód słodkich, jak i morskich. Będąc głównymi producentami, w dużej mierze determinują produktywność ryb ekosystemów wodnych. Dzięki aktywności fotosyntetycznej algi wzbogacają wodę w tlen i redukują ilość dwutlenku węgla. Mają wyjątkową zdolność gromadzenia różnorodnych szkodliwe substancje, a także uwalniają do środowiska metabolity, które hamują rozwój mikroorganizmów chorobotwórczych. Algi zmieniając skład chemiczny wody często przyczyniają się do jej oczyszczenia. Skład jakościowy i ilościowy grup glonów jest ważnym wskaźnikiem stanu ekologicznego jednolitych części wód. Wiele gatunków wykorzystuje się jako wskaźniki zanieczyszczenia wody.
Badanie glonów jest ważnym etapem w kształceniu specjalistów z zakresu marikultury, hodowli ryb i ekologii morza. Znajomość budowy, ekologii i systematyki glonów jest podstawą badań higrobiologii, ichtiologii, ekologii, ichtiotoksykologii; są także niezbędne do oceny bazy surowcowej zbiorników i sporządzania prognoz połowowych.
Ostatnio, dzięki nowoczesnym technikom metodologicznym, uzyskano nowe informacje na temat drobnej struktury, fizjologii i biochemii glonów, co spowodowało rewizję tradycyjnych pomysłów. Największym zmianom uległa taksonomia roślin niższych, do których zaliczają się glony. Jednocześnie współczesne informacje na temat systematyki i struktury glonów nie znajdują odzwierciedlenia w literaturze edukacyjnej z zakresu botaniki, a specjalna literatura z zakresu fykologii nie jest dostępna dla szerokiego grona studentów.
W tym podręcznik Dostarczane są najświeższe informacje na temat budowy, morfologii, taksonomii, ekologii i praktycznego znaczenia glonów. Podano opis najważniejszych taksonów glonów.
Podręcznik przeznaczony jest dla studentów studiów licencjackich na kierunkach „Biozasoby wodne i akwakultura” oraz „Ekologia i zarządzanie środowiskiem” studiów stacjonarnych i niestacjonarnych, magisterskich na kierunkach ekologia, ichtiologia, hodowla ryb i akwakultura.
W przygotowaniu materiałów do niniejszego podręcznika wzięli udział nauczyciele z Dalrybvtuz oraz specjaliści z zakresu hydrobiologii i fykologii Dalekowschodniego Oddziału Rosyjskiej Akademii Nauk.
1. STRUKTURA KOMÓREK ALG
Glony prokariotyczne mają podobną strukturę komórkową do bakterii: brakuje im organelli błonowych, takich jak jądro, chloroplasty, mitochondria, siateczka śródplazmatyczna i aparat Golgiego.
Glony eukariotyczne zawierają elementy strukturalne charakterystyczne dla wyższych komórek roślinnych (ryc. 1).

Ryż. 1. Dorosła komórka roślinna bez wtórnego pogrubienia ścianki (schemat) przy maksymalnym powiększeniu mikroskopu świetlnego (po: 1 – ściana komórkowa, 2 – płytka środkowa, 3 – przestrzeń międzykomórkowa, 4 – plazmodesmata, 5 – plazmalema, 6 – tonoplast, 7 – wakuola centralna, 9 – jądro, 10 – otoczka jądrowa, 11 – por w otoczce jądrowej, 12 – jąderko, 13 – chromatyna, 14 – chloroplast, 15 – grana w chloroplastie, 16 – ziarno skrobi w chloroplastie, 17 - mitochondrium, 18 - dyktosom, 19 - ziarnista siateczka śródplazmatyczna, 20 - kropla rezerwowego tłuszczu (lipidu) w cytoplazmie, 21 - mikrociało, 22 - cytoplazma (hialoplazma)
Podany schemat komórka roślinna ogólnie odzwierciedla budowę komórek glonów, jednakże wiele glonów wraz z typowymi organellami roślinnymi (plastydami, wakuolami z sokiem komórkowym) zawiera struktury charakterystyczne dla komórek zwierzęcych (wici, piętno, nietypowe dla komórek roślinnych błony).
Osłony komórek
Osłony komórek zapewniają odporność wewnętrznej zawartości komórek na wpływy zewnętrzne i nadają komórkom określony kształt. Osłony są przepuszczalne dla wody i rozpuszczonych w niej substancji drobnocząsteczkowych oraz łatwo przepuszczają światło słoneczne. Osłony komórkowe glonów charakteryzują się dużą różnorodnością morfologiczną i chemiczną. Należą do nich polisacharydy, białka, glikoproteiny, sole mineralne, pigmenty, lipidy i woda. W przeciwieństwie do roślin wyższych, w skorupach alg nie ma ligniny.
Struktura osłon komórkowych opiera się na plazmalemie, czyli błonie cytoplazmatycznej. U wielu przedstawicieli wici i ameboidów komórki na zewnątrz pokryte są jedynie plazmalemmą, która nie jest w stanie zapewnić stałego kształtu ciała. Takie komórki mogą tworzyć pseudopodia. Na podstawie morfologii wyróżnia się kilka typów pseudopodiów. Najczęściej spotykany w algach ryzopodia, które są nitkowatymi, długimi, cienkimi, rozgałęzionymi, czasami zespalającymi się wypustkami cytoplazmatycznymi. Wewnątrz ryzopodiów znajdują się mikrofilamenty. Lobopodia– szerokie zaokrąglone wypustki cytoplazmy. Występują w algach z ameboidalnym i monadycznym typem zróżnicowania plechy. Mniej powszechne w algach filopodia- cienkie mobilne formacje przypominające macki, które można wciągnąć do komórki.
Wiele bruzdnic ma ciała pokryte łuskami umieszczonymi na powierzchni komórki. Łuski mogą być pojedyncze lub zamknięte w ciągłej osłonie - płynący. Mogą być organiczne lub nieorganiczne. Organiczne łuski znajdują się na powierzchni zielonych, złotych, kryptofitowych alg. Skład płatków nieorganicznych może obejmować węglan wapnia lub krzemionkę. Płatki węglanu wapnia – kokolity– występuje głównie w algach morskich pierwiosnków.
Często komórki glonów wiciowatych i ameboidalnych znajdują się w domach, które są głównie pochodzenia organicznego. Ich ścianki mogą być cienkie i przezroczyste (rodzaj Dinobryona) lub bardziej trwałe i kolorowe ze względu na osadzanie się w nich soli żelaza i manganu (rodzaj Trachelomonas). Domy mają zwykle jeden otwór, przez który wici mogą wyjść, czasami może być ich kilka. Kiedy glony się rozmnażają, dom nie ulega zniszczeniu, najczęściej jedna z powstałych komórek opuszcza go i buduje nowy dom.
Pokrycie komórek glonów euglenoidalnych nazywa się błonką. Błonka to zbiór błony cytoplazmatycznej i znajdujących się pod nią pasm białkowych, mikrotubul i zbiorników retikulum endoplazmatycznego.
U alg dinofitów osłony komórkowe są reprezentowane przez amfiesmę. Amfiezma składa się z plazmalemy i znajdującego się pod nią zestawu spłaszczonych pęcherzyków, pod którymi znajduje się warstwa mikrotubul. Pęcherzyki wielu dinofitów mogą zawierać płytki celulozowe; nazywa się to amfiemią aktualny, Lub powłoka(narodziny Cerat, Perydynium).
U okrzemek na plazmalemie tworzy się specjalna osłona komórkowa - powłoka, składający się głównie z amorficznej krzemionki. Oprócz krzemionki otoczka zawiera domieszkę związków organicznych i niektórych metali (żelazo, aluminium, magnez).
W ścianach komórkowych alg zielonych, żółto-zielonych, czerwonych i brunatnych głównym składnikiem strukturalnym jest celuloza, która tworzy szkielet (podstawę strukturalną) zanurzoną w matrycy (pożywce półpłynnej) składającej się z pektyny, hemicelulozy, kwasu alginowego i inne substancje organiczne.
Wici
Monadyczne komórki wegetatywne i monadyczne etapy cyklu życiowego (zoospory i gamety) glonów są wyposażone w wici - długie i dość grube wyrostki komórkowe, pokryte zewnętrznie plazmalemmą. Ich liczba, długość, morfologia, miejsce przyczepienia i wzór ruchu są dość zróżnicowane u glonów, ale są stałe w obrębie pokrewnych grup.
Wici mogą być przyczepione do przedniego końca komórki (wierzchołkowy) lub lekko przesunięte w bok (podwierzchołkowe); można je przymocować z boku kuwety (bocznie) i po brzusznej stronie kuwety (brzusznie). Nazywa się wici identyczne pod względem morfologii izomorficzny, jeśli się różnią - heteromorficzny. Izokont- są to wici tej samej długości, heterokontnye– różne długości.
Wici mają jeden plan struktury. Wyróżnia się część wolną (undulipodium), strefę przejściową i ciało podstawne (kinetosom). Poszczególne części wici różnią się liczbą i rozmieszczeniem mikrotubul tworzących szkielet (ryc. 2).

Ryż. 2. Schemat budowy wici glonów (wg: L.L. Velikanov i in., 1981): 1 – przekrój podłużny wici; 2, 3 – przekrój poprzeczny przez czubek wici; 4 – przekrój poprzeczny przez undulipodium; 5 – strefa przejściowa; 6 – przekrój przez podstawę wici – kinetosom
Undulipodium(przetłumaczone z łaciny jako „wavepod”) jest w stanie wykonywać rytmiczne ruchy przypominające fale. Undulipodium to aksonem pokryty błoną. Aksonem składa się z dziewięciu par mikrotubul ułożonych w okrąg i pary mikrotubul pośrodku (ryc. 2). Wici mogą być gładkie lub pokryte łuskami lub mastigonemami (włosami), a u dinofitów i kryptofitów są pokryte zarówno łuskami, jak i włoskami. Wici prymnezjofitów, kryptofitów i zielonych alg mogą być pokryte łuskami o różnych kształtach i rozmiarach.
Strefa przejściowa. Funkcjonalnie odgrywa rolę we wzmocnieniu wici w miejscu jej wyjścia z komórki. W algach występuje kilka rodzajów struktur strefy przejściowej: płyta poprzeczna (dinofity), struktura w kształcie gwiazdy (zielona), spirala przejściowa (heterokontyna), cylinder przejściowy (primnezofity i dinofity).
Ciało podstawowe lub kinetosom. Ta część wici ma strukturę w postaci pustego cylindra, którego ściana jest utworzona przez dziewięć trójek mikrotubul. Funkcją kinetosomu jest połączenie wici z plazmalemmą komórki. Ciała podstawowe wielu glonów mogą brać udział w podziale jądrowym i stać się ośrodkami organizacji mikrotubul.
Mitochondria
Mitochondria występują w komórkach alg eukariotycznych. Kształt i struktura mitochondriów w komórkach glonów jest bardziej zróżnicowana w porównaniu do mitochondriów roślin wyższych. Mogą być okrągłe, nitkowate, siatkowe lub o nieregularnym kształcie. Ich kształt może się różnić w tej samej komórce na różnych etapach cyklu życia. Mitochondria są pokryte powłoką złożoną z dwóch błon. Macierz mitochondrialna zawiera rybosomy i mitochondrialny DNA. Wewnętrzna membrana tworzy fałdy - święta(ryc. 3).

Ryż. 3. Budowa mitochondriów roślinnych (wg:): A – obraz wolumetryczny; B – przekrój podłużny; B – część crista z grzybkowatymi wypustkami: 1 – błona zewnętrzna, 2 – błona wewnętrzna, 3 – crista, 4 – matrix, 5 – przestrzeń międzybłonowa, 6 – rybosomy mitochondrialne, 7 – granulka, 8 – mitochondrialne DNA, 9 – ATP-somy
Są algi cristae różne kształty: krążkowaty (algi euglena), rurkowaty (algi dinofitowe), blaszkowaty (algi zielone, czerwone, kryptomonady) (ryc. 4).

Ryż. 4. Różne rodzaje Cristae mitochondrialne (by:): A – blaszkowate; B – rurowy; B – w kształcie dysku; k - cristae
Cristae w kształcie dysku są uważane za najbardziej prymitywne.
Pigmenty
Wszystkie glony różnią się znacznie zestawem pigmentów fotosyntetycznych. Takie grupy w taksonomii roślin mają status działów.
Głównym pigmentem wszystkich alg jest zielony pigment chlorofil. Znane są cztery rodzaje chlorofilu, różniące się budową: chlorofilA– występuje we wszystkich algach i roślinach wyższych; chlorofil b– występuje w algach zielonych, charofitach, euglenoidach i roślinach wyższych: rośliny zawierające ten chlorofil zawsze mają jasnozieloną barwę; chlorofil c– występuje w algach heterokontowych; chlorofil d– rzadka forma, występująca u alg czerwonych i niebieskozielonych. Większość roślin fotosyntetyzujących zawiera dwa różne chlorofile, z których jeden jest zawsze chlorofilem A. W niektórych przypadkach zamiast drugiego chlorofilu są biliproteiny. Istnieją dwa rodzaje biliprotein występujących w niebiesko-zielonych i czerwonych algach: fikocyjanina– niebieski pigment, fikoerytryna- czerwony pigment.
Obowiązkowymi pigmentami zawartymi w błonach fotosyntetycznych są pigmenty żółte - karotenoidy. Różnią się od chlorofilów widmem pochłanianego światła i uważa się, że pełnią funkcję ochronną, chroniąc cząsteczki chlorofilu przed destrukcyjnym działaniem tlenu cząsteczkowego.
Oprócz wymienionych pigmentów algi zawierają również: fukoksantyna– złoty pigment; ksantofil- brązowy pigment.
Plastydy
Pigmenty w komórkach alg eukariotycznych znajdują się w plastydach, jak we wszystkich roślinach. W algach występują dwa rodzaje plastydów: kolorowe chloroplasty (chromatofory) i bezbarwne leukoplasty (amyloplast). Chloroplasty glonów, w przeciwieństwie do chloroplasty roślin wyższych, są znacznie bardziej zróżnicowane pod względem kształtu i struktury (ryc. 5).

Ryż. 5. Schemat budowy chloroplastów w glonach eukariotycznych (autor:): 1 – rybosomy; 2 – otoczka chloroplastowa; 3 – tylakoid pasowy; 4 – DNA; 5 – fikobilisomy; 6 – skrobia; 7 – dwie membrany z chloroplastowego EPS; 8 – dwie membrany otoczki chloroplastowej; 9 – blaszka; 10 – produkt zapasowy; 11 – rdzeń; 12 – jedna membrana z chloroplastowego EPS; 13 – lipid; 14 – ziarno; 15 – pirenoid. A – tylakoidy zlokalizowane są pojedynczo, nie ma CES – siateczki śródplazmatycznej chloroplastowej (Rhodophyta); B – blaszki dwutylakoidowe, dwie błony CES (Cryptophyta); B – blaszki tritylakoidowe, jedna błona CES (Dinophyta. Euglenophyta); D – blaszki tritylakoidowe, dwie błony CES (Heterokontophyta, Prymnesiophyta); D – blaszki dwu-, sześciotylakoidowe, bez CES (Chlorophyta)
Strukturalna jednostka fotosyntezy eukariontów i prokariotów to tylakoid- płaski worek membranowy. Błony tylakoidów zawierają układy pigmentowe i nośniki elektronów. Faza jasna fotosyntezy jest związana z tylakoidami. Ciemna faza fotosyntezy zachodzi w zrębie chloroplastu. Skorupa zielonych i czerwonych alg składa się z dwóch membran. W innych glonach chloroplast jest otoczony dodatkowym jednym lub dwoma błony siateczki śródplazmatycznej chloroplastów(ON). U euglenaceae i większości dinofitów chloroplast jest otoczony trzema błonami, a u heterokontaceae i kryptofitów - czterema (ryc. 5).
Jądro i aparat mitotyczny
Jądro glonów ma strukturę typową dla eukariontów. Liczba jąder w komórce może wahać się od jednego do kilku. Na zewnątrz jądro pokryte jest otoczką składającą się z dwóch błon, zewnętrzna błona pokryta jest rybosomami. Przestrzeń między błonami jądrowymi nazywa się okołojądrowe. Może zawierać chloroplasty lub leukoplasty, jak u heterokontów i kryptofitów. Macierz jądrowa zawiera chromatynę, która reprezentuje DNA w kompleksie z głównymi białkami - histonami. Wyjątkiem są dinofity, u których liczba histonów jest niewielka i nie ma organizacji chromatyny nukleosomalnej. Nici chromatyny tych alg ułożone są w formie ósemek. W jądrze znajduje się od jednego do kilku jąder, które zanikają lub utrzymują się podczas mitozy.
Mitoza - Pośredni podział glonów może przebiegać na różne sposoby, ale generalnie zachowany jest schemat tego procesu z 4 etapami (ryc. 6).

Ryż. 6. Kolejne fazy mitozy: 1 – interfaza; 2-4 – profaza; 5 – metafaza; 6– anafaza; 7-9 – telofaza; 10– cytokineza
Profaza– najdłuższa faza mitozy. Dokonują się w nim najważniejsze przemiany: jądro zwiększa swoją objętość, zamiast ledwo zauważalnej sieci chromatyny, pojawiają się w nim chromosomy w postaci cienkich, długich, zakrzywionych i słabo spiralnie skręconych nitek, tworzących rodzaj kuli. Od samego początku profazy widać, że chromosomy składają się z 2 nici (wynik ich replikacji w interfazie). Połówki chromosomów (chromatyd) są ułożone równolegle do siebie. W miarę rozwoju profazy nici stają się coraz bardziej spiralne, a powstałe chromosomy stają się coraz bardziej skracane i zagęszczane.
Pod koniec profazy ujawniają się indywidualne cechy morfologiczne chromosomów. Następnie jąderka znikają, błona jądrowa rozpada się na osobne, krótkie cysterny, nie do odróżnienia od elementów EPS, w wyniku czego nukleoplazma miesza się z hialoplazmą i powstaje myksoplazma; Włókna achromatyczne – wrzeciono rozszczepienia – powstają z substancji jądra i cytoplazmy.
Wrzeciono rozszczepienia jest dwubiegunowe i składa się z wiązek mikrotubul rozciągających się od jednego bieguna do drugiego. Po zniszczeniu błony jądrowej każdy chromosom jest przyłączany do nici wrzeciona za pomocą swojego centromeru. Po przyłączeniu się chromosomów do wrzeciona ustawiają się one w płaszczyźnie równikowej komórki, tak że wszystkie centromery znajdują się w tej samej odległości od jej biegunów.
Metafaza. W tej fazie mitozy chromosomy osiągają maksymalne zagęszczenie i uzyskują charakterystyczny kształt charakterystyczny dla każdego gatunku rośliny. Zwykle są dwuramienne iw takich przypadkach w miejscu przegięcia tzw centromer, chromosomy są połączone z włóknami achromatyny wrzeciona. W metafazie wyraźnie widać, że każdy chromosom składa się z dwóch chromatyd potomnych. Znajdują się one mniej więcej równolegle w płaszczyźnie równikowej komórki. Pod koniec tego etapu każdy chromosom dzieli się na dwie chromatydy, które pozostają połączone tylko w centromerze. Później centromery również podzieliły się na dwie siostrzane; podczas gdy siostrzane centromery i chromatydy są zwrócone w stronę przeciwnych biegunów.
Anafaza. Najkrótsza faza mitozy. Chromosomy potomne - chromatydy - rozchodzą się do przeciwnych biegunów komórki. Teraz wolne końce chromatyd są skierowane w stronę równika, a kinetochory - w stronę biegunów. Uważa się, że chromatydy rozdzielają się w wyniku skurczu włókien wrzeciona achromatyny, które znajdują się blisko centromeru. Chromosomy stają się mniej zauważalne z powodu odwijania i wydłużania. W środku komórki (wzdłuż równika) czasami już na tym etapie pojawiają się fragmenty ściany komórkowej – fragmoplast.
Telofaza. Proces rozwijania trwa - desspiralizacja i wydłużanie chromosomów. Wreszcie giną w polu widzenia mikroskopu optycznego. Błona jądrowa i jąderko zostają przywrócone. Ten sam proces zachodzi jak w profazie, tylko w odwrotnej kolejności. Chromosomy mają teraz po jednej chromatydzie. Przywrócona zostaje struktura jądra międzyfazowego, wrzeciono zmienia się z beczkowatego na stożkowy.
Tak to się kończy karotomia– potem następuje rozszczepienie jądrowe plazmotomia. Organelle cytoplazmatyczne rozmieszczone są pomiędzy komórkami potomnymi, a niektóre z nich (dyktosomes, mitochondria i plastydy) ulegają znaczącym modyfikacjom. Wreszcie to się dzieje cytokineza– powstawanie ściany komórkowej pomiędzy jądrami potomnymi. Z poprzedniej komórki powstały dwie nowe; każdy z nich ma jądro zawierające diploidalną liczbę chromosomów.
W zależności od zachowania błony jądrowej alg, istnieją Zamknięte, półzamknięte I otwarty mitozy. W zamkniętej mitozie segregacja chromosomów zachodzi bez uszkodzenia błony jądrowej. W mitozie półzamkniętej otoczka jądrowa utrzymuje się przez cały czas mitozy, z wyjątkiem stref polarnych. W otwartej mitozie błona jądrowa zanika w profazie. W zależności od kształtu wrzeciona wyróżnia się podziały pleuromitoza I ormitoza.
W pleuromitozie płytka metafazowa nie tworzy się w metafazie, a wrzeciono jest reprezentowane przez dwa półwrzeciona umieszczone pod kątem względem siebie na zewnątrz lub wewnątrz jądra. Podczas ormitozy w metafazie chromosomy ustawiają się w jednej linii z równikiem wrzeciona dwubiegunowego. W zależności od połączenia tych właściwości u glonów wyróżnia się następujące rodzaje mitozy (ryc. 7, 8):
Zamknięta mitoza zewnątrzjądrowa

Zamknięta mitoza wewnątrzjądrowa


Mitoza półzamknięta


Otwarta mitoza

Ryż. 7. Schemat głównych typów mitoz w algach (wg: S.A. Karpov, rok). Linie wewnątrz lub na zewnątrz jądra - mikrotubule wrzecionowe
Ośrodkami organizowania mikrotubul wrzeciona mitotycznego w ormitozie półzamkniętej mogą być kinetosomy i inne struktury:
– ormitoza otwarta, spotykana u kryptofitów, gorzkowca, gorzowatych;
– ormitoza półzamknięta, występująca w kolorze zielonym, czerwonym, brązowym itp.;
– ormitoza zamknięta, występująca u euglenoidów;
– u niektórych dinofitów występuje pleuromitoza zamknięta, wewnątrzjądrowa lub zewnątrzjądrowa;
– mitoza półzamknięta, podczas metafazy centriole nie znajdują się na biegunach, ale w rejonie płytki metafazowej; można zaobserwować u zielonych trebuxiaceae.



