Wiewiórki- wysoka masa cząsteczkowa związki organiczne, składający się z reszt α-aminokwasowych.
W skład białka obejmuje węgiel, wodór, azot, tlen, siarkę. Niektóre białka tworzą kompleksy z innymi cząsteczkami zawierającymi fosfor, żelazo, cynk i miedź.
Białka mają dużą masę cząsteczkową: albumina jaja - 36 000, hemoglobina - 152 000, miozyna - 500 000. Dla porównania: masa cząsteczkowa alkoholu wynosi 46, kwas octowy- 60, benzen - 78.
Skład aminokwasowy białek
Wiewiórki- polimery nieokresowe, których monomerami są α-aminokwasy. Zwykle 20 rodzajów α-aminokwasów nazywa się monomerami białkowymi, chociaż ponad 170 z nich występuje w komórkach i tkankach.
W zależności od tego, czy aminokwasy mogą być syntetyzowane w organizmie człowieka i innych zwierząt, wyróżnia się je: aminokwasy nieistotne- można syntetyzować; aminokwasy- nie można syntetyzować. Aminokwasy musi przedostać się do organizmu wraz z pożywieniem. Rośliny syntetyzują wszystkie rodzaje aminokwasów.
W zależności od składu aminokwasów białka są: kompletne- zawierają cały zestaw aminokwasów; wadliwy- w ich składzie brakuje niektórych aminokwasów. Jeśli białka składają się wyłącznie z aminokwasów, nazywa się je prosty. Jeśli białka zawierają oprócz aminokwasów składnik nieaminokwasowy (grupę prostetyczną), nazywa się je złożony. Grupę prostetyczną mogą reprezentować metale (metaloproteiny), węglowodany (glikoproteiny), lipidy (lipoproteiny), kwasy nukleinowe (nukleoproteiny).
Wszystko zawierają aminokwasy: 1) grupa karboksylowa (-COOH), 2) grupa aminowa (-NH2), 3) rodnik lub grupa R (reszta cząsteczki). Struktura rodnika różne rodzaje aminokwasy - różne. W zależności od liczby grup aminowych i grup karboksylowych wchodzących w skład aminokwasów wyróżnia się: neutralne aminokwasy posiadający jedną grupę karboksylową i jedną grupę aminową; podstawowe aminokwasy posiadający więcej niż jedną grupę aminową; aminokwasy kwasowe mające więcej niż jedną grupę karboksylową.
Aminokwasy są związki amfoteryczne , ponieważ w roztworze mogą działać zarówno jako kwasy, jak i zasady. W roztwory wodne aminokwasy występują w różnych formach jonowych.
Wiązanie peptydowe
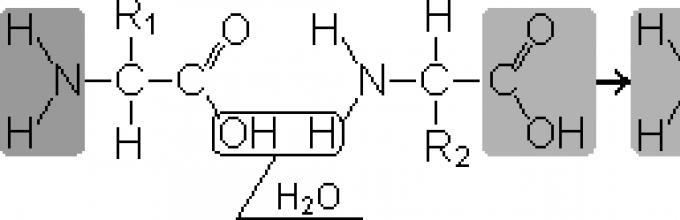
Peptydy- substancje organiczne składające się z reszt aminokwasowych połączonych wiązaniami peptydowymi.
Tworzenie peptydów następuje w wyniku reakcji kondensacji aminokwasów. Kiedy grupa aminowa jednego aminokwasu oddziałuje z grupą karboksylową innego, powstaje między nimi kowalencyjne wiązanie azot-węgiel, tzw. peptyd. W zależności od liczby reszt aminokwasowych zawartych w peptydzie istnieją dipeptydy, tripeptydy, tetrapeptydy itp. Tworzenie wiązania peptydowego można powtarzać wielokrotnie. Prowadzi to do formacji polipeptydy. Na jednym końcu peptydu znajduje się wolna grupa aminowa (zwana końcem N), a na drugim wolna grupa karboksylowa (zwana końcem C).

Organizacja przestrzenna cząsteczek białek
Spełnianie przez białka pewnych specyficznych funkcji zależy od przestrzennej konfiguracji ich cząsteczek, ponadto energetycznie niekorzystne dla komórki jest utrzymywanie białek w postaci niezłożonej, w postaci łańcucha, dlatego łańcuchy polipeptydowe ulegają fałdowaniu, uzyskując pewna trójwymiarowa struktura lub konformacja. Istnieją 4 poziomy organizacja przestrzenna białka.
Struktura pierwotna wiewiórka- kolejność ułożenia reszt aminokwasowych w łańcuchu polipeptydowym tworzącym cząsteczkę białka. Wiązanie między aminokwasami jest wiązaniem peptydowym.

Jeśli cząsteczka białka składa się tylko z 10 reszt aminokwasowych, liczba ta jest teoretycznie możliwe opcje cząsteczki białka różniące się kolejnością naprzemienności aminokwasów - 10 20. Mając 20 aminokwasów, można z nich tworzyć jeszcze bardziej różnorodne kombinacje. W organizmie człowieka znaleziono około dziesięciu tysięcy różnych białek, które różnią się zarówno między sobą, jak i od białek innych organizmów.
To pierwotna struktura cząsteczki białka określa właściwości cząsteczek białka i ich konfigurację przestrzenną. Zastąpienie jednego aminokwasu innym w łańcuchu polipeptydowym prowadzi do zmiany właściwości i funkcji białka. Na przykład zastąpienie szóstego aminokwasu glutaminowy waliną w podjednostce β hemoglobiny prowadzi do tego, że cząsteczka hemoglobiny jako całość nie może pełnić swojej głównej funkcji - transportu tlenu; W takich przypadkach u osoby rozwija się choroba zwana anemią sierpowatokrwinkową.
Struktura wtórna- uporządkowane złożenie łańcucha polipeptydowego w spiralę (wygląda jak rozciągnięta sprężyna). Zwoje helisy są wzmocnione wiązaniami wodorowymi, które powstają pomiędzy grupami karboksylowymi i grupami aminowymi. Prawie wszystkie grupy CO i NH biorą udział w tworzeniu wiązań wodorowych. Są słabsze od peptydowych, ale wielokrotnie powtarzane, nadają tej konfiguracji stabilność i sztywność. Na poziomie struktury wtórnej znajdują się białka: fibroina (jedwab, pajęczyna), keratyna (włosy, paznokcie), kolagen (ścięgna).



Struktura trzeciorzędowa- upakowanie łańcuchów polipeptydowych w kuleczki powstałe w wyniku ich wystąpienia wiązania chemiczne(wodór, jonowy, dwusiarczkowy) i powstawanie oddziaływań hydrofobowych pomiędzy rodnikami reszt aminokwasowych. Główną rolę w tworzeniu struktury trzeciorzędowej odgrywają oddziaływania hydrofilowo-hydrofobowe. W roztworach wodnych rodniki hydrofobowe mają tendencję do ukrywania się przed wodą, grupując się wewnątrz globuli, natomiast rodniki hydrofilowe w wyniku hydratacji (oddziaływania z dipolami wody) mają tendencję do pojawiania się na powierzchni cząsteczki. W niektórych białkach struktura trzeciorzędowa jest stabilizowana przez dwusiarczkowe wiązania kowalencyjne utworzone pomiędzy atomami siarki dwóch reszt cysteiny. Na poziomie struktury trzeciorzędowej znajdują się enzymy, przeciwciała i niektóre hormony.

Struktura czwartorzędowa charakterystyczne dla złożonych białek, których cząsteczki są utworzone przez dwie lub więcej kulek. Podjednostki są utrzymywane w cząsteczce poprzez oddziaływania jonowe, hydrofobowe i elektrostatyczne. Czasami podczas tworzenia struktury czwartorzędowej między podjednostkami występują wiązania dwusiarczkowe. Najbardziej zbadanym białkiem o strukturze czwartorzędowej jest hemoglobina. Tworzą go dwie podjednostki α (141 reszt aminokwasowych) i dwie podjednostki β (146 reszt aminokwasowych). Z każdą podjednostką związana jest cząsteczka hemu zawierająca żelazo.
Jeżeli z jakiegoś powodu konformacja przestrzenna białek odbiega od normy, białko nie może pełnić swoich funkcji. Na przykład przyczyną „choroby szalonych krów” (encefalopatii gąbczastej) jest nieprawidłowa konformacja prionów, białek powierzchniowych komórek nerwowych.
Właściwości białek
Decyduje o tym skład aminokwasowy i struktura cząsteczki białka nieruchomości. Białka łączą właściwości zasadowe i kwasowe, określone przez rodniki aminokwasowe: im więcej aminokwasów kwasowych w białku, tym wyraźniejsze są jego właściwości kwasowe. Określana jest zdolność do oddawania i dodawania H+ właściwości buforujące białek; Jednym z najsilniejszych buforów jest hemoglobina zawarta w czerwonych krwinkach, która utrzymuje pH krwi na stałym poziomie. Istnieją białka rozpuszczalne (fibrynogen) i białka nierozpuszczalne, które pełnią funkcje mechaniczne (fibroina, keratyna, kolagen). Istnieją białka chemicznie aktywne (enzymy), są białka chemicznie nieaktywne, odporne na różne warunki środowiskowe i takie, które są wyjątkowo niestabilne.
Czynniki zewnętrzne (ciepło, promieniowanie ultrafioletowe, metale ciężkie i ich sole, zmiany pH, promieniowanie, odwodnienie)
może powodować zaburzenie organizacji strukturalnej cząsteczki białka. Nazywa się proces utraty trójwymiarowej konformacji właściwej danej cząsteczce białka denaturacja. Przyczyną denaturacji jest zerwanie wiązań stabilizujących określoną strukturę białka. Początkowo zrywane są najsłabsze więzi, a w miarę zaostrzania warunków zrywane są nawet silniejsze. Dlatego najpierw tracone są struktury czwartorzędowe, potem trzeciorzędowe i wtórne. Zmiana konfiguracji przestrzennej prowadzi do zmiany właściwości białka i w efekcie uniemożliwia mu pełnienie przyrodzonych mu funkcji biologicznych. Jeśli denaturacji nie towarzyszy zniszczenie struktury pierwotnej, może tak być odwracalny w tym przypadku następuje samoodzyskiwanie charakterystycznej konformacji białka. Takiej denaturacji ulegają na przykład białka receptorów błonowych. Nazywa się proces przywracania struktury białka po denaturacji renaturyzacja. Jeżeli przywrócenie konfiguracji przestrzennej białka nie jest możliwe, wówczas nazywa się denaturację nieodwracalny.
Funkcje białek
| Funkcjonować | Przykłady i wyjaśnienia |
|---|---|
| Budowa | Białka biorą udział w tworzeniu struktur komórkowych i zewnątrzkomórkowych: są częścią błon komórkowych (lipoproteiny, glikoproteiny), włosów (keratyna), ścięgien (kolagen) itp. |
| Transport | Białko krwi, hemoglobina, wiąże tlen i transportuje go z płuc do wszystkich tkanek i narządów, a z nich przenosi dwutlenek węgla do płuc; W skład błon komórkowych wchodzą specjalne białka, które zapewniają aktywne i ściśle selektywne przenoszenie określonych substancji i jonów z komórki do środowiska zewnętrznego i z powrotem. |
| Regulacyjne | Hormony białkowe biorą udział w regulacji procesów metabolicznych. Na przykład hormon insuliny reguluje poziom glukozy we krwi, wspomaga syntezę glikogenu i zwiększa powstawanie tłuszczów z węglowodanów. |
| Ochronny | W odpowiedzi na przenikanie obcych białek lub mikroorganizmów (antygenów) do organizmu powstają specjalne białka – przeciwciała, które mogą je wiązać i neutralizować. Fibryna utworzona z fibrynogenu pomaga zatrzymać krwawienie. |
| Silnik | Białka kurczliwe, aktyna i miozyna, zapewniają skurcz mięśni u zwierząt wielokomórkowych. |
| Sygnał | W powierzchniową błonę komórkową wbudowane są cząsteczki białka, które w odpowiedzi na czynniki środowiskowe mają zdolność zmiany swojej trzeciorzędowej struktury, odbierając w ten sposób sygnały ze środowiska zewnętrznego i przekazując polecenia komórce. |
| Składowanie | W ciele zwierząt białka z reguły nie są magazynowane, z wyjątkiem albuminy jaja i kazeiny mleka. Ale dzięki białkom niektóre substancje mogą być magazynowane w organizmie, na przykład podczas rozkładu hemoglobiny żelazo nie jest usuwane z organizmu, ale jest magazynowane, tworząc kompleks z białkiem ferrytyną. |
| Energia | Kiedy 1 g białka rozpada się na produkty końcowe, uwalniane jest 17,6 kJ. Najpierw białka rozkładają się na aminokwasy, a następnie na produkty końcowe – wodę, dwutlenek węgla i amoniak. Jednakże białka są wykorzystywane jako źródło energii tylko wtedy, gdy zostaną wykorzystane inne źródła (węglowodany i tłuszcze). |
| Katalityczny | Jedna z najważniejszych funkcji białek. Dostarczane przez białka - enzymy przyspieszające reakcje biochemiczne zachodzące w komórkach. Na przykład karboksylaza wodorofosforanu rybulozy katalizuje wiązanie CO2 podczas fotosyntezy. |
Enzymy
Enzymy, Lub enzymy, stanowią specjalną klasę białek będących katalizatorami biologicznymi. Dzięki enzymom reakcje biochemiczne zachodzą z ogromną szybkością. Szybkość reakcji enzymatycznych jest dziesiątki tysięcy (a czasem miliony) większa niż szybkość reakcji zachodzących przy udziale katalizatorów nieorganicznych. Substancja, na którą działa enzym, nazywa się podłoże.
Enzymy to białka kuliste, cechy konstrukcyjne Enzymy można podzielić na dwie grupy: proste i złożone. Proste enzymy są białkami prostymi, tj. składają się wyłącznie z aminokwasów. Złożone enzymy są białkami złożonymi, tj. Oprócz części białkowej zawierają grupę o charakterze niebiałkowym - kofaktor. Niektóre enzymy wykorzystują witaminy jako kofaktory. Cząsteczka enzymu zawiera specjalną część zwaną centrum aktywnym. Aktywny ośrodek- niewielka część enzymu (od trzech do dwunastu reszt aminokwasowych), w której następuje wiązanie substratu lub substratów z utworzeniem kompleksu enzym-substrat. Po zakończeniu reakcji kompleks enzym-substrat rozpada się na enzym i produkt(y) reakcji. Niektóre enzymy mają (z wyjątkiem aktywnych) centra allosteryczne- obszary, do których przyłączone są regulatory szybkości enzymów ( enzymy allosteryczne).

Reakcje katalizy enzymatycznej charakteryzują się: 1) wysoką wydajnością, 2) ścisłą selektywnością i kierunkiem działania, 3) specyficznością substratową, 4) dokładną i precyzyjną regulacją. Specyficzność substratową i reakcyjną reakcji katalizy enzymatycznej wyjaśniają hipotezy E. Fischera (1890) i D. Koshlanda (1959).
E. Fisher (hipoteza zamka na klucz) zasugerowali, że konfiguracje przestrzenne centrum aktywnego enzymu i substratu muszą dokładnie sobie odpowiadać. Substrat porównywany jest do „klucza”, enzym do „zamka”.
D. Koshland (hipoteza rękawicy) zasugerowali, że zgodność przestrzenna pomiędzy strukturą substratu i centrum aktywnego enzymu powstaje dopiero w momencie ich wzajemnego oddziaływania. Ta hipoteza jest również nazywana hipoteza indukowanej korespondencji.
Szybkość reakcji enzymatycznych zależy od: 1) temperatury, 2) stężenia enzymu, 3) stężenia substratu, 4) pH. Należy podkreślić, że ponieważ enzymy są białkami, ich aktywność jest najwyższa pod względem fizjologicznym normalne warunki.
Większość enzymów może działać tylko w temperaturach od 0 do 40°C. W tych granicach szybkość reakcji wzrasta około 2 razy na każde 10°C wzrostu temperatury. W temperaturach powyżej 40°C białko ulega denaturacji i aktywność enzymu maleje. W temperaturach bliskich zamarzania enzymy ulegają inaktywacji.
Wraz ze wzrostem ilości substratu szybkość reakcji enzymatycznej wzrasta, aż liczba cząsteczek substratu zrówna się z liczbą cząsteczek enzymu. Przy dalszym wzroście ilości substratu prędkość nie wzrośnie, ponieważ centra aktywne enzymu są nasycone. Wzrost stężenia enzymu prowadzi do zwiększonej aktywności katalitycznej, gdyż przemianie ulega większa liczba cząsteczek substratu w jednostce czasu.

Dla każdego enzymu istnieje optymalna wartość pH, przy której wykazuje on maksymalną aktywność (pepsyna – 2,0, amylaza ślinowa – 6,8, lipaza trzustkowa – 9,0). Przy wyższych lub niższych wartościach pH aktywność enzymu maleje. Przy nagłych zmianach pH enzym ulega denaturacji.
Szybkość enzymów allosterycznych jest regulowana przez substancje przyłączające się do centrów allosterycznych. Jeżeli substancje te przyspieszają reakcję, nazywa się je aktywatory, jeśli zwolnią - inhibitory.
Klasyfikacja enzymów
Ze względu na rodzaj przemian chemicznych, które katalizują, enzymy dzielą się na 6 klas:
- oksyreduktazy(przeniesienie atomów wodoru, tlenu lub elektronów z jednej substancji na drugą – dehydrogenaza),
- transferazy(przeniesienie grupy metylowej, acylowej, fosforanowej lub aminowej z jednej substancji na drugą – transaminaza),
- hydrolazy(reakcje hydrolizy, podczas których z substratu powstają dwa produkty – amylaza, lipaza),
- liazy(niehydrolityczny dodatek do substratu lub oderwanie od niego grupy atomów, w którym to przypadku można rozerwać wiązania C-C, C-N, C-O, C-S - dekarboksylaza),
- izomerazy(przegrupowanie wewnątrzcząsteczkowe – izomeraza),
- ligazy(połączenie dwóch cząsteczek w wyniku powstawania Połączenia CC, C-N, C-O, C-S - syntetaza).
Klasy dzielą się z kolei na podklasy i podklasy. W aktualnej klasyfikacja międzynarodowa Każdy enzym ma specyficzny kod, składający się z czterech liczb oddzielonych kropkami. Pierwsza liczba to klasa, druga to podklasa, trzecia to podklasa, czwarta to numer seryjny enzym w tej podklasie, na przykład kod arginazy to 3.5.3.1.
Iść do wykłady nr 2„Budowa i funkcje węglowodanów i lipidów”
Iść do wykłady nr 4„Struktura i funkcje kwasy nukleinowe ATP”
Krótki opis:
Fragment podręcznika: Chemia biologiczna z ćwiczeniami i zadaniami: podręcznik / wyd. Członek korespondent RAMS SE Seweryna. M.: GEOTAR-Media, 2011. - 624 s.: il. MODUŁ 1: STRUKTURA, WŁAŚCIWOŚCI I FUNKCJE BIAŁEK
MODUŁ 1: STRUKTURA, WŁAŚCIWOŚCI I FUNKCJE BIAŁEK
Struktura modułu | Motywy |
Jednostka modułowa 1 | 1.1. Strukturalna organizacja białek. Etapy tworzenia konformacji białka natywnego 1.2. Podstawy funkcjonowania białek. Leki jako ligandy wpływające na funkcję białek 1.3. Denaturacja białek i możliwość ich samoistnej renatywacji |
Jednostka modułowa 2 | 1.4. Cechy budowy i funkcjonowania białek oligomerycznych na przykładzie hemoglobiny 1,5. Utrzymanie konformacji białka natywnego w warunkach komórkowych 1.6. Różnorodność białek. Rodziny białek na przykładzie immunoglobulin 1.7. Właściwości fizykochemiczne białek i metody ich rozdzielania |
Jednostka modułowa 1 ORGANIZACJA STRUKTURALNA BIAŁEK MONOMERYCZNYCH I PODSTAWY ICH DZIAŁANIA
Cele nauczania Być w stanie:
1. Wykorzystywać wiedzę o cechach strukturalnych białek i zależności funkcji białek od ich budowy do zrozumienia mechanizmów rozwoju proteinopatii dziedzicznych i nabytych.
2. Wyjaśniać mechanizmy działania terapeutycznego niektórych leków jako ligandów oddziałujących z białkami i zmieniających ich działanie.
3. Wykorzystywać wiedzę o strukturze i labilności konformacyjnej białek do zrozumienia ich niestabilności strukturalnej i funkcjonalnej oraz tendencji do denaturacji w zmieniających się warunkach.
4. Wyjaśniać zastosowanie środków denaturujących do sterylizacji materiałów i narzędzi medycznych oraz środków antyseptycznych.
Wiedzieć:
1. Poziomy organizacji strukturalnej białek.
2. Znaczenie struktury pierwszorzędowej białek, która decyduje o ich różnorodności strukturalnej i funkcjonalnej.
3. Mechanizm powstawania centrum aktywnego w białkach i jego specyficzne oddziaływanie z ligandem, które leży u podstaw funkcjonowania białek.
4. Przykłady wpływu egzogennych ligandów (leków, toksyn, trucizn) na konformację i aktywność funkcjonalną białek.
5. Przyczyny i skutki denaturacji białek, czynniki powodujące denaturację.
6. Przykłady zastosowania czynników denaturujących w medycynie jako środków antyseptycznych i środków do sterylizacji narzędzi medycznych.
TEMAT 1.1. ORGANIZACJA STRUKTURALNA BIAŁEK. ETAPY FORMOWANIA RODZIMYCH
KONFORMACJE BIAŁKOWE
Wiewiórki - są to heteropolimery Cząsteczki (tj. składający się z różnych monomerów). Monomery białkowe to 20 rodzajów α-aminokwasów połączonych ze sobą wiązaniami peptydowymi.
Zestaw i kolejność kombinacji aminokwasów w białku jest określona przez strukturę odpowiednich genów w DNA poszczególnych osób. Każde białko, zgodnie ze swoją specyficzną budową, pełni swoją funkcję. Zbiór białek danego organizmu ( proteom) decyduje o jego cechach fenotypowych, a także o występowaniu chorób dziedzicznych lub predyspozycji do ich rozwoju.
1. Aminokwasy tworzące białka. Wiązanie peptydowe.
Białka to heteropolimery zbudowane z monomerów – 20 α-aminokwasów.
Ogólny wzór aminokwasów przedstawiono poniżej.