Ryż. 8. Schemat porównujący mitozy (A) zamknięte, (B) metacentryczne i (C) otwarte (wg: L.E. Graham, L.W. Wilcox, 2000)
Podczas mitozy zmienia się również kształt wrzeciona i kształt biegunów wrzeciona, a także czas istnienia wrzeciona międzystrefowego. Szczyt mitoz występuje w ciemnej porze dnia. W komórkach wielojądrzastych podział jądrowy może zachodzić synchronicznie. asynchronicznie, falowo.
Pytania kontrolne
1. Wymień główne elementy strukturalne komórek roślinnych.
2. Różnica w budowie komórek glonów od komórek roślin wyższych.
3. Osłony komórkowe glonów.
4. Czym jest teka? W jakich algach występuje?
5. Główne pigmenty z alg. Lokalizacja pigmentów w komórkach glonów.
6. Struktura plastydów.
7. Cechy strukturalne plastydów alg.
8. Struktura mitochondriów.
9. Cechy budowy mitochondriów glonów.
10. Budowa jądra i błon jądrowych. Cechy błon jądrowych komórek glonów.
11. Schemat mitozy. Charakterystyka faz mitozy.
12. Rodzaje mitozy w komórkach glonów.
13. Jaka jest różnica pomiędzy pleuromitozą a ormitozą?
14. Rodzaje pseudopodiów glonów.
2. OGÓLNA CHARAKTERYSTYKA ALG
2.1. Rodzaje mocy
Głównym rodzajem odżywiania alg jest fototroficzny typ. We wszystkich działach glonów są przedstawiciele, którzy są ścisłymi (obowiązkowymi) fototrofami. Jednak wiele glonów dość łatwo przechodzi z fototroficznego rodzaju odżywiania na asymilację materia organiczna, Lub heterotroficzny rodzaj jedzenia. Jednak najczęściej przejście na odżywianie heterotroficzne w algach nie prowadzi do całkowitego zaprzestania fotosyntezy, to znaczy w takich przypadkach możemy mówić o miksotroficzny, lub mieszany rodzaj żywienia.
Wykazano zdolność do wzrostu na podłożach organicznych w ciemności lub przy świetle przy braku dwutlenku węgla w przypadku wielu niebiesko-zielonych, zielonych, żółto-zielonych, okrzemek itp. Zauważono, że w przypadku alg wzrost heterotroficzny jest wolniejszy niż wzrost autotroficzny w świetle.
Różnorodność i plastyczność sposobów żerowania glonów pozwala na ich szerokie rozpowszechnienie i zajmowanie różnych nisz ekologicznych.
2.2. Rodzaje thalli
Przedstawiono ciało wegetatywne glonów plecha, Lub plecha, niezróżnicowane na narządy - korzeń, łodyga, liść. W obrębie struktury plechy glony wyróżniają się bardzo dużym zróżnicowaniem morfologicznym (ryc. 9). Reprezentowane są przez organizmy jednokomórkowe, wielokomórkowe i niekomórkowe. Ich rozmiary są bardzo zróżnicowane: od najmniejszych organizmów jednokomórkowych po gigantyczne organizmy wielometrowe. Kształt ciała glonów jest również zróżnicowany: od najprostszych kulistych po kompleksowo rozcięte formy przypominające rośliny wyższe.
Ogromną różnorodność glonów można sprowadzić do kilku typów struktury morfologicznej: monadyczny, ryzopodialny, palmelloid, kokosowy, trójdzielny, heterotrychalny, miąższowy, syfonalny, syfonokladalny.
Monadyczny (wiciowy) typ struktury plechy
Najbardziej charakterystyczną cechą charakteryzującą ten typ budowy jest obecność wici, za pomocą których organizmy monadyczne aktywnie poruszają się w środowisku wodnym (ryc. 9, A). Ruchliwe formy wici są szeroko rozpowszechnione u glonów. Wśród wielu grup glonów dominują formy wiciowców: euglenofity, dinofity, kryptofity, raphidae, złote algi i występują w żółto-zielonych i zielonych algach. W stanie wegetatywnym algi brunatne nie mają struktury monadycznej, ale podczas rozmnażania (rozmnażania) powstają stadia monadyczne. Liczba wici, ich długość, charakter umieszczenia i ruchu są zróżnicowane i mają ważne znaczenie systematyczne.

Ryż. 9. Morfologiczne typy budowy plechy glonów (wg:): A– monadyczny ( Chlamydomonas); B– ameboid ( Rizochrisa); W– palmelloid ( Hydrorus); G– kokosowy ( Pediastrum); D– sarcynoid ( Chlorosarcyna); mi– nitkowate ( Ulotrix); I– wielonitkowe ( Fritchiela); Z, I– tkanina ( Furcellaria, Laminaria); DO– syfonalny ( Caulerpa); L– syfonokladalny ( Kladofora)
Mobilność glonów monadowych determinuje polarność struktury ich komórek i kolonii. Wici są zwykle przyczepione do przedniego bieguna komórki lub w jego pobliżu. Podstawowy kształt komórki ma kształt łzy z mniej lub bardziej zwężonym przednim biegunem wici. Jednakże organizmy monadyczne często odbiegają od tego podstawowego kształtu i mogą być asymetryczne, mieć kształt spiralny, mieć zwężający się tylny koniec itp.
Kształt ogniwa w dużej mierze zależy od osłon ogniw, które się różnią wielka różnorodność(plazmalemma; błonka; osłonka; składająca się z łusek organicznych, krzemionkowych lub wapiennych; dom; błona komórkowa). Dziwaczne zarysy komórek niektórych złotych alg tworzą rodzaj wewnątrzkomórkowego szkieletu, składającego się z pustych w środku rurek krzemionkowych. Błona komórkowa jest zwykle gładka, czasami ma różne wypustki lub jest inkrustowana solami żelaza lub wapnia i wtedy przypomina dom. W skorupie tworzą się tylko małe otwory, przez które wici wydostają się na zewnątrz.
Polaryzacja organizmów monadycznych przejawia się także w rozmieszczeniu struktur wewnątrzkomórkowych. Na przednim końcu komórki często znajduje się różnie ułożona gardło, zwykle pełniący funkcję wydalniczą. Tylko kilka fagotroficznych wiciowców ma gardło, które pełni funkcję ujścia komórki - cytostom.
Osobliwe organelle charakterystyczne dla alg o budowie monadowej to kurczliwe wakuole pełniąc funkcję osmoregulacyjną, ciała śluzowe I kłujące struktury. Kapsułki kłujące znajdują się w algach dinofitów, euglenoidów, złotych, rafidofitów, kryptofitów i pełnią funkcję ochronną. Pojedyncze jądro zajmuje centralne miejsce w komórkach. Chloroplasty, zróżnicowane pod względem kształtu i koloru, mogą być osiowe lub ścienne.
Tendencja do zwiększania rozmiarów ciała objawia się tworzeniem różnych kolonii. W najprostszych przypadkach kolonie powstają w wyniku braku rozłączenia dzielących się komórek. Obserwuje się kolonie o kształcie pierścieniowym, krzaczastym, drzewiastym i kulistym. Zielone organizmy monadyczne charakteryzują się głównie koloniami tego typu cenobiści ze stałą liczbą komórek dla każdego typu.
W niesprzyjających warunkach organizmy monadyczne zrzucają lub cofają wici, tracąc ruchliwość i otaczając się obfitą ilością śluzu.
Obiecujący okazał się typ monadyczny. Na jego podstawie opracowano inne, bardziej złożone konstrukcje.
Struktura typu ryzopodialnego (ameboidalnego).
Najważniejszymi cechami struktury typu ameboidalnego jest brak silnych osłon komórkowych i zdolność do tego ameboid ruch za pomocą wyrostków cytoplazmatycznych chwilowo utworzonych na powierzchni komórki - pseudopodium. Istnieje kilka rodzajów pseudopodiów, z których najczęściej obserwuje się glony ryzopodia I lodopodia, rzadziej aksopodia(ryc. 9, B).
Nie ma zasadniczych różnic w budowie i mechanizmie działania układów skurczowych, które determinują ruchliwość organizmów monadycznych i ameboidalnych na poziomie molekularnym. Ruch ameboidalny powstał prawdopodobnie w wyniku przystosowania się komórek wiciowych do uproszczonych warunków życia, co doprowadziło do uproszczenia budowy ciała.
Komórki glonów ameboidalnych zawierają jądra, plastydy, mitochondria i inne organelle charakterystyczne dla eukariontów: często obserwuje się kurczliwe wakuole, piętno i ciała podstawne zdolne do tworzenia wici.
Wiele organizmów ameboidalnych prowadzi przywiązany tryb życia. Potrafią budować domy o różnych kształtach i konstrukcjach: cienkie, delikatne lub szorstkie, grubościenne.
Typ ciała ameboidalnego nie jest tak rozpowszechniony jak typ monadyczny. Obserwuje się go tylko u alg złocistych i żółto-zielonych.
Struktura typu palmelloidowego (hemimonadalnego).
Charakterystyczne dla tego typu struktury jest połączenie stacjonarnego trybu życia roślin z obecnością organelli komórkowych charakterystycznych dla organizmów monadycznych: wakuoli kurczliwych, piętna, wici. Zatem komórki wegetatywne mogą posiadać wici, za pomocą których w ograniczonym stopniu poruszają się w śluzie kolonialnym, lub wici są zachowywane w komórkach nieruchomych w znacznie zredukowanej formie.
Komórki typu palmelloidowego (hemimonady) charakteryzują się budową polarną. Czasami klatki umieszczane są w domach.
Glony Hemimonad często tworzą kolonie. W najprostszym przypadku śluz jest bezstrukturalny, a komórki są w nim rozmieszczone w dowolnej kolejności. Dalsza złożoność takich kolonii objawia się zarówno różnicowaniem śluzu, jak i bardziej uporządkowanym rozmieszczeniem komórek w śluzie. Kolonie typu dendrytycznego (rodzaj Hydrorus) (ryc. 9, W).
Struktura typu palmelloidowego (hemimonada) była ważnym etapem ewolucji morfologicznej glonów w kierunku od mobilnych form monadycznych do typowo roślinnych form nieruchomych.
Struktura typu kokosowego
Ten typ łączy glony jednokomórkowe i kolonialne, nieruchome w stanie wegetatywnym. Komórki typu kokosowego są pokryte błoną i mają protoplast typu roślinnego (tonoplast bez kurczliwych wakuoli, znamion, wici). Utrata cech struktury monadycznej w strukturze komórkowej organizmów prowadzących roślinny, siedzący tryb życia i nabycie nowych struktur charakterystycznych dla komórek roślinnych jest kolejnym ważnym krokiem w ewolucji glonów według rodzaju rośliny.
Ogromna różnorodność glonów o strukturze typu kokosowego wiąże się z obecnością osłon komórkowych. Powłoka warunkuje obecność różnych komórek: kulistych, jajowatych, wrzecionowatych, elipsoidalnych, cylindrycznych, gwiaździstych, spiralnych, gruszkowatych itp. Różnorodność form potęguje także rzeźbiarska dekoracja powłoki komórkowej - kolce, kolce, włosie, procesy rogowe.
Algi kokosowe tworzą kolonie o różnych kształtach, w których komórki są zjednoczone ze śluzem lub bez.
Struktura typu kokosowego jest szeroko rozpowszechniona w prawie wszystkich podziałach glonów eukariotycznych (z wyjątkiem Euglenaceae).
W ujęciu ewolucyjnym strukturę kokoidową można uznać za wyjściową dla pojawienia się plech wielokomórkowych, a także struktur syfonalnych i syfonokladalnych (ryc. 9, G, D).
Struktura typu trichal (nitkowatego).
Cechą charakterystyczną struktury typu nitkowatego jest nitkowaty układ nieruchomych komórek, które powstają wegetatywnie w wyniku podziału komórek zachodzącego przeważnie w jednej płaszczyźnie. Komórki włókienkowe mają strukturę polarną i mogą rosnąć tylko w jednym kierunku, pokrywającym się z osią wrzeciona jądrowego.
W najprostszych przypadkach plechy o strukturze nitkowatej składają się z komórek podobnych morfologicznie do siebie. Jednocześnie u wielu glonów w obszarach włókien, które stają się cieńsze lub szersze w kierunku końców, komórki różnią się kształtem od pozostałych. W tym przypadku często dolna komórka, pozbawiona chloroplastów, zamienia się w bezbarwny ryzoid lub stopę. Gwinty mogą być proste lub rozgałęzione, jedno- lub wielorzędowe, wolno żyjące lub przyczepione.
Strukturę nitkowatą reprezentują algi zielone, czerwone, żółto-zielone i złote (ryc. 9, mi).
Struktura typu heterotrychalnego (niewłóknistego).
Typ heterofilamentowy powstał na bazie typu nitkowatego. Plecha heterofilamentowa składa się głównie z nitek poziomych pełzających po podłożu, pełniących funkcję przyczepu, oraz nitek pionowych wznoszących się ponad podłoże, pełniących funkcję asymilacyjną. Te ostatnie noszą narządy rozrodcze.
U niektórych glonów rozróżnia się włókna pionowe międzywęźle I węzły, od którego odchodzą okółki gałęzi bocznych, które również mają budowę segmentową. Ponadto z węzłów mogą wyrastać dodatkowe nici, tworząc skorupę skorupową międzywęźli. Funkcję mocowania do podłoża pełnią bezbarwne ryzoidy. Strukturę tę można spotkać u charofitów, alg zielonych, brązowych, czerwonych, niektórych żółtozielonych i złotych (ryc. 9, I).
Struktura miąższowa (tkankowa).
Jeden z kierunków ewolucji plechy heterofilamentowej wiązał się z pojawieniem się plechy miąższowej. Zdolność do nieograniczonego wzrostu i podziału komórek w różnych kierunkach doprowadziła do powstania obszernych makroskopowych plech z morfofunkcjonalnym zróżnicowaniem komórek w zależności od ich położenia w wzgórzu (kora, warstwa pośrednia, rdzeń).
W obrębie tego typu występuje stopniowe powikłanie plech od prostych płytek do złożonych, zróżnicowanych plech z prymitywnymi tkankami i narządami. Struktura miąższowa jest najwyższym etapem ewolucyjnym zróżnicowania morfologicznego ciała glonów. Jest szeroko reprezentowany w dużych algach: brunatnych, czerwonych i zielonych – tzw. glonach makrofitowych (ryc. 10).

Ryż. 10. Przekrój plechy brunatnicy (wg:): 1 – kora zewnętrzna; 2 – kora wewnętrzna; 3 – rdzeń
Konstrukcja typu syfonowego
Struktura typu syfonalnego (niekomórkowego) charakteryzuje się brakiem przegród komórkowych wewnątrz plechy, która osiąga stosunkowo duże, zwykle makroskopowe rozmiary i pewien stopień zróżnicowania, w obecności dużej liczby organelli. Przegrody w takiej plechy mogą pojawić się tylko przypadkowo, gdy jest ona uszkodzona lub podczas tworzenia się narządów rozrodczych. W obu przypadkach proces tworzenia przegród różni się od powstawania organizmu wielokomórkowego.
Struktura syfonalna występuje w niektórych algach zielonych i żółto-zielonych. Jednak ten kierunek ewolucji morfologicznej okazał się ślepą uliczką.
Konstrukcja typu syfonokladalnego
Główną cechą struktury typu syfonokladalnego jest zdolność do tworzenia kompleksowo ułożonych wzgórz, składających się głównie z segmentów wielojądrzastych, z pierwotnego wzgórza niekomórkowego. Na czym opiera się tworzenie takiego plechy podział segregacyjny, w którym mitoza nie zawsze kończy się cytokinezą.
Budowa syfonokladalna znana jest jedynie u niewielkiej grupy zielonych alg morskich.
2.3. Rozmnażanie się glonów
Rozmnażanie jest główną właściwością istot żywych. Jego istota polega na reprodukcji własnego rodzaju. W algach rozmnażanie może odbywać się bezpłciowo, wegetatywnie i płciowo.
Rozmnażanie bezpłciowe
Rozmnażanie bezpłciowe glonów odbywa się za pomocą wyspecjalizowanych komórek - spór. Zarodnikowaniu zwykle towarzyszy podział protoplastu na części i uwolnienie produktów podziału z otoczki komórki macierzystej. Ponadto przed podziałem protoplastu zachodzą w nim procesy prowadzące do jego odmłodzenia. Uwalnianie produktów podziału ze skorupy komórki macierzystej jest najbardziej znaczącą różnicą między prawdziwym rozmnażaniem bezpłciowym a rozmnażaniem wegetatywnym. Czasami w komórce powstaje tylko jeden zarodnik, ale nawet wtedy opuszcza on skorupę matki.
Zarodniki są zwykle produkowane w specjalnych komórkach zwanych sporangia, różniące się od zwykłych komórek wegetatywnych wielkością i kształtem. Powstają jako wyrostki zwykłych komórek i pełnią jedynie funkcję tworzenia zarodników. Czasami zarodniki powstają w komórkach, które nie różnią się kształtem i rozmiarem od zwykłych komórek wegetatywnych. Zarodniki różnią się także od komórek wegetatywnych kształtem i mniejszym rozmiarem. Liczba zarodników w sporangium waha się od jednego do kilkuset. Zarodniki są etapem rozprzestrzeniania się w cyklu życiowym glonów.
W zależności od konstrukcji wyróżnia się:
zoospory– ruchliwe zarodniki glonów zielonych i brunatnych, mogą posiadać jedną, dwie, cztery lub wiele wici, w tym drugim przypadku wici są umiejscowione w koronie na przednim końcu zarodnika lub parami na całej powierzchni;
hemizoospory– zoospory, które utraciły wici, ale zachowują kurczliwe wakuole i piętno;
aplanospory– nieruchliwe zarodniki, które pokrywają się błoną wewnątrz komórki macierzystej;
sporty motorowe– aplanospory, mające kształt komórki macierzystej;
hipnospory– nieruchliwe zarodniki o pogrubionych otoczkach, zaprojektowane tak, aby przetrwać niesprzyjające warunki środowiskowe.
W czerwonych algach rozmnażanie bezpłciowe odbywa się za pomocą monospora, bispor, tetraspora Lub polispora. Monospory nie mają wici ani błony. Po opuszczeniu komórki macierzystej są zdolne do ruchu ameboidalnego. Monospory różnią się od komórek wegetatywnych tym, że mają jajowaty lub kulisty kształt, są bogate w składniki odżywcze i intensywnie wybarwione.
Mają strukturę zarodników i rodzaje sporulacji bardzo ważne dla systematyki glonów, ponieważ odzwierciedlają one różnice w organizacji form przodków różnych grup glonów.
Rozmnażanie wegetatywne
Rozmnażanie wegetatywne w algach może przebiegać na kilka sposobów: prosty podział na dwie części, wielokrotny podział, pączkowanie, fragmentacja plechy, rozłogi, zawiązki czerwowe, paraspory, guzki, akinety.
Prosty podział.
Ta metoda rozmnażania występuje tylko w jednokomórkowych formach glonów. Podział zachodzi najłatwiej w komórkach o budowie ciała typu ameboidalnego.
Podział form ameboidalnych. Podział ameboidów jest możliwy w dowolnym kierunku. Rozpoczyna się od wydłużenia ciała ameby, a następnie na równiku zarysowuje się przegrodę, która dzieli ciało na dwie mniej więcej równe części. Podziałowi cytoplazmy towarzyszy podział jądra. Czasami podział poprzedza przejście do stanu stacjonarnego w wyniku cofnięcia nóg, a komórka nabiera kształtu kulistego. Jednocześnie protoplazma traci przezroczystość, a kurczliwa wakuola znika. Pod koniec podziału komórka jest rozciągana, splatana, po czym pojawiają się pseudonóżki.
Podział form biczowanych. W formach wiciowych występują najbardziej złożone rodzaje rozmnażania wegetatywnego. Rodzaje reprodukcji zależą od poziomu organizacji i stopnia polaryzacji komórek. W niektórych kryptofitach, złotych i zielonych algach rozmnażanie prosty podział rozwidlenie zachodzi w stanie ruchomym tylko wzdłuż osi podłużnej i zaczyna się od przedniego bieguna komórki. W takim przypadku wici mogą trafić tylko do jednej komórki lub zostać równo podzielone między nowe komórki. Komórka, która nie ma wici, sama ją tworzy. U większości alg Volvox i Euglena podczas rozmnażania błona komórkowa staje się śluzem, a podział zachodzi w stanie stacjonarnym. We wszystkich wiciowych formach, które mają skorupę, komórki są podzielone na dwie równe lub nierówne części. Po rozdzieleniu stara skorupa zostaje zrzucona i powstaje nowa.
Podział form kokosowych. U glonów o strukturze komórkowej typu kokosowego rozmnażanie wegetatywne nabiera typowych cech podziału stacjonarnej komórki roślinnej z dobrze określoną ścianą komórkową. W swojej prostocie zbliża się do rozmnażania wegetatywnego typu ameboidalnego i odbywa się poprzez prosty podział komórki na dwie części.
Początkujący.
Komórki nitkowatych rozgałęzionych alg charakteryzują się dwoma sposobami rozmnażania wegetatywnego: prostym podziałem na dwie części i pączkowaniem. Połączenie tych metod rozmnażania powoduje boczne rozgałęzianie glonów nitkowatych.
Podział.
Fragmentacja jest nieodłączna dla wszystkich grup glonów wielokomórkowych i objawia się w różnych postaciach: tworzeniu hormogonium, regeneracji oderwanych części plechy, samoistnej utracie gałęzi, odrastaniu ryzoidów. Przyczyną fragmentacji mogą być czynniki mechaniczne (fale, prądy, gryzienie zwierząt) lub śmierć niektórych komórek. Przykładem tej ostatniej metody fragmentacji jest powstawanie hormogonii w niebieskozielonych algach. Z każdego hormonu może powstać nowy osobnik. Rozmnażanie przez części plechy, charakterystyczne dla alg czerwonych i brunatnych, nie zawsze prowadzi do wznowienia normalnych roślin. Wodorosty rosnące na skałach i skałach są często częściowo lub całkowicie niszczone przez działanie fal. Ich oderwane fragmenty lub całe plechy nie są w stanie ponownie przyczepić się do gleby stałej ze względu na ciągły ruch wody. Ponadto narządy przyczepu nie tworzą się ponownie. Jeśli takie thalli znajdą się w spokojnych miejscach z błotnistym lub piaszczystym dnem, leżąc na ziemi, nadal rosną. Z biegiem czasu starsze części obumierają, a odchodzące od nich gałęzie zamieniają się w niezależne plechy; w takich przypadkach mówi się o niezwiązanych lub wolno żyjących formach odpowiedniego gatunku. Glony ulegają znacznym zmianom: ich gałęzie stają się cieńsze, węższe, a gałęzie słabsze. Nieprzyłączone formy glonów nie tworzą narządów rozmnażania płciowego i bezpłciowego i mogą rozmnażać się jedynie wegetatywnie.
Rozmnażanie przez pędy, rozłogi, pąki czerwowe, guzki, akinety.
W postaciach tkankowych zielonych, brunatnych i czerwonych alg rozmnażanie wegetatywne przybiera pełną formę, która niewiele różni się od rozmnażania wegetatywnego roślin wyższych. Zachowując zdolność do regeneracji części plechy, formy tkankowe uzyskują wyspecjalizowane formacje, które pełnią funkcję rozmnażania wegetatywnego. Wiele gatunków alg brunatnych, czerwonych, zielonych i chara ma pędy, na których wyrastają nowe plechy. Na plechach niektórych alg brunatnych i czerwonych rozwijają się pąki lęgowe (propagule), które opadają i wyrastają nowe plechy.
Za pomocą jednokomórkowych lub wielokomórkowych zimujących guzków następuje sezonowa odnowa glonów charofitycznych. Niektóre glony nitkowate (na przykład zielone algi ulothrix) rozmnażają się przez akinety - wyspecjalizowane komórki o pogrubionej skorupie i dużej ilości rezerwowych składników odżywczych. Są w stanie przetrwać niekorzystne warunki.
Rozmnażanie płciowe
Rozmnażanie płciowe u alg jest związane z procesem płciowym, który polega na połączeniu dwóch komórek, w wyniku czego powstaje zygota, z której wyrasta nowy osobnik lub wytwarza zoospory.
Istnieje kilka rodzajów rozmnażania płciowego u glonów:
hologamia(koniugacja) – bez tworzenia wyspecjalizowanych komórek;
gametogamia– za pomocą wyspecjalizowanych komórek – gamet.
Holologia. W najprostszym przypadku proces ten zachodzi poprzez fuzję dwóch nieruchomych komórek wegetatywnych pozbawionych błon komórkowych. W jednokomórkowych wiciowych formach alg proces seksualny odbywa się poprzez połączenie dwóch osobników.
Kiedy zawartość dwóch wiciowych komórek wegetatywnych łączy się, nazywa się proces płciowy koniugacja. Podczas koniugacji następuje fuzja dwóch komórek, które pełnią funkcję komórek rozrodczych - gamet. Fuzja zawartości komórek następuje poprzez specjalnie utworzony kanał koniugacyjny, w wyniku czego powstaje zygota, która następnie zostaje pokryta grubą błoną i przekształca się w zygosporę. Jeśli szybkość przepływu zawartości komórek jest taka sama, w kanale koniugacyjnym tworzy się zygota. W tym przypadku podział komórek na męskie i żeńskie jest warunkowy.
Gametogamia. Rozmnażanie płciowe u glonów, w tym jednokomórkowych, następuje najczęściej poprzez podział zawartości komórek i tworzenie w nich wyspecjalizowanych komórek rozrodczych - gamet. U wszystkich alg zielonych i brunatnych gamety męskie mają wici, ale gamety żeńskie nie zawsze je mają. U prymitywnych glonów gamety powstają w komórkach wegetatywnych. W bardziej zorganizowanych formach gamety znajdują się w specjalnych komórkach zwanych gametangiami. Komórka wegetatywna lub gametangium może zawierać od jednej do kilkuset gamet. W zależności od wielkości łączących się gamet wyróżnia się kilka rodzajów gametogamii: izogamia, heterogamia, oogamia.
Jeśli łączące się gamety mają ten sam kształt i rozmiar, ten proces seksualny nazywa się izogamia.
Jeśli łączące się gamety mają ten sam kształt, ale różne rozmiary (gameta żeńska jest większa niż męska), wówczas mówią o heterogamia.
Proces seksualny, podczas którego nieruchoma duża komórka łączy się - jajko i mobilna mała komórka męska - sperma, zwany oogamia. Nazywa się gametangia z jajami archegonia Lub oogonia i z plemnikami – pylniki. Gamety męskie i żeńskie mogą rozwijać się u tego samego osobnika (jednopienne) lub u różnych osobników (dwupienne). Zygota powstała w wyniku połączenia gamet po pewnych zmianach zamienia się w zygosporę. Ta ostatnia jest zwykle pokryta gęstą skorupą. Zygospora może pozostawać w stanie uśpienia przez długi czas (do kilku miesięcy) lub kiełkować bez okresu uśpienia.
Autogamia. Specjalny rodzaj procesu seksualnego. Polega na tym, że jądro komórkowe dzieli się mejotycznie, z czterech powstałych jąder dwa ulegają zniszczeniu, a pozostałe dwa jądra łączą się, tworząc zygotę, która bez okresu odpoczynku powiększa się i zamienia w auksosporę. W ten sposób ludzie się odmładzają.
2.4. Cykle życiowe glonów
Cykl życia lub cykl rozwoju, to zespół wszystkich etapów rozwoju organizmów, w wyniku których z pewnych osobników lub ich podstaw powstają nowe osobniki i podstawy do nich podobne. Etap starzenia się, prowadzący do śmierci jednostki i okresy odpoczynku wykraczają poza cykl życia. Cykl rozwojowy może być prosty lub złożony, co jest związane ze stosunkiem diploidalnych i haploidalnych faz jądrowych, lub formy rozwoju(ryc. 11).

Ryż. 11. Cykle życiowe glonów (wg:): I – haplobiont z redukcją zygotyczną; II – haplodiplobiont z redukcją sporadyczną; III – diplobiont z redukcją gametową; IV – haplodiplobiont z redukcją somatyczną. Faza dominująca w przypadkach I i III jest wielokomórkowa; jeśli jest jednokomórkowy, to jest długotrwały i zdolny do rozmnażania mitotycznego; 1 – faza haploidalna; 2 – faza diploidalna
Pojęcie cyklu życia wiąże się z przemianą pokoleń. Pod Pokolenie rozumieć ogół jednostek rozpatrywany w odniesieniu do przodków i potomków żyjących w bliskim czasie i genetycznie z nim spokrewnionych.
Cyjanobakterie charakteryzują się prostym cyklem życiowym, w którym nie występuje rozmnażanie płciowe. Ich cykle życiowe są zakończone ( duży) I mały. Mały cykl życiowy odpowiada pewnym gałęziom dużego cyklu i prowadzi do powtarzającego się powstawania pośrednich stanów wiekowych osobników cyjanobakterii . Cykl rozwojowy sinic obejmuje zatem pewne segmenty rozwoju jednego lub kilku kolejnych pokoleń określonej formy systematycznej: od zawiązka osobnika do pojawienia się nowych zawiązków tego samego typu.
U większości glonów z procesem płciowym, w zależności od pory roku i warunków zewnętrznych, obserwuje się różne formy rozmnażania (płciowe i bezpłciowe), ze zmianą haploidalnej i diploidalnej fazy jądrowej. Zmiany, jakim podlega jednostka pomiędzy tymi samymi fazami rozwoju, stanowią jej cykl życia.
Narządy rozmnażania płciowego i bezpłciowego mogą rozwijać się u tego samego osobnika lub u różnych osobników. Rośliny wytwarzające zarodniki nazywane są sporofity, a tworzące się gamety są gametofity. Nazywa się rośliny, które mogą wytwarzać zarówno zarodniki, jak i gamety gametosporofity. Gametosporofity są charakterystyczne dla wielu glonów: zielonych (Ulvacaceae), brunatnych (Ectocarpaceae) i czerwonych (Bangieaceae). Rozwój narządów rozrodczych tego czy innego typu zależy od temperatury otoczenia. Na przykład na plechach blaszkowatych czerwonych alg Porfira Tenera w temperaturach poniżej 15–17 °C tworzą się narządy rozmnażania płciowego, a w wyższych temperaturach tworzą się narządy rozmnażania bezpłciowego. Ogólnie rzecz biorąc, u wielu glonów gamety rozwijają się w niższej temperaturze niż zarodniki. Na rozwój niektórych narządów rozrodczych wpływają także inne czynniki: intensywność światła, długość dnia, skład chemiczny wody, w tym jej zasolenie.
Gametofity, gametosporofity i sporofity glonów nie mogą różnić się wyglądem lub mieć dobrze określone różnice morfologiczne. Wyróżnić izomorficzny(podobne) i heteromorficzny(różne) zmiany form rozwoju, które utożsamiane są z przemianą pokoleń. U większości gametosporofitów nie występuje przemiana pokoleń. Czasami gametofity i sporofity, nie różniąc się morfologicznie, żyją w różnych warunkach środowiskowych; w niektórych przypadkach różnią się one morfologicznie. Na przykład w czerwonych algach Porfira Tenera sporofity mają postać rozgałęzionych jednorzędowych włókien, które są osadzone w podłożu wapiennym (muszle mięczaków, skały). Rosną preferencyjnie przy słabym oświetleniu i wnikają w podłoże na duże głębokości. Gametofity tych glonów mają postać płytek i rosną w dobrym świetle w pobliżu brzegu wody oraz w strefie pływów.
Przy heteromorficznej przemianie pokoleń struktura sporofitów i gametofitów różni się w niektórych przypadkach dość znacząco. Tak więc w zielonych algach z rodzajów Akrosyfonia I Spongomorfa gametofit jest wielokomórkowy, ma kilka centymetrów wysokości, a sporofit jest jednokomórkowy, mikroskopijny. Możliwe są również inne proporcje wielkości gametofitów i sporofitów. W brunatnicach Cukry gametofit jest mikroskopijny, a sporofit ma długość do 12 m. U większości alg gametofity i sporofity są roślinami niezależnymi. U wielu gatunków krasnorostów sporofity rosną na gametofitach, a u niektórych alg brunatnych gametofity rozwijają się wewnątrz plechy sporofitów.
Heteromorficzna zmiana form rozwojowych, gdy obserwuje się wyraźnie określone oddzielenie sporofitu od gametofitu, jest charakterystyczna dla wyżej zorganizowanych grup glonów. W tym przypadku jedna z form, najczęściej gametofit, jest mikroskopijna. Uważa się, że heteromorficzny cykl rozwojowy glonów powstał z izomorficznego. W taksonomii glonów duże znaczenie mają metody rozwoju gametofitu i sporofitu. Najbardziej złożone i różnorodne cykle rozwojowe, niespotykane u innych alg, są charakterystyczne dla krasnorostów.
Zmiana faz jądrowych.
Podczas procesu seksualnego, w wyniku połączenia gamet i ich jąder, liczba chromosomów w jądrze podwaja się. Na pewnym etapie cyklu rozwojowego, podczas mejozy, liczba chromosomów ulega zmniejszeniu, w wyniku czego powstałe jądra otrzymują pojedynczy zestaw chromosomów. Sporofity wielu glonów są diploidalne, a mejoza w ich cyklu rozwojowym zbiega się z momentem powstania zarodników, z których rozwijają się haploidalne gametosporofity lub gametofity. Ta mejoza nazywa się redukcja sporadyczna. Sporofity bardziej prymitywnych krasnorostów (rodzaje Kladofora, Ektokarpus i wiele innych) wraz z zarodnikami haploidalnymi tworzą zarodniki diploidalne, które ponownie rozwijają się w sporofity. Zarodniki pojawiające się na gametosporofitach służą do samoodnowy roślin matecznych. Sporofity i gametofity glonów na najwyższych etapach ewolucji ściśle się zmieniają, bez samoodnawiania.
U wielu glonów mejoza zachodzi w zygocie. Ta mejoza nazywa się redukcja zygotyczna i występuje w wielu gatunkach alg zielonych i charofitowych. W słodkowodnych algach volvox i ulothrix sporofit jest reprezentowany przez jednokomórkową zygotę, która wytwarza do 32 zoospor, których masa jest wielokrotnie większa niż gamety rodzicielskie, tj. zasadniczo obserwuje się sporadyczną redukcję.
Niektóre grupy glonów mają redukcja gametyczna, co jest charakterystyczne dla zwierząt, a nie organizmów roślinnych. U tych alg mejoza zachodzi podczas tworzenia gamet, podczas gdy pozostałe komórki plechy pozostają diploidalne. Taka zmiana faz jądrowych jest charakterystyczna dla okrzemek i morszczynów brunatnych, które są szeroko rozpowszechnione na całym świecie (do których należą najbardziej rozpowszechnione gatunki alg morskich) oraz wśród zielonych alg z dużego rodzaju Kladofora. Uważa się, że rozwój polegający na redukcji jądra gametycznego daje tym algom pewną przewagę nad innymi.
Jeśli podział redukcyjny zachodzi w zarodniach przed utworzeniem zarodników rozmnażania bezpłciowego (redukcja zarodnikowa), wówczas następuje przemiana pokoleń - diploidalny sporofit i haploidalny gametofit. Ten typ cyklu życia nazywa się haplobiont ze sporami zmniejszenie. Jest charakterystyczny dla niektórych zielonych alg, wielu brunatnic i czerwonych alg.
Wreszcie u kilku glonów mejoza zachodzi w komórkach wegetatywnych diploidalnej plechy (redukcja somatyczna), z których następnie rozwijają się haploidalne plechy. Taki cykl życia z redukcja somatyczna znany z czerwonych i zielonych alg.
Pytania kontrolne
Rodzaje żywienia algami.
Rodzaje plechy glonów.
Charakterystyka monadycznej struktury morfologicznej.
Charakterystyka budowy morfologicznej ryzopodiów. Rodzaje procesów cytoplazmatycznych.
Charakterystyka budowy morfologicznej palmelloidu.
Charakterystyka budowy morfologicznej kokosa.
Charakterystyka budowy morfologicznej trichali.
Charakterystyka heterotrychalnej struktury morfologicznej.
Charakterystyka budowy morfologicznej miąższu.
Charakterystyka syfonalnej struktury morfologicznej.
Charakterystyka budowy morfologicznej syfonokladalnej.
12. Rozmnażanie bezpłciowe. Rodzaje sporów.
13. Rodzaje wegetatywnego rozmnażania glonów.
14. Rodzaje rozmnażania płciowego glonów.
15. Czym różnią się sporofity i gametofity?
16. Czym jest heteromorficzna i izomorficzna zmiana pokoleń?
17. Zmiana faz jądrowych w cyklu życiowym glonów. Redukcja sporyczna, zygotyczna i gametyczna.
3. EKOLOGICZNE GRUPY ALG
Glony występują na całym świecie i występują w różnych biotopach wodnych, lądowych i glebowych. Znane są różne grupy ekologiczne: glony siedlisk wodnych, glony lądowe, glony glebowe, glony gorących źródeł, glony śniegu i lodu, glony źródeł hipersalinowych.
3.1. Glony siedlisk wodnych
3.1.1. Fitoplankton
Termin „fitoplankton” oznacza zbiór organizmów roślinnych unoszących się w słupie wody. Glony planktonowe są głównym, a w niektórych przypadkach jedynym producentem pierwotnej materii organicznej, na bazie której istnieje całe życie w zbiorniku wodnym. Produktywność fitoplanktonu zależy od zespołu różnych czynników.
Glony planktonowe żyją w różnych zbiornikach wodnych - od oceanu po kałużę. Ponadto większe zróżnicowanie warunków środowiskowych w zbiornikach wód śródlądowych w porównaniu z morzami determinuje także znacznie większe zróżnicowanie składu gatunkowego i kompleksów ekologicznych planktonu słodkowodnego.
Fitoplankton ekosystemów słodkowodnych charakteryzuje się wyraźnie określoną sezonowością. W zbiorniku o każdej porze roku dominuje jedna lub kilka grup glonów, a w okresach intensywnego rozwoju często dominuje tylko jeden gatunek. Zatem zimą pod lodem (zwłaszcza gdy jest pokryty śniegiem) fitoplankton jest bardzo ubogi lub prawie nieobecny z powodu braku światła. Rozwój wegetatywny glonów planktonowych jako zbiorowisk rozpoczyna się w marcu - kwietniu, kiedy poziom światła słonecznego staje się wystarczający do fotosyntezy glonów nawet pod lodem. W tym czasie pojawiają się dość liczne małe wiciowce - euglenofity, dinofity, złote, a także zimnolubne okrzemki. W okresie pękania lodu, zanim nastąpi rozwarstwienie temperaturowe, co zwykle ma miejsce, gdy górna warstwa wody zostanie podgrzana do temperatury 10–12°C, rozpoczyna się szybki rozwój zimnolubnego kompleksu okrzemek. Latem, gdy temperatura wody przekracza 15°C, obserwuje się maksymalną produktywność sinic, euglenoidów i zielonych alg. W zależności od typu troficznego i limnologicznego zbiornika, w tym czasie może nastąpić „rozkwit” wody, spowodowany rozwojem sinic i zielonych alg.
Jedną z istotnych cech fitoplanktonu słodkowodnego jest obfitość w nim tymczasowych glonów planktonowych. Szereg gatunków, które są powszechnie uważane za typowo planktonowe, występujące w stawach i jeziorach, przechodzi w swoim rozwoju fazę denną lub peryfitonową (przyłączoną do jakiegoś obiektu).
Fitoplankton morski składa się głównie z okrzemek i dinofitów. Spośród okrzemek szczególnie liczni są przedstawiciele rodzajów Chaetoceros, Rhizosolenia, Thalassiosira i kilka innych nieobecnych w planktonie słodkowodnym. Skład wiciowych form glonów dinofitów w fitoplanktonie morskim jest bardzo zróżnicowany. Przedstawiciele primnezjofitów są bardzo liczni w fitoplanktonie morskim, reprezentowani m.in świeże wody tylko kilka typów. Chociaż środowisko morskie jest stosunkowo jednorodne na dużych obszarach, podobnej jednorodności nie obserwuje się w rozmieszczeniu fitoplanktonu morskiego. Różnice w składzie gatunkowym i liczebności są często wyraźne nawet na stosunkowo małych obszarach wody morskiej, ale są szczególnie wyraźnie odzwierciedlone w wielkoskalowej strefie geograficznej występowania. Tutaj przejawia się ekologiczny wpływ głównych czynników środowiskowych: zasolenia wody, temperatury, światła i zawartości składników odżywczych.
Glony planktonowe mają zwykle specjalne przystosowania do życia w zawiesinie. Niektóre mają różnego rodzaju narośla i przydatki ciała - kolce, włosie, narośla rogowe, błony. Inne tworzą kolonie obficie wydzielające śluz. Jeszcze inne gromadzą w swoich ciałach substancje zwiększające ich pływalność (kropelki tłuszczu w okrzemkach, wakuole gazu w niebiesko-zielonych). Formacje te są znacznie bardziej rozwinięte w fitoplankteriach morskich niż słodkowodnych. Jednym z przystosowań do istniejących zawieszonych w słupie wody jest niewielki rozmiar ciała glonów planktonowych.
3.1.2. Fitobentos
Fitobentos odnosi się do zbioru organizmów roślinnych przystosowanych do istnienia w stanie przyłączonym lub niezwiązanym na dnie zbiorników wodnych oraz na różnych obiektach, organizmach żywych i martwych w wodzie.
O możliwości rozwoju glonów bentosowych w określonych siedliskach decydują zarówno czynniki abiotyczne, jak i biotyczne. Wśród czynników biotycznych znaczącą rolę odgrywa konkurencja z innymi glonami oraz obecność konsumentów. Prowadzi to do tego, że niektóre rodzaje glonów bentosowych nie rosną na wszystkich głębokościach i nie we wszystkich zbiornikach wodnych o odpowiednich warunkach świetlnych i hydrochemicznych. Światło jest szczególnie ważne dla wzrostu glonów bentosowych jako organizmów fotosyntetyzujących. Jednak na stopień jego wykorzystania wpływają inne czynniki środowiskowe: temperatura, zawartość substancji biogennych i biologicznie aktywnych, źródła tlenu i węgla nieorganicznego, a co najważniejsze, szybkość wnikania tych substancji do plechy, która zależy od stężenia substancji i prędkość ruchu wody. Z reguły miejsca o intensywnym ruchu wody charakteryzują się bujnym rozwojem glonów bentosowych.
Algi bentosowe rosnące w warunkach aktywnych ruch wody zyskują przewagę nad glonami rosnącymi w wodach osiadłych. Ten sam poziom fotosyntezy mogą osiągnąć organizmy fitobentosu w warunkach przepływu przy mniejszej ilości światła, co sprzyja wzrostowi większych plech. Ruch wody zapobiega ponadto osadzaniu się cząstek mułu na skałach i kamieniach, co utrudnia zawiązanie pąków glonów, sprzyja wzrostowi glonów dennych, wypłukując z powierzchni gleby zwierzęta żywiące się glonami. Wreszcie, chociaż podczas silnych prądów lub silnych fal, plechy glonów ulegają uszkodzeniu lub wyrwaniu z podłoża, ruch wody nadal nie zapobiega osiedlaniu się mikroskopijnych gatunków glonów lub mikroskopijnych stadiów glonów makrofitów.
Wpływ ruchu wody na rozwój glonów bentosowych jest szczególnie zauważalny w rzekach, potokach i potokach górskich. W zbiornikach tych występuje grupa organizmów bentosowych preferujących miejsca z silnymi prądami. W jeziorach, gdzie nie ma silnych prądów, ruch fal nabiera pierwszorzędnego znaczenia. W morzach fale mają również istotny wpływ na życie glonów bentosowych, w szczególności na ich pionowe rozmieszczenie.
W morzach północnych na rozmieszczenie i liczebność glonów bentosowych wpływają: lód. W zależności od grubości, ruchu i kępowania zarośla glonów mogą zostać zniszczone (wymazane) do głębokości kilku metrów. Dlatego na przykład w Arktyce wieloletnie brunatnice ( Fukus, Laminaria) najłatwiej znaleźć w pobliżu brzegu, wśród głazów i półek skalnych utrudniających ruch lodu.
Na życie glonów bentosowych wpływa wiele czynników temperatura. Wraz z innymi czynnikami determinuje tempo ich wzrostu, tempo i kierunek rozwoju, moment kształtowania się narządów rozrodczych oraz strefę geograficzną rozmieszczenia.
Intensywnemu rozwojowi glonów sprzyja również umiarkowana zawartość wody. składniki odżywcze. W wodach słodkich takie warunki powstają w płytkich stawach, w strefie przybrzeżnej jezior, w rozlewiskach rzecznych, w morzach - w małych zatokach.
Jeśli w takich miejscach jest wystarczające oświetlenie, twarde gleby i słaby ruch wody, wówczas powstają optymalne warunki do życia fitobentosu. W przypadku braku ruchu wody i jej niedostatecznego wzbogacenia w składniki odżywcze glony bentosowe słabo rosną. Takie warunki panują w zatokach skalistych o dużym spadku dna i znacznych głębokościach w centrum, gdyż składniki odżywcze z osadów dennych nie są przenoszone do górnych poziomów. Ponadto w takich siedliskach może nie występować makroskopowe wodorosty, które służą jako substrat dla wielu małych form glonów bentosowych.
Źródłem składników odżywczych w wodzie są spływy przybrzeżne i osady denne. Rola tych ostatnich jest szczególnie duża jako akumulatorów pozostałości organicznych. W osadach dennych w wyniku żywotnej działalności bakterii i grzybów następuje mineralizacja pozostałości organicznych; złożone substancje organiczne przekształcają się w proste związki nieorganiczne, które mogą być wykorzystane przez rośliny fotosyntetyzujące.
Oprócz światła, ruchu wody, temperatury i zawartości składników odżywczych, wzrost glonów bentosowych zależy od obecność roślinożernych zwierząt wodnych – jeżowce, ślimaki, skorupiaki, ryby. Jest to szczególnie widoczne w zaroślach glonów wodorostów, które są duże. W morzach tropikalnych w niektórych miejscach ryby całkowicie zjadają zielone, brązowe i czerwone glony z miękką plechą. Ślimaki pełzające po dnie zjadają mikroskopijne glony i małe sadzonki gatunków makroskopowych.
Dominującymi glonami dennymi w kontynentalnych zbiornikach wodnych są okrzemki oraz glony nitkowate zielone, niebieskozielone i żółtozielone, przyczepione lub nieprzytwierdzone do podłoża.
Głównymi glonami bentosowymi mórz i oceanów są brązowe i czerwone, czasem zielone, makroskopowe formy plechowe. Wszystkie mogą być porośnięte małymi okrzemkami, niebieskozielonymi i innymi glonami.
W zależności od miejsca wzrostu wśród glonów bentosowych wyróżnia się następujące grupy ekologiczne:
epilitów– rosną na powierzchni twardego podłoża (skały, kamienie);
epipelity– zasiedlają powierzchnię gleb sypkich (piasek, muł);
narost– epizoity– żyją na powierzchni roślin/zwierząt;
endofity– endozoity lub endosymbionty– żyją w ciele roślin/zwierząt, ale odżywiają się samodzielnie (mają chloroplasty i dokonują fotosyntezy);
endolity– żyją w podłożu wapiennym (skały, muszle mięczaków, skorupiaków).
Czasami izolowana jest grupa organizmów zanieczyszczanie, Lub peryfiton. Organizmy zaliczane do tej grupy żyją na obiektach, które w większości poruszają się lub unoszą pod wodą. Ponadto są one usuwane z dna i wystawiane na działanie innych warunków świetlnych, pokarmowych i temperaturowych niż organizmy rzeczywiście żyjące na dnie.
Skład porostowy obejmuje mikroalgi i glony makrofitowe. Mikroskopijne algi (niebieskozielone i okrzemki) tworzą śluzową warstwę bakterio-glono-detrytyczną na podłożu wprowadzonym do środowiska wodnego. Następnie makroalgi (czerwone, brązowe i zielone) osadzają się na pierwotnym mikrofilmie wraz ze zwierzętami. Stwarza to poważną ingerencję w działalność gospodarczą człowieka. Z powodu zanieczyszczeń spada prędkość statków i wydajność urządzeń hydroakustycznych, wzrasta zużycie paliwa, a konstrukcje podwodne stają się cięższe i korodują. Ponadto śluzowaty film powstały w wyniku zanieczyszczania może zakłócać pracę rur wodociągowych, zatykać otwory poboru wody i rurociągów oraz zakłócać procesy wymiany ciepła w agregatach chłodniczych.
Przyłączone organizmy porostowe żyjące na konstrukcjach podwodnych w strefie pływów i na głębokościach do 1 m są zwykle eliminowane zimą w wyniku długotrwałego suszenia i ścierania przez lód. Dlatego co roku w okresie wiosenno-letnim tworzą się tu zbiorowiska porostowe, charakterystyczne dla pionierskiego etapu sukcesji biologicznej. Dominującym gatunkiem takich zbiorowisk, obok pąkli i mięczaków, są często glony makrofitowe. W strefie sublitoralnej budowli podwodnych – od głębokości 0,7-0,9 m do ich podstawy (6-12 m) – rozwija się wieloletnie osadzanie. W jego składzie dominują algi brunatne z rodzaju Sacharyna I Kostaryka. Biomasa tych dużych glonów w umiarkowanych szerokościach geograficznych może być bardzo znacząca i wynosić dziesiątki kilogramów na metr kwadratowy.
W powietrzu mogą również występować glony zanieczyszczające ( aerofiton). Wśród nich dominują glony zielone i niebieskozielone. W pewnych warunkach glony aerofitonowe mogą uszkodzić materiały przemysłowe i budowlane, zabytki architektury, obrazy itp., jeśli nie są chronione toksycznymi powłokami. Przyczyną uszkodzeń są produkty przemiany materii czynników zanieczyszczających, głównie kwasów organicznych. Glony Aerophyton są szczególnie powszechne w wilgotnych tropikach, gdzie jest wystarczająco dużo ciepła, wilgoci i kurzu pochodzenia organicznego, co jest pożywką dla ich rozwoju. Uszkodzenia biologiczne spowodowane przez nie mogą być znaczące.
Epility. Do tej grupy zaliczają się glony przyczepione. Zasiedlają powierzchnię kamieni, tworząc skorupopodobne powłoki lub płaskie poduszki lub mają specjalne narządy przyczepne - ryzoidy. Intensywny rozwój epilitów obserwuje się w zbiornikach o twardym dnie i szybko płynącej wodzie. Typowymi epilitami są przedstawiciele złocistych alg z rodzaju Hydrorus, brunatnice z rodzaju Saccharina, Kelp, Costaria itd.
Epipelity. Luźne glony rozprzestrzeniają się po dnie, wiążąc i wzmacniając podłoże. Często reprezentowane są przez okrzemki, aureusy, euglenoidy, kryptofity i dinofity swobodnie pełzające po podłożu. Narządami przyczepowymi epipelitów są czasami krótkie ryzoidy, które nie mogą się głęboko zakorzenić. Na błotnistym dnie dobrze rozwijają się jedynie glony charofitowe z długimi ryzoidami.
Zwykle narządami mocowania naskórków i naskórków są specjalne formacje - podeszwa, noga, stopa, błona śluzowa lub poduszka śluzowa, poduszka itp.
Epifity/epizoity. Glony wykorzystują organizmy żywe jako substrat. Epizoity to algi osiedlające się na zwierzętach. Na powierzchni muszli mięczaków znajdują się małe zielone ( Edogonium, Cladophora, Ulva) i czerwony ( Gelidium, Palmaria,) wodorosty; na gąbkach - zielonych, niebiesko-zielonych i okrzemkach. Epizoity żyją na skorupiakach, wrotkach, rzadziej na wodnych nieowadach lub larwach, robakach, a nawet większych zwierzętach. Epizoity obejmują gatunki glonów zielonych i charofitowych z rodzajów Chlorangiella, Charatiochloris, Korzhikoviella, Chlorangiopsis itp. Większość epizoitów nie może istnieć w izolacji od podłoża. Glony zwykle giną na martwych zwierzętach lub na ich muszlach zrzucanych podczas linienia.
Epifity to algi żyjące na roślinach. Pomiędzy rośliną substratową (bazyfitem) a rośliną epifitową powstają krótkotrwałe połączenia. Złożone i interesujące zjawisko epifityzmu jest wciąż słabo poznane. Często zdarzają się przypadki podwójnego, a nawet potrójnego epifityzmu, gdy jednych glonów osiada na innych, jest więcej duże formy, same są podłożem dla innych, mniejszych lub mikroskopijnych gatunków. Czasami dla rozwoju epifitów ważny jest stan fizjologiczny rośliny substratowej. Liczba epifitów z reguły wzrasta wraz ze starzeniem się glonów bazfitowych. Przykładowo największe bogactwo gatunkowe epifitycznych glonów aedogonia obserwuje się na martwych roślinach wodnych ( Manna, trzcina, turzyca).
Endofity/endozoity lub endosymbionty
Endosymbionty lub symbionty wewnątrzkomórkowe – algi żyjące w tkankach lub komórkach innych organizmów (zwierząt bezkręgowych lub glonów). Tworzą rodzaj grupy ekologicznej. Symbionty wewnątrzkomórkowe nie tracą zdolności do fotosyntezy i rozmnażania się wewnątrz komórek gospodarza. Endosymbiontami mogą być różne glony, ale najliczniejsze są endosymbiozy jednokomórkowych zielonych i żółto-zielonych alg ze zwierzętami jednokomórkowymi. Glony uczestniczące w takich symbiozach nazywane są odpowiednio zoochlorella I zooksantele. Zielone i żółto-zielone algi tworzą endosymbiozy z organizmami wielokomórkowymi: gąbkami, hydrami itp. Endosymbiozy niebiesko-zielonych alg z pierwotniakami nazywane są syncyjanozy. Często inne rodzaje sinic mogą osadzać się w śluzie niektórych niebiesko-zielonych gatunków. Zwykle korzystają z gotowych związków organicznych, które powstają w dużych ilościach podczas rozkładu śluzu kolonii rośliny żywicielskiej i intensywnie się rozmnażają.
Najczęstszymi endofitami są przedstawiciele goldenów (gatunki z rodzaju Chromulina, Myxochloris) i zielony (rodzaj Chlorochitrium, Chlamydomyks) glony zasiedlające ciało rzęsy i mchów torfowców. Rodzaj zielonych alg Karteria osadza się w komórkach naskórka robaka rzęskowego Skręcony, jeden gatunek z rodzaju Chlorella– w wakuolach pierwotniaków i gatunków z rodzaju Chlorokok– w komórkach alg kryptofitowych Cyjanofora.
3.1.3. Glony ekstremalnych ekosystemów wodnych
Algi gorącej wiosny. Nazywa się glony rosnące w temperaturach 35–85°C termofilny. Często wysokie temperatury otoczenia łączą się z dużą zawartością soli mineralnych lub substancji organicznych (silnie zanieczyszczone gorące ścieki z fabryk, fabryk, elektrowni lub elektrownie jądrowe). Typowymi mieszkańcami gorących wód są sinice oraz w mniejszym stopniu okrzemki i algi zielone.
Algi śniegu i lodu. Nazywa się glony rosnące na powierzchni lodu i śniegu kriofilny. Rozwijając się w dużych ilościach, mogą powodować zielone, żółte, niebieskie, czerwone, brązowe lub czarne „kwitnienie” śniegu lub lodu. Wśród glonów kriofilnych dominują algi zielone, niebieskozielone i okrzemkowe. Tylko kilka z tych glonów ma stadia uśpienia, większość nie posiada żadnych specjalnych adaptacji morfologicznych pozwalających tolerować niskie temperatury.
Glony ze zbiorników słonowodnych dostałem to imię halofilny Lub halobionty. Takie glony rosną przy wysokich stężeniach soli w wodzie, sięgających 285 g/l w jeziorach z przewagą soli kuchennej i 347 g/l w jeziorach glauberskich. Wraz ze wzrostem zasolenia maleje liczba gatunków glonów; Tylko nieliczne z nich tolerują bardzo duże zasolenie. W wodach zasolonych (hiperhalinowych) dominują jednokomórkowe, ruchliwe algi zielone - hiperhaloby, których komórki nie mają błony i są otoczone plazmalemmą ( Asteromonas, Pedinomonas). Wyróżniają się zwiększoną zawartością chlorku sodu w protoplazmie, wysokim wewnątrzkomórkowym ciśnieniem osmotycznym oraz akumulacją karotenoidów i gliceryny w komórkach. W niektórych zasiedlonych zbiornikach takie glony mogą powodować czerwone lub zielone „zakwitanie” wody. Dno zbiorników hiperhalinowych jest czasami całkowicie pokryte sinicami; Wśród nich dominują gatunki z rodzajów oscylatory, Spirulina itp. Wraz ze spadkiem zasolenia obserwuje się wzrost różnorodności gatunkowej glonów: oprócz niebiesko-zielonych alg pojawiają się okrzemki (gatunki z rodzaju Navicula, Nietzschego).
3.2. Glony siedlisk niewodnych
Chociaż głównym środowiskiem życia większości glonów jest woda, to ze względu na eurytopowy charakter tej grupy organizmów, z powodzeniem kolonizują one różnorodne siedliska pozawodne. W obecności przynajmniej okresowej wilgoci wiele z nich rozwija się na różnych obiektach gruntowych - skałach, korze drzew, płotach itp. Korzystnym siedliskiem dla glonów jest gleba. Ponadto znane są także zbiorowiska glonów endolitycznych, których głównym środowiskiem życia jest otaczające je podłoże wapienne.
Zbiorowiska utworzone przez glony w siedliskach pozawodnych dzielą się na aerofilne, edafilne i litofilne.
3.2.1. Algi aerofilne
Głównym środowiskiem życia glonów aerofilnych jest otaczające je powietrze. Typowe siedliska to powierzchnia różnych twardych podłoży pozaglebowych (skały, kamienie, kora drzew, ściany domów itp.). W zależności od stopnia zawilgocenia dzieli się je na dwie grupy: powietrzne i wodno-powietrzne. Algi powietrzneŻyją w warunkach jedynie wilgoci atmosferycznej i doświadczają ciągłych zmian wilgotności i suszenia. Algi wodno-powietrzne są narażone na ciągłe nawadnianie wodą (pod strumieniem wodospadów, w strefie surfingu itp.).
Warunki życia tych glonów są bardzo specyficzne i charakteryzują się przede wszystkim częstą zmianą dwóch czynników - wilgotności i temperatury. Glony żyjące w warunkach wyłącznie wilgoci atmosferycznej często zmuszone są do przejścia ze stanu nadmiernej wilgoci (na przykład po ulewie) do stanu minimalnej wilgoci w okresach suchych, kiedy wysychają tak bardzo, że można je zmielić na proszek. Glony wodne żyją w warunkach stosunkowo stałej wilgotności, jednak i u nich występują znaczne wahania tego czynnika. Na przykład glony żyjące na skałach nawadnianych przez strumienie wodospadów czas letni Kiedy przepływ znacznie się zmniejsza, następuje u nich deficyt wilgoci. Społeczności aerofilne są również podatne na ciągłe wahania temperatury. W ciągu dnia nagrzewają się bardzo, w nocy schładzają, a zimą zamarzają. To prawda, że niektóre glony aerofilne żyją w dość stałych warunkach (na ścianach szklarni). Ale ogólnie rzecz biorąc, stosunkowo niewiele glonów, reprezentowanych przez mikroskopijne jednokomórkowe, kolonialne i nitkowate formy niebiesko-zielonych i zielonych alg oraz, w znacznie mniejszym stopniu, okrzemki, przystosowały się do niesprzyjających warunków istnienia tej grupy. Wśród krasnorostów tego rodzaju znane są także formy aerofilne Porfiryd itd.; można je znaleźć na kamieniach i starych ścianach szklarni. Liczba gatunków występujących w grupach aerofilnych zbliża się do 300. Kiedy glony aerofilne rozwijają się w masowych ilościach, zwykle przybierają postać sypkich lub śluzowatych osadów, mas przypominających filc, miękkich lub twardych błon i skorup.
Na korze drzew zwykłymi osadnikami są wszechobecne zielone algi z rodzajów Pleurokoki, Chlorella, Chlorokok. Na drzewach znacznie rzadziej spotykane są sinice i okrzemki. Istnieją dowody na to, że na nagonasiennych rosną głównie glony zielone.
Skład systematyczny grup glonów żyjących na powierzchni odsłoniętych skał jest inny. Rozwijają się tu okrzemki i niektóre, przeważnie jednokomórkowe, zielone algi, ale w tych siedliskach najliczniej występują przedstawiciele sinic. Glony i towarzyszące im bakterie tworzą „opaleniznę górską” (filmy i skorupy skalne) na skałach krystalicznych różnych pasm górskich. W gruzie gromadzącym się w zagłębieniach skalnych żyją zazwyczaj jednokomórkowe glony zielone i sinice. Szczególnie obfite porosty glonów występują na powierzchni mokrych skał. Tworzą filmy i narośla o różnych kolorach. Z reguły żyją tu gatunki wyposażone w grube błony śluzowe. W zależności od natężenia światła śluz może zabarwić się mniej lub bardziej intensywnie, co decyduje o kolorze narośli. Mogą być jasnozielone, złote, brązowe, fioletowe, prawie czarne, w zależności od gatunku, który je tworzy. Szczególnie charakterystyczni dla nawodnionych skał są przedstawiciele sinic, m.in. gatunki z rodzaju Gleocapsa, Tolipotrix, Spirogyra itp. W naroślach na wilgotnych skałach można spotkać także okrzemki z rodzaju Frustulia, Achnantes itd.
Zatem zbiorowiska glonów aerofilnych są bardzo zróżnicowane i powstają zarówno w warunkach całkowicie sprzyjających, jak i ekstremalnych. Przystosowania zewnętrzne i wewnętrzne do tego stylu życia są zróżnicowane i podobne do przystosowań glonów glebowych, zwłaszcza tych rozwijających się na powierzchni gleby.
3.2.2. Algi edafilne
Głównym środowiskiem życia glonów edafofilnych jest gleba. Typowe siedliska to powierzchnia i grubość warstwy gleby, która ma fizyczny i chemiczny wpływ na bionty. W zależności od lokalizacji glonów i ich trybu życia, w obrębie tego typu wyróżnia się trzy grupy: algi lądowe, masowo rozwijający się na powierzchni gleby w warunkach wilgoci atmosferycznej; wodno-lądowe wodorost, rozwijający się masowo na powierzchni gleby, stale nasyconej wodą; glony glebowe, zasiedlający grubość warstwy gleby.
Gleba jako biotop przypomina siedliska wodne i powietrzne: zawiera powietrze, ale jest nasycona parą wodną, co zapewnia oddychanie powietrzem atmosferycznym bez zagrożenia wysychaniem. Nieruchomość. Tym, co zasadniczo odróżnia glebę od wymienionych biotopów, jest jej nieprzezroczystość. Czynnik ten ma decydujący wpływ na rozwój glonów. Jednak w grubości gleby, gdzie światło nie przenika, żywe glony występują na głębokości do 2 m na terenach dziewiczych i do 2,7 m na gruntach ornych. Wyjaśnia to zdolność niektórych glonów do przemieszczania się odżywianie heterotroficzne.
Niewielka liczba glonów znajduje się w głębokich warstwach gleby. Aby zachować żywotność, glony glebowe muszą wykazywać się odpornością na niestabilną wilgotność, nagłe wahania temperatury i silne nasłonecznienie. Właściwości te zapewnia szereg cech morfologicznych i fizjologicznych. Na przykład zauważono, że glony glebowe są stosunkowo małe w porównaniu z odpowiadającymi im formami wodnymi tego samego gatunku. W miarę zmniejszania się rozmiaru komórek wzrasta ich zdolność zatrzymywania wody i odporność na suszę. Ważną rolę w odporności glonów glebowych na suszę odgrywa zdolność wytwarzania obfitego śluzu – śluzowatych kolonii, osłon i otoczek składających się z hydrofilowych polisacharydów. Dzięki obecności śluzu glony szybko wchłaniają wodę po zwilżeniu i magazynują ją, spowalniając wysychanie. Glony glebowe przechowywane w stanie suchym na powietrzu w próbkach gleby wykazują niesamowitą żywotność. Jeśli taka gleba zostanie położona pożywka, wówczas będzie można zaobserwować rozwój glonów.
Charakterystyczną cechą glonów glebowych jest „efemeryczność” okresu wegetacyjnego - zdolność szybkiego przejścia ze stanu spoczynku do aktywnego życia i odwrotnie. Są także w stanie wytrzymać wahania temperatury w bardzo szerokim zakresie: od -200 do +84°C. Glony glebowe (głównie niebieskozielone) są odporne na promieniowanie ultrafioletowe i radioaktywne.
Zdecydowana większość glonów glebowych to formy mikroskopijne, ale często można je zobaczyć na powierzchni gleby gołym okiem. Masowy rozwój takich glonów może powodować zazielenianie zboczy wąwozów i poboczy leśnych dróg.
Pod względem składu systematycznego glony glebowe są dość zróżnicowane. Wśród nich niebiesko-zielone i zielone algi są reprezentowane w przybliżeniu w równych proporcjach. W glebach mniej zróżnicowane są żółtozielone algi i okrzemki.
3.2.3. Algi litofilne
Głównym środowiskiem życia glonów litofilnych jest otaczające je nieprzezroczyste, gęste podłoże wapienne. Typowe siedliska znajdują się głęboko w twardych skałach o określonym składzie chemicznym, otoczone powietrzem lub zanurzone w wodzie. Wyróżnia się dwie grupy glonów litofilnych: wiercenie glonów, które aktywnie wnikają w podłoże wapienne; algi tworzące tufy, odkładając wokół ciała wapno i żyjąc w peryferyjnych warstwach środowiska, które deponują, w granicach dostępnych dla wody i światła. W miarę gromadzenia się osadu zanika on.
Pytania kontrolne
1. Opisać główne grupy ekologiczne glonów występujących w siedliskach wodnych: fitoplankton i fitobentos.
2. Różnice pomiędzy fitolaktonem słodkowodnym i morskim. Przedstawiciele fitoplanktonu morskiego i słodkowodnego.
3. Morfologiczne adaptacje glonów do planktonicznego trybu życia.
4. Sezonowe zmiany wskaźników jakościowych i ilościowych fitoplanktonu słodkowodnego.
5. Różnice pomiędzy fitobentosem słodkowodnym i morskim. Skład systematyczny fitobentosu morskiego i słodkowodnego.
6. Grupy ekologiczne fitobentosu w zależności od podłoża (epility, epipelity, epifity, endofity).
7. Co to jest zanieczyszczenie? Jakie glony mogą wchodzić w skład tej grupy ekologicznej?
8. Algi aerofilne. Adaptacje do ekstremalnych warunków środowiskowych. Skład systematyczny glonów powietrznych.
9. Algi edafilne. Adaptacje do warunków środowiskowych. Skład systematyczny glonów glebowych.
10. Glony litofilne.
4. ROLA ALG W PRZYRODIE I ZNACZENIE PRAKTYCZNE
Rola glonów w ekosystemach naturalnych. W biocenozach wodnych glony pełnią rolę producentów. Wykorzystując energię świetlną, są w stanie syntetyzować substancje organiczne z nieorganicznych. Według datowania radiowęglowego średnia pierwotna produkcja oceanów w wyniku żywotnej aktywności glonów wynosi 550 kg węgla na 1 hektar rocznie. Całkowita wartość jego pierwotnej produkcji wynosi 550,2 miliardów ton (w surowej biomasie) rocznie, a według naukowców udział glonów w całkowitej produkcji węgla organicznego na naszej planecie waha się od 26 do 90%. Glony odgrywają ważną rolę w obiegu azotu. Potrafią wykorzystywać zarówno organiczne (mocznik, aminokwasy, amidy), jak i nieorganiczne (jony amonowe i azotanowe) źródła azotu. Wyjątkową grupę stanowią sinice, które potrafią wiązać gazowy azot, przekształcając go w związki dostępne dla innych roślin.
Algi - producenci tlenu. Glony w procesie swojej aktywności życiowej wydzielają tlen niezbędny do oddychania organizmów wodnych. W środowisku wodnym (szczególnie w morzach i oceanach) glony są praktycznie jedynymi producentami wolnego tlenu. Ponadto odgrywają dużą rolę w ogólnym bilansie tlenowym na Ziemi, ponieważ oceany służą jako główny regulator bilansu tlenowego w atmosferze ziemskiej.
Glony są pożywką dla innych organizmów wodnych. Tworząc podwodne lasy, glony makrofitowe tworzą wysoce produktywne ekosystemy, które zapewniają pożywienie, schronienie i ochronę wielu innym żywym organizmom. Ustalono, że słup wody o objętości 5 litrów zawierał jeden okaz brunatnic Cystoseira liczy aż 60 tysięcy osobników różnych bezkręgowców, m.in. mięczaków, roztoczy i skorupiaków.
Algi - pionierzy roślinności. Glony lądowe mogą osiedlać się na nagich skałach, piasku i innych jałowych miejscach. Po ich śmierci powstaje pierwsza warstwa przyszłej gleby. Glony glebowe biorą udział w procesach kształtowania struktury i żyzności gleby.
Glony jako czynnik geologiczny. Rozwój glonów w poprzednich epokach geologicznych doprowadził do powstania wielu skał. Razem ze zwierzętami glony brały udział w tworzeniu raf w oceanach. Osiedlone bliżej powierzchni wody utworzyły grzbiety tych raf. Struktury rafowe krasnorostów znane są na Krymie jako szczyty Yayla i inne.Niebiesko-zielone algi brały udział w tworzeniu wapieni stromatolitowych, glony chara - w tworzeniu wapieni charocytowych (podobne złoża znaleziono w Tuwie). Kokolitofory biorą udział w powstawaniu skał kredowych (skały kredowe składają się w 95% z pozostałości muszli tych glonów). Masowe nagromadzenie muszli okrzemek doprowadziło do powstania diatomitu (mąki górskiej), którego duże złoża odkryto na Terytorium Primorskim, Uralu i Sachalinie. Glony były materiałem wyjściowym do ciekłych i stałych związków ropopochodnych - sapropelów, gorących łupków, węgla.
Obecnie w niektórych regionach odnotowano aktywną aktywność glonów w tworzeniu skał. Absorbują węglan wapnia i tworzą produkty zmineralizowane. Procesy te są szczególnie aktywne w wodach tropikalnych o wysokich temperaturach i niskim ciśnieniu parcjalnym.
Największe znaczenie w niszczeniu skał mają glony nudne. Powoli i trwale rozluźniają podłoża wapienne, czyniąc je podatnymi na wietrzenie, kruszenie i erozję.
Symbiotyczne relacje z innymi organizmami. Glony tworzą kilka ważnych symbioz. Po pierwsze, tworzą porosty z grzybami, po drugie, jako zooksanty, żyją razem z niektórymi bezkręgowcami, takimi jak gąbki, żółwie i koralowce rafowe. Wiele cyjanofitów tworzy skojarzenia z roślinami wyższymi.
Glony mają ogromne znaczenie praktyczne w życiu codziennym i działalności gospodarczej człowieka, przynosząc zarówno korzyści, jak i szkody. Duże, głównie wodorosty są znane od czasów starożytnych i od dawna wykorzystywane w rolnictwie człowieka.
Algi jako produkt spożywczy. Ludzie jedzą głównie wodorosty, z których szczególnie chętnie korzystają mieszkańcy Azji Południowo-Wschodniej i wysp. Pacyfik. W Chinach stosowanie alg w diecie znane jest już od IX wieku p.n.e. mi. Wśród glonów makrofitowych (wielokomórkowych zielonych, brunatnych i czerwonych) nie ma gatunków trujących, gdyż nie zawierają alkaloidów – substancji o działaniu narkotycznym i trującym. W pożywieniu wykorzystuje się około 160 gatunków różne algi. Pod względem wartości odżywczych glony nie ustępują wielu uprawom rolnym. Zawierają duży procent białka, węglowodanów i tłuszczów. Algi są doskonałym źródłem witamin C, A, D, z grupy B, ryboflawiny, kwasu pantotenowego i foliowego oraz mikroelementów.
Spośród mikroskopijnych alg jako pokarm wykorzystuje się niebiesko-zielone gatunki lądowe z rodzaju. Nostok, które służą jako żywność w Chinach i Ameryce Południowej. W Japonii je się chleb jęczmienny „tengu” – są to grube warstwy gęstej galaretowatej masy na zboczach niektórych wulkanów, składające się z niebieskozielonych alg z rodzaju Gleocapsa, Geoteke, Microcystis z domieszką bakterii. Spirulina używany przez Azteków już w XVI wieku do przygotowywania ciast z suszonych wodorostów, a ludność regionu Jeziora Czad w Ameryka północna Z tych alg nadal wytwarza się produkt zwany dikhe. Spirulina zawiera duża liczba białka i jest szeroko uprawiany w wielu krajach.
Algi jako nawóz. Glony zawierają wystarczającą ilość substancji organicznych i mineralnych, dlatego od dawna są stosowane jako nawozy. Zaletą takich nawozów jest to, że nie zawierają nasion chwastów i zarodników grzybów fitopatogennych, a zawartość potasu przewyższa prawie wszystkie stosowane rodzaje nawozów. Niebiesko-zielone algi wiążące azot są powszechnie stosowane na polach ryżowych zamiast nawozów azotowych. Wykazano, że nawozy z alg mogą zwiększać kiełkowanie nasion, plon i odporność na choroby.
Właściwości lecznicze alg. Algi są szeroko stosowane w medycynie ludowej jako środek przeciwrobaczy oraz do leczenia wielu chorób, takich jak wole, zaburzenia nerwowe, stwardnienie rozsiane, reumatyzm, krzywica itp. Wykazano, że ekstrakty z wielu rodzajów alg zawierają substancje antybiotykowe i mogą obniżać ciśnienie krwi. Wyciągi z Sargassum, wodorosty i Saharina w eksperymentach na myszach hamowały wzrost mięsaka i komórek białaczkowych. W USA i Japonii uzyskano od nich leki, które pomagają usuwać radionuklidy z organizmu. Wydajność takich sorbentów sięga 90–95%.
Algi jako źródło surowców przemysłowych. Od ubiegłego wieku algi są wykorzystywane do produkcji sody i jodu. Obecnie z alg pozyskuje się kwas alginowy i jego sole – alginiany, a także karageny i agar.
Alkohol mannitolowy otrzymywany jest z alg brunatnych – niezbędny surowiec do celów farmakologicznych i Przemysł spożywczy w produkcji leków i artykułów spożywczych dla diabetyków.
Negatywna rola glonów. Szereg glonów (niebieskozielonych, dinofitowych, złotych, zielonych) wytwarza toksyny, które mogą powodować różne choroby u zwierząt, roślin i ludzi, a niektóre z nich mogą być śmiertelne. Wśród glonów dinofitów powodujących „czerwone przypływy” na rozległych obszarach morskich gatunki z tego rodzaju są toksyczne Gymnodinium, Noctiluca, Amphidinium itp. Najwięcej gatunków toksycznych zidentyfikowano wśród sinic. Działanie toksyn sinicowych jest kilkukrotnie silniejsze niż trucizn takich jak kurara i botulina. Toksyczność glonów objawia się masową śmiercią organizmów wodnych, ptactwa wodnego, zatruciami i innymi chorobami ludzi, które powstają w wyniku wdychania, spożycia wody, spożycia skorupiaków, ryb itp.
Przy silnym rozwoju - „kwitnięciu zbiorników wodnych”, niektóre glony (złote, żółto-zielone, niebiesko-zielone) mogą nadawać wodzie nieprzyjemny zapach i smak, czyniąc ją niezdatną do picia.
Nadmierny rozwój glonów może uniemożliwić przepływ wody przez filtry struktur ujęcia wody. Wiadomo, że zanieczyszczanie statków glonami znacznie zwiększa koszty operacyjne. Makrofity mogą przyczyniać się do korozji materiałów na platformach wiertniczych i innych podwodnych konstrukcjach morskich.
Problem zanieczyszczenia jest prawdopodobnie najstarszym problemem eksploracji oceanów. Każdy przedmiot, który wejdzie w kontakt ze środowiskiem morskim, wkrótce zostaje pokryty masą przyczepionych do niego organizmów: zwierząt i glonów. Całkowita powierzchnia zatopionych podłoży wynosi około 20% powierzchni górnej półki. Całkowita biomasa zanieczyszczeń wynosi miliony ton, a szkody z nią związane sięgają miliardów dolarów (Zwiagincew, 2005). W aspekcie biologicznym jest to naturalny proces stanowiący integralną część życia hydrosfery. Jednocześnie zjawisko porastania podsunęło człowiekowi pomysł hodowli szeregu cennych gatunków mięczaków w rybołówstwie morskim na skalę przemysłową ( Ostrygi, małże, przegrzebki, małże perłowe) i algi ( Sacharyny, Porfir, Gracilaria, Euchema itd.). Glony są pionierskimi organizmami porastającymi. Mikroalgi wraz z bakteriami tworzą pierwotny mikrofilm na powierzchni sztucznych podłoży dodawanych do wody, który służy jako podłoże do sedymentacji innych hydrobiontów. Makroglony wraz ze skorupiakami, mięczakami, hydroidami i innymi zwierzętami często tworzą początkowe stadia wieloletnich zbiorowisk porastających.
Pytania kontrolne
1. Rola glonów w zwiększaniu żyzności gleby.
2. Rola glonów w ekosystemach wodnych.
3. Rola glonów w ekosystemach lądowych.
4. Znaczenie glonów w procesach geologicznych.
5. Wartość odżywcza i biologiczna alg. Jakie wodorosty można jeść?
6. Właściwości lecznicze alg.
7. Dlaczego rozwój glonów złocistych i żółto-zielonych w zbiornikach jest niepożądany? Co to jest „kwitnienie” zbiorników wodnych?
8. Glony powodujące zatrucie zwierząt i ludzi.
9. Zjawisko zanieczyszczania. Rola glonów w zbiorowiskach zanieczyszczających.
5. WSPÓŁCZESNA SYSTEMATYKA ALG
Klasyfikacja organizmów żywych zajmuje umysły ludzi od czasów Arystotelesa. Szwedzki botanik Carl Linnaeus jako pierwszy w XVIII wieku zastosował nazwę Algi do grupy roślin i zaczął fykologia(z greckiego fikos – algi i logo – nauczanie) jako nauka. Wśród glonów Linneusz wyróżnił tylko cztery rodzaje: Chara, Fucus, Ulva i Conferva. W XIX wieku opisano większość (kilka tysięcy) współczesnych rodzajów glonów. Duża liczba nowych rodzajów wymagała pogrupowania ich w taksony wyższego rzędu. Początkowe próby klasyfikacji opierały się wyłącznie na zewnętrznych cechach plechy. Pierwszym, który zaproponował kolor plechy glonów jako cechę podstawową przy ustalaniu dużych grup taksonomicznych, czyli megataksonów, był angielski naukowiec W. Harvey (Harvey, 1836). Wyróżnił duże serie: Chlorospermeae – algi zielone, Melanospermeae – algi brunatne i Rhodospermeae – algi czerwone. Później przemianowano je odpowiednio na Chlorophyceae, Phaeophyceae i Rhodophyceae.
Podwaliny współczesnej taksonomii glonów założył w pierwszej połowie XX wieku czeski uczony A. Pascher. Ustalił 10 klas glonów: niebieskozielone, czerwone, zielone, złociste, żółto-zielone, okrzemkowe, brunatne, Dinophyte, Cryptophyte i Euglenaceae. Każda klasa charakteryzuje się specyficznym zestawem pigmentów, produktów rezerwowych i budową wici. Te ciągłe różnice między dużymi taksonami skłoniły nas do uznania ich za niezależne grupy filogenetyczne, niepowiązane i porzucenia koncepcji glonów - Algi jako specyficznej jednostki taksonomicznej.
Zatem słowo „algi” w rzeczywistości nie jest koncepcją systematyczną, ale ekologiczną i dosłownie oznacza „to, co rośnie w wodzie”. Glony to rośliny niższe, zawierające zdecydowaną większość chlorofilu, zdolne do odżywiania się fototroficznego i żyjące głównie w wodzie. Wszystkie glony, z wyjątkiem charofitów, w przeciwieństwie do roślin wyższych, nie mają wielokomórkowych narządów rozrodczych z osłoną sterylnych komórek.
Nowoczesne systemy różnią się głównie liczbą i objętością megataksonów - podziałów i królestw. Liczba wydziałów waha się od 4 do 10-12. W rosyjskiej literaturze fykologicznej prawie każda z powyższych klas odpowiada działowi. W literaturze zagranicznej zauważalna jest tendencja do konsolidacji wydziałów i co za tym idzie zmniejszania ich liczby.