Aminokwasy różnią się budową, wielkością i właściwościami fizykochemicznymi rodników przyłączonych do atomu węgla α. Grupy funkcyjne aminokwasów określają charakterystykę właściwości różnych α-aminokwasów. Rodniki występujące w α-aminokwasach można podzielić na kilka grup:
Prolina, W przeciwieństwie do pozostałych 19 monomerów białkowych, nie jest to aminokwas, ale iminokwas; rodnik w prolinie jest związany zarówno z atomem węgla α, jak i grupą iminową
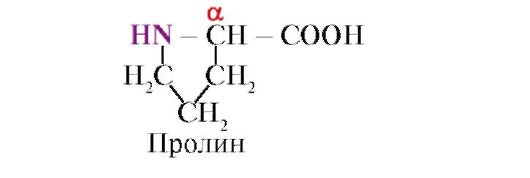
Aminokwasy różnią się rozpuszczalnością w wodzie. Wynika to ze zdolności rodników do interakcji z wodą (hydratem).
DO hydrofilowy obejmują rodniki zawierające nienaładowane anionowe, kationowe i polarne grupy funkcyjne.
DO hydrofobowy obejmują rodniki zawierające grupy metylowe, łańcuchy lub pierścienie alifatyczne.
2. Wiązania peptydowe łączą aminokwasy, tworząc peptydy. Podczas syntezy peptydów grupa α-karboksylowa jednego aminokwasu oddziałuje z grupą α-aminową innego aminokwasu, tworząc wiązanie peptydowe:
Białka są polipeptydami, tj. liniowe polimery α-aminokwasów połączone wiązaniem peptydowym (ryc. 1.1.)
 Ryż. 1.1. Terminy używane do opisu struktury peptydów
Ryż. 1.1. Terminy używane do opisu struktury peptydów
Nazywa się monomery aminokwasów tworzące polipeptydy reszty aminokwasowe.Łańcuch powtarzających się grup - NH-CH-CO- formy szkielet peptydowy. Resztę aminokwasu zawierającą wolną grupę α-aminową nazywa się N-końcową, a resztę zawierającą wolną grupę α-karboksylową nazywa się C-końcową. Peptydy są zapisywane i odczytywane od N-końca do C-końca.
Wiązanie peptydowe utworzone przez grupę iminową proliny różni się od innych wiązań peptydowych: atom azotu grupy peptydowej nie zawiera wodoru,
zamiast tego dochodzi do wiązania z rodnikiem, w wyniku czego jedna strona pierścienia zostaje włączona do szkieletu peptydowego:

Peptydy różnią się składem aminokwasów, liczbą aminokwasów i kolejnością połączeń aminokwasów, na przykład Ser-Ala-Glu-Gis i His-Glu-Ala-Ser to dwa różne peptydy.
Wiązania peptydowe są bardzo mocne, a ich chemiczna, nieenzymatyczna hydroliza wymaga rygorystycznych warunków: analizowane białko ulega hydrolizie w stężonym kwas chlorowodorowy w temperaturze około 110° przez 24 godziny. W żywej komórce Wiązania peptydowe można rozerwać Enzymy proteolityczne, zwany proteazy Lub hydrolazy peptydowe.
3. Podstawowa struktura białek. Reszty aminokwasowe w łańcuchach peptydowych różnych białek nie zmieniają się losowo, ale są ułożone w określonej kolejności. Nazywa się sekwencją liniową lub kolejnością naprzemienną reszt aminokwasowych w łańcuchu polipeptydowym pierwotna struktura białka.
Podstawowa struktura każdego pojedynczego białka jest kodowana w cząsteczce DNA (w regionie zwanym genem) i jest realizowana podczas transkrypcji (kopiowanie informacji na mRNA) i translacji (synteza pierwszorzędowej struktury białka). Zatem pierwotną strukturą białek pojedynczego człowieka jest informacja przekazywana dziedzicznie z rodziców na dzieci, która określa cechy strukturalne białek danego organizmu, od których zależy funkcja istniejących białek (ryc. 1.2.).
 Ryż. 1.2. Związek pomiędzy genotypem a konformacją białek syntetyzowanych w organizmie człowieka
Ryż. 1.2. Związek pomiędzy genotypem a konformacją białek syntetyzowanych w organizmie człowieka
Każde z około 100 000 pojedynczych białek w organizmie człowieka ma unikalny struktura pierwotna. Cząsteczki tego samego rodzaju białka (na przykład albuminy) mają tę samą przemianę reszt aminokwasowych, co odróżnia albuminę od innych pojedynczych białek.
Sekwencję reszt aminokwasowych w łańcuchu peptydowym można uznać za formę zapisu informacji. Informacje te określają przestrzenne rozmieszczenie liniowego łańcucha peptydowego w bardziej zwartą trójwymiarową strukturę zwaną struktura wiewiórka. Nazywa się proces tworzenia funkcjonalnie aktywnej konformacji białka składanie
4. Konformacja białka. Swobodna rotacja w szkielecie peptydowym jest możliwa pomiędzy atomem azotu grupy peptydowej a sąsiadującym atomem węgla α, a także pomiędzy atomem węgla α a węglem grupy karbonylowej. Dzięki oddziaływaniu grup funkcyjnych reszt aminokwasowych pierwotna struktura białek może nabrać bardziej złożonych struktur przestrzennych. W białkach globularnych istnieją dwa główne poziomy fałdowania konformacji łańcuchów peptydowych: wtórny I struktura trzeciorzędowa.
Struktura wtórna białek to struktura przestrzenna powstająca w wyniku utworzenia wiązań wodorowych pomiędzy grupami funkcyjnymi -C=O i -NH- szkieletu peptydowego. W tym przypadku łańcuch peptydowy może uzyskać regularne struktury dwóch typów: α-helisy I Struktury β.
W α-helisy wiązania wodorowe powstają między atomem tlenu grupy karbonylowej a wodorem azotu amidowego czwartego aminokwasu; łańcuchy boczne reszt aminokwasowych
znajdują się wzdłuż obwodu spirali, nie uczestnicząc w tworzeniu struktury wtórnej (ryc. 1.3.).
Rodniki masowe lub rodniki niosące równe ładunki zapobiegają tworzeniu się α-helisy. Reszta proliny, która ma strukturę pierścieniową, przerywa α-helisę, ponieważ z powodu braku wodoru przy atomie azotu w łańcuchu peptydowym niemożliwe jest utworzenie wiązania wodorowego. Wiązanie między azotem i atomem węgla α jest częścią pierścienia proliny, więc szkielet peptydowy ulega w tym miejscu wygięciu.
Struktura β powstaje pomiędzy liniowymi regionami szkieletu peptydowego jednego łańcucha polipeptydowego, tworząc w ten sposób złożone struktury. Mogą tworzyć się łańcuchy polipeptydowe lub ich części równoległy Lub antyrównoległe struktury β. W pierwszym przypadku N- i C-końce oddziałujących łańcuchów peptydowych pokrywają się, w drugim mają przeciwny kierunek (ryc. 1.4).
Ryż. 1.3. Struktura drugorzędowa białka - α-helisa
 Ryż. 1.4. Struktury β-arkuszowe równoległe i antyrównoległe
Ryż. 1.4. Struktury β-arkuszowe równoległe i antyrównoległe
Struktury β zaznaczono szerokimi strzałkami: A – Struktura β antyrównoległa. B - Równoległe struktury β-arkuszowe
W niektórych białkach struktury β mogą powstawać w wyniku tworzenia się wiązań wodorowych pomiędzy atomami szkieletu peptydowego różnych łańcuchów polipeptydowych.
Występuje także w białkach obszary o nieregularnym wtórnym struktura, która obejmuje zagięcia, pętle i zwoje szkieletu polipeptydowego. Często lokalizują się w miejscach, w których zmienia się kierunek łańcucha peptydowego, np. gdy tworzy się równoległa struktura β-kartki.
Na podstawie obecności α-helis i β-struktur białka globularne można podzielić na cztery kategorie.
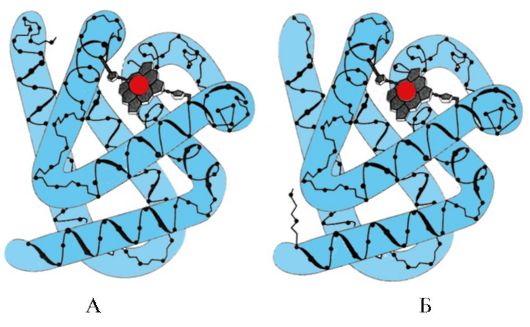
Ryż. 1,5. Wtórna struktura mioglobiny (A) i łańcucha β hemoglobiny (B), zawierająca osiem α-helis
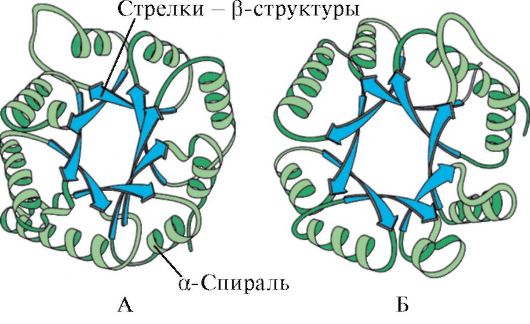 Ryż. 1.6. Struktura drugorzędowa domeny izomerazy triozofosforanowej i kinazy pirogronianowej
Ryż. 1.6. Struktura drugorzędowa domeny izomerazy triozofosforanowej i kinazy pirogronianowej
 Ryż. 1.7. Struktura drugorzędowa domeny stałej immunoglobuliny (A) i enzymu dysmutazy ponadtlenkowej (B)
Ryż. 1.7. Struktura drugorzędowa domeny stałej immunoglobuliny (A) i enzymu dysmutazy ponadtlenkowej (B)
W czwarta kategoria obejmowały białka zawierające niewielką ilość regularnych struktur drugorzędowych. Białka te obejmują małe białka bogate w cysteinę lub metaloproteiny.
Trzeciorzędowa struktura białka- rodzaj konformacji powstający w wyniku oddziaływań pomiędzy rodnikami aminokwasów, które w łańcuchu peptydowym mogą znajdować się w znacznej odległości od siebie. Większość białek tworzy przestrzenną strukturę przypominającą globulę (białka globularne).
Ponieważ hydrofobowe rodniki aminokwasowe mają tendencję do łączenia się poprzez tzw oddziaływania hydrofobowe i międzycząsteczkowe siły van der Waalsa wewnątrz globulki białkowej tworzy się gęsty hydrofobowy rdzeń. Hydrofilowe rodniki zjonizowane i niezjonizowane zlokalizowane są głównie na powierzchni białka i decydują o jego rozpuszczalności w wodzie.
 Ryż. 1.8. Rodzaje wiązań powstających pomiędzy rodnikami aminokwasów podczas tworzenia trzeciorzędowej struktury białka
Ryż. 1.8. Rodzaje wiązań powstających pomiędzy rodnikami aminokwasów podczas tworzenia trzeciorzędowej struktury białka
1 - wiązanie jonowe- występuje pomiędzy dodatnio i ujemnie naładowanymi grupami funkcyjnymi;
2 - wiązanie wodorowe- występuje pomiędzy hydrofilową grupą nienaładowaną i dowolną inną grupą hydrofilową;
3 - oddziaływania hydrofobowe- powstają pomiędzy rodnikami hydrofobowymi;
4 - wiązanie disiarczkowe- powstają w wyniku utleniania grup SH reszt cysteiny i ich wzajemnego oddziaływania
Hydrofilowe reszty aminokwasowe znajdujące się wewnątrz hydrofobowego rdzenia mogą oddziaływać ze sobą za pomocą joński I wiązania wodorowe(ryc. 1.8).
Wiązania jonowe, wodorowe i oddziaływania hydrofobowe są słabe: ich energia jest niewiele wyższa od energii ruchu termicznego cząsteczek w temperatura pokojowa. Konformacja białka jest utrzymywana poprzez tworzenie wielu takich słabych wiązań. Ponieważ atomy tworzące białko są w ciągłym ruchu, możliwe jest rozerwanie niektórych słabych wiązań i utworzenie innych, co prowadzi do nieznacznych ruchów poszczególnych odcinków łańcucha polipeptydowego. Nazywa się tę właściwość białek zmiany konformacji w wyniku rozrywania niektórych i tworzenia innych słabych wiązań labilność konformacyjna.
Ciało ludzkie ma systemy, które wspierają homeostaza- konsystencja środowisko wewnętrzne w pewnych dopuszczalnych granicach dla zdrowego organizmu. W warunkach homeostazy niewielkie zmiany w konformacji nie zakłócają ogólnej struktury i funkcji białek. Nazywa się funkcjonalnie aktywną konformację białka konformacja natywna. Zmiany w środowisku wewnętrznym (na przykład stężenie glukozy, jonów Ca, protonów itp.) prowadzą do zmian w konformacji i zakłócenia funkcji białek.
Trzeciorzędowa struktura niektórych białek jest ustabilizowana wiązania disiarczkowe, powstaje w wyniku oddziaływania grup -SH dwóch reszt
 Ryż. 1.9. Tworzenie wiązania dwusiarczkowego w cząsteczce białka
Ryż. 1.9. Tworzenie wiązania dwusiarczkowego w cząsteczce białka
cysteina (ryc. 1.9). Większość białek wewnątrzkomórkowych nie ma kowalencyjnych wiązań dwusiarczkowych w swojej trzeciorzędowej strukturze. Ich obecność jest charakterystyczna dla białek wydzielanych przez komórkę, co zapewnia ich większą stabilność w warunkach zewnątrzkomórkowych. Zatem wiązania dwusiarczkowe są obecne w cząsteczkach insuliny i immunoglobulin.
Insulina- hormon białkowy syntetyzowany w komórkach β trzustki i wydzielany do krwi w odpowiedzi na wzrost stężenia glukozy we krwi. W strukturze insuliny występują dwa wiązania dwusiarczkowe łączące łańcuchy polipeptydowe A i B oraz jedno wiązanie dwusiarczkowe w obrębie łańcucha A (ryc. 1.10).
 Ryż. 1.10. Wiązania dwusiarczkowe w strukturze insuliny
Ryż. 1.10. Wiązania dwusiarczkowe w strukturze insuliny
5. Nadwtórna struktura białek. Czasami wykrywa się je w białkach o różnej strukturze pierwszorzędowej i funkcjach podobne kombinacje i względne położenia struktur drugorzędnych, które nazywane są strukturą nadwtórną. Zajmuje pozycję pośrednią między strukturami drugorzędowymi i trzeciorzędowymi, ponieważ jest specyficzną kombinacją elementów struktury drugorzędowej w tworzeniu trzeciorzędowej struktury białka. Struktury superwtórne mają specyficzne nazwy, takie jak „α-helisa-zmieniająca helisę”, „zamek leucynowy”, „palce cynkowe” itp. Takie struktury superwtórne są charakterystyczne dla białek wiążących DNA.
„Zamek leucynowy”. Ten typ struktury superwtórnej służy do łączenia ze sobą dwóch białek. Na powierzchni oddziałujących białek znajdują się regiony α-helikalne zawierające co najmniej cztery reszty leucyny. Reszty leucyny w α-helisie są oddalone od siebie o sześć aminokwasów. Ponieważ każdy zwój α-helisy zawiera 3,6 reszt aminokwasowych, rodniki leucyny znajdują się na powierzchni co drugiego zwoju. Reszty leucyny α-helisy jednego białka mogą oddziaływać z resztami leucyny innego białka (oddziaływania hydrofobowe), łącząc je ze sobą (ryc. 1.11.). Wiele białek wiążących DNA funkcjonuje w kompleksach oligomerycznych, w których poszczególne podjednostki są połączone ze sobą „zamkami leucynowymi”.
 Ryż. 1.11. „Zamek leucynowy” pomiędzy regionami α-helikalnymi dwóch białek
Ryż. 1.11. „Zamek leucynowy” pomiędzy regionami α-helikalnymi dwóch białek
Przykładem takich białek są histony. Histony- białka jądrowe, które zawierają dużą liczbę dodatnio naładowanych aminokwasów - argininę i lizynę (do 80%). Cząsteczki histonów łączą się w oligomeryczne kompleksy zawierające osiem monomerów za pomocą „zamków leucynowych”, pomimo znacznego homonimicznego ładunku tych cząsteczek.
„Palec cynkowy”- wariant struktury nadwtórnej, charakterystyczny dla białek wiążących DNA, ma postać wydłużonego fragmentu na powierzchni białka i zawiera około 20 reszt aminokwasowych (ryc. 1.12). Kształt „wydłużonego palca” wspiera atom cynku związany z czterema rodnikami aminokwasowymi – dwiema resztami cysteinowymi i dwiema resztami histydyny. W niektórych przypadkach zamiast reszt histydyny występują reszty cysteiny. Dwie blisko leżące reszty cysteiny są oddzielone od pozostałych dwóch reszt Gisili sekwencją Cys składającą się z około 12 reszt aminokwasowych. Ten region białka tworzy α-helisę, której rodniki mogą specyficznie wiązać się z regionami regulatorowymi głównego rowka DNA. Indywidualna specyficzność wiązania
 Ryż. 1.12. Podstawowa struktura regionu białek wiążących DNA tworzących strukturę „palca cynkowego” (litery wskazują aminokwasy tworzące tę strukturę)
Ryż. 1.12. Podstawowa struktura regionu białek wiążących DNA tworzących strukturę „palca cynkowego” (litery wskazują aminokwasy tworzące tę strukturę)
Białko regulatorowe wiążące DNA zależy od sekwencji reszt aminokwasowych zlokalizowanych w regionie palca cynkowego. Struktury takie zawierają w szczególności receptory dla hormonów steroidowych biorące udział w regulacji transkrypcji (odczycie informacji z DNA na RNA).
TEMAT 1.2. PODSTAWY DZIAŁANIA BIAŁEK. LEKI JAKO LIGANDY WPŁYWAJĄCE NA FUNKCJĘ BIAŁKA
1. Centrum aktywne białka i jego oddziaływanie z ligandem. Podczas tworzenia struktury trzeciorzędowej na powierzchni funkcjonalnie aktywnego białka tworzy się region, zwykle we wnęce utworzonej przez rodniki aminokwasów, które są daleko od siebie oddalone w strukturze pierwszorzędowej. Region ten, który ma unikalną dla danego białka strukturę i jest zdolny do specyficznego oddziaływania z konkretną cząsteczką lub grupą podobnych cząsteczek, nazywany jest miejscem wiązania białko-ligand lub miejscem aktywnym. Ligandy to cząsteczki oddziałujące z białkami.
Wysoka specyficzność Oddziaływanie białka z ligandem zapewnia komplementarność struktury centrum aktywnego do struktury liganda.
Komplementarność- jest to zgodność przestrzenna i chemiczna oddziałujących powierzchni. Centrum aktywne musi nie tylko odpowiadać przestrzennie zawartemu w nim ligandowi, ale także muszą powstać wiązania (oddziaływania jonowe, wodorowe i hydrofobowe) pomiędzy grupami funkcyjnymi rodników wchodzących w skład centrum aktywnego a ligandem, które utrzymują ligand w centrum aktywnym (ryc. 1.13 ).
 Ryż. 1.13. Uzupełniające oddziaływanie białka z ligandem
Ryż. 1.13. Uzupełniające oddziaływanie białka z ligandem
Niektóre ligandy po przyłączeniu do centrum aktywnego białka pełnią pomocniczą rolę w funkcjonowaniu białek. Takie ligandy nazywane są kofaktorami, a białka zawierające część niebiałkową złożone białka(w przeciwieństwie do białek prostych, składających się wyłącznie z części białkowej). Część niebiałkowa, trwale połączona z białkiem, nazywa się grupa protetyczna. Na przykład mioglobina, hemoglobina i cytochromy zawierają grupę prostetyczną, hem, zawierającą jon żelaza, mocno związaną z centrum aktywnym. Złożone białka zawierające hem nazywane są hemoproteinami.
Kiedy do białek przyłączają się specyficzne ligandy, manifestuje się funkcja tych białek. Zatem albumina, najważniejsze białko osocza krwi, spełnia swoją funkcję transportową poprzez przyłączenie do centrum aktywnego ligandów hydrofobowych, takich jak kwasy tłuszczowe, bilirubina, niektóre leki itp. (ryc. 1.14)
Ligandami oddziałującymi z trójwymiarową strukturą łańcucha peptydowego mogą być nie tylko drobnocząsteczkowe cząsteczki organiczne i nieorganiczne, ale także makrocząsteczki:
DNA (przykłady z białkami wiążącymi DNA omówione powyżej);
Polisacharydy;
 Ryż. 1.14. Związek między genotypem a fenotypem
Ryż. 1.14. Związek między genotypem a fenotypem
Unikalna pierwotna struktura białek ludzkich, zakodowana w cząsteczce DNA, realizowana jest w komórkach w postaci unikalnej konformacji, struktury centrum aktywnego i funkcji białek
W takich przypadkach białko rozpoznaje specyficzny region ligandu, który jest proporcjonalny i komplementarny do miejsca wiązania. Zatem na powierzchni hepatocytów znajdują się białka receptorowe dla hormonu insuliny, który również ma strukturę białkową. Oddziaływanie insuliny z receptorem powoduje zmianę jej konformacji i aktywację układów sygnalizacyjnych, co prowadzi do magazynowania składników odżywczych w hepatocytach po posiłkach.
Zatem, Funkcjonowanie białek opiera się na specyficznym oddziaływaniu centrum aktywnego białka z ligandem.
2. Struktura domeny i jej rola w funkcjonowaniu białek. Długie łańcuchy polipeptydowe białek globularnych często składają się na kilka zwartych, stosunkowo niezależnych regionów. Mają niezależną strukturę trzeciorzędową, przypominającą białka globularne i nazywane są domeny. Ze względu na strukturę domenową białek łatwiej jest uformować ich strukturę trzeciorzędową.
W białkach domenowych miejsca wiązania ligandów często znajdują się pomiędzy domenami. Zatem trypsyna jest enzymem proteolitycznym wytwarzanym przez zewnątrzwydzielniczą część trzustki i jest niezbędna do trawienia białek pokarmowych. Ma budowę dwudomenową, a centrum wiązania trypsyny z jej ligandem – białkiem pokarmowym – znajduje się w rowku pomiędzy obiema domenami. W centrum aktywnym powstają warunki niezbędne do skutecznego wiązania określonego miejsca białka spożywczego i hydrolizy jego wiązań peptydowych.
Różne domeny w białku mogą przemieszczać się względem siebie, gdy centrum aktywne oddziałuje z ligandem (ryc. 1.15).
Heksokinaza- enzym katalizujący fosforylację glukozy przy użyciu ATP. Miejsce aktywne enzymu znajduje się w szczelinie pomiędzy dwiema domenami. Kiedy heksokinaza wiąże się z glukozą, otaczające ją domeny zamykają się, a substrat zostaje uwięziony, gdzie następuje fosforylacja (patrz ryc. 1.15).
 Ryż. 1,15. Wiązanie domen heksokinazy z glukozą
Ryż. 1,15. Wiązanie domen heksokinazy z glukozą
W niektórych białkach domeny pełnią niezależne funkcje, wiążąc się z różnymi ligandami. Takie białka nazywane są wielofunkcyjnymi.
3. Leki to ligandy wpływające na funkcję białek. Oddziaływanie białek z ligandami jest specyficzne. Jednakże, ze względu na labilność konformacyjną białka i jego centrum aktywnego, możliwe jest wybranie innej substancji, która również mogłaby oddziaływać z białkiem w centrum aktywnym lub innej części cząsteczki.
Nazywa się substancję o strukturze podobnej do naturalnego ligandu Strukturalny analog ligandu lub nienaturalny ligand. Oddziałuje również z białkiem w miejscu aktywnym. Strukturalny analog liganda może zarówno wzmacniać funkcję białka (agonista), i zmniejsz go (antagonista). Ligand i jego analogi strukturalne konkurują ze sobą o wiązanie z białkiem w tym samym miejscu. Takie substancje nazywane są konkurencyjne modulatory(regulatory) funkcji białek. Wiele leków działa jako inhibitory białek. Część z nich otrzymywana jest poprzez chemiczną modyfikację naturalnych ligandów. Inhibitorami funkcji białek mogą być leki i trucizny.
Atropina jest konkurencyjnym inhibitorem receptorów M-cholinergicznych. Acetylocholina jest neuroprzekaźnikiem odpowiedzialnym za przekazywanie impulsów nerwowych przez synapsy cholinergiczne. Aby przeprowadzić wzbudzenie, uwolniono w szczelina synaptyczna acetylocholina musi oddziaływać z białkiem receptorowym błony postsynaptycznej. Znaleziono dwa typy receptory cholinergiczne:
Receptor M oprócz acetylocholiny oddziałuje selektywnie z muskaryną (toksyną muchomora). M - receptory cholinergiczne występują na mięśniach gładkich i podczas interakcji z acetylocholiną powodują ich skurcz;
Receptor H specyficznie wiążący się z nikotyną. Receptory N-cholinergiczne znajdują się w synapsach prążkowanych mięśni szkieletowych.
Specyficzny inhibitor Receptory M-cholinergiczne jest atropina. Występuje w roślinach belladonna i lulek.
 Atropina posiada grupy funkcyjne podobne budową do acetylocholiny i ich rozmieszczeniem przestrzennym, dlatego jest konkurencyjnym inhibitorem receptorów M-cholinergicznych. Biorąc pod uwagę, że wiązanie acetylocholiny z receptorami M-cholinergicznymi powoduje skurcz mięśni gładkich, atropina jest stosowana jako lek łagodzący ich skurcze (przeciwskurczowy). Dlatego wiadomo, że atropina służy do rozluźniania mięśni oka podczas oglądania dna oka, a także do łagodzenia skurczów podczas kolki żołądkowo-jelitowej. Receptory M-cholinergiczne są również obecne w ośrodku system nerwowy(OUN), dlatego duże dawki atropiny mogą powodować niepożądane reakcje ze strony ośrodkowego układu nerwowego: pobudzenie ruchowe i psychiczne, omamy, drgawki.
Atropina posiada grupy funkcyjne podobne budową do acetylocholiny i ich rozmieszczeniem przestrzennym, dlatego jest konkurencyjnym inhibitorem receptorów M-cholinergicznych. Biorąc pod uwagę, że wiązanie acetylocholiny z receptorami M-cholinergicznymi powoduje skurcz mięśni gładkich, atropina jest stosowana jako lek łagodzący ich skurcze (przeciwskurczowy). Dlatego wiadomo, że atropina służy do rozluźniania mięśni oka podczas oglądania dna oka, a także do łagodzenia skurczów podczas kolki żołądkowo-jelitowej. Receptory M-cholinergiczne są również obecne w ośrodku system nerwowy(OUN), dlatego duże dawki atropiny mogą powodować niepożądane reakcje ze strony ośrodkowego układu nerwowego: pobudzenie ruchowe i psychiczne, omamy, drgawki.
Ditilin jest konkurencyjnym agonistą receptorów H-cholinergicznych, hamującym funkcję synaps nerwowo-mięśniowych.
 Synapsy nerwowo-mięśniowe mięśni szkieletowych zawierają receptory H-cholinergiczne. Ich interakcja z acetylocholiną prowadzi do skurczów mięśni. Podczas niektórych zabiegów chirurgicznych, a także w badaniach endoskopowych stosuje się leki powodujące rozluźnienie mięśni szkieletowych (leki zwiotczające mięśnie). Należą do nich ditilina, która jest strukturalnym analogiem acetylocholiny. Przyłącza się do receptorów H-cholinergicznych, ale w przeciwieństwie do acetylocholiny jest bardzo powoli niszczona przez enzym acetylocholinoesterazę. W wyniku długotrwałego otwarcia kanałów jonowych i utrzymującej się depolaryzacji błony dochodzi do zaburzenia przewodzenia impulsów nerwowych i rozluźnienia mięśni. Początkowo właściwości te odkryto w truciźnie kurary, dlatego nazywane są takimi lekami podobny do kurary.
Synapsy nerwowo-mięśniowe mięśni szkieletowych zawierają receptory H-cholinergiczne. Ich interakcja z acetylocholiną prowadzi do skurczów mięśni. Podczas niektórych zabiegów chirurgicznych, a także w badaniach endoskopowych stosuje się leki powodujące rozluźnienie mięśni szkieletowych (leki zwiotczające mięśnie). Należą do nich ditilina, która jest strukturalnym analogiem acetylocholiny. Przyłącza się do receptorów H-cholinergicznych, ale w przeciwieństwie do acetylocholiny jest bardzo powoli niszczona przez enzym acetylocholinoesterazę. W wyniku długotrwałego otwarcia kanałów jonowych i utrzymującej się depolaryzacji błony dochodzi do zaburzenia przewodzenia impulsów nerwowych i rozluźnienia mięśni. Początkowo właściwości te odkryto w truciźnie kurary, dlatego nazywane są takimi lekami podobny do kurary.
TEMAT 1.3. DENATURACJA BIAŁEK I MOŻLIWOŚĆ ICH SPONTANICZNEJ RENATURACJI
1. Ponieważ natywna konformacja białek jest utrzymywana na skutek słabych oddziaływań, zmiany składu i właściwości środowiska otaczającego białko, narażenie na odczynniki chemiczne i czynniki fizyczne powodują zmianę ich konformacji (właściwość labilności konformacyjnej). Luka duża ilość wiązania prowadzą do zniszczenia natywnej konformacji i denaturacji białek.
Denaturacja białek- jest to zniszczenie ich natywnej konformacji pod wpływem czynników denaturujących, spowodowane zerwaniem słabych wiązań stabilizujących strukturę przestrzenną białka. Denaturacji towarzyszy zniszczenie unikalnej trójwymiarowej struktury i centrum aktywnego białka oraz utrata jego aktywności biologicznej (ryc. 1.16).
Wszystkie zdenaturowane cząsteczki jednego białka uzyskują losową konformację, która różni się od innych cząsteczek tego samego białka. Rodniki aminokwasowe tworzące centrum aktywne okazują się być od siebie przestrzennie odległe, tj. specyficzne miejsce wiązania białka z ligandem ulega zniszczeniu. Podczas denaturacji pierwotna struktura białek pozostaje niezmieniona.
Zastosowanie środków denaturujących w badaniach biologicznych i medycynie. W badania biochemiczne Przed oznaczeniem w materiale biologicznym związków o niskiej masie cząsteczkowej zazwyczaj najpierw usuwa się z roztworu białka. Najczęściej wykorzystuje się w tym celu kwas trichlorooctowy (TCA). Po dodaniu TCA do roztworu zdenaturowane białka wytrącają się i można je łatwo usunąć przez filtrację (tab. 1.1.).
W medycynie środki denaturujące są często stosowane do sterylizacji narzędzi i materiałów medycznych w autoklawach (środkiem denaturującym jest wysoka temperatura) oraz jako środki antyseptyczne (alkohol, fenol, chloramina) do leczenia zanieczyszczonych powierzchni zawierających patogenną mikroflorę.