Najbardziej powszechnym w schematach klasyfikacyjnych jest schemat Parkera (Parker, 1982). Uznaje podział na formy prokariotyczne i eukariotyczne. Formy prokariotyczne nie mają w swoich komórkach organelli otoczonych błoną. Do prokariotów zaliczają się bakterie i sinice (cyjanobakterie). Formy eukariotyczne obejmują wszystkie inne glony i rośliny. Podział glonów jest od dawna przedmiotem dyskusji. Harvey (1836) podzielił glony głównie ze względu na kolor. Chociaż obecnie rozpoznaje się znacznie więcej podziałów, duże znaczenie przywiązuje się do składu pigmentów, cech biochemicznych i strukturalnych struktury komórkowej. P. Silva (1982) wyróżnia 16 głównych klas. Klasy różnią się pigmentacją, produktami przechowywania, cechami ściany komórkowej i ultrastrukturą wici, jądra, chloroplastów, pyrenoidów i oczek.
Nowe informacje na temat ultrastruktury glonów, uzyskane w ostatnich dziesięcioleciach metodami mikroskopii elektronowej, genetyki i biologii molekularnej, umożliwiają badanie najdrobniejszych szczegółów budowy komórki. „Eksplozje” informacji okresowo skłaniają naukowców do ponownego rozważenia ustalonych tradycyjnych poglądów na temat taksonomii glonów. Ciągły napływ nowych informacji stymuluje nowe podejścia do klasyfikacji, a każdy proponowany schemat nieuchronnie pozostaje przybliżony. Według współczesnych danych organizmy tradycyjnie zaliczane do roślin niższych wykraczają poza zakres Królestwa Roślin. Należą do dużej liczby niezależnie rozwijających się grup. Tabela przedstawia megataksony, do których zaliczają się glony, w różnych interpretacjach. Jak widać, w różnych gromadach można znaleźć różne taksony glonów; ten sam typ może łączyć różne grupy ekologiczne i troficzne organizmów (tabela).
Ponad 100 lat temu K.A. Timiryazev przenikliwie zauważył, że „nie ma ani rośliny, ani zwierzęcia, jest tylko jeden, nierozerwalny świat organiczny. Rośliny i zwierzęta to tylko wartości przeciętne, tylko typowe wyobrażenia, które tworzymy, abstrahując od znanych cech organizmów, do jednych przywiązując wyjątkową wagę, zaniedbując inne. Teraz nie możemy nie podziwiać jego niesamowitej biologicznej intuicji.
Współczesny system glonów przedstawiony w tym podręczniku obejmuje 9 działów: niebiesko-zielone, czerwone, okrzemki, heterokonty, haptofity, kryptofity, dinofity, zielone, charofity i euglenofity. Podobieństwo w składzie pigmentów, strukturze aparatu fotosyntetycznego i wici posłużyło jako podstawa do połączenia klas glonów o złotobrązowym zabarwieniu w jedną dużą grupę - Heterokontae, czyli glony heteroflagellate (Ochrophyta).
Megasystem organizmów zaliczanych do roślin niższych
|
Imperium |
Królestwo |
Dział (typ) |
Trofogrupa |
|
Eubakterie/Prokariota |
Sinice/bakterie |
Sinice/cyjanobakterie |
Wodorost |
|
Wykopuje/Eukaryota |
Euglenobionty/pierwotniaki |
Euglenophyta/Euglenozoa Acrasiomycota |
Wodorost Myxomycetes |
|
Rhizaria/Eukariont |
Cercozoa/Plantae |
Chlorarachniophyta Plasmodiophoromy-cota |
Algi Myxomycetes |
|
Rhizaria/Eukariont |
Myxogasteromycota Dictyosteliomycota |
Myxomycetes Myxomycetes |
|
|
Choromalveola-tes/Eukaryota |
Straminopilae/Chromista/Heterokontobiontes |
Labirynthulomycota -Oomycota Heterokontophyta |
Algi grzybów Myxomycetes |
|
Choromalveola-tes/Eukaryota |
Haptofity/Chromista |
Prymnesiophyta/Haptophyta |
Wodorost |
|
Choromalveola-tes/Eukaryota |
Kryptofity/Chromista |
Wodorost |
|
|
Choromalveola-tes/Eukaryota |
Pęcherzyki/pierwotniaki |
Dinophyta/Myzozoa |
Wodorost |
|
Rośliny/Eukarionty |
Glaukofity/Planty |
Glaucocystophyta/glaucophyta |
Wodorost |
|
Rośliny/Eukarionty |
Rodobionty/Planty |
Cyanidiophyta Rhodophyta |
Algi Algi |
|
Rośliny/Eukarionty |
Chlorobiontes/Plantae |
Chlorophyta Charophyta |
Algi Algi |
WODOROST
ogólna charakterystyka
Glony stanowią dużą grupę roślin niższychnium - od mikroskopijnych małych organizmów jednokomórkowych powielokomórkowi giganci. Termin „algi” nie jest systememjednostka matyczna. Pod nazwą „algi” łączy siękilka systematycznych podziałów roślin niższych, mających różne pochodzenie, scharakteryzowanych w podobny sposóbżycie i autotroficzny sposób odżywiania. Jak sama nazwa wskazuje,Glony zazwyczaj prowadzą wodny tryb życia.
Ciało glonów, podobnie jak innych roślin niższych, jestplecha - plecha niepodzielna na korzeń lub łodygęi liścia, a u większości gatunków ma prostą budowę anatomiczną. Glony zawierają chlorofil w swoich komórkach i są do tego zdolnefotosynteza. Strukturakomórki, ciała, metody rozmnażania są różne u poszczególnych przedstawicieliprzedstawiciele tej grupy wydziałów roślinnych.
Większość komórek glonów ma ścianę komórkową celulozy; Istnieją komórki, których zawartość jest otoczona jedynie cienką błoną ograniczającą. Ściany komórkowe glonów są zróżnicowane pod względem budowy i skład chemiczny. Podstawa powłokijest białko- kompleks węglowodanów. Powłoka charakteryzuje się niejednorodnością i warstwowością. Warstwy różnią się między sobą m.ingrubość, gęstość i skład chemiczny. Często muszle ulegają nasyceniu związki organiczne(lignina i kutyna).
W skorupie znajdują się specjalne otwory zwane porami. Oprócz porów ściany komórkowe wielu glonów są wyposażone w różnego rodzaju struktury.stami - włosie, kolce i łuski.
W protoplastach komórki dzielą się na jądro i cytoplazmę. Boli cię?Większość glonów ma tylko jedno jądro w komórce, ale czasamijest ich 2, 3 lub więcej. Komórkom niebiesko-zielonych alg brakuje uformowanego jądra. Kształt, wielkość i położenie jądra w komórceróżnią się znacznie między różnymi glonami. W rdzeniu alg jestzawierają takie same struktury jak w jądrach innych roślin: łupina,sok jądrowy, jąderko, wtrącenia chromatyny.
Cytoplazma składa się z substancji głównej (zrębu) i zanurzonych w niej ciał (organelli). Osobliwość klejPrzepływ glonów jest słabym rozwojem retikulum endoplazmatycznego.
W określonych organellach – chloroplasty (tzw. chromatofory) to ciała bogate w substancje białkowe,zwane pyrenoidami. Pyrenoid jest otoczony wyściółką w postaci pierścienia lub oddzielnych płytek, zwykle skrobiowychNatura. Chloroplasty zawierają zielony pigment chlorofil,występujący w kilku formach.
Oprócz chlorofilu algi zawierają także inne pigmenty, którektóre często swoją obecnością maskują zielony kolor wodydorastanie. Najbardziej charakterystyczne i najczęściej występujące pigmentyalgi tami, z wyjątkiem chlorofilu (zielony), karotenu (żółto-pomarańczowy) i ksantofil (żółty), są fikocyjanami, fiko-erytryna i fukoksantyna. Pigment fikocyjanowy, rozpuszczalny w wodzie,barwi algi na niebiesko (niebiesko-zielone algi);fikoerytryna, również rozpuszczalna w wodzie, nadaje czerwony kolor(algi czerwone); fukoksantyna powoduje brązową barwę(algi brunatne).
Posiadające pigmenty glony nadają podłożu, na którymosiedlają się w różnych kolorach. Na przykład na Antarktydzie radzieccy naukowcy odkryli 3 jeziora o różnych kolorachwoda: niebieska, zielona i czerwona. Określono kolor wody w tych jeziorachobecność w nich bardzo małych glonów o odpowiednim kolorze.Wraz z nadejściem wiosny oblodzone wybrzeża Grenlandii w wielu miejscachtemperatury ocieplenia są zabarwione na czerwono (krew), co można wytłumaczyć masowym namnażaniem się glonów o czerwonym zabarwieniu.skromny.
Nazwa Morza Czerwonego wynika z wielkiego wzrostuilość glonów w tym morzu, które mają czerwonawy odcień.Różne kolory alg mają znaczenie adaptacyjnecja. Światło dzienne dla wodorostów głęboko w wodziezawsze przyjeżdża w zmienionym składzie. Woda jest stosunkowo dobraprzepuszcza kolor niebieski i zielony oraz silnie pochłania kolor czerwony i żółtypromienie. Na dużych głębokościach zatrzymanie promieni zielonych o jedenchlorofil jest zły, z pomocą przychodzi czerwieńpigment łatwo pochłaniający zielone promienie. Dlatego czerwieńlub fioletowe, algi powszechne na dużych głębokościachmorza.
Glony są bardzo zróżnicowane pod względem kształtu i wielkości. PoznaćIstnieją mikroskopijnie małe i osiągające ogromne rozmiary (do 50 m lub więcej) formy jednokomórkowe i wielokomórkowe. Etap przejściowy między nimi zajmują glony kolonialne.Kolonie składają się z kilku luźno połączonych jednorodnychkomórki. Pewnie, że wraz ze wzrostem wielkości glonówrajskie zróżnicowanie ich ciał. Na przykład w algach brunatnychkorowanie duże rozmiary, plecha jest silnie rozcięta. Takie glony przyczepiają się do podłoża cienkimi, bezbarwnymi nitkami,które nazywane są ryzoidami.
Istnieją jednokomórkowe glony, do których dociera organizmduże rozmiary 0,5 m i więcej. Mają dużą liczbęjądra i chromatofory. Zewnętrznie ciało takich glonów ma następujące cechy:Bez wątpienia, ale wewnątrz nie ma przegród, czyli jest jednokomorowy. Przykładem takich gigantycznych jednokomórkowych glonów jestżywa kalerpa morska.
Rozmnażanie glonów może być wegetatywne, bezpłciowe i płciowe. Rozmnażanie wegetatywne odbywa się w częściachciała glonów. Rozmnażanie bezpłciowe następuje w wyniku tworzenia zoospor, rzadziej zarodników. Proces seksualny w różnych typachglony reprezentowane są przez: izogamię, heterogamię, oogamię iautogamia.
W niektórych bardziej zorganizowanych algach (czerwonych,brązowy) następuje naprzemienność pokoleń płciowych i bezpłciowych.
Glony charakteryzują się dużą różnorodnością. Ogólnyliczba ich gatunków wynosi ponad 20 tysięcy.Klasyfikacja glonów jest bardzojest złożony i obecnie nie został jeszcze ukończony. Gatunki łączy się w większe jednostki systematyczne - rodzaje, rodzinystany, rozkazy, klasy, działy (typy).
Całą odmianę glonów zwykle łączy się w 6...10 dużych działów (typów), których przedstawiciele różnią się budowąniya, metoda rozmnażania, pochodzenie, ale najczęściejUchwyty tych sekcji różnią się od siebie kolorem.
Podręcznik ten omawia 6 działów glonów: błękitno-zielony, heteroflagellates, okrzemki, zielony, brązowy i czerwonynowy
Algi mają bardzo starożytne pochodzenie, w którymwiele jest nadal niejasnych. Nie ma jednego punktu widzenia na temat powiązanych relacji między działami glonów. Niektórzy z ich przedstawicielinajwyraźniej pochodzi od prostszych organizmów, takich jak wiciowce, a same glony są przodkami niektórych więcej niż razposkręcane rośliny (grzyby, mchy).
Przedstawiciele współczesnego różnorodnego świata algmają różną starożytność w pochodzeniu, pojawiały się w różnych epokach geologicznych. Uwzględniane są najstarszeniebiesko-zielone algi.
Ewolucja glonów przeszła z form mobilnych do nieruchomych.Rozważane są bardziej prymitywne i starożytne grupy glonówci, którzy spędzają życie w stanie mobilnym; bardziej organiczneTak zwane glony charakteryzują się siedzącym trybem życia. Ścieżkaewolucja glonów wyszła od pierwotniaków - jednokomórkowych mikroformy mikroskopijne do form złożonych wielokomórkowych.
Jak już wspomniano, zdecydowana większość glonówżyją w środowisku wodnym. Niektóre z nich rosną w słonej wodzie oceanówi morzach, inne - w rzekach, stawach i jeziorach słodkowodnych. Bólznaczący wpływ na rozmieszczenie glonów w głębinach zbiorników wodnychzapewnia światło, a więc w powierzchniowych warstwach wody ilośćZawsze jest więcej glonów. Czerwone i brązowe wodorostyczęsto tworzą ogromne podwodne zarośla zajmujące dziesiątki kilometrów. W zależności od siedliska glonówdzielą się na 2 duże grupy: bentosową i planktonową.
Do dennych lub dennych glonów żyją przyczepioneobiekty denne lub podwodne i tworzą głównie zaroślaV pas przybrzeżny. Glony planktonowe żyją w wodziew stanie zawieszonym, bez mocowania do dna, można je transportowaćnapędzany ruchem wody.
Glony żyją nie tylko w wodzie, można je znaleźć na jej powierzchniw glebie, na drzewach, na stosach, ale zawsze w wilgociw niektórych miejscach. Często zwłaszcza wczesną wiosną, gleby „kwitną”,lub „zmienia kolor na zielony”, co tłumaczy się rozwojem kolosalnej colijakość mikroskopijnie małych glonów. „Kwitnienie” gleby zglony występują na różnych obszarach związek Radziecki - na północy, stepie, a nawet pustyni. Rosnące w dużych ilościach stwarzają warunki do rozwoju bakterii i nie tylkomikroorganizmy.
Algi mają ogromne znaczenie w przyrodzie i są szeroko stosowane.zwany osobą.
Jako rośliny autotroficzne przetwarzają ogromne ilości colijakość minerałów i dwutlenku węgla w masie organicznej. Glony tworzą ogromne rezerwy pożywienia dla zwierzęciaświat mórz, oceanów i zbiorników słodkowodnych. Zatem z 1 hektara zarośli glonów można uzyskać plon równy 100 tonom mokrej lub 10 ton suchej glebyszerokie rzesze.
Glony wytwarzają ogromne ilości tlenu.
Rozwój rybołówstwa jest nierozerwalnie związany z glonami. Absorbując dużo dwutlenku węgla i uwalniając tlen, glonyczyste zbiorniki wodne.
W krajach przybrzeżnych (Anglia, Francja, Norwegia, Irlandiaitp.), szczególnie w Japonii, powszechnie spożywa się wodorosty(wodorosty itp.) oraz na paszę dla zwierząt gospodarskich (w postaci surowej, suszonej i kiszonki).forma kąpieli). Niektóre glony wykorzystuje się jako nawózpola.
Wiele glonów gromadzi duże ilości jodu i bromu. Jod zawarty jest w popiele z alg w ilości około 0,2% suchej masy.Głównym źródłem jodu są często algi. Gotując algi czerwone i brunatne uzyskują cenną substancję – agar-agar,który jest stosowany w mikrobiologii jako pożywkado hodowli mikroorganizmów, a także w cukiernictwieprzemysł w produkcji marmolady itp.
W starożytnych epokach geologicznych okrzemki razemz krzemionką utworzyła się skała osadowa - Trypolis, którastosowany do produkcji dynamitu, cegieł i do polerowaniaobiekty itp.
W zbiornikach słodkowodnych glony uczestniczą w tworzeniu sapropula lub osad organiczny. Sapropel zawiera dużą ilośćw substancjach organicznych i jest często stosowany w terapii błotnej.Sapropel, który zawiera dużą ilość soli wapniajon, fosfor, żelazo, mogą być stosowane w paszach rolniczychzwierzęta hodowlane.
Glony, zwłaszcza wodorosty, mogą wyrządzić szkody, gdypokrywać w dużych ilościach podwodne części statku. Namasowa śmierć glonów, następuje psucie się wody i, jakw konsekwencji śmierć ryb.
Zakład niebiesko-zielonych alg
Są to niebiesko-zielone algi, zwane cyjanaminajbardziej prymitywne i najstarsze pochodzenieorganizmy. Na podstawie danych paleobotanicznychOczywiste jest, że współczesne niebiesko-zielone algi nie różnią się zbytniood swoich skamieniałych przodków. W większości przypadków są one reprezentowane przez formy jednokomórkowe, choć są też wielokomórkowewszelkie formy zebrane w koloniach.
Jak sama nazwa tego działu glonów wskazuje, dla nichcharakterystyczny niebiesko-zielony kolor o różnych odcieniach w zależnościzależy od stosunku pigmentów - chlorofilu, karotenu, fiko-cyjanina i fikoerytryna.
Ściana komórkowa sinic składa się z pektyngęstą substancją i jest pokryty na zewnątrz śluzową substancją. W ich klatcenie ma morfologicznie odrębnego jądra i chromatoforów,cytoplazma jest nasycona pigmentami i dlatego nazywa się ją chromatoosocze.
W procesie asymilacji, zamiast zwykłego załamania się roślin,powstaje niewielka ilość glikogenu węglowodanowego (skrobi zwierzęcej).
Niebiesko-zielone algi rozmnażają się bardzo szybko i w prosty sposóbdzieląc komórki na pół. Ich proces rozmnażania płciowegonieobecny.
Zgodnie ze strukturą i charakterem podziału komórkowego, niebiesko-zielona woda-wzrosty są podobne do bakterii. Podobnie jak bakterie nie mają jasno określonego jądra, nie mają wiciowych stadiów rozwojuistnieć. Niektóre niebiesko-zielone algi (nitkowate) mają inne właściwościŻycie występuje na obszarach, na które te glony się rozpadają. Takie obszary nazywane są hormonami. Kiedy niekorzystnew przyjemnych warunkach ze zwykłych komórek powstają zarodniki, którepokryty pogrubioną skorupą. Chroni zawartośćprzed niekorzystnymi warunkami, dzięki którym glony się zachowujążywotność przez długi czas. Kiedy awansujeszsprzyjających warunkach, zarodniki kiełkują i dają początek nowymklatka szybowa.
Niebieskozielone algi żyją głównie w wodach słodkichwody – stawy, jeziora, rzeki, ale spotykane także w morzach,na powierzchni gleby, na skałach. Puszka niebiesko-zielonych algżyć tak, jakby niskie temperatury na śniegu i lodzie oraz na wysokościachniskie temperatury (do 80°C) w gorących źródłach. Po śmiercimasa komórkowa glonów w postaci płatków o brudnej zielonkawej barwiewypływa na powierzchnię wody.
Starożytne pochodzenie, niezróżnicowana struktura komórkiprąd, brak utworzonego jądra i procesu płciowego, rozmnażanieprosty podział komórek, zdolność do tworzenia zarodników -wszystkie te cechy wskazują na prymitywność niebiesko-zielonychwodorost Uproszczony na swój sposóbdo nowej struktury są one znacząceróżni się od innych glonówsą najbliżej bakterii.
Niebiesko-zielone algi są bardzo elastycznezdolność przystosowania się do różnych warunkówmniam środowisko, Jakprzyczynił się do ich zachowaniado dziś bez żadnego specjalnegozmiany.
Dział niebiesko-zielonego wodorusli jednoczy około 1400 viDow. Przedstawicielami tego działu mogą być:rosły jak Chroococcus, Oscillatorium, Nostoc itp.
Chrookok- jednokomórkowe sharo z algwyróżniające się kształtem, czasami glony te tworzą kolonie.Często takie glony zbiera się w grupach po 2...4, rozdzielonychcienka przegroda i otoczona ogólnie dość grubym śluzemgruba warstwa.
Chroococcus jest szeroko rozpowszechniony na bagnach wśród ras wodnychcienie, pomiędzy kępami i wśród błota.
Oscylator- nitkowate niebiesko-zieloneglony powszechnie występujące w zbiornikach wody stojącej, częstotworzy ciemnozielony film na powierzchni wody lub na mulistym dnie. Komórki tych glonów mają kształt cylindryczny, ściśle przylegającypołączone w jeden wątek.
Nostoc - nitkowate niebiesko-zielone algi, nitkilub jego łańcuchy są połączone w kolonie, często o kształcie kulistym,wielkości owocu śliwki. Na zewnątrz kolonie te są pokryte galaretowatą powłokąmasa. Nostok żyje wzdłuż brzegów stawów i jezior, na wilgotnej glebiei na dnie zbiorników.
Tworzą się sinice i grzybyróżne rodzaje porostów.
Dział wieloflarowy wodorost
Multiflagellates lub żółto-zielone,glony charakteryzują się tym, że posiadają zoospory2 nierówne wici, krótka wici gładka, a długa wici pierzaste. Chromatofory są żółto-zielone i mają kształt dysku.Organizmy jednokomórkowe, kolonialne, nitkowate i niekomórkowe.Przykładem jest botryd.
Botridium- najbardziej charakterystyczny predsadzarka glonów heteroflagellatowych. Plecha to zielony pęcherzyk o średnicy 1...2 mm znajdujący się na spodniej stronie pęcherzykawystępują bezbarwne rozgałęzione odrosty - ryzoidy, którealgi wnikają do gleby. Jest to jednokomórkowy organizm wielojądrowywodorost Środek pęcherzyka jest wypełniony sokiem komórkowym, cytoplazma znajduje się od ściany do ściany. Chromatofory zawierają dużo karotinoidy, dlatego botridium ma żółto-zielony kolor.Nie ma pyrenoidów.
Rozmnaża się głównie przez zoospory, które są wytwarzanew kolosalnych ilościach w środku bańki. Żyje na wilgocina ziemi, wzdłuż krawędzi kałuż, tworzy na glebie ciemnozielone osadyzabarwienie.
Podział okrzemek wodorost
Okrzemki, okrzemki, czy krzemionka, czy bakcylaria, algi – skrajnośćherbata różnorodna, w większości mikroskopijnaprzypadkach organizmy jednokomórkowe. Ten dział glonówcharakteryzuje się unikalną strukturą komórkową. Komórka jest całkowicie pokrytaotoczka w postaci pektyny, galaretowata błonka, na zewnątrzjest pokryty krzemionkową skorupą, która składa się z dwóch częścipołówki stojące, tzw. skrzydła. Jeden z tychskała zakrywa drugą niczym wieko pudełka. Wzdłuż połówekSkorupa ma po obu stronach otwór przypominający szczelinę. Przez tootwarcie cytoplazmy komórkowej komunikuje się z otoczenie zewnętrzne. Celki mają wyjątkową siłę. Nie trawiąodporne na zwierzęta i ptaki, nie zapadają się nawet po podgrzaniuw ogniu. Istnieje ponad 5000 gatunków. Charakterystyczne widokiGłównymi objawami są kształt komórek i różne zgrubieniamuszelki w postaci blizn, oczek itp.
Kształt komórek poszczególnych gatunków okrzemek jest zróżnicowanywydłużony kwadrat, eliptyczny, okrągły, w kształcie gwiazdy, w formiewstążki, spirale itp. Komórki zawierają cytoplazmę, jądro i jednolub kilka chromatoforów. Oprócz chlorofilu i fukoksantyny,Chromatofory zawierają także inne żółto-brązowe pigmenty,dlatego chromatofory mają żółty. W komórkach okrzemek nie ma skrobi, są to substancje rezerwoweolejek Leny.
Rozmnażają się głównie przez bezpośredni podział, któryChodzą w specyficzny sposób. Podczas podziału każda komórka potomnaotrzymuje rdzeń, jeden chromatofor i tylko jedną z klapek powłoki,drugi zawór jest formowany od nowa. Oprócz bezpośredniego podziału, diatowodorosty rozmnażają się płciowo, gdy się łącządwie komórki, które wcześniej zrzuciły swoje skorupy.
Okrzemki żyją w wodach morskich i słodkich,często są głównymi część integralna planktonu i służą jako cenny pokarm dla zwierząt. Płatki martwych komórek glonów opadają na dno i stopniowo tworzą ogromne osady,znana pod nazwą „mąka górska, okrzemkowa, trypolisowa”.
Duże złoża okrzemek w postaci okrzemek ila koncentrują się w polarnych obszarach oceanów, w pobliżu Alaski,Wyspy Aleuckie na Morzu Ochockim i Beringu. Dostępnysą także na Morzu Bałtyckim.
Do okrzemek należą algi słodkowodne piniusRija,Fragilariai kartę zgłoszeniaRia; glony te tworzą kolonie w postaci wstążeklub łańcuchy;naviculakrzew colo rośnienija.
Dział zielonych alg
Glony zielone są jedną z najbardziej różnorodnych grup glonów, obejmującą około 5000 gatunków.Przedstawiciele tego działu charakteryzują się zielonym kolorem,który jest powodowany przez chlorofil i nie jest przez żaden maskowanylub inne pigmenty. Zielone algi są reprezentowane przez formy jednokomórkowe, wielokomórkowe i kolonialne. Warzywaglony najczęściej mają strukturę nitkowatą, z której składają się nicijeden rząd komórek. Komórka ma błonę celulozową, cytoplazma, jądro i chromatofory. Rozmnażanie odbywa się bezpłciowosposób wegetatywny i seksualnyalgi zielone żyją w wodzie,Ale część z nich żyje na lądzie, natematy
Większość zaprezentujebędę żyć dalejdrzewa, skałynyah w wilgotnych, zacienionych miejscach. Tworzą się w stawach i rzekachbłoto.
Dział zielonych alg dzieli się na kilka klas, z którychRozważmy najpierw równowiciowce, a właściwie zielone, spoiste glony lub koniugatyi characeae, czyli promienieKlasa równowiciowców Wśród wód zielonychTa klasa jest najbardziej rozbudowana. Przedstawiciele są zróżnicowaniwyglądem i strukturą wewnętrzną. To dla nich typoweobecność dwóch identycznych wici. Ta klasa łączy jednąformy komórkowe i kolonialne, mobilne i nieruchome.Klasa jest podzielona na 8 rzędów. Rozważmy przedstawicieli 4wydziwianie.
Zamówienie volvox . Bardzocharakterystycznymi przedstawicielami rzędu Volvoxidae sąChlamydomonas i Volvox.
Chlamydomonas jest jednokomórkowy,algi mobilne. Żyje w dużych ilościachw słodkich, płytkich zbiornikach wodnych - kałuże, stawy, rowywzdłuż dróg. Obficie rozmnażany, zmienia kolor wody na zielonykolor. Jest to mikroskopijnie mała owalna komórkalub okrągły kształt. Na jednym (przednim) końcu komórka jest wydłużonaw formie dziobka, na tym samym końcu znajdują się 2 równe wici ułatwiające przemieszczanie się glonów. Cała jama komórkowa jest wypełnionanie objęte cytoplazmą, w której znajduje się jądro, bliżej dziobka
Volvoks, lubszczyt, jest charakterystycznym przykładem kolonialnych form mikroskopijnych glonów. Kolonię tych glonów widać gołym okiem, sięgawielkości główki od szpilki i ma kształt kuli. Taka kulista kolonia Volvox składa się z ogromnej liczby komórek (do 50 tysięcy), rozmieszczonych w jednej warstwie na obwodzie kuli.Każda komórka ma 2 wici. Wszystkie wici znajdują się wzdłużperyferie i przyczyniają się do ruchu całej kolonii. Ruchwici wszystkich komórek są zawsze spójne. Wnęka kuli jest wypełnionanie pokryte płynnym śluzem. Volvox rozmnaża się wegetatywnie iseksualnie. Podczas rozmnażania wegetatywnego w organizmie matkikolonia (kula) Tworzy się 8...15 kolonii potomnych (kulek). NaW miarę dojrzewania kolonii potomnych ściany dorosłej kuli pękająwyłaniają się młode kolonie, po których następuje okrężnica macierzystaNiya umiera.
Rozmnażanie płciowe - oogamia - występuje tylko u większościlee większe komórki. Jednocześnie w komórkach kolonii VolvoxRozwijają się nagie gamety dwuwiciowe, łączą się tylko gamety różnych osobników glonów.
Istnieje wiele rodzajów Volvoxów. Wszystkie są powszechnegłównie w stojącej, dobrze podgrzanej wodzie słodkiejzbiorniki wodne ze stojącą wodą - stawy, jeziora, rzadziej rzeki
Chlorellamikroskopijny jednokomórkowyglony kuliste. Chlorella charakteryzuje się szybkim działaniemreprodukcja i to bardzo aktywny proces fotosynteza. Dziękiobecność dużej liczby plastydów, chlorella ma inny efektaktywne wykorzystanie energii słonecznej, kultura konwencjonalnarośliny zużywają tylko 0,1% energii słonecznej w procesach biochemicznychprzemiany iczne i chlorella -2,5%, czyli 25 razy więcej.Inny pozytywna jakość chlorella – bardzo szybka zmianażycie
Ulotrix to nitkowata, nierozgałęziona alga,składający się z jednego rzędu komórek. Dolna bezbarwna komórka nicima specyficzny wydłużony kształtDzięki tej komórce algi przyczepiają się do podwodnych obiektów.
Włókno ulotrix wydłuża się w wyniku poprzecznego podziału komóreki może rosnąć w nieskończoność. Komórki są jednorodne,w skrócie, każdy z nich cytoplazma, jądro, chromatoformaz pyrenoidami. Ulothrix rozmnaża się głównie bezpłciowosposób, tworząc 4-wiciowe zoospory. Te zoospory są niektóreporuszać się przez chwilę, a następnie osiąść na jakimś podwodnym obiekciei wyrośnie nowy wątek.Proces seksualny to izogamia. Co więcej, gamety męskie i żeńskie nie są rozróżnialne zewnętrznie, ale pod względem fizycznymlogicznie są one różne i pochodzą z różnych wątków, a więc różneGamety są oznaczone znakiem + i znakiem -.
Ulothrix żyje w zbiornikach słodkowodnych, pod którymi rośnieobiekty wodne (skały, stosy), które zmieniają kolor na jasnozielonykolor.
Cladophora to nitkowata alga zielona.Plecha składa się z rozgałęzionych nici uformowanych w jednym rzędziewielojądrzastych, poszczególne rośliny wyglądają jak podwodnekrzewy Komórki są duże, wielojądrowe, o grubej,warstwowa skorupa. Występuje w wodach słodkich i słonych
Dział czerwony algi lub fioletowe naschiIstnieje ponad 600 rodzajów i około 4000 gatunków. Z innych algróżnią się kolorem farby, który zależy od obecnościOprócz chlorofilu mają jeszcze dwa pigmenty - fikoerytrynę (czerwony) i fikocyjan (niebiesko-zielony). Ze stosunkuKolor alg zależy od stężenia tych pigmentów (różni się w zależności odczerwony do prawie czarnego). Przedstawiciele działu w zdecydowanej większości organizmów wielokomórkowychmałe organizmy o złożonej budowie i tylko te najbardziej prymitywnemają plechę jednokomórkową lub kolonialną. Częściej plechama kształt krzaków, talerzy, u niektórych jest bardzo silnie rozcięty.
Algi czerwone żyją w morzach na większych głębokościach niżzielony i brązowy. Dzieje się tak dlatego, że czerwony pigment jestpomaga uchwycić zielone i niebieskie promienie na dużych głębokościachspektrum, a tym samym usprawnia proces fotosyntezy. W czerwonymW algach nie odkłada się zwykła skrobia. w odróżnieniuzwykła skrobia, jod zabarwia ją nie na niebiesko, ale na czerwonobrązowy kolor.
Algom czerwonym brakuje zoospor i plemników.Rozmnażanie bezpłciowe odbywa się za pomocą nieruchomegospór. Rozmnażanie płciowe - oogamia, ale zamiast plemnikówpowstają komórki męskie - plemniki, które są przenoszone do komórek jajowych przez wodę. Jaja powstają w specjalnych narządach -karpogony.
_Algi czerwone obejmująbezseriyaz plechą w kształcie krzaka; polisyfoniaw którym plecha ma wygląd rozgałęzionej nici.
Dział brązowy wodorost
Dla przedstawicieli działu, charakter brunatnictarnina ma brązową barwę ze względu na obecność w chromatoforachbrązowy pigment - fukoksantyna. Obecność fukoksantynymaskuje zielony kolor i nadaje tym algom brązowy kolorróżne odcienie. Oprócz fukoksantyny zawierają ksantofilo i karoten. Dział brunatnic jednoczy się900 gatunków.
Brunatnice wyróżniają się zwykle dużą wielokomórkową plechą. Najwięksi przedstawiciele glonów występują wśród brunatnic. Niektóre z nich, takie jak makrocystis, osiągają 60 m długości, ale zdarzają się również formy małekilka milimetrów od siebie.
Zamiast skrobi komórki brunatnic zawierają glukodla substancji słodkich - przynęt i wodorostów, które dająTe wodorosty mają słodkawy smak po ugotowaniu. JakCzęsto odkładają oleje jako substancję rezerwową.
Plecha brunatnic jest wieloletnia, ale ma kształt liściapłytki obumierają co roku i odrastają wczesną wiosną.
Złożony struktura zewnętrzna przyczyny brunatnicNaich i zróżnicowanie budowy anatomicznej (mająróżne kształty komórek). Niektórzy naukowcy uważają, że teglony mają nawet różne tkanki.
Brązowce rozmnażają się na różne sposoby. Nekoniektóre z nich rozmnażają się za pomocą prymitywnych środków seksualnych - isogamiia, gdy łączą się 2 gamety o identycznym kształcie. Inni,bardziej rozwinięte glony (kelp) są bardziej złożoneproces seksualny - oogamia, w którym duże jajołączy się z małą, ruchomą gametą męską - plemnikiemdom.
W brunatnicach następuje rozmnażanie bezpłciowezoospory, które powstają w dużych ilościach w zoosporachszeregi. Brunatniki mają dość jasno określoną przemianę pokoleń; aseksualny i seksualny. Na ostrzach w kształcie liścizebrane te algi tworzą jednokomórkowe zoosporangieuformowane w grupy, wśród których znajdują się sterylne nici.Każdy zoosporangium wytwarza 16...64 lub więcejpływka. Zoospory mają identyczny wygląd, ale różnią się fizjologicznie.Niektóre z nich kiełkują i tworzą mikroskopijnie małe żonymęskie gametofity, podczas gdy inne są męskimi gametofitami. O męskich gametofitachnastępnie tworzą się pylniki, a w każdym z nich znajduje się jeden plemnikzoid, a na żeńskich gametofitach tworzą się oogonia, niosącejedno jajko na raz. Po fuzji plemnika i komórki jajowejpowstaje zygota, z której rozwija się pokolenie bezpłciowe -sporofit
Glony brunatne to stworzenia morskie, których jest wielesą bardziej powszechne w morzach północnych, często tworząW morzach i oceanach są ogromne zarośla. W północnej części Atlantydyocean, na Morzu Sargassowym, w ogromnej liczbieIstnieje rodzaj brunatnic - Sargassum. Te glony są częściejznajdują się w stanie pływającym ze względu na obecnośćspecjalne bąbelki wypełnione powietrzem.
Brązowce są uważane za starożytną grupę roślinznaleziono więcej wysoki stopień zróżnicowanie nie tylkozewnętrzne, ale także wewnętrzne części plechy. Zewnętrznie są podobne do wysokichwspaniałe rośliny, więc niektórzy botanicy uważają, że teglony mogły dać początek roślinom wyższym.
Dział alg brunatnych składa się z 4 rzędów. Rozważmy wcześniejrośliny dwóch rzędów: wodorosty i morszczyny.
Zamów Laminarię. Tenbardzo duże glony, czasami osiągające 60 m lub więcej.Ich plecha jest silnie rozcięta, a ponadto dobrze rozwiniętarozgałęzione ryzoidy, za pomocą których glony mocno przylegają do dna morskiego. Laminaria żyją w przybrzeżnym pasie mórzna głębokości 5...10 m i często tworzą ogromne podwodne zarośla„lasy”.
Laminariaceae obejmują rodzaj Laminaria (obejmuje 30 gatunków), rodzaj Lessonia (obejmuje5 gatunki) i rodzaj Macrocystis. Glony te to rośliny wieloletnie, które różnią się od siebiestruktura plechy.
Dział śluzowata pleśń
Plasmodium powstaje w wyniku fuzji nagiej amebyróżne komórki śluzowca i w niektórych typach śluzowca docierająwielkości ludzkiej dłoni.On zwykle w kolorze jasnożółtymkolor i ma zdolność bardzo powolnego transferu przypominającego amebęprzesunąć (0,1 mm/min). Podczas ruchu plazmodium ma tendencjęchowaj się przed światłem i kieruj się w stronę źródła wilgoci. Poznaćśluzowce zwykle występują w zacienionych lasach; na zgniłych roślinach, pomiędzykora i drewno, w pęknięciach pniaków, pod opadłymi liśćmi.Rozmnażają się przez zarodniki.
Pozycja śluzowców w systemie filogenetycznym jest niejasna,Najwyraźniej pochodzą od niektórych wiciowców. Od tegodziale, spójrzmy na plazmodiofor.
Choroba rozprzestrzenia się przez glebę i rozwija się szczególnie szybkorośnie na glebach kwaśnych.
Glony zielone są najliczniejszą ze wszystkich grup glonów, licząc według różnych szacunków od 4 do 13 - 20 tysięcy gatunków. Wszystkie mają zielony kolor plechy, co wynika z przewagi chlorofilu w chloroplastach A I B nad innymi pigmentami. Komórki niektórych przedstawicieli zielonych alg ( Chlamydomonas, Trentepolia, Hematococcus) mają barwę czerwoną lub pomarańczową, co jest związane z gromadzeniem się pigmentów karotenoidowych i ich pochodnych poza chloroplastem.
Morfologicznie są bardzo zróżnicowane. Wśród zielonych alg są przedstawiciele jednokomórkowi, kolonialni, wielokomórkowi i niekomórkowi, aktywnie mobilni i nieruchomi, przywiązani i wolno żyjący. Rozpiętość ich rozmiarów jest również niezwykle duża – od kilku mikrometrów (co jest porównywalne wielkością z komórkami bakteryjnymi) do 1–2 metrów.
Komórki są jednojądrzaste lub wielojądrowe, z jednym lub większą liczbą chromatoforów zawierających chlorofil i karotenoidy. Chloroplasty są pokryte dwiema błonami i zwykle mają piętno, czyli ocellus, filtr przewodzący światło niebieskie i zielone do fotoreceptora. Oko składa się z kilku rzędów kuleczek lipidowych. Tylakoidy - struktury, w których zlokalizowane są pigmenty fotosyntetyczne - gromadzą się w stosy (lamele) po 2–6. W strefie przejściowej wici znajduje się formacja gwiaździsta. Najczęściej występują dwie wici. Głównym składnikiem ściany komórkowej jest celuloza.
Chlorofity mają różne rodzaje odżywiania: fototroficzne, miksotroficzne i heterotroficzne. Rezerwowy polisacharyd zielonych alg, skrobia, osadza się wewnątrz chloroplastu. Chlorofity mogą również akumulować lipidy, które odkładają się w postaci kropelek w zrębie chloroplastów i cytoplazmie.
Plechy wielokomórkowe są nitkowate, rurkowate, blaszkowate, krzaczaste lub o innej budowie i różnych kształtach. Ze znanych typów organizacji plechy u zielonych alg nieobecny jest tylko typ ameboidalny.
Są szeroko rozpowszechnione w wodach słodkich i morskich, w glebie i siedliskach lądowych (na glebie, skałach, korze drzew, ścianach domów itp.). Około 1/10 ogólnej liczby gatunków występuje w morzach, zwykle rosnąc w górnych warstwach wody do 20 m. Wśród nich występują formy planktonowe, peryfitoniczne i bentosowe. Innymi słowy, zielone algi opanowały trzy główne siedliska organizmów żywych: woda – ziemia – powietrze.
Zielone algi wykazują fototaksję dodatnią (ruch w kierunku źródła światła) i ujemną (ruch od jasnego źródła światła). Oprócz intensywności światła temperatura wpływa na fototaksję. Zoospory gatunków z tego rodzaju wykazują pozytywną fototaksję w temperaturze 160°C Hematococcus, Ulothrix, Ulva, a także niektóre rodzaje glonów desmidianowych, w których ruch komórek odbywa się poprzez wydzielanie śluzu przez pory w skorupce.
Reprodukcja. Zielone glony charakteryzują się występowaniem wszystkich znanych metod rozmnażania: wegetatywnego, bezpłciowego i płciowego .
Rozmnażanie wegetatywne w postaciach jednokomórkowych komórka dzieli się na pół. Kolonialne i wielokomórkowe formy chlorofitu rozmnażają się przez części ciała (talus lub plecha).
Rozmnażanie bezpłciowe w zielonych algach jest szeroko reprezentowany. Częściej dokonują tego ruchliwe zoospory, rzadziej nieruchome aplanospory i hipnospory. Komórki, w których tworzą się zarodniki (sporangia), w większości przypadków nie różnią się od pozostałych komórek wegetatywnych plechy, rzadziej mają inny kształt i większe rozmiary. Tworzące się zoospory mogą być nagie lub pokryte sztywną ścianą komórkową. Liczba wici w zoosporach waha się od 2 do 120. Zoospory mają różne kształty: kulisty, elipsoidalny lub gruszkowy, jednojądrowy, bez oddzielnej otoczki, z 2–4 wiciami na przednim, bardziej spiczastym końcu i chloroplastem na końcu rozszerzony koniec tylny. Zwykle mają pulsujące wakuole i piętno. Zoospory powstają pojedynczo lub częściej z kilku z wewnętrznej zawartości komórki macierzystej, wychodzą przez okrągły lub szczelinowy otwór utworzony w skorupce, rzadziej w wyniku jej ogólnego śluzu. W momencie wyjścia z komórki macierzystej zoospory są czasami otoczone cienkim pęcherzem śluzowym, który wkrótce się rozpuszcza (rodzaj Ulotrix).
U wielu gatunków zamiast zoospor lub wraz z nimi powstają nieruchome zarodniki - aplanospory. Aplanospory to zarodniki rozmnażające się bezpłciowo, które nie mają wici, ale mają kurczliwe wakuole. Aplanospory uważa się za komórki, w których zawieszony jest dalszy rozwój zoospor. Powstają również z protoplastu komórki, jednego lub więcej, ale nie wytwarzają wici, ale po przyjęciu kulistego kształtu są pokryte własną skorupą, w tworzeniu której skorupa komórki macierzystej nie uczestniczy. Aplanospory są uwalniane w wyniku pęknięcia błon śluzowych komórek macierzystych i kiełkują po pewnym okresie spoczynku. Aplanospory o bardzo grubych błonach nazywane są hipnosporami. Przeważnie przejmują funkcję fazy spoczynkowej. Autospory, które są mniejszymi kopiami nieruchliwych komórek wegetatywnych, nie mają kurczliwych wakuoli. Powstawanie autosporów koreluje z pokonaniem warunków lądowych, w których woda nie zawsze może być obecna w wystarczającej ilości.
Rozmnażanie płciowe przeprowadzana przez gamety powstałe w niezmienionych, nieznacznie zmienionych lub znacząco przekształconych komórkach - gametangia. Ruchliwe gamety o strukturze monadycznej, dwuwiciowe. Proces seksualny w zielonych algach jest reprezentowany przez różne formy: hologamię, koniugację, izogamię, heterogamię, oogamię. W przypadku izogamii gamety są morfologicznie całkowicie do siebie podobne, a różnice między nimi są czysto fizjologiczne. Zygota pokryta jest grubą skorupą, często z rzeźbionymi naroślami, zawiera dużą ilość substancji rezerwowych i kiełkuje natychmiast lub po pewnym okresie spoczynku. Podczas kiełkowania zawartość zygoty u większości gatunków dzieli się na cztery części, które wyłaniają się z muszli i wyrastają na nowe osobniki. Znacznie rzadziej gamety rozwijają się w nowy organizm bez fuzji, samodzielnie, bez tworzenia zygoty. Ten rodzaj reprodukcji nazywa się partenogeneza i zarodniki utworzone z poszczególnych gamet partenospory.
W heterogamii obie gamety różnią się od siebie wielkością, a czasem kształtem. Gamety większe, często mniej mobilne, uważa się za żeńskie, mniejsze i bardziej mobilne – męskie. W niektórych przypadkach różnice te są niewielkie, a wtedy mówią po prostu o heterogamii, w innych są bardzo znaczące.
Jeśli gameta żeńska jest nieruchoma i przypomina bardziej jajo, wówczas ruchliwa gameta męska staje się plemnikiem, a proces seksualny nazywa się oogamią. Nazywa się gametangie, w których powstają jaja oogonia, Różnią się od komórek wegetatywnych zarówno kształtem, jak i rozmiarem. Gametangie, w których powstają plemniki, nazywane są gametangiami pylniki. Zygota powstała w wyniku zapłodnienia komórki jajowej przez plemnik tworzy grubą skorupę i nazywa się oospora.
W typowej oogamii jaja są duże, nieruchome i najczęściej rozwijają się pojedynczo w oogonii; plemniki są małe, ruchliwe i powstają w dużych ilościach w antheridium. Na jednym osobniku mogą rozwinąć się oogonia i antheridia, w tym przypadku glony są jednopienne; jeśli rozwiną się na różnych osobnikach, są dwupienne. Zapłodnione jajo pokryte jest grubą brązową skorupą; Często sąsiadujące z nim komórki wytwarzają krótkie gałęzie, które przerastają oosporę, oplatając ją jednowarstwową korą.
Cykle życia. Większość przedstawicieli glonów zielonych ma haplobiontowy cykl życiowy z redukcją zygotyczną. U takich gatunków jedynie zygota jest stadium diploidalnym – komórką powstałą w wyniku zapłodnienia komórki jajowej przez plemnik. Inny typ cyklu życiowego – haplodiplobiont z redukcją sporadyczną – występuje u Ulvoceae, Cladophoraceae i niektórych Trentepoliaceae. Glony te charakteryzują się naprzemiennością diploidalnego sporofitu i haploidalnego gametofitu. Cykl życiowy haplodiplobiontów z redukcją somatyczną jest znany tylko w Prasiole. Kwestionuje się obecność diplobiontowego cyklu życiowego u Bryopsidae i Dasycladiaceae.
U niektórych Ulothrixidae ten sam osobnik może dać początek zarówno zoosporom, jak i gametom. W innych przypadkach zoospory i gamety powstają u różnych osobników, tj. Cykl życiowy glonów obejmuje zarówno formy rozwoju płciowe (gametofit), jak i bezpłciowe (sporofity). Sporofit jest zwykle diploidalny, tj. ma w komórkach podwójny zestaw chromosomów, gametofit jest haploidalny, tj. ma pojedynczy zestaw chromosomów. Obserwuje się to w przypadkach, gdy mejoza zachodzi podczas tworzenia zarodników (redukcja sporyczna), a część cyklu życiowego glonów od zygoty do powstania zarodników odbywa się w diplofazie, a część od zarodnika do tworzenia gamet w haplofaza. Ten cykl rozwojowy jest typowy dla gatunków z rodzaju Ulva.
W obrębie alg Ulothrix powszechna jest redukcja zygotyczna, gdy podczas kiełkowania zygoty następuje mejoza. W tym przypadku tylko zygota jest diploidalna, reszta cyklu życiowego zachodzi w haplofazie. Redukcja gamet występuje znacznie rzadziej, gdy podczas tworzenia gamet zachodzi mejoza. W tym przypadku tylko gamety są haploidalne, a reszta cyklu jest diploidalna.
Taksonomia
Nadal nie ma jednego ustalonego systemu zielonych alg, zwłaszcza jeśli chodzi o grupowanie rzędów w różne proponowane klasy. Przez bardzo długi czas przy rozróżnianiu rzędów u zielenic główne znaczenie przywiązywano do rodzaju zróżnicowania plechy. Jednak ostatnio, w związku z nagromadzeniem danych na temat cech ultrastrukturalnych komórek wiciowych, rodzaju mitozy i cytokinezy itp., Niejednorodność wielu z tych rzędów jest oczywista.
Dział obejmuje 5 klas: Ulvophyceae, Brypsodaceae – Bryopsidophyceae, Chlorophyceae – Chlorophyceae, Trebouxiophyceae, Prasinophyceae – Prasinophyceae.
Klasa Ulvophyceae –Ulvophyceae
Znanych jest około 1 tysiąca gatunków. Nazwa klasy pochodzi od rodzaju typu Ulwa. Obejmuje gatunki z plechą nitkowatą i blaszkowatą. Cykle życiowe są zróżnicowane. Gatunki są przeważnie morskie, rzadziej słodkowodne i lądowe. Niektóre są częścią porostów. U przedstawicieli morskich w ścianach komórkowych może osadzać się wapno.
Zamów Ulotrix –Ulotrichales.
Rodzaj Ulotrix(ryc. 54). Rodzaje Ulotrix Częściej żyją w wodach słodkich, rzadziej w morzach, zbiornikach słonawych i w glebie. Przywiązują się do obiektów podwodnych, tworząc jasnozielone krzewy o wielkości do 10 cm lub większej. Nierozgałęzione wątki Ulotrix, składające się z pojedynczego rzędu cylindrycznych komórek z grubymi błonami celulozowymi, są przymocowane do podłoża za pomocą bezbarwnej stożkowej komórki podstawnej, która pełni funkcję ryzoidu. Charakterystyczna jest budowa chromatoforu, który ma postać płytki ściennej tworzącej otwarty pas lub pierścień (cylinder).
Ryż. 54. UlothricC (by:): 1 – plecha nitkowata, 2 – zoospora, 3 – gameta, 4 – kopulacja gamet
Rozmnażanie bezpłciowe Ulotrix można przeprowadzić na dwa sposoby: poprzez rozpad włókna na krótkie odcinki, z których powstaje nowe włókno, lub poprzez utworzenie w komórkach zoospor czterowiciowych. Zoospory wyłaniają się z komórki macierzystej, zrzucają jedną po drugiej wici, przyczepiają się bokiem do podłoża, pokrywają się cienką błoną celulozową i kiełkują, tworząc nowe włókno. Proces seksualny jest izogamiczny. Po zapłodnieniu zygota najpierw unosi się na wodzie, następnie osiada na dnie, traci wici, tworzy gęstą skorupę i śluzową łodygę, którymi przyczepia się do podłoża. To sporofit spoczynkowy. Po okresie spoczynku następuje podział jądra redukcyjnego i zygota kiełkuje w postaci zoospor. Tak w cyklu życia Ulotrix następuje przemiana pokoleń lub zmiana seksualnych i bezpłciowych form rozwoju: nitkowaty wielokomórkowy gametofit (pokolenie tworzące gamety) zostaje zastąpiony jednokomórkowym sporofitem - pokoleniem reprezentowanym przez rodzaj zygoty na łodydze i jest zdolny do tworzenia zarodników.
Zamawiaj Ulvaceae -Ulvales. Mają blaszkowatą, workowatą, rurkowatą lub rzadko nitkowatą plechę w różnych odcieniach zieleni. Krawędzie płytek mogą być faliste lub złożone, do mocowania do podłoża są wyposażone w krótką łodygę lub podstawę z małym dyskiem podstawowym. Gatunki morskie i słodkowodne. Najczęstszymi gatunkami rodzajów w wodach przybrzeżnych mórz Dalekiego Wschodu są Ulva, Monostroma, Cornmannia I Ulwaria.
Rodzaj Ulwa(ryc. 55). Plecha to jasnozielona lub jasnozielona, cienka dwuwarstwowa, często perforowana płytka lub jednowarstwowa pusta rurka, przymocowana do podłoża za pomocą podstawy zwężonej w krótki ogonek.
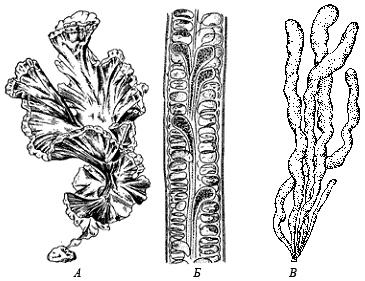
Ryż. 55. Ulwa: A- wygląd Ulva fenestrowana, B– przekrój wzgórza, W- wygląd Ulwa jelitowa
Zmiana form rozwoju w cyklu życia Ulwa redukuje się do izomorficznego, gdy etap bezpłciowy (sporofit) i etap płciowy (gametofit) są do siebie morfologicznie podobne, i heteromorficzny, gdy są morfologicznie różne. Gametofit jest wielokomórkowy, blaszkowy, sporofit jest jednokomórkowy. Gametofity wytwarzają gamety dwuwiciowe, a sporofity wytwarzają zoospory czterowiciowe.
Gatunki z rodzaju występują w morzach wszystkich stref klimatycznych, chociaż preferują ciepłe wody. Na przykład w płytkich wodach Morza Czarnego i Morza Japońskiego Ulva jest jednym z najliczniejszych rodzajów glonów. Dużo typów Ulwa tolerować odsalanie wody; często można je spotkać w ujściach rzek.
Klasa Bryopsidae–Bryopsidophyceae
Znanych jest około 500 gatunków. Plecha jest bezkomórkowa. Tworzą je proste lub splecione nitki syfonu, tworzące złożone struktury. Plecha w postaci bąbelków, krzewów, gąbczastych, dychotomicznie rozgałęzionych krzewów. Plecha jest podzielona na segmenty, co symuluje wielokomórkowość, składającą się z kilku lub wielu komórek jądrowych. Nici i krzewy wszystkich odcieni zieleni lub brązu.
Zamów Bryopsidae– Bryopsidales
Większość gatunków występuje w wodach słodkich i słonawych. Część z nich rośnie na ziemi, na kamieniach, piasku, czasem na słonych bagnach.
Rodzaj Bryopsis– nitkowate krzewy dorastające do 6-8 cm wys., pierzasto lub nieregularnie rozgałęzione, górne gałęzie ze zwężeniami u nasady. Plecha ma syfoniczną strukturę niekomórkową. Rośnie w pojedynczych krzakach lub małych kępach w strefie przybrzeżnej, żyje w morzach ciepłych i umiarkowanych (Załącznik 7B).
Rodzaj Kod– sznurkowate, dychotomicznie rozgałęzione krzewy o wysokości 10–20 cm, gąbczaste. miękkie, mocowane na podeszwie w kształcie krążka. Wewnętrzną część wzgórza tworzą kompleksowo splecione nici syfonu. Rośnie na glebach miękkich i twardych w strefie sublitoralnej do głębokości 20 m w pojedynczych roślinach lub w małych grupach (załącznik 7A, B).
Rodzaj Caulerpa obejmuje około 60 gatunków wodorostów, pełzające części plechy rozłożone na ziemi mają postać rozgałęzionych cylindrów, osiągających długość kilkudziesięciu centymetrów. W pewnych odstępach w dół od nich wychodzą obficie rozgałęzione ryzoidy, zakotwiczające roślinę w glebie, a w górę - płaskie, liściaste pionowe pędy, w których skupiają się chloroplasty.

Ryż. 56. Caulerpa: A – wygląd plechy; B – przekrój plechy z belkami celulozowymi
Plecha caulerpa, pomimo dużych rozmiarów, nie ma struktura komórkowa– całkowicie pozbawiony jest przegród poprzecznych i formalnie reprezentuje jedną komórkę olbrzymią (ryc. 56). Ta struktura wzgórza nazywa się syfon. Wewnątrz wzgórza caulerpa znajduje się centralna wakuola otoczona warstwą cytoplazmy zawierającą liczne jądra i chloroplasty. Różne części wzgórza wyrastają na końcach, gdzie gromadzi się cytoplazma. Centralną wnękę we wszystkich częściach plechy przecinają cylindryczne pasma szkieletowe - belki celulozowe, które nadają ciału glonów wytrzymałość mechaniczną.
Caulerpa łatwo rozmnaża się wegetatywnie: gdy obumierają starsze części plechy, jej poszczególne odcinki z pionowymi pędami stają się niezależnymi roślinami. Gatunki tego rodzaju żyją głównie w morzach tropikalnych, a tylko nieliczne wkraczają na szerokości subtropikalne, na przykład powszechne w Morzu Śródziemnym Kiełkuje Caulerpa. Glony te preferują płytkie, spokojne wody, na przykład laguny chronione przed działaniem ciągłych fal przez rafy koralowe i osiadają na różnych twardych podłożach - kamieniach, rafach, skałach, glebie piaszczystej i błotnistej.
Klasa Chlorophyceae–Chlorophyceae
Znanych jest około 2,5 tysiąca gatunków. Plecha jest jednokomórkowa lub kolonialna monnadyczna, wolno żyjąca.
Zamów Volvoxidae -Volvocales.
Rodzaj Chlamydomonas(ryc. 57) obejmuje ponad 500 gatunków jednokomórkowych glonów żyjących w świeżych, płytkich, dobrze nagrzanych i zanieczyszczonych zbiornikach wodnych: stawach, kałużach, rowach itp. Kiedy rozmnażają się masowo, woda zmienia kolor na zielony. Chlamydomonasżyje także na ziemi i śniegu. Jego ciało jest owalne, gruszkowate lub kuliste. Komórka pokryta jest gęstą skorupą, często pozostającą w tyle za protoplastem, z dwiema identycznymi wiciami na przednim końcu; z ich pomocą Chlamydomonas aktywnie porusza się w wodzie. Protoplast zawiera 1 jądro, chromatofor w kształcie miseczki, piętno i pulsujące wakuole.