2. Spontaniczna reaktywacja białek- dowód determinizmu pierwotnej struktury, konformacji i funkcji białek. Poszczególne białka są produktami jednego genu, które mają identyczną sekwencję aminokwasów i uzyskują w komórce tę samą konformację. Zasadniczy wniosek, że pierwotna struktura białka zawiera już informację o jego konformacji i funkcji, został wysunięty na podstawie zdolności niektórych białek (w szczególności rybonukleazy i mioglobiny) do samoistnej renatywacji – przywracania ich natywnej konformacji po denaturacji.
Tworzenie przestrzennych struktur białkowych odbywa się metodą samoorganizacji – spontanicznego procesu, w którym łańcuch polipeptydowy, posiadający unikalną strukturę pierwotną, ma tendencję do przyjmowania konformacji o najniższej energii swobodnej w roztworze. Zdolność do renatywacji białek, które po denaturacji zachowują swoją pierwotną strukturę, opisano w eksperymencie z enzymem rybonukleazą.
Rybonukleaza jest enzymem rozkładającym wiązania pomiędzy poszczególnymi nukleotydami w cząsteczce RNA. To globularne białko ma jeden łańcuch polipeptydowy, którego trzeciorzędowa struktura jest stabilizowana przez wiele słabych i cztery wiązania dwusiarczkowe.
Traktowanie rybonukleazy mocznikiem, który rozrywa wiązania wodorowe w cząsteczce, oraz czynnikiem redukującym, który rozrywa wiązania disiarczkowe, prowadzi do denaturacji enzymu i utraty jego aktywności.
Usunięcie czynników denaturujących poprzez dializę prowadzi do przywrócenia konformacji i funkcji białka, tj. do odrodzenia. (ryc. 1.17).
 Ryż. 1.17. Denaturacja i renatywacja rybonukleazy
Ryż. 1.17. Denaturacja i renatywacja rybonukleazy
A - natywna konformacja rybonukleazy, w której trzeciorzędowej strukturze znajdują się cztery wiązania dwusiarczkowe; B - zdenaturowana cząsteczka rybonukleazy;
B - reaktywowana cząsteczka rybonukleazy z przywróconą strukturą i funkcją
1. Wypełnij tabelę 1.2.
Tabela 1.2. Klasyfikacja aminokwasów ze względu na polarność rodników
2. Zapisz wzór tetrapeptydu:
Asp – Pro – Fen – Liz
a) podkreślić powtarzające się grupy w peptydzie, które tworzą szkielet peptydu i grupy zmienne reprezentowane przez rodniki aminokwasowe;
b) oznaczyć końce N i C;
c) podkreślić wiązania peptydowe;
d) napisz kolejny peptyd składający się z tych samych aminokwasów;
e) policzyć liczbę możliwych wariantów tetrapeptydu o podobnym składzie aminokwasów.
3. Wyjaśnij rolę pierwszorzędowej struktury białek na przykładzie analizy porównawczej dwóch strukturalnie podobnych i bliskich ewolucyjnie hormonów peptydowych neuroprzysadki ssaków – oksytocyny i wazopresyny (tab. 1.3).
Tabela 1.3. Struktura i funkcje oksytocyny i wazopresyny
 Dla tego:
Dla tego:
a) porównać skład i sekwencję aminokwasów dwóch peptydów;
b) znaleźć podobieństwo struktury pierwszorzędowej obu peptydów i podobieństwo ich działania biologicznego;
c) znaleźć różnice w budowie dwóch peptydów i różnice w ich funkcjach;
d) wyciągnąć wniosek na temat wpływu struktury pierwszorzędowej peptydów na ich funkcje.
4. Opisać główne etapy powstawania konformacji białek globularnych (struktury drugorzędowe, trzeciorzędowe, koncepcja struktury superwtórnej). Wskaż rodzaje wiązań biorących udział w tworzeniu struktur białkowych. Które rodniki aminokwasowe mogą brać udział w tworzeniu oddziaływań hydrofobowych, jonowych, wiązań wodorowych.
Daj przykłady.
5. Zdefiniować pojęcie „labilności konformacyjnej białek”, wskazać przyczyny jego istnienia i znaczenie.
6. Rozwiń znaczenie wyrażenia: „Funkcjonowanie białek opiera się na ich specyficznej interakcji z ligandem”, używając terminów i wyjaśniając ich znaczenie: konformacja białka, centrum aktywne, ligand, komplementarność, funkcja białka.
7. Na jednym przykładzie wyjaśnij, czym są domeny i jaka jest ich rola w funkcjonowaniu białek.
ZADANIA SAMOKONTROLI
1. Mecz.
Grupa funkcyjna rodnika aminokwasowego:
A. Grupa karboksylowa B. Grupa hydroksylowa C Grupa guanidynowa D. Grupa tiolowa E. Grupa aminowa
2. Wybierz prawidłowe odpowiedzi.
Aminokwasy z polarnymi nienaładowanymi rodnikami to:
A. Cis B. Asn
B. Glu G. Trzy
3. Wybierz prawidłowe odpowiedzi.
Rodniki aminokwasowe:
A. Podaj specyfikę struktury pierwotnej. B. Weź udział w tworzeniu struktury trzeciorzędowej
B. Znajdujące się na powierzchni białka wpływają na jego rozpuszczalność. D. Tworzą centrum aktywne
D. Uczestniczyć w tworzeniu wiązań peptydowych
4. Wybierz prawidłowe odpowiedzi.
Oddziaływania hydrofobowe mogą tworzyć się pomiędzy rodnikami aminokwasów:
A. Tre Lay B. Pro Three
B. Met Ile G. Tir Ala D. Val Fen
5. Wybierz prawidłowe odpowiedzi.
Wiązania jonowe mogą tworzyć się pomiędzy rodnikami aminokwasów:
A. Gln Asp B. Apr Liz
B. Liz Glu G. Gis Asp D. Asn Apr
6. Wybierz prawidłowe odpowiedzi.
Wiązania wodorowe mogą tworzyć się pomiędzy rodnikami aminokwasów:
A. Ser Gln B. Cis Tre
B. Asp Liz G. Glu Asp D. Asn Tre
7. Mecz.
Rodzaj wiązania biorącego udział w tworzeniu struktury białka:
A. Struktura pierwotna B. Struktura wtórna
B. Struktura trzeciorzędowa
D. Struktura nadwtórna. E. Konformacja.
1. Wiązania wodorowe pomiędzy atomami szkieletu peptydowego
2. Słabe wiązania pomiędzy grupami funkcyjnymi rodników aminokwasów
3. Wiązania pomiędzy grupami α-aminowymi i α-karboksylowymi aminokwasów
8. Wybierz prawidłowe odpowiedzi. Trypsyna:
A. Enzym proteolityczny B. Zawiera dwie domeny
B. Hydrolizuje skrobię
D. Aktywna witryna znajduje się pomiędzy domenami. D. Składa się z dwóch łańcuchów polipeptydowych.
9. Wybierz prawidłowe odpowiedzi. Atropina:
A. Neuroprzekaźnik
B. Strukturalny analog acetylocholiny
B. Oddziałuje z receptorami H-cholinergicznymi
D. Wzmacnia przewodzenie impulsów nerwowych przez synapsy cholinergiczne
D. Konkurencyjny inhibitor receptorów M-cholinergicznych
10. Wybierz poprawne stwierdzenia. W białkach:
A. Struktura pierwotna zawiera informacje o strukturze jej miejsca aktywnego
B. Centrum aktywne powstaje na poziomie struktury pierwotnej
B. Konformacja jest sztywno utwierdzona wiązaniami kowalencyjnymi
D. Miejsce aktywne może oddziaływać z grupą podobnych ligandów
ze względu na labilność konformacyjną białek D. Zmiana środowisko, może wpływać na powinowactwo substancji czynnej
centrum do liganda
1. 1-B, 2-G, 3-B.
3. A, B, C, D.
7. 1-B, 2-D, 3-A.
8. A, B, C, D.
PODSTAWOWE TERMINY I POJĘCIA
1. Białko, polipeptyd, aminokwasy
2. Pierwotne, drugorzędowe i trzeciorzędowe struktury białkowe
3. Konformacja, konformacja białka natywnego
4. Wiązania kowalencyjne i słabe w białkach
5. Labilność konformacyjna
6. Miejsce aktywne białka
7. Ligandy
8. Zwijanie białek
9. Strukturalne analogi ligandów
10. Białka domenowe
11. Białka proste i złożone
12. Denaturacja białek, środki denaturujące
13. Reaktywacja białek
Rozwiązywać problemy
„Strukturalna organizacja białek i podstawy ich funkcjonowania”
1. Główną funkcją białka – hemoglobiny A (HbA) jest transport tlenu do tkanek. W populacji ludzkiej znanych jest wiele form tego białka o zmienionych właściwościach i funkcjach – tzw. nieprawidłowe hemoglobiny. Na przykład stwierdzono, że hemoglobina S, występująca w czerwonych krwinkach pacjentów z niedokrwistością sierpowatokrwinkową (HbS), ma słabą rozpuszczalność w warunkach niskiego ciśnienia parcjalnego tlenu (jak ma to miejsce w przypadku krwi żylnej). Prowadzi to do powstania agregatów tego białka. Białko traci swoją funkcję, wytrąca się i nabywają czerwone krwinki nieregularny kształt(niektóre z nich przybierają kształt sierpa) i ulegają szybszemu niż zwykle zniszczeniu w śledzionie. W rezultacie rozwija się anemia sierpowatokrwinkowa.
Jedyną różnicę w pierwszorzędowej strukturze HbA stwierdzono w N-końcowym regionie łańcucha β hemoglobiny. Porównaj regiony N-końcowe nici β i pokaż, jak zmiany w strukturze pierwszorzędowej białka wpływają na jego właściwości i funkcje.
 Dla tego:
Dla tego:
a) napisz wzory aminokwasów, którymi różni się HbA i porównaj właściwości tych aminokwasów (polarność, ładunek).
b) wyciągnąć wniosek co do przyczyny zmniejszenia rozpuszczalności i zakłócenia transportu tlenu do tkanek.
2. Rysunek przedstawia schemat budowy białka, które posiada centrum wiążące z ligandem (centrum aktywne). Wyjaśnij, dlaczego białko dokonuje selektywnego wyboru ligandu. Dla tego:
a) przypomnij sobie, czym jest centrum aktywne białka i rozważ budowę centrum aktywnego białka pokazanego na rysunku;
b) napisz wzory rodników aminokwasowych tworzących centrum aktywne;
c) narysuj ligand, który mógłby specyficznie oddziaływać z miejscem aktywnym białka. Wskaż na nim grupy funkcyjne, które mogą tworzyć wiązania z rodnikami aminokwasów tworzącymi centrum aktywne;
d) wskazać rodzaje wiązań, które powstają pomiędzy ligandem i rodnikami aminokwasowymi centrum aktywnego;
e) wyjaśnić, na czym polega specyfika oddziaływania białko-ligand.
 3.
Rysunek przedstawia miejsce aktywne białka i kilka ligandów.
3.
Rysunek przedstawia miejsce aktywne białka i kilka ligandów.
Określ, który ligand najprawdopodobniej oddziałuje z miejscem aktywnym białka i dlaczego.
 Jakie rodzaje wiązań powstają podczas tworzenia kompleksu białko-ligand?
Jakie rodzaje wiązań powstają podczas tworzenia kompleksu białko-ligand?
4. Strukturalne analogi naturalnych ligandów białkowych można stosować jako leki modyfikujące aktywność białek.
Acetylocholina jest mediatorem przekazywania pobudzenia w synapsach nerwowo-mięśniowych. Kiedy acetylocholina wchodzi w interakcję z białkami - receptorami błony postsynaptycznej mięśni szkieletowych, otwierają się kanały jonowe i następuje skurcz mięśni. Ditilin to lek stosowany w niektórych operacjach w celu rozluźnienia mięśni, ponieważ zakłóca przekazywanie impulsów nerwowych przez synapsy nerwowo-mięśniowe. Wyjaśnij mechanizm działania ditiliny jako środka zwiotczającego mięśnie. Dla tego:
a) napisz wzory acetylocholiny i dityliny i porównaj ich budowę;
b) opisać mechanizm relaksującego działania ditiliny.
5. W niektórych chorobach wzrasta temperatura ciała pacjenta, co uważa się za reakcję obronną organizmu. Jednak wysokie temperatury są szkodliwe dla białek organizmu. Wyjaśnij, dlaczego w temperaturach powyżej 40°C funkcja białek zostaje zakłócona i powstaje zagrożenie dla życia człowieka. Aby to zrobić, pamiętaj:
1) Struktura białek i wiązania utrzymujące ich strukturę w natywnej konformacji;
2) Jak zmienia się struktura i funkcja białek wraz ze wzrostem temperatury?;
3) Czym jest homeostaza i dlaczego jest ważna dla utrzymania zdrowia człowieka.
Jednostka modułowa 2 BIAŁKA OLIGOMERYCZNE JAKO CELE DZIAŁAŃ REGULACYJNYCH. STRUKTURALNA I FUNKCJONALNA RÓŻNORODNOŚĆ BIAŁEK. METODY SEPARACJI I OCZYSZCZANIA BIAŁEK
Cele nauczania Być w stanie:
1. Wykorzystywać wiedzę o cechach budowy i funkcjach białek oligomerycznych do zrozumienia adaptacyjnych mechanizmów regulacji ich funkcji.
2. Wyjaśniać rolę białek opiekuńczych w syntezie i utrzymaniu konformacji białek w warunkach komórkowych.
3. Wyjaśniać różnorodność przejawów życia poprzez różnorodność struktur i funkcji białek syntetyzowanych w organizmie.
4. Analizować związek pomiędzy strukturą białek a ich funkcją na przykładach porównania pokrewnych hemoprotein – mioglobiny i hemoglobiny, a także przedstawicieli pięciu klas białek z rodziny immunoglobulin.
5. Wykorzystać wiedzę o osobliwościach właściwości fizycznych i chemicznych białek do doboru metod ich oczyszczania z innych białek i zanieczyszczeń.
6. Interpretować wyniki badań ilościowych i jakość składu białek osocza krwi w celu potwierdzenia lub wyjaśnienia diagnozy klinicznej.
Wiedzieć:
1. Cechy budowy białek oligomerycznych i mechanizmy adaptacyjne regulujące ich funkcje na przykładzie hemoglobiny.
2. Struktura i funkcje białek opiekuńczych oraz ich znaczenie dla utrzymania natywnej konformacji białek w warunkach komórkowych.
3. Zasady łączenia białek w rodziny na podstawie podobieństwa ich budowy i funkcji na przykładzie immunoglobulin.
4. Metody rozdziału białek w oparciu o charakterystykę ich właściwości fizykochemicznych.
5. Elektroforeza osocza krwi jako metoda oceny jakościowego i ilościowego składu białek.
TEMAT 1.4. CECHY STRUKTURY I FUNKCJONOWANIA BIAŁEK OLIGOMERYCZNYCH NA PRZYKŁADZIE HEMOGLOBINY
1. Wiele białek zawiera kilka łańcuchów polipeptydowych. Takie białka nazywane są oligomeryczny, i poszczególne łańcuchy - protomery. Protomery w białkach oligomerycznych są połączone wieloma słabymi wiązaniami niekowalencyjnymi (hydrofobowymi, jonowymi, wodorowymi). Interakcja
protomery realizowane są dzięki komplementarność ich powierzchnie stykowe.
Liczba protomerów w białkach oligomerycznych może się znacznie różnić: hemoglobina zawiera 4 protomery, enzym aminotransferaza asparaginianowa ma 12 protomerów, a białko wirusa mozaiki tytoniu zawiera 2120 protomerów połączonych wiązaniami niekowalencyjnymi. W konsekwencji białka oligomeryczne mogą mieć bardzo wysokie masy cząsteczkowe.
Można rozważyć interakcję jednego protomera z innymi szczególny przypadek interakcje białko-ligand, ponieważ każdy protomer służy jako ligand dla innych protomerów. Nazywa się liczbę i sposób łączenia protomerów w białku czwartorzędowa struktura białka.
Białka mogą zawierać protomery o tej samej lub różnej strukturze, na przykład homodimery to białka zawierające dwa identyczne protomery, a heterodimery to białka zawierające dwa różne protomery.
Jeśli białka zawierają różne protomery, wówczas mogą powstać na nich centra wiążące z różnymi ligandami różniącymi się budową. Kiedy ligand wiąże się z miejscem aktywnym, manifestuje się funkcja tego białka. Centrum znajdujące się na innym protomerze nazywa się allosterycznym (innym niż centrum aktywne). Kontaktowanie się allosteryczny ligand lub efektor, pełni funkcję regulacyjną (ryc. 1.18). Oddziaływanie centrum allosterycznego z efektorem powoduje zmiany konformacyjne w strukturze całego białka oligomerycznego ze względu na jego labilność konformacyjną. Wpływa to na powinowactwo miejsca aktywnego do określonego ligandu i reguluje funkcję tego białka. Zmiana konformacji i funkcji wszystkich protomerów podczas interakcji białka oligomerycznego z co najmniej jednym ligandem nazywana jest kooperacyjnymi zmianami konformacyjnymi. Nazywa się efektory wzmacniające funkcję białka aktywatory, i efektory, które hamują jego funkcję - inhibitory.
Zatem białka oligomeryczne, a także białka o strukturze domenowej, mają nową właściwość w porównaniu do białek monomerycznych - zdolność do allosterycznej regulacji funkcji (regulacja poprzez przyłączenie różnych ligandów do białka). Można to zobaczyć porównując struktury i funkcje dwóch blisko spokrewnionych białek złożonych, mioglobiny i hemoglobiny.
 Ryż. 1.18. Schemat struktury białka dimerycznego
Ryż. 1.18. Schemat struktury białka dimerycznego
2. Tworzenie struktur przestrzennych i funkcjonowanie mioglobiny.
Mioglobina (Mb) to białko występujące w mięśniach czerwonych, którego główną funkcją jest tworzenie rezerw O 2 niezbędnych do intensywnej pracy mięśni. Mb jest białkiem złożonym, zawierającym część białkową – apoMb i część niebiałkową – hem. Pierwotna struktura apoMB determinuje jego zwartą konformację kulistą i strukturę centrum aktywnego, do którego przyłączona jest niebiałkowa część mioglobiny – hem. Tlen docierający z krwi do mięśni wiąże się z hemami Fe+ 2 w mioglobinie. Mb jest białkiem monomerycznym, które ma bardzo duże powinowactwo do O 2, dlatego uwalnianie tlenu przez mioglobinę następuje tylko podczas intensywnej pracy mięśni, kiedy ciśnienie parcjalne O 2 gwałtownie maleje.
Tworzenie konformacji Mv. W mięśniach czerwonych na rybosomach podczas translacji syntetyzowana jest pierwotna struktura MB, reprezentowana przez specyficzną sekwencję 153 reszt aminokwasowych. Wtórna struktura Mb zawiera osiem α-helis, zwanych łacińskimi literami od A do H, pomiędzy którymi znajdują się obszary niehelikalne. Trzeciorzędowa struktura Mb ma postać zwartej globuli, w której wnęce znajduje się centrum aktywne pomiędzy α-helisami F i E (ryc. 1.19).
 Ryż. 1.19. Struktura mioglobiny
Ryż. 1.19. Struktura mioglobiny
3. Cechy budowy i funkcjonowania ośrodka czynnego SN. Centrum aktywne Mb utworzone jest głównie przez hydrofobowe rodniki aminokwasowe, szeroko od siebie oddalone w strukturze pierwszorzędowej (na przykład Tri 3 9 i Fen 138) Słabo rozpuszczalne w wodzie ligandy – hem i O 2 – przyłączają się do centrum aktywnego. Hem jest specyficznym ligandem apoMB (ryc. 1.20), którego podstawę stanowią cztery pierścienie pirolowe połączone mostkami metenylowymi; w centrum znajduje się atom Fe+ 2 połączony z atomami azotu pierścieni pirolu czterema wiązaniami koordynacyjnymi. W centrum aktywnym Mb oprócz hydrofobowych rodników aminokwasowych znajdują się także reszty dwóch aminokwasów z rodnikami hydrofilowymi - GisE 7(Rdz 64) i GIS F8(Jego 93) (ryc. 1.21).
 Ryż. 1,20. Struktura hemu - niebiałkowej części mioglobiny i hemoglobiny
Ryż. 1,20. Struktura hemu - niebiałkowej części mioglobiny i hemoglobiny
 Ryż. 1.21. Lokalizacja hemu i O2 w miejscu aktywnym protomerów apomioglobiny i hemoglobiny
Ryż. 1.21. Lokalizacja hemu i O2 w miejscu aktywnym protomerów apomioglobiny i hemoglobiny
Hem jest kowalencyjnie związany z His F8 poprzez atom żelaza. O 2 przyłącza się do żelaza po drugiej stronie płaszczyzny hemu. Jego E 7 jest niezbędny do prawidłowej orientacji O 2 i ułatwia dodanie tlenu do hemu Fe + 2
GIS F8 tworzy wiązanie koordynacyjne z Fe+ 2 i mocno wiąże hem w centrum aktywnym. GisE 7 niezbędny do prawidłowej orientacji w centrum aktywnym innego liganda - O 2 podczas jego interakcji z hemem Fe + 2. Mikrośrodowisko hemu stwarza warunki do silnego, ale odwracalnego wiązania O 2 z Fe + 2 i zapobiega przedostawaniu się wody do hydrofobowego miejsca aktywnego, co może prowadzić do jego utlenienia do Fe + 3.
Monomeryczna struktura Mb i jego centrum aktywnego decyduje o wysokim powinowactwie białka do O2.
4. Struktura oligomeryczna Hb i regulacja powinowactwa Hb do ligandów O 2. Hemoglobiny ludzkie- rodzina białek, takich jak mioglobina, spokrewniona z białkami złożonymi (hemoproteinami). Mają budowę tetrameryczną i zawierają dwa łańcuchy α, ale różnią się budową dwóch pozostałych łańcuchów polipeptydowych (łańcuchy 2α, 2x). Struktura drugiego łańcucha polipeptydowego determinuje cechy funkcjonowania tych form Hb. Około 98% hemoglobiny w czerwonych krwinkach dorosłego człowieka to hemoglobina hemoglobina A(łańcuchy 2α, 2p).
Podczas rozwoju płodu działają dwa główne typy hemoglobin: embrionalna Hb(2α, 2ε), który znajduje się na wczesne stadia rozwój płodu i hemoglobina F (płód)- (2α, 2γ), która zastępuje wczesną hemoglobinę płodową w szóstym miesiącu rozwoju wewnątrzmacicznego i dopiero po urodzeniu zastępuje ją Hb A.
HB A to białko spokrewnione z mioglobiną (MB) występujące w czerwonych krwinkach dorosłego człowieka. Struktura jej poszczególnych protomerów jest podobna do struktury mioglobiny. Struktury drugorzędowe i trzeciorzędowe protomerów mioglobiny i hemoglobiny są bardzo podobne, mimo że w strukturze pierwszorzędowej ich łańcuchów polipeptydowych identyczne są jedynie 24 reszty aminokwasowe (drugorzędowa struktura protomerów hemoglobiny, podobnie jak mioglobiny, zawiera osiem α-helis, oznaczone literami łacińskimi od A do H, a struktura trzeciorzędowa ma postać zwartej globuli). Jednak w przeciwieństwie do mioglobiny hemoglobina ma strukturę oligomeryczną, składającą się z czterech łańcuchów polipeptydowych połączonych wiązaniami niekowalencyjnymi (ryc. 1.22).
Każdy protomer Hb jest powiązany z częścią niebiałkową – hemem i sąsiednimi protomerami. Połączenie części białkowej Hb z hemem jest podobne do połączenia mioglobiny: w centrum aktywnym białka hydrofobowe części hemu są otoczone hydrofobowymi rodnikami aminokwasowymi, z wyjątkiem His F 8 i His E 7, które znajdują się po obu stronach płaszczyzny hemu i odgrywają podobną rolę w funkcjonowaniu białka i jego wiązaniu z tlenem (patrz budowa mioglobiny).
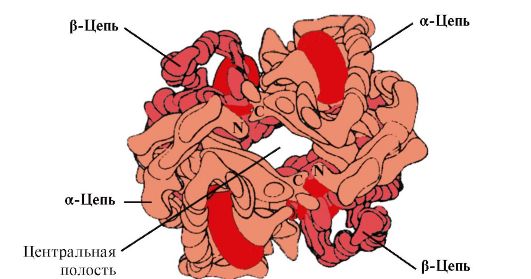 Ryż. 1,22. Oligomeryczna struktura hemoglobiny
Ryż. 1,22. Oligomeryczna struktura hemoglobiny
Oprócz, GisE 7 pełni ważne dodatkowa rola w funkcjonowaniu Nv. Wolny hem ma 25 000 razy większe powinowactwo do CO niż do O2. CO powstaje w organizmie w małych ilościach i ze względu na jego duże powinowactwo do hemu może zakłócać transport O 2 niezbędny do życia komórek. Jednak w składzie hemoglobiny powinowactwo hemu do tlenku węgla przekracza powinowactwo do O 2 tylko 200 razy ze względu na obecność His E 7 w centrum aktywnym. Pozostała część tego aminokwasu stwarza optymalne warunki wiązania hemu z O 2 i osłabia oddziaływanie hemu z CO.
5. Główną funkcją HB jest transport O2 z płuc do tkanek. W przeciwieństwie do monomerycznej mioglobiny, która ma bardzo duże powinowactwo do O2 i pełni funkcję magazynowania tlenu w czerwonych mięśniach, oligomeryczna struktura hemoglobiny zapewnia:
1) szybkie nasycenie HB tlenem w płucach;
2) zdolność HB do uwalniania tlenu w tkankach przy stosunkowo wysokim ciśnieniu cząstkowym O 2 (20–40 mm Hg);
3) możliwość regulacji powinowactwa Hb do O 2.
6. Wspólne zmiany w konformacji protomerów hemoglobiny przyspieszają wiązanie O 2 w płucach i jego uwalnianie do tkanek. W płucach wysokie ciśnienie parcjalne O2 sprzyja jego wiązaniu z Hb w miejscu aktywnym czterech protomerów (2α i 2β). Aktywne centrum każdego protomeru, podobnie jak w mioglobinie, znajduje się pomiędzy dwiema α-helisami (F i E) w kieszeni hydrofobowej. Zawiera część niebiałkową - hem, połączoną z częścią białkową wieloma słabymi oddziaływaniami hydrofobowymi i jednym silnym wiązaniem pomiędzy hemem Fe 2 + i His F 8 (patrz ryc. 1.21).
W deoksyhemoglobinie, dzięki temu wiązaniu z His F 8, atom Fe 2 + wystaje z płaszczyzny hemu w kierunku histydyny. Wiązanie O 2 z Fe 2 + zachodzi po drugiej stronie hemu w regionie His E 7 przy użyciu pojedynczego wolnego wiązania koordynacyjnego. Jego E 7 zapewnia optymalne warunki wiązania O 2 z żelazem hemowym.
Dodatek O2 do atomu Fe+2 jednego protomeru powoduje jego ruch do płaszczyzny hemu, po czym następuje związana z nim reszta histydyny
 Ryż. 1,23. Zmiana konformacji protomeru hemoglobiny w połączeniu z O2
Ryż. 1,23. Zmiana konformacji protomeru hemoglobiny w połączeniu z O2
Prowadzi to do zmiany konformacji wszystkich łańcuchów polipeptydowych ze względu na ich labilność konformacyjną. Zmiana konformacji pozostałych łańcuchów ułatwia ich interakcję z kolejnymi cząsteczkami O 2 .
Czwarta cząsteczka O 2 przyłącza się do hemoglobiny 300 razy łatwiej niż pierwsza (ryc. 1.24).
 Ryż. 1,24. Spółdzielcze zmiany w konformacji protomerów hemoglobiny podczas jej interakcji z O2
Ryż. 1,24. Spółdzielcze zmiany w konformacji protomerów hemoglobiny podczas jej interakcji z O2
W tkankach każda kolejna cząsteczka O 2 jest odcinana łatwiej niż poprzednia, także na skutek kooperatywnych zmian w konformacji protomerów.
7. CO 2 i H+ powstające podczas katabolizmu materia organiczna, zmniejszają powinowactwo hemoglobiny do O 2 proporcjonalnie do ich stężenia. Energia niezbędna do funkcjonowania komórek wytwarzana jest przede wszystkim w mitochondriach podczas utleniania substancji organicznych przy użyciu O 2 dostarczanego z płuc przez hemoglobinę. W wyniku utleniania substancji organicznych powstają końcowe produkty ich rozkładu: CO 2 i K 2 O, których ilość jest proporcjonalna do intensywności zachodzących procesów utleniania.
CO 2 dyfunduje z komórek do krwi i przenika do czerwonych krwinek, gdzie pod wpływem enzymu karbanhydrazy przekształca się w kwas węglowy. Ten słaby kwas dysocjuje na proton i jon wodorowęglanowy.
H+ są w stanie przyłączyć się do Jego rodników 14 6 w łańcuchach α i β hemoglobiny, tj. w obszarach odległych od hemu. Protonowanie hemoglobiny zmniejsza jej powinowactwo do O 2, sprzyja usuwaniu O 2 z oksyHb, tworzeniu deoksyHb i zwiększa dopływ tlenu do tkanek proporcjonalnie do liczby utworzonych protonów (ryc. 1.25).
Wzrost ilości uwalnianego tlenu w zależności od wzrostu stężenia H+ w czerwonych krwinkach nazywany jest efektem Bohra (nazwanym na cześć duńskiego fizjologa Christiana Bohra, który jako pierwszy odkrył ten efekt).
W płucach wysokie ciśnienie parcjalne tlenu sprzyja jego wiązaniu z deoksyHb, co zmniejsza powinowactwo białka do H +. Uwolnione protony pod działaniem kwasu węglowego reagują z wodorowęglanami, tworząc CO 2 i H 2 O

 Ryż. 1,25. Zależność powinowactwa Hb do O 2 od stężenia CO 2 i protonów (efekt Bohra):
Ryż. 1,25. Zależność powinowactwa Hb do O 2 od stężenia CO 2 i protonów (efekt Bohra):
A- wpływ stężenia CO 2 i H+ na uwalnianie O 2 z kompleksu z HB (efekt Bohra); B- natlenienie deoksyhemoglobiny w płucach, tworzenie i uwalnianie CO2.
Powstały CO 2 przedostaje się do przestrzeni pęcherzykowej i jest usuwany wraz z wydychanym powietrzem. Zatem ilość tlenu uwalnianego przez hemoglobinę w tkankach jest regulowana przez produkty katabolizmu substancji organicznych: im intensywniejszy rozkład substancji, np. podczas wysiłku fizycznego, tym wyższe stężenie CO 2 i H + oraz tym więcej tlenu tkanki otrzymują w wyniku zmniejszenia powinowactwa Hb do O2.
8. Allosteryczna regulacja powinowactwa Hb do O2 przez ligand – 2,3-bisfosfoglicerynian. W erytrocytach allosteryczny ligand hemoglobiny, 2,3-bisfosfoglicerynian (2,3-BPG), jest syntetyzowany z produktu utleniania glukozy - 1,3-bisfosfoglicerynianu. W normalnych warunkach stężenie 2,3-BPG jest wysokie i porównywalne ze stężeniem Hb. 2,3-BPG ma silny ładunek ujemny -5.
 Bisfosfoglicerynian w naczyniach włosowatych tkanek, wiążąc się z deoksyhemoglobiną, zwiększa uwalnianie tlenu do tkanek, zmniejszając powinowactwo Hb do O2.
Bisfosfoglicerynian w naczyniach włosowatych tkanek, wiążąc się z deoksyhemoglobiną, zwiększa uwalnianie tlenu do tkanek, zmniejszając powinowactwo Hb do O2.
W środku tetramerycznej cząsteczki hemoglobiny znajduje się wnęka. Tworzą go reszty aminokwasowe wszystkich czterech protomerów (patrz ryc. 1.22). W naczyniach włosowatych tkanek protonowanie Hb (efekt Bohra) prowadzi do zerwania wiązania pomiędzy żelazem hemowym i O2. W cząsteczce
deoksyhemoglobina w porównaniu do oksyhemoglobiny są dodatkowe wiązania jonowe, łącząc protomery, w wyniku czego wymiary jamy centralnej zwiększają się w porównaniu z oksyhemoglobiną. Jama centralna jest miejscem przyłączania 2,3-BPG do hemoglobiny. Ze względu na różnicę w wielkości jamy centralnej, 2,3-BPG może przyłączać się jedynie do deoksyhemoglobiny.
2,3-BPG oddziałuje z hemoglobiną w miejscu odległym od aktywnych centrów białka i do którego należy allosteryczny(regulacyjne) ligandy i centralna wnęka Hb centrum allosteryczne. 2,3-BPG ma silny ładunek ujemny i oddziałuje z pięcioma dodatnio naładowanymi grupami dwóch łańcuchów β Hb: N-końcową grupą α-aminową Val i rodnikami Lys 82 His 143 (ryc. 1.26).
 Ryż. 1,26. BPG w centralnej jamie deoksyhemoglobiny
Ryż. 1,26. BPG w centralnej jamie deoksyhemoglobiny
BPG wiąże się z trzema dodatnio naładowanymi grupami na każdej nici β.
W naczyniach włosowatych powstająca deoksyhemoglobina oddziałuje z 2,3-BPG i pomiędzy dodatnio naładowanymi rodnikami łańcuchów β a ujemnie naładowanymi ligandami tworzą się wiązania jonowe, które zmieniają konformację białka i zmniejszają powinowactwo Hb do O2 . Zmniejszenie powinowactwa Hb do O 2 przyczynia się do bardziej wydajnego uwalniania O 2 do tkanki.
W płucach, przy wysokim ciśnieniu parcjalnym, tlen oddziałuje z Hb, łącząc żelazo hemowe; w tym przypadku zmienia się konformacja białka, zmniejsza się wnęka centralna i 2,3-BPG zostaje wyparty z centrum allosterycznego
Zatem białka oligomeryczne mają nowe właściwości w porównaniu do białek monomerycznych. Przyłączanie ligandów w miejscach
odległe od siebie przestrzennie (allosteryczne), mogą powodować zmiany konformacyjne w całej cząsteczce białka. W wyniku interakcji z ligandami regulatorowymi następuje zmiana konformacji i adaptacja funkcji cząsteczki białka do zmian środowiskowych.
TEMAT 1.5. UTRZYMANIE NAtywnej KONFORMACJI BIAŁEK W WARUNKACH KOMÓRKOWYCH
W komórkach podczas syntezy łańcuchów polipeptydowych, ich transportu przez błony do odpowiednich części komórki, podczas procesu fałdowania (tworzenia konformacji natywnej) i podczas składania białek oligomerycznych, a także podczas ich funkcjonowania, pośrednie W strukturze białka powstają podatne na agregację, niestabilne konformacje. Rodniki hydrofobowe, zwykle ukryte wewnątrz cząsteczki białka w konformacji natywnej, pojawiają się na powierzchni w konformacji niestabilnej i mają tendencję do łączenia się z grupami innych białek, które są słabo rozpuszczalne w wodzie. W komórkach wszystkich znanych organizmów odkryto specjalne białka, które zapewniają optymalne zwijanie białek komórkowych, stabilizują ich natywną konformację podczas funkcjonowania i, co najważniejsze, utrzymują strukturę i funkcje białek wewnątrzkomórkowych, gdy zaburzona jest homeostaza. Białka te nazywane są „opiekunowie” co po francusku oznacza „nianię”.
1. Chaperony molekularne i ich rola w zapobieganiu denaturacji białek.
Chaperony (CH) są klasyfikowane według masy ich podjednostek. Chaperony o dużej masie cząsteczkowej mają masę od 60 do 110 kDa. Wśród nich najczęściej badane są trzy klasy: Sh-60, Sh-70 i Sh-90. Każda klasa obejmuje rodzinę powiązanych białek. Zatem Sh-70 obejmuje białka o masie cząsteczkowej od 66 do 78 kDa. Chaperony o niskiej masie cząsteczkowej mają masę cząsteczkową od 40 do 15 kDa.
Wśród opiekunów są składowy białka, których wysoka podstawowa synteza nie zależy od wpływu stresu na komórki organizmu, oraz indukowalny, którego synteza w normalnych warunkach jest słaba, ale gwałtownie wzrasta pod wpływem stresu. Indukowalne białka opiekuńcze nazywane są także „białkami szoku cieplnego”, ponieważ po raz pierwszy odkryto je w komórkach narażonych na działanie wysokich temperatur. W komórkach, ze względu na duże stężenie białek, spontaniczna reaktywacja częściowo zdenaturowanych białek jest utrudniona. Sh-70 może zapobiegać denaturacji i pomagać w przywracaniu natywnej konformacji białek. Chaperony molekularne-70- wysoce konserwatywna klasa białek występująca we wszystkich częściach komórki: cytoplazmie, jądrze, retikulum endoplazmatycznym, mitochondriach. Na końcu karboksylowym pojedynczego łańcucha polipeptydowego Ш-70 znajduje się region będący rowkiem zdolnym do interakcji z peptydami o długości
od 7 do 9 reszt aminokwasowych wzbogaconych rodnikami hydrofobowymi. Takie regiony w białkach globularnych występują w przybliżeniu co 16 aminokwasów. Sh-70 jest w stanie chronić białka przed inaktywacją temperaturową oraz przywracać konformację i aktywność częściowo zdenaturowanych białek.
2. Rola białek opiekuńczych w zwijaniu białek. Podczas syntezy białka na rybosomie, N-końcowy region polipeptydu jest syntetyzowany przed C-końcowym. Aby utworzyć natywną konformację, wymagana jest pełna sekwencja aminokwasów białka. W procesie syntezy białek chaperony-70, dzięki strukturze swojego centrum aktywnego, są w stanie zamykać obszary polipeptydu podatne na agregację, wzbogacone w hydrofobowe rodniki aminokwasowe, aż do zakończenia syntezy (Rysunek 1.27, A ).
 Ryż. 1,27. Udział białek opiekuńczych w fałdowaniu białek
Ryż. 1,27. Udział białek opiekuńczych w fałdowaniu białek
A - udział chaperonów-70 w zapobieganiu oddziaływaniom hydrofobowym pomiędzy odcinkami syntetyzowanego polipeptydu; B - tworzenie natywnej konformacji białka w kompleksie chaperonowym
Wiele białek wielkocząsteczkowych o złożonej konformacji, takiej jak struktura domeny, składa się w specjalnej przestrzeni utworzonej przez Sh-60. Ř-60 funkcjonują jako kompleks oligomeryczny składający się z 14 podjednostek. Tworzą dwa puste pierścienie, z których każdy składa się z siedmiu podjednostek, pierścienie te są ze sobą połączone. Każda podjednostka Sh-60 składa się z trzech domen: wierzchołkowej (wierzchołkowej), wzbogaconej rodnikami hydrofobowymi skierowanymi w stronę wnęki pierścienia, pośredniej i równikowej (ryc. 1.28).
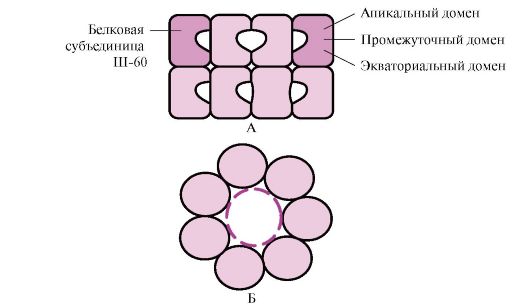 Ryż. 1,28. Struktura kompleksu opiekuńczego składającego się z 14 Ш-60
Ryż. 1,28. Struktura kompleksu opiekuńczego składającego się z 14 Ш-60
A - widok z boku; B - widok z góry
Zsyntetyzowane białka, które mają na powierzchni elementy charakterystyczne dla rozwiniętych cząsteczek, w szczególności rodniki hydrofobowe, dostają się do wnęki pierścieni opiekuńczych. W specyficznym środowisku tych wnęk poszukuje się możliwych konformacji, aż do znalezienia jedynej, najkorzystniejszej energetycznie (rys. 1.27, B). Towarzyszy tworzeniu się konformacji i uwalnianiu białek Hydroliza ATP w rejonie równikowym. Zazwyczaj takie fałdowanie zależne od opiekunów wymaga znacznej ilości energii.
Oprócz udziału w tworzeniu trójwymiarowej struktury białek i renatywacji częściowo zdenaturowanych białek, chaperony są również niezbędne do zachodzenia tak podstawowych procesów jak składanie białek oligomerycznych, rozpoznawanie i transport zdenaturowanych białek do lizosomów, transport białek przez błony i udział w regulacji aktywności kompleksów białkowych.
TEMAT 1.6. RÓŻNORODNOŚĆ BIAŁEK. RODZINY BIAŁEK: PRZYKŁAD IMMUNOGLOBULIN
1. Białka odgrywają kluczową rolę w życiu poszczególnych komórek i wszystkim organizm wielokomórkowy, a ich funkcje są zaskakująco różnorodne. Decydują o tym cechy struktury pierwszorzędowej i konformacji białek, unikalna struktura centrum aktywnego oraz zdolność wiązania określonych ligandów.
Tylko bardzo mała część wszystkich możliwych wariantów łańcuchów peptydowych może przyjąć stabilną strukturę przestrzenną; większość
z nich może przyjmować wiele konformacji o w przybliżeniu tej samej energii Gibbsa, ale o różnych właściwościach. Wybrano strukturę pierwotną większości znanych białek ewolucja biologiczna, zapewnia wyjątkową stabilność jednej z konformacji, która określa specyfikę funkcjonowania tego białka.
2. Rodziny białek. W obrębie tego samego gatunku biologicznego podstawienia reszt aminokwasowych mogą prowadzić do powstania różnych białek, które pełnią powiązane funkcje i mają sekwencje homologiczne aminokwasy. Takie pokrewne białka mają uderzająco podobne konformacje: liczbę i względne położenie α-helis i/lub struktur β, a większość zwojów i zagięć łańcuchów polipeptydowych jest podobna lub identyczna. Białka z homologicznymi regionami łańcucha polipeptydowego, podobną konformacją i powiązanymi funkcjami dzieli się na rodziny białek. Przykłady rodzin białek: proteinazy serynowe, rodzina immunoglobulin, rodzina mioglobin.
Proteinazy serynowe- rodzina białek pełniących funkcję enzymów proteolitycznych. Należą do nich enzymy trawienne – chymotrypsyna, trypsyna, elastaza i wiele czynników krzepnięcia krwi. Białka te mają identyczne aminokwasy w 40% swoich pozycji i bardzo podobną konformację (ryc. 1.29).