Ryż. 57. Budowa i rozwój Chlamydomonas: A – osobnik wegetatywny; B – stadium palmelle; B – rozmnażanie (młode osobniki wewnątrz komórki macierzystej)
Chlamydomonas rozmnażają się głównie bezpłciowo. Kiedy zbiornik wyschnie, rozmnażają się, dzieląc komórkę na pół. Komórki zatrzymują się, tracą wici, ich ściany komórkowe stają się śluzem i w tym nieruchomym stanie komórki zaczynają się dzielić. Ściany powstałych komórek potomnych zawierają także śluz, dzięki czemu ostatecznie tworzy się system zagnieżdżonych w sobie błon śluzowych, w których grupami rozmieszczone są nieruchome komórki. To jest palmelle stan glonów. Kiedy dostaną się do wody, komórki ponownie tworzą wici, opuszczają komórkę macierzystą w postaci zoospor i przechodzą w samotny stan monadyczny.
W sprzyjających warunkach Chlamydomonas rozmnaża się intensywnie w inny sposób - komórka zatrzymuje się, a jej protoplast nieco za ścianą dzieli się sukcesywnie wzdłużnie na dwie, cztery lub osiem części. Te komórki potomne tworzą wici i pojawiają się jako zoospory, które wkrótce zaczynają się ponownie rozmnażać.
Proces rozrodczy u Chlamydomonas jest izogamiczny lub oogamiczny. Mniejsze gamety powstają wewnątrz komórki macierzystej w taki sam sposób jak zoospory, ale w większej liczbie (16, 32 lub 64). Zapłodnienie następuje w wodzie. Zapłodnione jajo zostaje przykryte wielowarstwową membraną i osiada na dnie zbiornika. Po okresie spoczynku zygota dzieli się mejotycznie, tworząc 4 osobniki haploidalnej córki Chlamydomonas.
Rodzaj Volvox- najbardziej zorganizowani przedstawiciele zakonu tworzą gigantyczne kolonie składające się z setek i tysięcy komórek. Kolonie mają wygląd kulek śluzowych o średnicy do 2 mm, w warstwie obwodowej których znajduje się do 50 tysięcy komórek z wiciami, połączonymi ze sobą bocznymi ścianami śluzowymi i połączonymi plazmodesmami (ryc. 58). Wnęka wewnętrzna