Ryż. 1,29. Struktury przestrzenne elastaza (A) i chymotrypsyna (B)
Niektóre podstawienia aminokwasów doprowadziły do zmian w specyficzności substratowej tych białek i pojawienia się różnorodności funkcjonalnej w obrębie rodziny.
3. Rodzina immunoglobulin. W funkcjonowaniu układu odpornościowego ogromną rolę odgrywają białka z nadrodziny immunoglobulin, do której zaliczają się trzy rodziny białek:
Przeciwciała (immunoglobuliny);
Receptory limfocytów T;
Białka głównego kompleksu zgodności tkankowej - klasy MHC 1 i 2 (Major Histocompatibility Complex).
Wszystkie te białka mają budowę domenową, składają się z homologicznych domen immunopodobnych i pełnią podobne funkcje: oddziałują z obcymi strukturami, albo rozpuszczonymi we krwi, limfie, płynie międzykomórkowym (przeciwciała), albo znajdującymi się na powierzchni komórek (własnych lub zagraniczny).
4. Przeciwciała- specyficzne białka wytwarzane przez limfocyty B w odpowiedzi na przedostanie się obcej struktury do organizmu, tzw antygen.
Cechy struktury przeciwciał
Najprostsze cząsteczki przeciwciał składają się z czterech łańcuchów polipeptydowych: dwóch identycznych lekkich - L, zawierających około 220 aminokwasów i dwóch identycznych ciężkich - H, składających się z 440-700 aminokwasów. Wszystkie cztery łańcuchy w cząsteczce przeciwciała są połączone wieloma wiązaniami niekowalencyjnymi i czterema wiązaniami dwusiarczkowymi (ryc. 1.30).
Łańcuchy lekkie przeciwciał składają się z dwóch domen: domeny zmiennej (VL), zlokalizowanej w regionie N-końcowym łańcucha polipeptydowego i domeny stałej (CL), zlokalizowanej na C-końcu. Łańcuchy ciężkie mają zwykle cztery domeny: jedną zmienną (VH), zlokalizowaną na N-końcu i trzy domeny stałe (CH1, CH2, CH3) (patrz ryc. 1.30). Każda domena immunoglobulinowa ma nadbudowę arkusza β, w której dwie reszty cysteinowe są połączone wiązaniem dwusiarczkowym.
Pomiędzy dwiema domenami stałymi CH1 i CH2 znajduje się region zawierający duża liczba reszty proliny, które zapobiegają tworzeniu się struktury drugorzędowej i oddziaływaniu sąsiadujących łańcuchów H w tym segmencie. Ten region zawiasowy zapewnia elastyczność cząsteczki przeciwciała. Pomiędzy domenami zmiennymi łańcucha ciężkiego i lekkiego znajdują się dwa identyczne miejsca wiązania antygenu (miejsca aktywne wiązania antygenów), dlatego takie przeciwciała często nazywane są biwalenty. W wiązaniu antygenu z przeciwciałem nie bierze udziału cała sekwencja aminokwasów regionów zmiennych obu łańcuchów, ale jedynie 20-30 aminokwasów znajdujących się w regionach hiperzmiennych każdego łańcucha. To właśnie te regiony określają unikalną zdolność każdego typu przeciwciał do interakcji z odpowiednim antygenem komplementarnym.
Przeciwciała są jedną z linii obrony organizmu przed inwazją obcych organizmów. Ich funkcjonowanie można podzielić na dwa etapy: pierwszy etap to rozpoznanie i związanie antygenu na powierzchni obcych organizmów, co jest możliwe dzięki obecności w strukturze przeciwciała miejsc wiążących antygen; drugi etap to inicjacja procesu inaktywacji i zniszczenia antygenu. Specyficzność drugiego etapu zależy od klasy przeciwciał. Wyróżnia się pięć klas łańcuchów ciężkich, różniących się między sobą budową domen stałych: α, δ, ε, γ i μ, według których wyróżnia się pięć klas immunoglobulin: A, D, E, G i M.
Cechy strukturalne łańcuchów ciężkich nadają regionom zawiasowym i regionom C-końcowym łańcuchów ciężkich konformację charakterystyczną dla każdej klasy. Po związaniu antygenu z przeciwciałem zmiany konformacyjne w domenach stałych określają ścieżkę usuwania antygenu.
 Ryż. 1. 30. Struktura domenowa IgG
Ryż. 1. 30. Struktura domenowa IgG
Immunoglobuliny M
Immunoglobuliny M mają dwie formy.
Forma monomeryczna- I klasa przeciwciał wytwarzanych przez rozwijające się limfocyty B. Następnie wiele limfocytów B zaczyna wytwarzać przeciwciała innej klasy, ale z tym samym miejscem wiązania antygenu. IgM jest osadzone w błonie i działa jako receptor rozpoznający antygen. Integracja IgM z błoną komórkową jest możliwa dzięki obecności 25 hydrofobowych reszt aminokwasowych w części ogonowej regionu.
Wydzielnicza postać IgM zawiera pięć monomerycznych podjednostek połączonych ze sobą wiązaniami dwusiarczkowymi i dodatkowym polipeptydowym łańcuchem J (ryc. 1.31). Łańcuchy ciężkie monomerów tej postaci nie zawierają ogona hydrofobowego. Pentamer ma 10 miejsc wiązania antygenu i dlatego skutecznie rozpoznaje i usuwa antygen, który jako pierwszy dostanie się do organizmu. Wydzielnicza postać IgM to główna klasa przeciwciał wydzielanych do krwi podczas pierwotnej odpowiedzi immunologicznej. Wiązanie IgM z antygenem zmienia konformację IgM i indukuje jej związanie z pierwszym białkowym składnikiem układu dopełniacza (układ dopełniacza to zespół białek biorących udział w niszczeniu antygenu) i aktywację tego układu. Jeśli antygen znajduje się na powierzchni mikroorganizmu, układ dopełniacza powoduje naruszenie integralności Błona komórkowa i śmierć komórki bakteryjnej.
Immunoglobuliny G
Ilościowo ta klasa immunoglobulin przeważa we krwi (75% wszystkich Ig). IgG – monomery, główna klasa przeciwciał wydzielanych do krwi podczas wtórnej odpowiedzi immunologicznej. Po interakcji IgG z antygenami powierzchniowymi mikroorganizmów kompleks antygen-przeciwciało jest w stanie wiązać i aktywować białka układu dopełniacza lub oddziaływać ze specyficznymi receptorami makrofagów i neutrofili. Interakcja z fagocytami prowadzi
 Ryż. 1.31. Struktura wydzielniczej formy IgM
Ryż. 1.31. Struktura wydzielniczej formy IgM
na absorpcję kompleksów antygen-przeciwciało i ich zniszczenie w fagosomach komórkowych. IgG to jedyna klasa przeciwciał, która jest w stanie przeniknąć przez barierę łożyskową i zapewnić wewnątrzmaciczną ochronę płodu przed infekcjami.
Immunoglobuliny A
Główna klasa przeciwciał występujących w wydzielinach (mleko, ślina, wydzieliny dróg oddechowych i jelit). IgA jest wydzielana głównie w postaci dimerycznej, gdzie monomery są połączone ze sobą dodatkowym łańcuchem J (ryc. 1.32).
IgA nie oddziałuje z układem dopełniacza i komórkami fagocytarnymi, ale wiążąc się z mikroorganizmami, przeciwciała uniemożliwiają ich przyłączenie do komórek nabłonkowych i przenikanie do organizmu.
Immunoglobuliny E
Immunoglobuliny E są reprezentowane przez monomery, w których ciężkie łańcuchy ε zawierają, podobnie jak łańcuchy μ immunoglobulin M, jedną domenę zmienną i cztery stałe. Po wydzieleniu IgE wiąże się ze swoimi
 Ryż. 1,32. Struktura IgA
Ryż. 1,32. Struktura IgA
Regiony C-końcowe z odpowiednimi receptorami na powierzchni komórek tucznych i bazofilów. W rezultacie stają się receptorami dla antygenów na powierzchni tych komórek (ryc. 1.33).
 Ryż. 1,33. Oddziaływanie IgE z antygenem na powierzchni komórek tucznych
Ryż. 1,33. Oddziaływanie IgE z antygenem na powierzchni komórek tucznych
Po związaniu się antygenu z odpowiednimi miejscami wiązania antygenu IgE, komórki otrzymują sygnał do biologicznego wydzielania substancje czynne(histamina, serotonina), które w dużej mierze odpowiadają za rozwój reakcji zapalnej i manifestację reakcji alergicznych, takich jak astma, pokrzywka, katar sienny.
Immunoglobuliny D
Immunoglobuliny D występują w bardzo małych ilościach w surowicy i są monomerami. Ciężkie łańcuchy δ mają jedną domenę zmienną i trzy domeny stałe. IgD działają jako receptory dla limfocytów B; inne funkcje są nadal nieznane. Oddziaływanie specyficznych antygenów z receptorami na powierzchni limfocytów B (IgD) prowadzi do przekazania tych sygnałów do komórki i aktywacji mechanizmów zapewniających proliferację danego klonu limfocytów.
TEMAT 1.7. WŁAŚCIWOŚCI FIZYCZNE I CHEMICZNE BIAŁEK ORAZ METODY ICH SEPARACJI
1. Poszczególne białka różnią się właściwościami fizycznymi i chemicznymi:
Kształt cząsteczek;
Waga molekularna;
Całkowity ładunek, którego wielkość zależy od stosunku grup anionowych i kationowych aminokwasów;
Stosunek polarnych i niepolarnych rodników aminokwasowych na powierzchni cząsteczek;
Stopnie odporności na różne czynniki denaturujące.
2. Rozpuszczalność białka zależy od właściwości wymienionych białek, a także od składu środowiska, w którym białko jest rozpuszczane (wartości pH, skład soli, temperatura, obecność innych substancji organicznych mogących oddziaływać z białkiem). Wielkość ładunku cząsteczek białek jest jednym z czynników wpływających na ich rozpuszczalność. Kiedy ładunek w punkcie izoelektrycznym zostanie utracony, białka łatwiej agregują i wytrącają się. Jest to szczególnie typowe dla białek zdenaturowanych, w których na powierzchni pojawiają się hydrofobowe rodniki aminokwasowe.
Na powierzchni cząsteczki białka znajdują się zarówno dodatnio, jak i ujemnie naładowane rodniki aminokwasów. Liczba tych grup, a co za tym idzie całkowity ładunek białek, zależy od pH ośrodka, tj. stosunek stężeń grup H+ - i OH -. W kwaśnym środowisku Wzrost stężenia H+ prowadzi do zahamowania dysocjacji grup karboksylowych -COO - + H+ > - COOH i zmniejszenia ładunku ujemnego białek. W środowisku zasadowym wiązanie nadmiaru OH - przez protony powstałe podczas dysocjacji grup aminowych -NH 3 + + OH - - NH 2 + H 2 O z utworzeniem wody prowadzi do zmniejszenia ładunku dodatniego białek . Nazywa się wartość pH, przy której białko ma ładunek zerowy netto punkt izoelektryczny (IEP). W IET liczba grup naładowanych dodatnio i ujemnie jest taka sama, tj. białko jest w stanie izoelektrycznym.
3. Rozdzielanie poszczególnych białek. Cechy budowy i funkcjonowania organizmu zależą od zestawu syntetyzowanych w nim białek. Badanie struktury i właściwości białek nie jest możliwe bez ich izolacji z komórki i oczyszczenia z innych białek organiczne molekuły. Etapy izolacji i oczyszczania poszczególnych białek:
Zniszczenie komórek badanej tkanki i uzyskania homogenatu.
Podział homogenatu na frakcje przez wirowanie, uzyskując frakcję jądrową, mitochondrialną, cytozolową lub inną zawierającą pożądane białko.
Selektywna denaturacja termiczna- krótkotrwałe ogrzewanie roztworu białka, podczas którego można usunąć część zdenaturowanych zanieczyszczeń białkowych (o ile białko jest stosunkowo termostabilne).
Solenie. Różne białka wytrącają się przy różnym stężeniu soli w roztworze. Stopniowo zwiększając stężenie soli, można otrzymać szereg odrębnych frakcji, w których w jednej z nich przeważa zawartość izolowanego białka. Do frakcjonowania białek najczęściej stosuje się siarczan amonu. Białka o najmniejszej rozpuszczalności wytrącają się przy niskich stężeniach soli.
Filtracja żelowa- metoda przesiewania cząsteczek przez spęczniałe granulki Sephadexu (trójwymiarowe łańcuchy polisacharydowe dekstranu posiadające pory). Szybkość, z jaką białka przechodzą przez kolumnę wypełnioną Sephadexem, będzie zależała od ich masy cząsteczkowej: im mniejsza masa cząsteczek białka, tym łatwiej wnikają one do granulek i pozostają tam dłużej niż więcej masy, tym szybciej wymywają się z kolumny.
Ultrawirowanie- metoda polegająca na umieszczeniu białek w probówce wirówkowej w rotorze ultrawirówki. Kiedy rotor się obraca, szybkość sedymentacji białek jest proporcjonalna do ich masy cząsteczkowej: frakcje białek cięższych znajdują się bliżej dna probówki, lżejsze - bliżej powierzchni.
Elektroforeza- metoda oparta na różnicach prędkości ruchu białek w polu elektrycznym. Wartość ta jest proporcjonalna do ładunku białek. Elektroforezę białek przeprowadza się na papierze (w tym przypadku prędkość ruchu białek jest proporcjonalna tylko do ich ładunku) lub w żelu poliakryloamidowym o określonej wielkości porów (szybkość ruchu białek jest proporcjonalna do ich ładunku i masy cząsteczkowej) .
Chromatografia jonowymienna- metoda frakcjonowania polegająca na wiązaniu zjonizowanych grup białek z przeciwnie naładowanymi grupami żywic jonowymiennych (nierozpuszczalnych materiałów polimerowych). Siła wiązania białka z żywicą jest proporcjonalna do ładunku białka. Białka zaadsorbowane na polimerze jonowymiennym można wypłukać wraz ze wzrostem stężenia roztworów NaCl; im niższy ładunek białka, tym niższe stężenie NaCl wymagane do wypłukania białka związanego z grupami jonowymi żywicy.
Chromatografii powinowactwa- najbardziej specyficzna metoda izolacji poszczególnych białek.Ligand białka jest kowalencyjnie przyłączony do obojętnego polimeru. Kiedy roztwór białka przepuszczany jest przez kolumnę z polimerem, na kolumnie adsorbowane jest wyłącznie białko specyficzne dla danego liganda, ze względu na komplementarne wiązanie białka z ligandem.
Dializa- metoda służąca do usuwania związków drobnocząsteczkowych z roztworu izolowanego białka. Metoda opiera się na niezdolności białek do przejścia przez półprzepuszczalną membranę, w przeciwieństwie do substancji o niskiej masie cząsteczkowej. Służy do oczyszczania białek z zanieczyszczeń drobnocząsteczkowych, np. soli po wysalaniu.
ZADANIA NA PRACĘ POZASZKOLNĄ
1. Wypełnij tabelę. 1.4.
Tabela 1.4. Analiza porównawcza struktura i funkcje pokrewnych białek - mioglobiny i hemoglobiny
a) zapamiętaj strukturę aktywnego centrum Mb i Hb. Jaką rolę odgrywają hydrofobowe rodniki aminokwasowe w tworzeniu centrów aktywnych tych białek? Opisać budowę centrum aktywnego Mb i Hb oraz mechanizmy przyłączania do niego ligandów. Jaką rolę odgrywają reszty His F 8 i His E 7 w funkcjonowaniu centrum aktywnego Mv iHv?
b) jakie nowe właściwości w porównaniu z monomeryczną mioglobiną ma blisko spokrewnione białko oligomeryczne, hemoglobina? Wyjaśnić rolę kooperatywnych zmian w konformacji protomerów w cząsteczce hemoglobiny, wpływ stężenia CO 2 i protonów na powinowactwo hemoglobiny do tlenu, a także rolę 2,3-BPG w allosterycznej regulacji funkcji Hb .
2. Scharakteryzuj chaperony molekularne, zwracając uwagę na związek pomiędzy ich strukturą i funkcją.
3. Jakie białka dzielimy na rodziny? Na przykładzie rodziny immunoglobulin zidentyfikuj podobne cechy strukturalne i powiązane funkcje białek tej rodziny.
4. Oczyszczone pojedyncze białka są często wymagane do celów biochemicznych i leczniczych. Wyjaśnij które fizyczne i chemiczne właściwości białka opierają się na metodach stosowanych do ich rozdzielania i oczyszczania.
ZADANIA SAMOKONTROLI
1. Wybierz prawidłowe odpowiedzi.
Funkcje hemoglobiny:
A. Transport O 2 z płuc do tkanek B. Transport H + z tkanek do płuc
B. Utrzymanie stałego pH krwi. D. Transport CO 2 z płuc do tkanek
D. Transport CO 2 z tkanek do płuc
2. Wybierz prawidłowe odpowiedzi. Ligandα -protomer Hb to: A. Hem
B. Tlen
B. CO G. 2,3-BPG
D. β-Protomer
3. Wybierz prawidłowe odpowiedzi.
Hemoglobina w przeciwieństwie do mioglobiny:
A. Ma strukturę czwartorzędową
B. Strukturę drugorzędową reprezentują jedynie α-helisy
B. Należy do białek złożonych
D. Oddziałuje z ligandem allosterycznym. D. Kowalencyjnie związany z hemem
4. Wybierz prawidłowe odpowiedzi.
Powinowactwo Hb do O2 maleje:
A. Gdy dodana zostanie jedna cząsteczka O 2. B. Gdy zostanie usunięta jedna cząsteczka O 2
B. Podczas interakcji z 2,3-BPG
D. Po przyłączeniu do protomerów H + D. Gdy stężenie 2,3-BPG maleje
5. Mecz.
Typy HB charakteryzują się:
A. W formie deoksy tworzy agregaty włókniste B. Zawiera dwa łańcuchy α i dwa δ
B. Dominująca forma Hb w erytrocytach dorosłych D. Zawiera hem z Fe+ 3 w centrum aktywnym
D. Zawiera dwa łańcuchy α i dwa łańcuchy γ 1. HbA 2.
6. Mecz.
Ligandy Hb:
A. Wiąże się z Hb w centrum allosterycznym
B. Ma bardzo duże powinowactwo do miejsca aktywnego Hb
B. Łącząc zwiększa powinowactwo Hb do O 2 G. Utlenia Fe+ 2 do Fe+ 3
D. Formularze wiązanie kowalencyjne z gisF8
7. Wybierz prawidłowe odpowiedzi.
Opiekunowie:
A. Białka obecne we wszystkich częściach komórki
B. Synteza wzrasta pod wpływem stresu
B. Uczestniczyć w hydrolizie zdenaturowanych białek
D. Uczestniczyć w utrzymaniu natywnej konformacji białek
D. Tworzą organelle, w których powstaje konformacja białek.
8. Mecz. Immunoglobuliny:
A. Forma wydzielnicza jest pentameryczna.
B. Klasa Ig, która przenika przez barierę łożyskową
B. Ig - receptor komórek tucznych
D. Główna klasa Ig obecna w wydzielinach komórek nabłonkowych. D. Receptor limfocytów B, którego aktywacja zapewnia proliferację komórek
9. Wybierz prawidłowe odpowiedzi.
Immunoglobuliny E:
A. Wytwarzane przez makrofagi. B. Mają ciężkie łańcuchy ε.
B. Osadzone w błonie limfocytów T
D. Działają jako błonowe receptory antygenów na komórkach tucznych i bazofilach
D. Odpowiedzialny za reakcje alergiczne
10. Wybierz prawidłowe odpowiedzi.
Metoda rozdziału białek opiera się na różnicach w ich masie cząsteczkowej:
A. Filtracja żelowa
B. Ultrawirowanie
B. Elektroforeza w żelu poliakryloamidowym. D. Chromatografia jonowymienna
D. Chromatografia powinowactwa
11. Wybierz poprawną odpowiedź.
Metoda rozdziału białek opiera się na różnicach w ich rozpuszczalności w wodzie:
A. Filtracja żelowa B. Wysalanie
B. Chromatografia jonowymienna. D. Chromatografia powinowactwa
D. Elektroforeza w żelu poliakryloamidowym
STANDARDY ODPOWIEDZI NA „ZADANIA SAMOKONTROLI”
1. A, B, C, D
2. A, B, C, D
5. 1-B, 2-A, 3-G
6. 1-B, 2-B, 3-A
7. A, B, D, D
8. 1-G; 2-B, 3-B
PODSTAWOWE TERMINY I POJĘCIA
1. Białka oligomeryczne, protomer, czwartorzędowa struktura białek
2. Kooperacyjne zmiany w konformacji protomeru
3. Efekt Bohra
4. Allosteryczna regulacja funkcji białek, centrum allosteryczne i efektor allosteryczny
5. Chaperony molekularne, białka szoku cieplnego
6. Rodziny białek (proteazy serynowe, immunoglobuliny)
7. Zależność IgM-, G-, E-, A-struktura-funkcja
8. Ładunek całkowity białek, punkt izoelektryczny białek
9. Elektroforeza
10. Solenie
11. Filtracja żelowa
12. Chromatografia jonowymienna
13. Ultrawirowanie
14. Chromatografia powinowactwa
15. Elektroforeza białek osocza krwi
ZADANIA DO PRACY W KLASIE
1. Porównaj zależności stopnia nasycenia hemoglobiny (Hb) i mioglobiny (Mb) tlenem od jej ciśnienia parcjalnego w tkankach
 Ryż. 1,34. Zależność nasycenia Mv iNHtlenu z jego ciśnienia cząstkowego
Ryż. 1,34. Zależność nasycenia Mv iNHtlenu z jego ciśnienia cząstkowego
Należy pamiętać, że kształt krzywych nasycenia białka tlenem jest inny: dla mioglobiny - hiperbola, dla hemoglobiny - kształt esowaty.
1. porównać wartości ciśnienia cząstkowego tlenu, przy którym Mb i Hb są nasycone O 2 o 50%. Które z tych białek ma większe powinowactwo do O2?
2. Jakie cechy strukturalne Mb decydują o jego wysokim powinowactwie do O 2?
3. Jakie cechy strukturalne HB pozwalają na uwalnianie O2 w kapilarach spoczynkowych tkanek (przy stosunkowo wysokim ciśnieniu parcjalnym O2) i gwałtownie zwiększają to uwalnianie w pracujących mięśniach? Jaka właściwość białek oligomerycznych zapewnia taki efekt?
4. Oblicz, jaką ilość O 2 (w%) utlenionej hemoglobiny oddaje mięśnie odpoczywające i pracujące?
5. wyciągać wnioski na temat związku pomiędzy strukturą białka a jego funkcją.
2. Ilość tlenu uwalnianego przez hemoglobinę w naczyniach włosowatych zależy od intensywności procesów katabolicznych w tkankach (efekt Bohra). W jaki sposób zmiany w metabolizmie tkanek regulują powinowactwo Hb do O2? Wpływ CO 2 i H+ na powinowactwo Hb do O 2
 1. opisać efekt Bohra.
1. opisać efekt Bohra.
2. w jakim kierunku przebiega proces pokazany na schemacie:
a) w naczyniach włosowatych płuc;
b) w naczyniach włosowatych tkankowych?
3. Jakie jest fizjologiczne znaczenie efektu Bohra?
4. Dlaczego oddziaływanie Hb z H+ w miejscach odległych od hemu zmienia powinowactwo białka do O 2?
3. Powinowactwo Hb do O2 zależy od stężenia jej ligandu – 2,3-bisfosfoglicerynianu, który jest allosterycznym regulatorem powinowactwa Hb do O2. Dlaczego interakcja ligandu w miejscu odległym od miejsca aktywnego wpływa na funkcję białka? W jaki sposób 2,3-BPG reguluje powinowactwo Hb do O2? Aby rozwiązać problem, odpowiedz na następujące pytania:
1. gdzie i z czego syntetyzowany jest 2,3-bisfosfoglicerynian (2,3-BPG)? Napisz jego wzór, wskaż ładunek tej cząsteczki.
2. Z jaką formą hemoglobiny (oksy czy deoksy) wchodzi w interakcję BPG i dlaczego? W jakiej części cząsteczki Hb zachodzi interakcja?
3. w jakim kierunku przebiega proces pokazany na schemacie?
a) w naczyniach włosowatych tkankowych;
b) w naczyniach włosowatych płuc?
4. gdzie stężenie kompleksu powinno być wyższe
Nv-2,3-BFG:
a) w naczyniach włosowatych mięśni w stanie spoczynku,
b) w naczyniach włosowatych pracujących mięśni (pod warunkiem takiego samego stężenia BPG w erytrocytach)?
5. Jak zmieni się powinowactwo HB do tlenu, gdy człowiek przystosuje się do warunków wysokogórskich, jeśli wzrośnie stężenie BPG w erytrocytach? Jakie jest fizjologiczne znaczenie tego zjawiska?
4. Zniszczenie 2,3-BPG podczas przechowywania zakonserwowanej krwi upośledza funkcje HB. Jak zmieni się powinowactwo HB do O 2 w zakonserwowanej krwi, jeśli stężenie 2,3-BPG w erytrocytach może spaść z 8 do 0,5 mmol/l. Czy można przetaczać taką krew ciężko chorym, jeśli stężenie 2,3-BPG zostanie przywrócone dopiero po trzech dniach? Czy możliwe jest przywrócenie funkcji czerwonych krwinek poprzez dodanie do krwi 2,3-BPG?
5. Zapamiętaj strukturę najprostszych cząsteczek immunoglobulin. Jaką rolę odgrywają immunoglobuliny w układzie odpornościowym? Dlaczego Igs często nazywane są biwalentami? Jak struktura Ig wiąże się z ich funkcją? (Opisz na przykładzie klasy immunoglobulin.)
Właściwości fizykochemiczne białek i metody ich rozdzielania.
6. Jak ładunek netto białka wpływa na jego rozpuszczalność?
a) określić całkowity ładunek peptydu przy pH 7
Ala-Glu-Tre-Pro-Asp-Liz-Cis
b) jak zmieni się ładunek tego peptydu przy pH >7, pH<7, рН <<7?
c) jaki jest punkt izoelektryczny białka (IEP) i w jakim środowisku się ono znajduje?
IET tego peptydu?
d) przy jakiej wartości pH będzie zaobserwowana najmniejsza rozpuszczalność tego peptydu.
7. Dlaczego kwaśne mleko, w przeciwieństwie do świeżego mleka, „zsiada się” po ugotowaniu (tj. wytrąca się kazeina białka mleka)? W świeżym mleku cząsteczki kazeiny mają ładunek ujemny.
8. Do separacji poszczególnych białek stosuje się filtrację żelową. Mieszaninę zawierającą białka A, B, C o masach cząsteczkowych odpowiednio 160 000, 80 000 i 60 000 poddano analizie metodą filtracji żelowej (ryc. 1.35). Spęcznione granulki żelu przepuszczają białka o masie cząsteczkowej mniejszej niż 70 000. Na jakiej zasadzie leży ta metoda separacji? Który wykres poprawnie przedstawia wyniki frakcjonowania? Wskaż kolejność, w jakiej białka A, B i C są uwalniane z kolumny.
 Ryż. 1,35. Zastosowanie filtracji żelowej do separacji białek
Ryż. 1,35. Zastosowanie filtracji żelowej do separacji białek
9. Na ryc. 1.36, A przedstawia schemat elektroforezy na papierze białek surowicy krwi osoby zdrowej. Względne ilości frakcji białkowych otrzymanych tą metodą wynoszą: albuminy 54-58%, α1-globuliny 6-7%, α2-globuliny 8-9%, β-globuliny 13%, γ-globuliny 11-12%.
 Ryż. 1.36 Elektroforeza na papierze białek osocza krwi osoby zdrowej (A) i pacjenta (B)
Ryż. 1.36 Elektroforeza na papierze białek osocza krwi osoby zdrowej (A) i pacjenta (B)
I - γ-globuliny; II - β-globuliny; III -α 2 -globulina; IV -α 2 -globulina; V - albuminy
Wielu chorobom towarzyszą ilościowe zmiany w składzie białek surowicy (dyproteinemia). Charakter tych zmian jest brany pod uwagę przy stawianiu diagnozy oraz ocenie ciężkości i stadium choroby.
1. Dlaczego białka uważa się za polimery?
Odpowiedź. Białka to polimery, czyli cząsteczki zbudowane jak łańcuchy z powtarzających się jednostek lub podjednostek monomeru, składające się z aminokwasów połączonych w określonej sekwencji wiązaniem peptydowym. Są podstawowymi i niezbędnymi składnikami wszystkich organizmów.
Istnieją białka proste (białka) i białka złożone (proteidy). Białka to białka, których cząsteczki zawierają wyłącznie składniki białkowe. Po całkowitej hydrolizie powstają aminokwasy.
Proteidy to złożone białka, których cząsteczki różnią się znacznie od cząsteczek białek tym, że oprócz samego składnika białkowego zawierają składnik o niskiej masie cząsteczkowej o charakterze niebiałkowym
2. Jakie znasz funkcje białek?
Odpowiedź. Białka pełnią funkcje: konstrukcyjną, energetyczną, katalityczną, ochronną, transportową, skurczową, sygnalizacyjną i inne.
Pytania po § 11
1. Jakie substancje nazywane są białkami?
Odpowiedź. Białka lub białka to polimery biologiczne, których monomerami są aminokwasy. Wszystkie aminokwasy posiadają grupę aminową (-NH2) i grupę karboksylową (-COOH) oraz różnią się budową i właściwościami rodników. Aminokwasy są połączone ze sobą wiązaniami peptydowymi, dlatego białka nazywane są także polipeptydami.
Odpowiedź. Cząsteczki białek mogą przybierać różne formy przestrzenne – konformacje, które reprezentują cztery poziomy ich organizacji. Liniowa sekwencja aminokwasów w łańcuchu polipeptydowym reprezentuje pierwotną strukturę białka. Jest unikalny dla każdego białka i określa jego kształt, właściwości i funkcje.
3. Jak powstają drugorzędowe, trzeciorzędowe i czwartorzędowe struktury białkowe?
Odpowiedź. Struktura drugorzędowa białka powstaje w wyniku tworzenia wiązań wodorowych pomiędzy grupami -CO- i -NH-. W tym przypadku łańcuch polipeptydowy skręca się w spiralę. Helisa może uzyskać konfigurację kulistą, ponieważ pomiędzy rodnikami aminokwasów w helisie powstają różne wiązania. Globula jest trzeciorzędową strukturą białka. Jeśli kilka globul połączy się w jeden złożony kompleks, powstaje struktura czwartorzędowa. Na przykład hemoglobina w ludzkiej krwi składa się z czterech kulek.
4. Co to jest denaturacja białek?
Odpowiedź. Naruszenie naturalnej struktury białka nazywa się denaturacją. Pod wpływem szeregu czynników (chemicznych, radioaktywnych, temperatury itp.) czwartorzędowe, trzeciorzędowe i drugorzędowe struktury białka mogą zostać zniszczone. Jeśli działanie czynnika ustanie, białko może przywrócić swoją strukturę. Jeśli działanie czynnika wzrasta, zniszczeniu ulega również pierwotna struktura białka - łańcuch polipeptydowy. Jest to proces nieodwracalny – białko nie jest w stanie przywrócić swojej struktury
5. Na jakiej podstawie dzieli się białka na proste i złożone?
Odpowiedź. Białka proste składają się wyłącznie z aminokwasów. Białka złożone mogą obejmować inne substancje organiczne: węglowodany (wówczas zwane glikoproteinami), tłuszcze (lipoproteiny), kwasy nukleinowe (nukleoproteiny).
6. Jakie znasz funkcje białek?
Odpowiedź. Funkcja konstrukcyjna (plastikowa). Białka są składnikiem strukturalnym błon biologicznych i organelli komórkowych, a także wchodzą w skład struktur nośnych ciała, włosów, paznokci i naczyń krwionośnych. Funkcja enzymatyczna. Białka pełnią rolę enzymów, czyli biologicznych katalizatorów, które przyspieszają tempo reakcji biochemicznych dziesiątki i setki milionów razy. Przykładem jest amylaza, która rozkłada skrobię na monosacharydy. Funkcja skurczowa (motoryczna). Odbywa się to za pomocą specjalnych białek kurczliwych, które zapewniają ruch komórek i struktur wewnątrzkomórkowych. Dzięki nim chromosomy poruszają się podczas podziału komórki, a wici i rzęski przemieszczają komórki pierwotniaków. Właściwości skurczowe białek aktyny i miozyny leżą u podstaw funkcjonowania mięśni. Funkcja transportowa. Białka biorą udział w transporcie cząsteczek i jonów w organizmie (hemoglobina przenosi tlen z płuc do narządów i tkanek, albumina surowicy bierze udział w transporcie kwasów tłuszczowych). Funkcja ochronna. Polega na ochronie organizmu przed uszkodzeniami i inwazją obcych białek i bakterii. Białka przeciwciał wytwarzane przez limfocyty tworzą obronę organizmu przed obcymi infekcjami; trombina i fibryna biorą udział w tworzeniu skrzepów krwi, pomagając w ten sposób organizmowi uniknąć dużych strat krwi. Funkcja regulacyjna. Odbywa się to za pomocą białek hormonalnych. Uczestniczą w regulacji aktywności komórek i wszystkich procesów życiowych organizmu. W ten sposób insulina reguluje poziom cukru we krwi i utrzymuje go na określonym poziomie. Funkcja sygnału. Białka wbudowane w błonę komórkową mają zdolność zmiany swojej struktury w odpowiedzi na podrażnienie. W ten sposób sygnały są przesyłane ze środowiska zewnętrznego do komórki. Funkcja energii. Jest ona realizowana przez białka niezwykle rzadko. Przy całkowitym rozkładzie 1 g białka można uwolnić 17,6 kJ energii. Białka są jednak związkiem bardzo cennym dla organizmu. Dlatego też rozkład białek następuje najczęściej na aminokwasy, z których budowane są nowe łańcuchy polipeptydowe. Białka hormonalne regulują aktywność komórki i wszystkie procesy życiowe organizmu. Tym samym w organizmie człowieka somatotropina bierze udział w regulacji wzrostu organizmu, insulina utrzymuje poziom glukozy we krwi na stałym poziomie.
7. Jaką rolę odgrywają białka hormonalne?
Odpowiedź. Funkcja regulacyjna jest nieodłącznie związana z białkami hormonalnymi (regulatorami). Regulują różne procesy fizjologiczne. Na przykład najbardziej znanym hormonem jest insulina, która reguluje poziom glukozy we krwi. Kiedy w organizmie nie ma wystarczającej ilości insuliny, pojawia się choroba zwana cukrzycą.
8. Jaką funkcję pełnią białka enzymatyczne?
Odpowiedź. Enzymy są katalizatorami biologicznymi, czyli przyspieszają reakcje chemiczne setki milionów razy. Enzymy mają ścisłą specyficzność wobec substancji, która reaguje. Każda reakcja jest katalizowana przez własny enzym.
9. Dlaczego białka są rzadko wykorzystywane jako źródło energii?
Odpowiedź. Monomery białkowe aminokwasów są cennymi surowcami do budowy nowych cząsteczek białek. Dlatego rzadko zdarza się całkowity rozkład polipeptydów na substancje nieorganiczne. W związku z tym funkcję energetyczną, polegającą na uwolnieniu energii po całkowitym rozpadzie, białka pełnią dość rzadko.
Białko jaja jest typowym białkiem. Dowiedz się, co się z nim stanie, jeśli zostanie wystawiony na działanie wody, alkoholu, acetonu, kwasu, zasady, oleju roślinnego, wysokiej temperatury itp.
Odpowiedź. W wyniku działania wysokiej temperatury na białko jaja, nastąpi denaturacja białka. Pod wpływem alkoholu, acetonu, kwasów lub zasad dzieje się mniej więcej to samo: białko koaguluje. Jest to proces, w którym trzeciorzędowa i czwartorzędowa struktura białka zostaje rozerwana na skutek zerwania wiązań wodorowych i jonowych.
W wodzie i oleju roślinnym białko zachowuje swoją strukturę.
Zmiel surową bulwę ziemniaka na miąższ. Weź trzy probówki i włóż do każdej niewielką ilość posiekanych ziemniaków.
Pierwszą probówkę należy umieścić w zamrażarce lodówki, drugą na dolnej półce lodówki, a trzecią w słoiku z ciepłą wodą (t = 40°C). Po 30 minutach wyjąć probówki i wrzucić do każdej niewielką ilość nadtlenku wodoru. Obserwuj, co dzieje się w każdej probówce. Wyjaśnij swoje wyniki
Odpowiedź. Doświadczenie to ilustruje aktywność enzymu katalazy w żywej komórce na nadtlenku wodoru. W wyniku reakcji wydziela się tlen. Dynamikę uwalniania pęcherzyków można wykorzystać do oceny aktywności enzymu.
Doświadczenie pozwoliło nam odnotować następujące wyniki:
Aktywność katalazy zależy od temperatury:
1. Probówka 1: nie ma pęcherzyków - dzieje się tak dlatego, że w niskich temperaturach komórki ziemniaka zapadają się.
2. Probówka nr 2: jest mała liczba pęcherzyków – ponieważ aktywność enzymu w niskich temperaturach jest niska.
3. Probówka 3: jest dużo pęcherzyków, temperatura jest optymalna, katalaza jest bardzo aktywna.
Do pierwszej probówki z ziemniakami wpuść kilka kropli wody, do drugiej kilka kropli kwasu (ocet stołowy), a do trzeciej zasady.
Obserwuj, co dzieje się w każdej probówce. Wyjaśnij swoje wyniki. Wyciągać wnioski.
Odpowiedź. Po dodaniu wody nic się nie dzieje, po dodaniu kwasu następuje pewne ciemnienie, po dodaniu zasady następuje „pienienie” - hydroliza alkaliczna.
Jak wiadomo, białka są podstawą powstania życia na naszej planecie. Ale to kropelka koacerwatu, składająca się z cząsteczek peptydów, stała się podstawą powstania żywych istot. Nie ulega to wątpliwości, ponieważ analiza składu wewnętrznego dowolnego przedstawiciela biomasy pokazuje, że substancje te są obecne we wszystkim: roślinach, zwierzętach, mikroorganizmach, grzybach, wirusach. Co więcej, są one bardzo różnorodne i mają charakter makromolekularny.
Struktury te mają cztery nazwy, wszystkie są synonimami:
- białka;
- białka;
- polipeptydy;
- peptydy.
Cząsteczki białka
Ich liczba jest naprawdę niezliczona. W tym przypadku wszystkie cząsteczki białka można podzielić na dwie duże grupy:
- proste - składają się wyłącznie z sekwencji aminokwasów połączonych wiązaniami peptydowymi;
- złożony - strukturę i strukturę białka charakteryzują dodatkowe grupy protolityczne (protetyczne), zwane także kofaktorami.
Jednocześnie złożone cząsteczki mają również swoją własną klasyfikację.
Gradacja złożonych peptydów
- Glikoproteiny są blisko spokrewnionymi związkami białka i węglowodanów. Grupy prostetyczne mukopolisacharydów są wplecione w strukturę cząsteczki.
- Lipoproteiny są złożonym związkiem białek i lipidów.
- Metaloproteiny - jony metali (żelaza, manganu, miedzi i innych) pełnią rolę grupy prostetycznej.
- Nukleoproteiny stanowią połączenie pomiędzy białkiem i kwasami nukleinowymi (DNA, RNA).
- Fosfoproteiny - konformacja białka i reszty kwasu ortofosforowego.
- Chromoproteiny są bardzo podobne do metaloprotein, jednak elementem wchodzącym w skład grupy prostetycznej jest cały kompleks kolorowy (czerwony - hemoglobina, zielony - chlorofil i tak dalej).
W każdej rozpatrywanej grupie struktura i właściwości białek są odmienne. Funkcje, które pełnią, różnią się również w zależności od rodzaju cząsteczki.