Ryż. 58. Pojawienie się kolonii Volvox
Kulka wypełniona jest płynnym śluzem. W kolonii istnieje specjalizacja komórek: jej obwodową część tworzą komórki wegetatywne, a pomiędzy nimi rozproszone są większe komórki rozrodcze.
Około tuzina komórek kolonii to gonidia, komórki rozmnażania bezpłciowego. W wyniku powtarzających się podziałów dają początek młodym koloniom potomnym, które wpadają do kuli matki i uwalniają się dopiero po jej zniszczeniu. Proces seksualny to oogamia. Oogonia i antheridia powstają również z komórek rozrodczych. Kolonie są jednopienne i dwupienne. Gatunki z rodzaju spotykane są w stawach i starorzeczach rzek, gdzie w okresie intensywnego rozmnażania powodują „zakwitanie” wody.
Klasa Trebuxiaceae –Trebouxiophyceae
Klasa nazwana na cześć rodzaju typu Trebuksja. Obejmuje głównie jednokomórkowe formy kokosowe. Są przedstawiciele sarcinoidów i nitkowatych. Formy słodkowodne i lądowe, rzadziej morskie, wiele z nich tworzy symbiozy. Około 170 gatunków.
Zamów chlorellę -Chlorele. Jednoczy przedstawicieli autospor kokoidowych.
Rodzaj Chlorella- algi jednokomórkowe w postaci stacjonarnej kuli. Komórka pokryta jest gładką skorupą; zawiera jedno jądro i ścianę, cały, wycięty lub płatkowany chromatofor z pirenoidem. Ściana komórkowa wielu gatunków, wraz z celulozą, zawiera sporopoleninę, substancję niezwykle odporną na działanie różnych enzymów, która występuje także w ziarnach pyłku i zarodnikach roślin wyższych. Chlorella rozmnaża się bezpłciowo, tworząc do 64 nieruchomych autospor. Nie ma rozmnażania płciowego. Chlorella szeroko rozpowszechniony w różnych zbiornikach wodnych, spotykany na wilgotnej glebie, korze drzew i części porostów.
Zamawiaj Trebuxiaceae - Trebouxiales . Obejmuje rodzaje i gatunki zaliczane do porostów.
Rodzaj Trebuksja- algi jednokomórkowe. Komórki kuliste mają pojedynczy osiowy chloroplast gwiaździsty z pojedynczym pyrenoidem. Rozmnażanie bezpłciowe odbywa się za pomocą nagich zoospor. Występuje w postaci wolno żyjącej w siedliskach lądowych (na korze drzew) lub jako fotobiont porostów.
Zajęcia Prazyne –Prasinophyceae
Nazwa klasy pochodzi z języka greckiego. prasino - zielony. Wiciowce lub rzadziej jednokomórkowe organizmy kokosowe lub palmellooidalne.
Zamów Pyramidonidae - Piramimonady. Komórki mają 4 lub więcej wici i trzy warstwy łusek. Mitoza jest otwarta, wrzeciono pozostaje w telofazie, cytokineza zachodzi w wyniku tworzenia się bruzdy szczelinowej.
Rodzaj Piramimony– organizmy jednokomórkowe (ryc. 59). Z przedniego końca komórki znajduje się 4–16 wici, które mogą być pięciokrotnie dłuższe od komórki. Chloroplast jest zwykle pojedynczy, z jednym pirenoidem i jednym lub większą liczbą oczek. Komórki i wici pokryte są kilkoma warstwami łusek. Szeroko rozpowszechniony w wodach słodkich, słonawych i morskich. Występują w planktonie i bentosie i mogą powodować zakwity wody.

Ryż. 59. Wygląd glonów Piramimony
Zamów Chlorodendraceae– Chlorodendrale. Komórki są skompresowane, z czterema wiciami, pokryte osłonką, mitoza jest zamknięta, zachodzi cytokineza w wyniku utworzenia bruzdy podziału.
Rodzaj Tetraselmis mogą występować jako ruchliwe komórki czterowiciowe lub jako komórki nieruchliwe przyczepione łodygami śluzowymi. Komórki pokryte są osłonką. Kiedy komórki dzielą się, wokół każdej komórki potomnej tworzy się nowa osłonka w osłonce komórki macierzystej. Na przednim końcu komórki wici wychodzą przez otwór w osłonce, który jest pokryty włoskami i łuskami. Jest jeden chloroplast z podstawowym pirenodem. Komórki są zwykle zielone, ale czasami zmieniają kolor na czerwony w wyniku nagromadzenia karotenoidów. Przedstawiciele morscy mogą żyć w płazińcach morskich.
Ekologia i znaczenie
Zielone algi są szeroko rozpowszechnione na całym świecie. Większość z nich można znaleźć w zbiornikach słodkowodnych, ale istnieje wiele form słonawych i morskich. Nitkowate zielone algi, przyczepione lub niezwiązane, wraz z okrzemkami i niebieskozieleniną, są dominującymi glonami bentosowymi w kontynentalnych zbiornikach wodnych. Występują w zbiornikach o różnej troficzności (od dystroficznej do eutroficznej) i o różnej zawartości substancji organicznych (od kseno- do polisaprobowych), jonów wodorowych (od zasadowych do kwaśnych), w różnych temperaturach (termo-, mezo- i kriofile). .
Wśród zielonych glonów występują formy planktonowe, peryfitoniczne i bentosowe. W grupie pikoplanktonu morskiego, algi prasinowe Ostreokoki uważana za najmniejszą wolno żyjącą komórkę eukariotyczną. Istnieją gatunki zielonych alg, które przystosowały się do życia w glebie i siedliskach lądowych. Można je znaleźć na korze drzew, skałach, różnych budynkach, na powierzchni gleby i w powietrzu. Przedstawiciele rodzajów są szczególnie pospoliti w tych siedliskach Trentepoly I Trebuksja. Zielone algi rosną w gorących źródłach o temperaturze 35–52°C, a w niektórych przypadkach nawet do 84°C i wyższej, często ze zwiększoną zawartością soli mineralnych lub substancji organicznych (silnie zanieczyszczone gorące ścieki z fabryk, fabryk, elektrowni lub elektrownie jądrowe). Przeważają także wśród gatunków glonów kriofilnych. Mogą powodować zielone, żółte, niebieskie, czerwone, brązowe, brązowe lub czarne „zakwity” śniegu lub lodu. Glony te występują w powierzchniowych warstwach śniegu lub lodu i intensywnie namnażają się w stopionej wodzie o temperaturze około 0°C. Tylko kilka gatunków przechodzi stadia spoczynku, podczas gdy większości brakuje żadnych specjalnych adaptacji morfologicznych do niskich temperatur.
W zbiornikach zasolonych dominują jednokomórkowe, mobilne algi zielone - hiperhaloby, których komórki nie mają błony i są otoczone jedynie plazmalemą. Glony te charakteryzują się zwiększoną zawartością chlorku sodu w protoplazmie, wysokim wewnątrzkomórkowym ciśnieniem osmotycznym, akumulacją karotenoidów i gliceryny w komórkach oraz dużą labilnością układów enzymatycznych i procesów metabolicznych. W słonych zbiornikach wodnych często rozwijają się w dużych ilościach, powodując czerwone lub zielone „kwitnienie” słonych zbiorników wodnych.
Mikroskopijne jednokomórkowe, kolonialne i nitkowate formy zielonych alg przystosowały się do niesprzyjających warunków życia w powietrzu. W zależności od stopnia zawilgocenia dzieli się je na 2 grupy: glony powietrzne, które żyją w warunkach wyłącznie wilgoci atmosferycznej i dlatego podlegają ciągłej zmianie wilgotności i suszeniu; algi wodne narażone na ciągłe nawadnianie wodą (pod strumieniem wodospadu, falami itp.). Warunki istnienia glonów w zbiorowiskach aerofilnych są bardzo wyjątkowe i charakteryzują się przede wszystkim częstymi i gwałtownymi zmianami dwóch czynników - wilgotności i temperatury.
W warstwie gleby żyją setki gatunków zielonych alg. Gleba jako biotop przypomina siedliska wodne i powietrzne: zawiera powietrze, ale jest nasycona parą wodną, co zapewnia oddychanie powietrzem atmosferycznym bez zagrożenia wysychaniem. Intensywny rozwój glonów jako organizmów fototroficznych możliwy jest jedynie w granicach przenikania światła. Na glebach dziewiczych jest to wierzchnia warstwa gleby o grubości do 1 cm, na glebach uprawnych jest ona nieco grubsza. Jednak w grubości gleby, gdzie światło nie przenika, żywe glony znajdują się na głębokości do 2 m na glebach dziewiczych i do 3 m na glebach ornych. Wyjaśnia to zdolność niektórych glonów do przejścia na odżywianie heterotroficzne w ciemności. Wiele glonów pozostaje w glebie w stanie uśpionym.
Aby utrzymać swoje funkcje życiowe, algi glebowe mają pewne cechy morfologiczne i fizjologiczne. Należą do nich stosunkowo niewielki rozmiar gatunków glebowych, a także zdolność do wytwarzania obfitego śluzu – śluzowatych kolonii, osłon i zawijasów. Dzięki obecności śluzu glony szybko wchłaniają wodę po zwilżeniu i magazynują ją, spowalniając wysychanie. Charakterystyczną cechą glonów glebowych jest „efemeryczność” ich okresu wegetacyjnego - zdolność szybkiego przejścia ze stanu spoczynku do aktywnego życia i odwrotnie. Są również w stanie tolerować różne wahania temperatury gleby. Zakres przeżycia wielu gatunków mieści się w zakresie od -200 do +84°C i więcej. Glony lądowe stanowią ważną część roślinności Antarktydy. Są prawie czarne, więc temperatura ich ciała jest wyższa niż temperatura otoczenia. Glony glebowe są także ważnym składnikiem biocenoz strefy suchej (suchej), gdzie latem gleba nagrzewa się do 60–80°C. Ciemne osłonki śluzowe wokół komórek stanowią ochronę przed nadmiernym nasłonecznieniem.
Wyjątkową grupę stanowią algi endolitofilne związane z podłożem wapiennym. Po pierwsze, są to nudne glony. Na przykład glony z rodzaju Gomontia Wwiercają się w skorupy jęczmienia perłowego i bezzębnych chrząszczy oraz penetrują wapienne podłoże zbiorników słodkowodnych. Sprawiają, że podłoże wapienne staje się luźne, podatne na działanie różnych czynników chemicznych i fizycznych. Po drugie, wiele glonów występujących w wodach słodkich i morskich ma zdolność przekształcania soli wapnia rozpuszczonych w wodzie w nierozpuszczalne i osadzania ich na plechach. Szereg tropikalnych zielonych alg, m.in. Galimeda, odkłada węglan wapnia w plechy. Biorą czynny udział w budowie raf. Gigantyczne złoża szczątków Halimedy, czasami osiągające 50 m wysokości, występują w wodach szelfu kontynentalnego związanych z Wielką Rafą Koralową w Australii i innych regionach, na głębokościach od 12 do 100 m.
Zielone algi Trebuxia, wchodzące w symbiotyczny związek z grzybami, są częścią porostów. Około 85% porostów zawiera jednokomórkowe i nitkowate zielone algi jako fotobionty, 10% zawiera sinice, a 4% (lub więcej) zawiera zarówno niebiesko-zielone, jak i zielone algi. Występują jako endosymbionty w komórkach pierwotniaków, alg kryptofitowych, hydr, gąbek i niektórych płazińce. Nawet chloroplasty poszczególnych glonów syfonowych, m.in. Kod, stają się symbiontami ślimaków nagoskrzelnych. Zwierzęta te żywią się glonami, których chloroplasty pozostają żywotne w komórkach jamy oddechowej, a przy świetle bardzo skutecznie fotosyntetyzują. Na sierści ssaków rozwijają się liczne glony zielone. Endosymbionty podlegające zmianom morfologicznym w porównaniu z przedstawicielami wolno żyjącymi nie tracą zdolności do fotosyntezy i rozmnażania się wewnątrz komórek gospodarza.
Znaczenie gospodarcze. Powszechne występowanie glonów zielonych decyduje o ich ogromnym znaczeniu w biosferze i działalności gospodarczej człowieka. Ze względu na zdolność do fotosyntezy tak jest główni producenci ogromna ilość materia organiczna w zbiornikach wodnych, które są szeroko stosowane przez zwierzęta i ludzi. Pochłaniając z wody dwutlenek węgla, zielone algi nasycają ją tlenem, niezbędnym wszystkim organizmom żywym. Ich rola w biologicznym cyklu substancji jest ogromna. Szybkie rozmnażanie i bardzo wysokie tempo asymilacji (około 3-5 razy wyższe niż u roślin lądowych) powodują, że masa glonów zwiększa się ponad 10 razy dziennie. Jednocześnie w komórkach chlorelli gromadzą się węglowodany (w szczepach selekcji ich zawartość sięga 60%), lipidy (do 85%), witaminy B, C i K. Białko chlorelli, które może stanowić nawet 50% suchej masy masa komórki, zawiera wszystkie niezbędne aminokwasy. Unikalna zdolność gatunku Chlorella Przyswojenie od 10 do 18% energii świetlnej (w porównaniu z 1–2% w roślinach lądowych) pozwala na wykorzystanie tej zielonej algi do regeneracji powietrza w zamkniętych biologicznych systemach podtrzymywania życia człowieka podczas długotrwałych lotów kosmicznych i nurkowania.
Wiele gatunków zielonych alg wykorzystuje się jako organizmy wskaźnikowe w systemie monitorowania ekosystemów wodnych. Oprócz fototroficznego sposobu odżywiania wiele jednokomórkowych zielonych alg (chlamydomonas) jest w stanie wchłaniać przez skorupę substancje organiczne rozpuszczone w wodzie, co przyczynia się do aktywnego oczyszczania zanieczyszczonych wód, w których rozwijają się te gatunki. Dlatego się je wykorzystuje Dla czyszczenie i obróbka końcowa zanieczyszczonej wody , a także jak karmić w zbiornikach rybackich.
Niektóre rodzaje zielonych alg są wykorzystywane przez ludność kilku krajów dla jedzenia. Na przykład do celów spożywczych w Japonii gatunki z tego rodzaju są specjalnie uprawiane Ulwa. Wodorosty te są szeroko stosowane, zwłaszcza w krajach Azji Południowo-Wschodniej, pod nazwą Sea Salad. Ulvaceae mają zauważalnie wyższą zawartość białka (do 20%) niż inne rodzaje glonów. Niektóre rodzaje zielonych alg są stosowane w jako producenci substancji fizjologicznie czynnych. Glony zielone są dobrym obiektem modelowym do różnorodnych badań biologicznych. Gatunki Hematococcus hoduje się w celu uzyskania astaksantyny, Botryococcus - w celu uzyskania lipidów. Jednocześnie śmierć ryb wiąże się z „rozkwitem” wody jednego z jezior na Tajwanie, spowodowanym przez Botryococcus.
Rodzaje porodu Chlorella I Chlamydomonas - obiekty modelowe do badania fotosyntezy w komórkach roślinnych. Chlorella, ze względu na bardzo wysokie wskaźniki reprodukcji, jest przedmiotem masowej uprawy z przeznaczeniem na różne pola
Warstwy powierzchniowe zielonych alg mają duże rozmiary wartość przeciwerozyjna. Niektóre jednokomórkowe gatunki zielonych alg, które wydzielają obfite ilości śluzu, mają działanie wiążące. Substancje śluzowe błon komórkowych sklejają ze sobą cząsteczki gleby. Rozwój glonów wpływa na strukturę drobnej ziemi, nadając ją wodoodporność i zapobieganie usuwaniu z warstwy powierzchniowej. Wilgotność gleby pod błonami glonów jest zwykle wyższa niż tam, gdzie ich nie ma. Ponadto folie zmniejszają przepuszczalność gleby i spowalniają parowanie wody, co również wpływa na reżim zasolenia gleby. Zmniejsza się wymywanie łatwo rozpuszczalnych soli z gleby; ich zawartość pod makroporostami glonów jest wyższa niż na innych obszarach. Jednocześnie spowalnia się wypływ soli z głębokich warstw gleby.
Glony glebowe wpływają także na wzrost i rozwój roślin wyższych. Uwalniając fizjologicznie substancje czynne przyspieszają wzrost sadzonek, zwłaszcza ich korzeni.
Wśród zielonych alg żyjących w wodach zanieczyszczonych dominują glony chlorokokowe, odporne na długotrwałe narażenie na wiele substancji toksycznych.
Komórki glonów są zdolne do gromadzenia różnych pierwiastki chemiczne, a ich stopa akumulacji jest dość wysoka. Słodkowodne algi zielone, zwłaszcza algi nitkowate, są silnymi koncentratorami. Jednocześnie intensywność akumulacji metali jest w nich znacznie wyższa niż w innych słodkowodnych organizmach wodnych. Bardzo interesująca jest zdolność glonów do koncentracji pierwiastków radioaktywnych. Martwe komórki glonów zatrzymują nagromadzone pierwiastki nie mniej mocno niż żywe, a w niektórych przypadkach desorpcja z martwych komórek jest mniejsza niż z żywych. Zdolność wielu rodzajów ( Chlorella, Scenedesmus itp.) koncentrują i mocno zatrzymują w swoich komórkach pierwiastki chemiczne i radionuklidy, co pozwala na ich wykorzystanie w wyspecjalizowanych systemach oczyszczania odkażenie przemysłowy Ścieki na przykład do dodatkowego oczyszczania niskoaktywnych ścieków z elektrowni jądrowych.
Niektóre zielone glony są antagoniści wirusa grypy, wirusa polio itp. Ważną rolę odgrywają substancje biologicznie czynne uwalniane przez glony dezynfekcja wody i tłumienie aktywności patogennej mikroflory.
W specjalnych stawach biologicznych korzystają zbiorowiska glonów i bakterii do rozkładu i detoksykacji herbicydów. Udowodniono zdolność wielu zielonych alg do hydrolizy herbicydu propanilu, który jest szybciej niszczony przez bakterie.
Pytania kontrolne
Wymień charakterystyczne cechy budowy komórek zielonych alg.
Jakie pigmenty i rodzaje składników odżywczych są znane w zielonych algach?
Jak rozmnażają się zielone algi? Co to są zoospory, aplanospory i autospory?
Jakie są klasy glonów zielonych?
Wymień charakterystyczne cechy zielonych alg z klasy Ulvophyceae.
Wymień charakterystyczne cechy zielonych alg z klasy Bryopsidae.
Wymień charakterystyczne cechy zielonych alg z klasy Chlorophyceae.
Wymień charakterystyczne cechy zielonych alg z klasy Trebuxiaceae.
Wymień charakterystyczne cechy zielonych alg z klasy Prasin.
W jakich siedliskach występują zielone algi? Opisz ich główne grupy ekologiczne.
Rola i znaczenie zielonych glonów w przyrodzie.
Jakie to jest znaczenie gospodarcze zielone algi?
Co to jest „rozkwit wody”? Udział zielonych glonów w biologicznym oczyszczaniu wody.
Zielone algi jako nietradycyjne źródła energii.
Glony, jednokomórkowe i wielokomórkowe formy glonów bentosowych. Występują tu wszystkie typy morfologiczne plechy, z wyjątkiem jednokomórkowych ryzopodialnych i dużych form wielokomórkowych z złożona struktura. Wiele nitkowatych zielone algi Przyczepiają się do podłoża dopiero we wczesnych stadiach rozwoju, następnie stają się wolno żyjące, tworząc maty lub kule.
| Zielone algi | ||||||||||||
| Różnorodność glonów syfonowych. Ilustracja z książki Ernsta Haeckela Kunstformen der Natur, 1904 |
||||||||||||
| Klasyfikacja naukowa | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Międzynarodowa nazwa naukowa | ||||||||||||
|
Chlorofit Paschera, 1914 |
||||||||||||
|
||||||||||||
Najbardziej rozległe są glony zielone dany czas dział glonów: według przybliżonych szacunków obejmuje od 13 000 do 20 000 gatunków. Wszystkie wyróżniają się przede wszystkim czysto zielonym kolorem plech, zbliżonym do koloru roślin wyższych i spowodowanym przewagą chlorofilu nad innymi pigmentami.
Struktura
Komórki wici zielonych alg są izokontami - wici mają podobną strukturę, chociaż mogą różnić się długością. Zwykle są dwie wici, ale może być ich również cztery lub wiele. Wici zielonych alg nie mają mastigonemów (w przeciwieństwie do heterokontów), ale mogą mieć wdzięczne włosy lub łuski.
Cykle życia
Cykle życiowe zielonych alg są bardzo zróżnicowane. Jest tu cała gama typów.
Haplobiont z redukcją zygotyczną ( Hydrodictyon reticulatum, Eudorina). Gamety dwuwiciowe są uwalniane z komórki rodzicielskiej przez pory w jej błonie, a fuzja gamet odbywa się za pomocą rurki. Następnie zygota przekształca się w zygosporę spoczynkową, która po okresie fizjologicznego spoczynku kiełkuje, tworząc 4 zoospory (w wyniku podziału mejotycznego). Każda zoospora tworzy wielościan i kiełkuje, tworząc małe kuliste sieci przylegających zoospor.
Haplo-diplobiont z redukcją sporadyczną ( Ulwa, Ulothrix, niektóre typy Kladofora). Izogamety biflagellatowe wyłaniają się z komórki macierzystej, po czym gamety utworzone przez różne włókna łączą się w wodzie. Tworzy się zygota z czterema wiciowcami, która aktywnie unosi się w wodzie. Następnie opada na jakieś podłoże i pokrywa się gęstą skorupą, przekształcając się w komórkę maczugowatą (codiolum), po czym następuje etap fizjologicznego odpoczynku. W sprzyjających warunkach kiełkuje w 4-16 zoospor lub aplanospor, które po krótkim okresie pływania przyczepiają się do podłoża i wyrastają w nowe włókna. Różne czynniki aktywują wyjście ze stanu uśpienia: wzrost temperatury, zmiana otoczenia itp.
Diplobiont z redukcją gamet ( Bryopsis). Planozygota osiada i wyrasta w nitkowatą plechę z dużym jądrem; jądro dzieli się, tworząc zoospory stephanokont, które wyrastają na plechę wegetatywną.
Szczególnie dużo zielonych glonów rozwija się wiosną, kiedy wszystkie kamienie w strefie przybrzeżnej pokryte są ciągłą szmaragdową powłoką zielonych alg, ostro kontrastującą z białym śniegiem zalegającym na przybrzeżnych kamieniach. W wyniku rozwijających się włókienek - ulotrix ( Ulothrix) i urospora ( Urospora). Latem często rozwija się dużo Egagropyli ( Aegagropila linnaei) (syn. Cladophora aegagropila), która często wygląda jak zielona masa śluzowa. Na otwartym skalistym wybrzeżu jasnozielone, rozgałęzione krzewy tworzą akrosyfonię ( Akrosyfonia).
Rola w przyrodzie i użytkowaniu
Niektóre zielone algi (takie jak ulva) są powszechnie spożywane. Chlorella służy jako wskaźnik poziomu zanieczyszczenia wody i jest zawarta w: statki kosmiczne, łodzie podwodne do oczyszczania powietrza z dwutlenku węgla.