Struktura chemiczna białek
Z tego punktu widzenia białka to długi, masywny łańcuch reszt aminokwasowych połączonych ze sobą specyficznymi wiązaniami zwanymi wiązaniami peptydowymi. Od bocznych struktur kwasów odchodzą gałęzie zwane rodnikami. Tę strukturę molekularną odkrył E. Fischer na początku XXI wieku.
Później bardziej szczegółowo zbadano białka, strukturę i funkcje białek. Stało się jasne, że strukturę peptydu tworzy tylko 20 aminokwasów, ale można je łączyć na różne sposoby. Stąd różnorodność struktur polipeptydowych. Ponadto w procesie życia i pełnieniu swoich funkcji białka mogą ulegać szeregowi przemian chemicznych. W rezultacie zmieniają strukturę i pojawia się zupełnie nowy rodzaj połączenia.
Aby rozerwać wiązanie peptydowe, czyli rozbić białko i strukturę łańcuchów, należy wybrać bardzo rygorystyczne warunki (wysokie temperatury, kwasy lub zasady, katalizator). Wynika to z dużej siły w cząsteczce, a mianowicie w grupie peptydowej.

Wykrywanie struktury białka w laboratorium odbywa się za pomocą reakcji biuretowej - ekspozycji na świeżo wytrącony polipeptyd (II). Kompleks grupy peptydowej i jonu miedzi daje jasny fioletowy kolor.
Istnieją cztery główne organizacje strukturalne, z których każda ma swoje własne cechy strukturalne białek.
Poziomy organizacji: struktura pierwotna
Jak wspomniano powyżej, peptyd to sekwencja reszt aminokwasowych z inkluzjami lub bez, koenzymami. Zatem pierwotna jest struktura cząsteczki, która jest naturalnym, naturalnym, prawdziwie aminokwasem połączonym wiązaniami peptydowymi i niczym więcej. Oznacza to, że jest to polipeptyd o strukturze liniowej. Ponadto cechy strukturalne białek tego typu polegają na tym, że taka kombinacja kwasów decyduje o spełnianiu funkcji cząsteczki białka. Dzięki obecności tych cech możliwa jest nie tylko identyfikacja peptydu, ale także przewidywanie właściwości i roli zupełnie nowego, jeszcze nieodkrytego. Przykładami peptydów o naturalnej strukturze pierwszorzędowej są insulina, pepsyna, chymotrypsyna i inne.

Konformacja wtórna
Struktura i właściwości białek tej kategorii są nieco zróżnicowane. Taka struktura może powstać początkowo w naturze lub gdy pierwotna zostanie poddana silnej hydrolizie, temperaturze lub innym warunkom.
Ta konformacja ma trzy odmiany:
- Gładkie, regularne, stereoregularne skręty, zbudowane z reszt aminokwasowych, które owijają się wokół głównej osi połączenia. Łączą je tylko te, które powstają pomiędzy tlenem jednej grupy peptydowej i wodorem drugiej. Ponadto konstrukcję uważa się za poprawną ze względu na fakt, że zwoje powtarzają się równomiernie co 4 ogniwa. Taka konstrukcja może być leworęczna lub praworęczna. Jednak w większości znanych białek dominuje izomer prawoskrętny. Takie konformacje są zwykle nazywane strukturami alfa.
- Skład i struktura białek kolejnego typu różni się od poprzedniego tym, że wiązania wodorowe powstają nie pomiędzy resztami sąsiadującymi z jedną stroną cząsteczki, ale pomiędzy resztami znacznie odległymi i w dość dużej odległości. Z tego powodu cała struktura przybiera postać kilku falistych, wężowych łańcuchów polipeptydowych. Jest jedna cecha, którą musi wykazywać białko. Struktura aminokwasów na gałęziach powinna być jak najkrótsza, jak na przykład glicyna lub alanina. Ten typ konformacji wtórnej nazywany jest arkuszami beta ze względu na ich zdolność do sklejania się, tworząc wspólną strukturę.
- W biologii trzeci typ struktury białka to złożone, heterogenicznie rozproszone, nieuporządkowane fragmenty, które nie posiadają stereoregularności i są zdolne do zmiany struktury pod wpływem warunków zewnętrznych.
Nie zidentyfikowano żadnych przykładów białek, które w sposób naturalny mają strukturę drugorzędową.

Wykształcenie wyższe
Jest to dość złożona konformacja zwana „globulą”. Co to jest za białko? Jej budowa opiera się na strukturze drugorzędowej, jednak dodawane są nowe rodzaje oddziaływań pomiędzy atomami grup, a cała cząsteczka sprawia wrażenie fałdowania, skupiając się tym samym na tym, że grupy hydrofilowe są skierowane do globuli, a hydrofobowe te na zewnątrz.
To wyjaśnia ładunek cząsteczki białka w koloidalnych roztworach wody. Jakie rodzaje interakcji tu występują?
- Wiązania wodorowe - pozostają niezmienione pomiędzy tymi samymi częściami, co w strukturze wtórnej.
- interakcje - zachodzą, gdy polipeptyd jest rozpuszczony w wodzie.
- Przyciągania jonowe powstają pomiędzy różnie naładowanymi grupami reszt aminokwasowych (rodnikami).
- Oddziaływania kowalencyjne - mogą tworzyć się pomiędzy określonymi miejscami kwasowymi - cząsteczkami cysteiny, a raczej ich ogonami.
Zatem skład i strukturę białek o strukturze trzeciorzędowej można opisać jako łańcuchy polipeptydowe złożone w kuleczki, które zachowują i stabilizują swoją konformację na skutek różnego rodzaju oddziaływań chemicznych. Przykłady takich peptydów: kenaza fosfoglicerynianowa, tRNA, alfa-keratyna, fibroina jedwabiu i inne.
Struktura czwartorzędowa
Jest to jedna z najbardziej złożonych globul, jakie tworzą białka. Budowa i funkcje białek tego typu są bardzo różnorodne i specyficzne.
Jaka jest ta konformacja? Jest to kilka (w niektórych przypadkach dziesiątki) dużych i małych łańcuchów polipeptydowych, które powstają niezależnie od siebie. Ale potem, z powodu tych samych interakcji, które rozważaliśmy dla struktury trzeciorzędowej, wszystkie te peptydy skręcają się i przeplatają ze sobą. W ten sposób otrzymuje się złożone globule konformacyjne, które mogą zawierać atomy metali, grupy lipidowe i węglowodany. Przykłady takich białek: polimeraza DNA, otoczka białkowa wirusa tytoniowego, hemoglobina i inne.

Wszystkie badane przez nas struktury peptydowe posiadają własne metody identyfikacji w laboratorium, oparte na nowoczesnych możliwościach stosowania chromatografii, wirowania, mikroskopii elektronowej i optycznej oraz zaawansowanych technologiach komputerowych.
Wykonywane funkcje
Struktura i funkcje białek są ze sobą ściśle powiązane. Oznacza to, że każdy peptyd odgrywa określoną rolę, wyjątkową i specyficzną. Są też takie, które są w stanie wykonać kilka znaczących operacji jednocześnie w jednej żywej komórce. Można jednak wyrazić w uogólnionej formie główne funkcje cząsteczek białka w organizmach żywych:
- Zapewnienie ruchu. Organizmy jednokomórkowe, organelle lub niektóre typy komórek są zdolne do ruchu, kurczenia się i ruchu. Zapewniają to białka tworzące strukturę ich aparatu motorycznego: rzęski, wici i błona cytoplazmatyczna. Jeśli mówimy o komórkach niezdolnych do ruchu, to białka mogą przyczyniać się do ich skurczu (miozyna mięśniowa).
- Funkcja odżywcza lub rezerwowa. Polega na gromadzeniu się cząsteczek białka w jajach, zarodkach i nasionach roślin w celu dalszego uzupełnienia brakujących składników odżywczych. Po rozkładzie peptydy wytwarzają aminokwasy i substancje biologicznie czynne, które są niezbędne do prawidłowego rozwoju organizmów żywych.
- Funkcja energii. Oprócz węglowodanów, białka mogą również zapewnić siłę organizmowi. Rozpad 1 g peptydu uwalnia 17,6 kJ użytecznej energii w postaci kwasu adenozynotrójfosforowego (ATP), który jest zużywany na procesy życiowe.
- Sygnalizacja polega na uważnym monitorowaniu zachodzących procesów i przekazywaniu sygnałów z komórek do tkanek, z nich do narządów, z tych ostatnich do układów i tak dalej. Typowym przykładem jest insulina, która ściśle reguluje ilość glukozy we krwi.
- Funkcja receptora. Odbywa się to poprzez zmianę konformacji peptydu po jednej stronie membrany i zaangażowanie drugiego końca w restrukturyzację. Jednocześnie przesyłany jest sygnał i niezbędne informacje. Najczęściej białka takie osadzone są w błonach cytoplazmatycznych komórek i sprawują ścisłą kontrolę nad wszystkimi przechodzącymi przez nie substancjami. Dostarczają również informacji o zmianach chemicznych i fizycznych w środowisku.
- Funkcja transportowa peptydów. Odbywa się to za pośrednictwem białek kanałowych i białek transportowych. Ich rola jest oczywista – przenoszenie niezbędnych cząsteczek do miejsc o niskim stężeniu z części o większym stężeniu. Typowym przykładem jest transport tlenu i dwutlenku węgla przez narządy i tkanki przez białko hemoglobinę. Dokonują także dostarczania związków o niskiej masie cząsteczkowej przez błonę komórkową do wnętrza.
- Funkcja strukturalna. Jedna z najważniejszych funkcji pełnionych przez białko. Strukturę wszystkich komórek i ich organelli zapewniają peptydy. To one, niczym rama, wyznaczają kształt i strukturę. Ponadto wspierają go i modyfikują w razie potrzeby. Dlatego do wzrostu i rozwoju wszystkie żywe organizmy potrzebują białek w swojej diecie. Do takich peptydów zalicza się elastynę, tubulinę, kolagen, aktynę, keratynę i inne.
- Funkcja katalityczna. Dokonują tego enzymy. Liczne i różnorodne, przyspieszają wszelkie reakcje chemiczne i biochemiczne w organizmie. Bez ich udziału zwykłe jabłko w żołądku mogłoby zostać strawione zaledwie w dwa dni, najprawdopodobniej gnijąc przy tym. Pod wpływem katalazy, peroksydazy i innych enzymów proces ten zachodzi w ciągu dwóch godzin. Ogólnie rzecz biorąc, dzięki tej roli białek zachodzi anabolizm i katabolizm, czyli plastyka i

Rola ochronna
Istnieje kilka rodzajów zagrożeń, przed którymi białka mają chronić organizm.
Po pierwsze, traumatyczne odczynniki, gazy, cząsteczki, substancje o różnym spektrum działania. Peptydy potrafią z nimi oddziaływać chemicznie, przekształcając je w nieszkodliwą formę lub po prostu neutralizując.
Po drugie, fizyczne zagrożenie ranami - jeśli białko fibrynogen nie zostanie w odpowiednim czasie przekształcone w fibrynę w miejscu urazu, wówczas krew nie będzie krzepnąć, co oznacza, że nie nastąpi zator. Wręcz przeciwnie, będziesz potrzebować plazminy peptydowej, która może rozpuścić skrzep i przywrócić drożność naczynia.
Po trzecie, zagrożenie dla immunitetu. Niezwykle ważna jest struktura i znaczenie białek tworzących obronę immunologiczną. Przeciwciała, immunoglobuliny, interferony – to wszystko są ważne i istotne elementy układu limfatycznego i odpornościowego człowieka. Każda obca cząstka, szkodliwa cząsteczka, martwa część komórki lub cała struktura podlega natychmiastowemu badaniu przez związek peptydowy. Dlatego człowiek może samodzielnie, bez pomocy leków, codziennie chronić się przed infekcjami i prostymi wirusami.
Właściwości fizyczne
Struktura białka komórkowego jest bardzo specyficzna i zależy od pełnionej funkcji. Ale właściwości fizyczne wszystkich peptydów są podobne i sprowadzają się do następujących cech.
- Masa cząsteczki dochodzi do 1 000 000 daltonów.
- Układy koloidalne powstają w roztworze wodnym. Tam struktura nabiera ładunku, który może się różnić w zależności od kwasowości środowiska.
- Pod wpływem trudnych warunków (napromieniowanie, kwas lub zasada, temperatura itp.) są w stanie przejść do innych poziomów konformacji, czyli denaturacji. W 90% przypadków proces ten jest nieodwracalny. Istnieje jednak również przesunięcie odwrotne – renaturyzacja.
Są to główne właściwości właściwości fizycznych peptydów.
Są to wielkocząsteczkowe związki organiczne, biopolimery, zbudowane z 20 rodzajów reszt L-a-aminokwasowych połączonych w określonej kolejności w długie łańcuchy. Masa cząsteczkowa białek waha się od 5 tysięcy do 1 miliona. Nazwę „białka” po raz pierwszy nadano substancji znajdującej się w ptasich jajach, która po podgrzaniu krzepnie w białą, nierozpuszczalną masę. Termin ten został później rozszerzony na inne substancje o podobnych właściwościach izolowane od zwierząt i roślin.
Ryż. 1. Najbardziej złożonymi biopolimerami są białka. Ich makrocząsteczki składają się z monomerów, które są aminokwasami. Każdy aminokwas ma dwie grupy funkcyjne: grupę karboksylową i grupę aminową. Cała różnorodność białek powstaje w wyniku różnych kombinacji 20 aminokwasów.
Białka przeważają nad wszystkimi innymi związkami obecnymi w organizmach żywych i stanowią zwykle ponad połowę ich suchej masy. Zakłada się, że w przyrodzie istnieje kilka miliardów pojedynczych białek (przykładowo w samej bakterii E. coli występuje ponad 3 tysiące różnych białek).
Białka odgrywają kluczową rolę w procesach życiowych każdego organizmu. Do białek zalicza się enzymy, przy udziale których zachodzą w komórce wszystkie przemiany chemiczne (metabolizm); kontrolują działanie genów; przy ich udziale realizowane jest działanie hormonów, odbywa się transport przezbłonowy, w tym wytwarzanie impulsów nerwowych. Stanowią integralną część układu odpornościowego (immunoglobuliny) i układu krzepnięcia, stanowią podstawę tkanki kostnej i łącznej, biorą udział w przemianie i wykorzystaniu energii.
Historia badań nad białkami
Pierwsze próby izolacji białek podejmowano już w XVIII wieku. Na początku XIX wieku pojawiły się pierwsze prace dotyczące chemicznych badań białek. Francuscy naukowcy Joseph Louis Gay-Lussac i Louis Jacques Thénard podjęli próbę ustalenia składu pierwiastkowego białek pochodzących z różnych źródeł, co zapoczątkowało systematyczne badania analityczne, dzięki którym stwierdzono, że wszystkie białka są podobne w zestawie pierwiastków wchodzących w skład białek ich skład. W 1836 roku holenderski chemik G. J. Mulder zaproponował pierwszą teorię budowy substancji białkowych, zgodnie z którą wszystkie białka mają pewien hipotetyczny rodnik (C 40 H 62 N 10 O 12), związany w różnych proporcjach z atomami siarki i fosforu. Nazwał to radykalne „białkiem” (od greckiego białka - po pierwsze, główne). Teoria Muldera przyczyniła się do wzrostu zainteresowania badaniem białek i udoskonalenia metod chemii białek. Opracowano techniki izolacji białek metodą ekstrakcji roztworami soli obojętnych i po raz pierwszy otrzymano białka w postaci krystalicznej (niektóre białka roślinne). Do analizy białek zaczęto stosować wstępne trawienie kwasami i zasadami.
W tym samym czasie zaczęto zwracać coraz większą uwagę na badanie funkcji białek. Jens Jakob Berzelius jako pierwszy zasugerował w 1835 roku, że pełnią one rolę biokatalizatorów. Wkrótce odkryto enzymy proteolityczne – pepsynę (T. Schwann, 1836) i trypsynę (L. Corvisart, 1856), co zwróciło uwagę na fizjologię trawienia i analizę produktów powstających podczas rozkładu składników odżywczych. Dalsze badania struktury białek i prace nad chemiczną syntezą peptydów zaowocowały pojawieniem się hipotezy peptydowej, według której wszystkie białka zbudowane są z aminokwasów. Pod koniec XIX wieku zbadano większość aminokwasów tworzących białka.
Na początku XX wieku niemiecki chemik Emil Hermann Fischer jako pierwszy zastosował metody chemii organicznej do badania białek i udowodnił, że białka składają się z β-aminokwasów połączonych ze sobą wiązaniem amidowym (peptydowym). Później, dzięki zastosowaniu fizykochemicznych metod analizy, określono masę cząsteczkową wielu białek, ustalono kulisty kształt białek globularnych, przeprowadzono analizę dyfrakcji rentgenowskiej aminokwasów i peptydów oraz opracowano metody analizy chromatograficznej rozwinięty (patrz chromatografia).
Wyizolowano pierwszy hormon białkowy (Frederick Grant Banting, John James Rickard McLeod, 1922), udowodniono obecność gamma globulin w przeciwciałach i opisano enzymatyczną funkcję miozyny białka mięśniowego (Vladimir Aleksandrovich Engelhardt, M. N. Lyubimova, 1939). . Po raz pierwszy otrzymano enzymy w postaci krystalicznej - ureazę (J.B. Saliner, 1926), pepsynę (J.H. Nortron, 1929), lizozym (E.P. Abraham, Robert Robinson, 1937).

Ryż. 2. Schemat trójwymiarowej struktury enzymu lizozymu. Kręgi - aminokwasy; nici - wiązania peptydowe; zacienione prostokąty to wiązania dwusiarczkowe. Widoczne są spiralne i wydłużone odcinki łańcucha polipeptydowego.
W latach pięćdziesiątych XX wieku udowodniono trójpoziomową organizację cząsteczek białka - obecność struktury pierwszorzędowej, drugorzędowej i trzeciorzędowej; stworzył automatyczny analizator aminokwasów (Stanford Moore, William Howard Stein, 1950). W latach 60. podjęto próby chemicznej syntezy białek (insulina, rybonukleaza). Metody analizy dyfrakcji promieni rentgenowskich zostały znacznie ulepszone; stworzono urządzenie – sekwencer (P. Edman, G. Begg, 1967), które umożliwiło określenie sekwencji aminokwasów w łańcuchu polipeptydowym. Konsekwencją tego było ustalenie struktury kilkuset białek pochodzących z różnych źródeł. Należą do nich enzymy proteolityczne (pepsyna, trypsyna, chymotrypsyna, subtylizyna, karboksypeptydazy), mioglobiny, hemoglobiny, cytochromy, lizozymy, immunoglobuliny, histony, neurotoksyny, białka otoczki wirusa, hormony białkowo-peptydowe. W rezultacie pojawiły się przesłanki do rozwiązania palących problemów w enzymologii, immunologii, endokrynologii i innych obszarach chemii biologicznej.
Pod koniec XX wieku nastąpił znaczący postęp w badaniu roli białek w syntezie matrycy biopolimerów, poznaniu mechanizmów ich działania w różnych procesach życiowych organizmów oraz ustaleniu powiązań między ich strukturą i funkcją. Duże znaczenie miało doskonalenie metod badawczych i pojawienie się nowych metod rozdziału białek i peptydów.
Opracowanie skutecznej metody analizy sekwencji nukleotydów w kwasach nukleinowych umożliwiło znaczne uproszczenie i przyspieszenie wyznaczania sekwencji aminokwasów w białkach. Okazało się to możliwe, ponieważ o kolejności aminokwasów w białku decyduje kolejność nukleotydów w genie kodującym to białko (fragment). W konsekwencji, znając układ nukleotydów w tym genie oraz kod genetyczny, można dokładnie przewidzieć, w jakiej kolejności aminokwasy znajdują się w łańcuchu polipeptydowym białka. Wraz z postępem analizy strukturalnej białek osiągnięto znaczące wyniki w badaniu ich organizacji przestrzennej, mechanizmów powstawania i działania kompleksów supramolekularnych, w tym rybosomów i innych organelli komórkowych, chromatyny, wirusów itp.
Struktura białka
Prawie wszystkie białka zbudowane są z 20 α-aminokwasów należących do serii L i są takie same w prawie wszystkich organizmach. Aminokwasy w białkach są połączone ze sobą wiązaniem peptydowym -CO-NH-, które jest utworzone przez grupę karboksylową i -aminową sąsiadujących reszt aminokwasowych: dwa aminokwasy tworzą dipeptyd, w którym końcowy karboksyl (-COOH) i grupa aminowa (H2N-) pozostają wolne, do których można dodać nowe aminokwasy, tworząc łańcuch polipeptydowy.
Sekcja łańcucha, na której znajduje się końcowa grupa H2 N, nazywana jest N-końcową, a część naprzeciwko niej nazywana jest C-końcową. O ogromnej różnorodności białek decyduje kolejność ułożenia i liczba zawartych w nich reszt aminokwasowych. Chociaż nie ma jasnego rozróżnienia, krótkie łańcuchy są zwykle nazywane peptydami lub oligopeptydami (od oligo...), a polipeptydy (białka) są zwykle rozumiane jako łańcuchy składające się z 50 lub więcej. Najpopularniejsze białka to te zawierające 100-400 reszt aminokwasowych, ale są też takie, których cząsteczki składają się z 1000 i więcej reszt. Białka mogą składać się z kilku łańcuchów polipeptydowych. W takich białkach każdy łańcuch polipeptydowy nazywany jest podjednostką.
Struktura przestrzenna białek

Ryż. 3. Białko we wszystkich organizmach składa się z 20 rodzajów aminokwasów. Każde białko charakteryzuje się pewnym asortymentem i stosunkiem ilościowym aminokwasów. W cząsteczkach białka aminokwasy są połączone ze sobą wiązaniami peptydowymi (- CO - NH -) w liniowej sekwencji, stanowiącej tzw. strukturę pierwszorzędową białka. Górna linia - wolne aminokwasy z grupami bocznymi R1, R2, R3; podsumowując - aminokwasy są połączone wiązaniami peptydowymi.
Łańcuch polipeptydowy jest zdolny do samoistnego tworzenia i utrzymywania specjalnej struktury przestrzennej. Ze względu na kształt cząsteczek białek dzielimy je na włókniste i kuliste. W białkach globularnych jeden lub więcej łańcuchów polipeptydowych jest złożonych w zwartą kulistą strukturę lub kulkę. Zazwyczaj białka te są dobrze rozpuszczalne w wodzie. Należą do nich prawie wszystkie enzymy, białka transportujące krew i wiele białek magazynujących. Białka włókniste to nitkowate cząsteczki utrzymywane razem za pomocą wiązań poprzecznych i tworzące długie włókna lub struktury warstwowe. Mają wysoką wytrzymałość mechaniczną, są nierozpuszczalne w wodzie i pełnią głównie funkcje strukturalne i ochronne. Typowymi przedstawicielami takich białek są keratyny włosów i wełny, fibroina jedwabiu i kolagen ścięgien.
Kolejność kowalencyjnie połączonych aminokwasów w łańcuchu polipeptydowym nazywana jest sekwencją aminokwasów lub pierwotną strukturą białek. Podstawowa struktura każdego białka, kodowana przez odpowiedni gen, jest stała i niesie ze sobą wszystkie informacje niezbędne do tworzenia struktur wyższego poziomu. Potencjalna liczba białek, jakie można zbudować z 20 aminokwasów, jest praktycznie nieograniczona.
W wyniku oddziaływania bocznych grup reszt aminokwasowych poszczególne stosunkowo małe odcinki łańcucha polipeptydowego przyjmują taką lub inną konformację (rodzaj fałdowania), znaną jako struktura drugorzędowa białek. Jego najbardziej charakterystycznymi elementami są okresowo powtarzająca się struktura α-helisy i β. Struktura wtórna jest bardzo stabilna. Ponieważ jest to w dużej mierze zdeterminowane przez sekwencję aminokwasów odpowiedniego regionu białka, możliwe jest przewidzenie tego z pewnym stopniem prawdopodobieństwa. Termin „β-helisa” został wprowadzony przez amerykańskiego biochemika, fizyka i chemika Linusa Carla Paulinga, który opisał ułożenie łańcucha polipeptydowego w białku a-keratyny w postaci prawoskrętnej helisy (β-helisa może porównać do kabla telefonicznego). Na każdy zwój takiej helisy w białku przypada 3,6 reszt aminokwasowych. Oznacza to, że grupa -C=O jednego wiązania peptydowego tworzy wiązanie wodorowe z grupą -NH innego wiązania peptydowego, cztery reszty aminokwasowe odległe od pierwszego. Średnio każdy region α-helisy zawiera do 15 aminokwasów, co odpowiada 3-4 zwojom helisy. Ale w każdym pojedynczym białku długość helisy może znacznie różnić się od tej wartości. W przekroju poprzecznym α-helisa ma kształt dysku, z którego skierowane są na zewnątrz łańcuchy boczne aminokwasów.
Struktura, czyli? -złożona warstwa, może być utworzona przez kilka odcinków łańcucha polipeptydowego. Sekcje te są rozciągane i układane równolegle do siebie, połączone ze sobą wiązaniami wodorowymi, które występują pomiędzy wiązaniami peptydowymi. Mogą być zorientowane w tych samych lub przeciwnych kierunkach (za kierunek ruchu wzdłuż łańcucha polipeptydowego zwykle uważa się od N-końca do C-końca). W pierwszym przypadku złożoną warstwę nazywa się równoległą, w drugim - antyrównoległą. Ten ostatni powstaje, gdy łańcuch peptydowy gwałtownie zawraca, tworząc zagięcie (a-zagięcie). Czy łańcuchy boczne aminokwasów są zorientowane prostopadle do płaszczyzny? -warstwa.
Treść względna? -sekcje spiralne i? -struktury mogą się znacznie różnić pomiędzy różnymi białkami. Istnieją białka z przewagą α-helis (około 75% aminokwasów w mioglobinie i hemoglobinie), a głównym rodzajem fałdowania łańcucha w wielu białkach fibrylarnych (m.in. fibroinie jedwabiu, β-keratynie) jest α-helisa. -Struktura. Regiony łańcucha polipeptydowego, których nie można zaklasyfikować do żadnej z opisanych powyżej konformacji, nazywane są pętlami łączącymi. O ich strukturze decydują głównie interakcje pomiędzy łańcuchami bocznymi aminokwasów, a w cząsteczce dowolnego białka mieści się ono w ściśle określony sposób.
Struktura trzeciorzędowa nazywa się struktura przestrzenna białek globularnych. Ale często koncepcja ta odnosi się do metody składania łańcucha polipeptydowego w przestrzeni, charakterystycznej dla każdego konkretnego białka. Struktura trzeciorzędowa jest tworzona przez łańcuch polipeptydowy białka spontanicznie, najwyraźniej wzdłuż określonej ścieżki krzepnięcia ze wstępnym utworzeniem elementów struktury drugorzędowej. Jeśli stabilność struktury drugorzędowej wynika z wiązań wodorowych, wówczas strukturę trzeciorzędową ustala zróżnicowany układ oddziaływań niekowalencyjnych: oddziaływania wodorowe, jonowe, międzycząsteczkowe, a także kontakty hydrofobowe między łańcuchami bocznymi niepolarnych grup aminowych pozostałości kwasu.
W niektórych białkach struktura trzeciorzędowa jest dodatkowo stabilizowana przez tworzenie wiązań dwusiarczkowych (wiązania -S-S-) pomiędzy resztami cysteiny. Z reguły wewnątrz globuli białkowej znajdują się łańcuchy boczne aminokwasów hydrofobowych złożone w rdzeń (ich transport do globuli białkowej jest korzystny termodynamicznie), a na obrzeżach znajdują się reszty hydrofilowe i część hydrofobowych. Globula białkowa otoczona jest kilkomaset cząsteczkami wody hydratacyjnej, która jest niezbędna dla stabilności cząsteczki białka i często bierze udział w jej funkcjonowaniu. Struktura trzeciorzędowa jest ruchoma, jej poszczególne sekcje mogą się przesuwać, co prowadzi do przejść konformacyjnych, które odgrywają znaczącą rolę w interakcji białka z innymi cząsteczkami.
Struktura trzeciorzędowa jest podstawą właściwości funkcjonalnych białka. Decyduje o tworzeniu się zespołów grup funkcyjnych w białku – centrach aktywnych i strefach wiązania, nadaje im niezbędną geometrię, pozwala na utworzenie środowiska wewnętrznego, które jest warunkiem zajścia wielu reakcji oraz zapewnia interakcję z innymi białkami .
Trzeciorzędowa struktura białek wyraźnie odpowiada ich strukturze pierwotnej; prawdopodobnie istnieje jeszcze nierozszyfrowany kod stereochemiczny, który określa naturę zwijania białek. Jednak ten sam sposób rozmieszczenia przestrzennego zwykle nie odpowiada pojedynczej strukturze pierwotnej, ale całej rodzinie struktur, w których tylko niewielka część (do 20-30%) reszt aminokwasowych może się pokrywać, ale w pewnych miejscach w łańcuchu zachowane jest podobieństwo reszt aminokwasowych. W rezultacie powstają duże rodziny białek charakteryzujące się podobną trzeciorzędową i mniej więcej podobną strukturą pierwszorzędową i z reguły wspólną funkcją. Są to na przykład białka organizmów różnych gatunków, które pełnią tę samą funkcję i są ze sobą powiązane ewolucyjnie: mioglobiny i hemoglobiny, trypsyna, chymotrypsyna, elastaza i inne proteinazy zwierzęce.

Ryż. 4. W wyniku połączenia kilku makrocząsteczek białkowych o strukturze trzeciorzędowej powstaje czwartorzędowa struktura białkowa w złożony kompleks. Przykładem takich złożonych białek jest hemoglobina, składająca się z czterech makrocząsteczek.
Często, zwłaszcza w dużych białkach, fałdowanie łańcucha polipeptydowego następuje poprzez tworzenie przez poszczególne odcinki łańcucha mniej lub bardziej autonomicznych elementów struktury przestrzennej - domen, które mogą mieć autonomię funkcjonalną, odpowiadając za tę lub inną aktywność biologiczną białka. białko. Zatem domeny N-końcowe białek krzepnięcia krwi zapewniają ich przyłączenie do błony komórkowej.
Istnieje wiele białek, których cząsteczki stanowią zespół globul (podjednostek) połączonych ze sobą oddziaływaniami hydrofobowymi, wiązaniami wodorowymi lub jonowymi. Takie kompleksy nazywane są białkami oligomerycznymi, multimerycznymi lub podjednostkowymi. Układ podjednostek w funkcjonalnie aktywnym kompleksie białkowym nazywany jest czwartorzędową strukturą białka. Niektóre białka są zdolne do tworzenia struktur wyższego rzędu, na przykład kompleksy wieloenzymowe, struktury rozciągnięte (białka płaszcza bakteriofaga), kompleksy supramolekularne, które funkcjonują jako pojedyncza całość (na przykład rybosomy lub składniki mitochondrialnego łańcucha oddechowego).
Struktura czwartorzędowa umożliwia tworzenie cząsteczek o nietypowych geometriach. Zatem ferrytyna zbudowana z 24 podjednostek posiada wewnętrzną wnękę, dzięki której białko jest w stanie związać aż 3000 jonów żelaza. Ponadto struktura czwartorzędowa pozwala na pełnienie kilku różnych funkcji w jednej cząsteczce. Syntetaza tryptofanu łączy w sobie enzymy odpowiedzialne za kilka kolejnych etapów syntezy aminokwasu tryptofanu.
Metody badania struktury białek
Podstawowa struktura białek determinuje wszystkie pozostałe poziomy organizacji cząsteczki białka. Dlatego przy badaniu funkcji biologicznych różnych białek ważna jest znajomość tej struktury. Pierwszym białkiem, dla którego ustalono sekwencję aminokwasów, był hormon trzustki – insulina. Prace te, które trwały 11 lat, przeprowadził angielski biochemik Frederick Sanger (1954). Określił lokalizację 51 aminokwasów w cząsteczce hormonu i wykazał, że składa się ona z 2 łańcuchów połączonych wiązaniami dwusiarczkowymi. Później większość prac nad ustaleniem pierwotnej struktury białek została zautomatyzowana.
Wraz z rozwojem metod inżynierii genetycznej możliwe stało się dalsze przyspieszenie tego procesu poprzez określenie struktury pierwszorzędowej białek zgodnie z wynikami analizy sekwencji nukleotydowej w genach kodujących te białka. Do badania struktury drugorzędowej i trzeciorzędowej białek stosuje się dość złożone metody fizyczne, na przykład dichroizm kołowy lub analizę dyfrakcji promieni rentgenowskich kryształów białek. Struktura trzeciorzędowa została po raz pierwszy ustalona przez angielskiego biochemika Johna Cowdery'ego Kendrew (1957) dla mioglobiny białka mięśniowego.

Ryż. 5. Model cząsteczki mioglobiny (konfiguracja przestrzenna cząsteczki)
Denaturacja białek
Stosunkowo słabe wiązania odpowiedzialne za stabilizację drugorzędowych, trzeciorzędowych i czwartorzędowych struktur białka łatwo ulegają zniszczeniu, czemu towarzyszy utrata jego aktywności biologicznej. Zniszczenie pierwotnej (natywnej) struktury białka, zwane denaturacją, następuje w obecności kwasów i zasad, podczas ogrzewania, zmian siły jonowej i innych wpływów. Z reguły zdenaturowane białka są słabo lub wcale nierozpuszczalne w wodzie. Dzięki krótkotrwałemu działaniu i szybkiej eliminacji czynników denaturujących możliwa jest renaturacja białek z całkowitym lub częściowym przywróceniem pierwotnej struktury i właściwości biologicznych.
Klasyfikacja białek
Złożoność budowy cząsteczek białek oraz ogromna różnorodność funkcji, jakie pełnią, powodują, że trudno jest stworzyć ich jednolitą i jednoznaczną klasyfikację, choć próby tego podejmowano wielokrotnie od końca XIX wieku. Ze względu na skład chemiczny białka dzielimy na proste i złożone (czasami nazywane proteidami). Cząsteczki tego pierwszego składają się wyłącznie z aminokwasów. Oprócz samego łańcucha polipeptydowego złożone białka zawierają składniki niebiałkowe reprezentowane przez węglowodany (glikoproteiny), lipidy (lipoproteiny), kwasy nukleinowe (nukleoproteiny), jony metali (metaloproteiny), grupy fosforanowe (fosfoproteiny), pigmenty (chromoproteiny), itp. .
W zależności od pełnionych funkcji wyróżnia się kilka klas białek. Najbardziej zróżnicowaną i najbardziej wyspecjalizowaną klasą są białka o funkcji katalitycznej – enzymy, które mają zdolność przyspieszania reakcji chemicznych zachodzących w organizmach żywych. W tej roli białka uczestniczą we wszystkich procesach syntezy i rozkładu różnych związków podczas metabolizmu, w biosyntezie białek i kwasów nukleinowych, regulacji rozwoju i różnicowania komórek. Białka transportowe mają zdolność selektywnego wiązania kwasów tłuszczowych, hormonów i innych związków organicznych i nieorganicznych oraz jonów, a następnie transportu ich prądem w wybrane miejsce (przykładowo hemoglobina bierze udział w przenoszeniu tlenu z płuc do wszystkich komórek organizmu Ciało). Białka transportowe realizują także aktywny transport jonów, lipidów, cukrów i aminokwasów przez błony biologiczne.
Białka strukturalne pełnią funkcję wspierającą lub ochronną; biorą udział w tworzeniu szkieletu komórkowego. Najpopularniejsze z nich to kolagen tkanki łącznej, keratyna, paznokcie i pióra, elastyna komórek naczyniowych i wiele innych. W połączeniu z lipidami stanowią podstawę strukturalną błon komórkowych i wewnątrzkomórkowych.
Szereg białek pełni funkcję ochronną. Na przykład immunoglobuliny (przeciwciała) kręgowców, mające zdolność wiązania obcych patogennych mikroorganizmów i substancji, neutralizują ich patogenne działanie na organizm i zapobiegają proliferacji komórek. Fibrynogen i trombina biorą udział w procesie krzepnięcia krwi. Wiele substancji białkowych wydzielanych przez bakterie, a także składniki niektórych bezkręgowców, zalicza się do toksyn.
Niektóre białka (regulacyjne) biorą udział w regulacji aktywności fizjologicznej organizmu jako całości, poszczególnych narządów, komórek lub procesów. Kontrolują transkrypcję genów i syntezę białek; obejmują one hormony peptydowo-białkowe wydzielane przez gruczoły dokrewne. Białka zapasowe nasion dostarczają składników odżywczych na początkowe etapy rozwoju zarodka. Należą do nich także kazeina, albumina białka jaja (albumina jaja kurzego) i wiele innych. Dzięki białkom komórki mięśniowe nabywają zdolność do kurczenia się i ostatecznie zapewnienia ruchu organizmowi. Przykładami takich białek kurczliwych są aktyna i miozyna mięśni szkieletowych, a także tubulina, które są składnikami rzęsek i wici organizmów jednokomórkowych; Zapewniają także rozbieżność chromosomów podczas podziału komórki.
Białka receptorowe są celem hormonów i innych związków biologicznie aktywnych. Za ich pomocą komórka odbiera informacje o stanie środowiska zewnętrznego. Odgrywają ważną rolę w przekazywaniu pobudzenia nerwowego i ukierunkowanym ruchu komórek (chemotaksja). Przekształcenie i wykorzystanie energii docierającej do organizmu, a także energii, następuje również przy udziale białek układu bioenergetycznego (na przykład barwnika wizualnego rodopsyny, cytochromów łańcucha oddechowego). Istnieje również wiele białek o innych, czasem dość nietypowych funkcjach (na przykład osocze niektórych ryb antarktycznych zawiera białka o właściwościach przeciw zamarzaniu).
Biosynteza białek
Cała informacja o budowie konkretnego białka jest „przechowywana” w odpowiednich genach w postaci sekwencji nukleotydów i wykorzystywana w procesie syntezy matrycy. Najpierw informacja jest przekazywana (odczytwana) z cząsteczki DNA do informacyjnego RNA (mRNA) za pomocą enzymu polimerazy RNA zależnej od DNA, a następnie w rybosomie na mRNA, jak na matrycy zgodnej z kodem genetycznym, przy udziale transportowych RNA dostarczających aminokwasy powstaje łańcuch polipeptydowy.
Zsyntetyzowane łańcuchy polipeptydowe wyłaniające się z rybosomu, samoistnie zwijające się, przyjmują konformację charakterystyczną dla białka i mogą podlegać modyfikacji potranslacyjnej. Łańcuchy boczne poszczególnych aminokwasów mogą ulegać modyfikacjom (hydroksylacja, fosforylacja itp.). Dlatego w kolagenie znajdują się na przykład hydroksyprolina i hydroksylizyna (patrz). Modyfikacji może towarzyszyć także zerwanie wiązań polipeptydowych. W ten sposób dochodzi np. do powstania aktywnej cząsteczki insuliny, składającej się z dwóch łańcuchów połączonych wiązaniami dwusiarczkowymi.

Ryż. 6. Ogólny schemat biosyntezy białek.
Znaczenie białek w żywieniu
Białka są najważniejszym składnikiem pożywienia zwierząt i ludzi. Wartość odżywcza białek zależy od zawartości niezbędnych aminokwasów, które nie są wytwarzane w samym organizmie. Pod tym względem białka roślinne są mniej wartościowe niż białka zwierzęce: są uboższe w lizynę, metioninę i tryptofan oraz są trudniej trawione w przewodzie pokarmowym. Brak niezbędnych aminokwasów w pożywieniu prowadzi do poważnych zaburzeń metabolizmu azotu.
Białka rozkładają się na wolne aminokwasy, które po wchłonięciu w jelicie przedostają się i są rozprowadzane do wszystkich komórek. Część z nich rozkłada się na proste związki z wyzwoleniem energii wykorzystywanej przez komórkę na różne potrzeby, a część idzie do syntezy nowych białek charakterystycznych dla danego organizmu. (RA Matveeva, Encyklopedia Cyryl i Metody)
Liczenie białek
- amyloid - amyloid;
- anionowy - anionowy;
- program antywirusowy - antywirusowy;
- autoimmunologiczny - autoimmunologiczny;
- autologiczny - autologiczny;
- bakteryjny - bakteryjny;
- białko Bence'a Jonesa;
- wywołane wirusem - wywołane wirusem;
- wirusowy - wirus;
- wirusowy niestrukturalny - wirusowy niestrukturalny;
- struktura wirusa - struktura wirusa;
- specyficzny dla wirusa - specyficzny dla wirusa;
- wysoka masa cząsteczkowa - wysoka masa cząsteczkowa;
- zawierający hem - hem;
- heterologiczny - obcy;
- hybrydowy - hybrydowy;
- glikozylowany - glikowany;
- kulisty - kulisty;
- zdenaturowany - zdenaturowany;
- zawierający żelazo - żelazo;
- żółtko - żółtko;
- białko zwierzęce – białko zwierzęce;
- ochronny - defensywny;
- odporny - odporny;
- immunogenny - istotny immunologicznie;
- wiązanie wapnia;
- kwaśny - kwaśny;
- korpuskularny - korpuskularny;
- membrana - membrana;
- szpiczak - szpiczak;
- mikrosomalny - mikrosomalny;
- białko mleka - białko mleka;
- monoklonalny - immunoglobulina monoklonalna;
- białko mięśniowe - białko mięśniowe;
- rodzimy - rodzimy;
- nonhiston - niehiston;
- uszkodzony - częściowy;
- nierozpuszczalny - nierozpuszczalny;
- niestrawny - nierozpuszczalny;
- nieenzymatyczny - nieenzym;
- niska masa cząsteczkowa - niska masa cząsteczkowa;
- nowe białko - nowe białko;
- ogólne - całe;
- onkogenny - onkoproteina;
- białko fazy głównej – anionowe;
- białko ostrej fazy (zapalenia) - białko ostrej fazy;
- jedzenie jedzenie;
- białko osocza krwi - białko osocza;
- łożysko - łożysko;
- rozprzęganie - rozprzęganie;
- białko regenerującego się nerwu;
- regulacyjny - regulacyjny;
- rekombinacja - rekombinowana;
- receptor - receptor;
- rybosomalny - rybosomalny;
- wiązanie - wiązanie;
- białko wydzielnicze - białko wydzielnicze;
- C-reaktywny - C-reaktywny;
- białko serwatkowe – białko serwatkowe, laktoproteina;
- tkanka - tkanka;
- toksyczny - toksyczny;
- chimeryczny - chimeryczny;
- cały - cały;
- cytozolowy - cytozolowy;
- białko alkaliczne – białko anionowe;
- egzogenny - egzogenny;
- endogenne - białko endogenne.
Przeczytaj więcej na temat białek w literaturze:
- Volkenshtein M.V., Molecules and, M., 1965, rozdz. 3 - 5;
- Gaurowitz F., Chemia i funkcje białek, przeł. z języka angielskiego, Moskwa, 1965;
- Sissakyan N. M. i Gladilin K. L., Biochemiczne aspekty syntezy białek, w książce: Postępy w chemii biologicznej, t. 7, M., 1965, s. 10-10. 3;
- Stepanov V. M. Biologia molekularna. Struktura i funkcja białek. M., 1996;
- Shamin A. N., Rozwój chemii białek, M., 1966;
- Białka i peptydy. M., 1995-2000. T. 1-3;
- Biosynteza białek i kwasów nukleinowych, wyd. AS Spirina, M., 1965;
- Wprowadzenie do biologii molekularnej, przeł. z języka angielskiego, M., 1967
- Cząsteczki i komórki. [sobota Art.], przeł. z jęz. angielskiego, M., 1966, s. 25. 7 - 27, 94 - 106;
- Podstawy biochemii: Tłumaczenie z języka angielskiego M., 1981. T. 1;
- Problem białka. M., 1995. T. 1-5;
- Białka. Nowy Jork, 1975–79. 3 wyd. V. 1-4.
Znajdź coś innego interesującego:







