Komórki mięśniowe i gruczołowe są przenoszone przez specjalną formację strukturalną - synapsę.
Synapsa- konstrukcja zapewniająca przewodzenie sygnału od jednego do drugiego. Termin został wprowadzony przez angielskiego fizjologa C. Sherringtona w 1897 roku.
Struktura synapsy
Synapsy składają się z trzech głównych elementów: błony presynaptycznej, błony postsynaptycznej i szczeliny synaptycznej (ryc. 1).
Ryż. 1. Struktura synapsy: 1 - mikrotubule; 2 - mitochondria; 3 - pęcherzyki synaptyczne z nadajnikiem; 4 - błona presynaptyczna; 5 - błona postsynaptyczna; 6 - receptory; 7 - szczelina synaptyczna
Niektóre elementy synaps mogą mieć inne nazwy. Na przykład płytka synaptyczna to synapsa pomiędzy, płytka końcowa to błona postsynaptyczna, płytka motoryczna to presynaptyczne zakończenie aksonu na włóknie mięśniowym.

Błona presynaptyczna pokrywa rozszerzone zakończenie nerwowe, które jest aparatem neurosekrecyjnym. Część presynaptyczna zawiera pęcherzyki i mitochondria, które zapewniają syntezę mediatorów. Mediatory osadzają się w postaci granulek (pęcherzyków).
Błona postsynaptyczna - pogrubiona część błony komórkowej, z którą styka się błona presynaptyczna. Posiada kanały jonowe i jest w stanie generować potencjały czynnościowe. Ponadto istnieją specjalne struktury białkowe- receptory odbierające działanie mediatorów.
Szczelina synaptyczna to przestrzeń pomiędzy błoną presynaptyczną i postsynaptyczną, wypełniona cieczą o składzie podobnym do.

Ryż. Budowa synapsy i procesy zachodzące podczas transmisji sygnału synaptycznego
Rodzaje synaps
Synapsy są klasyfikowane według lokalizacji, charakteru działania i sposobu przekazywania sygnału.
Według lokalizacji Wyróżniają synapsy nerwowo-mięśniowe, neurogruczołowe i neuronalne; te ostatnie z kolei dzielą się na aksoaksonalne, akso-dendrytyczne, akso-somatyczne, dendrosomatyczne, dendro-dendrotyczne.
Ze względu na charakter akcji Synapsy na strukturze percepcyjnej mogą być pobudzające lub hamujące.
Według metody transmisji sygnału Synapsy dzielą się na elektryczne, chemiczne i mieszane.
Tabela 1. Klasyfikacja i rodzaje synaps

Klasyfikacja synaps i mechanizm przekazywania pobudzenia
Synapsy dzieli się na:
- według lokalizacji - peryferyjne i centralne;
- z natury ich działania - ekscytujące i hamujące;
- metodą transmisji sygnału - chemiczną, elektryczną, mieszaną;
- według mediatora, przez który odbywa się transmisja - cholinergicznego, adrenergicznego, serotoninergicznego itp.
Podniecenie jest przekazywane przez mediatorzy(pośrednicy).
Mediatorzy- Cząsteczki substancje chemiczne, które zapewniają przekazywanie wzbudzenia w synapsach. Innymi słowy, substancje chemiczne biorące udział w przenoszeniu wzbudzenia lub hamowania z jednej komórki pobudliwej do drugiej.
Właściwości mediatorów
- Syntetyzowany w neuronie
- Gromadzi się na końcu komórki
- Uwalniany, gdy jon Ca2+ pojawia się w terminalu presynaptycznym
- Mają specyficzny wpływ na błonę postsynaptyczną
Przez struktura chemiczna Mediatory można podzielić na aminy (noradrenalina, dopamina, serotonina), aminokwasy (glicyna, kwas gamma-aminomasłowy) i polipeptydy (endorfiny, enkefaliny). Acetylocholina znana jest głównie jako neuroprzekaźnik pobudzający i występuje w różnych częściach ośrodkowego układu nerwowego. Nadajnik znajduje się w pęcherzykach zgrubienia presynaptycznego (płytki synaptycznej). Mediator jest syntetyzowany w komórkach neuronowych i może być resyntetyzowany z metabolitów powstałych w wyniku jego rozszczepienia w szczelinie synaptycznej.
Kiedy zakończenia aksonów są wzbudzone, błona płytki synaptycznej ulega depolaryzacji, powodując przepływ jonów wapnia ze środowiska zewnątrzkomórkowego do zakończenia nerwowego poprzez kanały wapniowe. Jony wapnia stymulują ruch pęcherzyków synaptycznych do błony presynaptycznej, ich fuzję z nią i późniejsze uwolnienie przekaźnika do błony presynaptycznej szczelina synaptyczna. Po wniknięciu w szczelinę nadajnik dyfunduje do błony postsynaptycznej zawierającej na jej powierzchni receptory. Oddziaływanie nadajnika z receptorami powoduje otwarcie kanałów sodowych, co przyczynia się do depolaryzacji błony postsynaptycznej i pojawienia się pobudzającego potencjału postsynaptycznego. W synapsie nerwowo-mięśniowej potencjał ten nazywa się potencjał płyty końcowej. Pomiędzy depolaryzowaną błoną postsynaptyczną a sąsiadującymi spolaryzowanymi odcinkami tej samej błony powstają lokalne prądy, które depolaryzują błonę do poziomu krytycznego, po czym następuje wygenerowanie potencjału czynnościowego. Potencjał czynnościowy rozprzestrzenia się po wszystkich błonach np. włókna mięśniowego i powoduje jego skurcz.
Nadajnik uwolniony do szczeliny synaptycznej wiąże się z receptorami błony postsynaptycznej i jest rozszczepiany przez odpowiedni enzym. W ten sposób cholinoesteraza niszczy neuroprzekaźnik acetylocholinę. Następnie pewna ilość produktów rozpadu mediatorów dostaje się do płytki synaptycznej, gdzie ponownie syntetyzuje się z nich acetylocholinę.
Ciało zawiera nie tylko synapsy pobudzające, ale także hamujące. Zgodnie z mechanizmem transmisji pobudzenia są one podobne do synaps pobudzających. W synapsach hamujących przekaźnik (na przykład kwas gamma-aminomasłowy) wiąże się z receptorami na błonie postsynaptycznej i sprzyja w niej otwarciu. W tym przypadku aktywowane jest przenikanie tych jonów do wnętrza komórki i rozwija się hiperpolaryzacja błony postsynaptycznej, powodując pojawienie się hamującego potencjału postsynaptycznego.
Obecnie odkryto, że jeden mediator może wiązać się z kilkoma różnymi receptorami i wywoływać różne reakcje.
Synapsy chemiczne
Właściwości fizjologiczne synaps chemicznych
Synapsy z chemiczną transmisją wzbudzenia mają pewne właściwości:
- wzbudzenie odbywa się w jednym kierunku, ponieważ nadajnik jest uwalniany tylko z płytki synaptycznej i oddziałuje z receptorami na błonie postsynaptycznej;
- rozprzestrzenianie się wzbudzenia przez synapsy następuje wolniej niż wzdłuż włókna nerwowego (opóźnienie synaptyczne);
- przekazywanie wzbudzenia odbywa się za pomocą określonych mediatorów;
- rytm zmian wzbudzenia w synapsach;
- synapsy mogą się zmęczyć;
- synapsy są bardzo wrażliwe na różne substancje chemiczne i niedotlenienie.
Jednokierunkowa transmisja sygnału. Sygnał przekazywany jest wyłącznie z błony presynaptycznej do błony postsynaptycznej. Wynika to z cech strukturalnych i właściwości struktur synaptycznych.
Powolna transmisja sygnału. Spowodowane opóźnieniem synaptycznym w przekazywaniu sygnału z jednej komórki do drugiej. Opóźnienie wynika z czasu potrzebnego na procesy uwolnienia przekaźnika, jego dyfuzji do błony postsynaptycznej, związania się z receptorami błony postsynaptycznej, depolaryzacji i konwersji potencjału postsynaptycznego na AP (potencjał czynnościowy). Czas trwania opóźnienia synaptycznego wynosi od 0,5 do 2 ms.
Umiejętność podsumowania wpływu sygnałów docierających do synapsy. Sumowanie to pojawia się, jeśli kolejny sygnał dotrze do synapsy w krótkim czasie (1-10 ms) po poprzednim. W takich przypadkach amplituda EPSP wzrasta i na neuronie postsynaptycznym można wygenerować wyższą częstotliwość AP.
Transformacja rytmu podniecenia. Częstotliwość impulsów nerwowych docierających do błony presynaptycznej zwykle nie odpowiada częstotliwości AP generowanych przez neuron postsynaptyczny. Wyjątkiem są synapsy, które przekazują wzbudzenie z włókna nerwowego do mięśnia szkieletowego.
Niska labilność i duże zmęczenie synaps. Synapsy mogą przewodzić 50-100 impulsów nerwowych na sekundę. Jest to 5–10 razy mniej niż maksymalna częstotliwość AP, jaką mogą odtwarzać włókna nerwowe pod wpływem stymulacji elektrycznej. Jeśli włókna nerwowe uważa się za praktycznie niestrudzone, wówczas zmęczenie synaps rozwija się bardzo szybko. Dzieje się tak z powodu wyczerpania rezerw nadajnika, zasobów energii, rozwoju trwałej depolaryzacji błony postsynaptycznej itp.
Wysoka wrażliwość synaps na działanie substancji biologicznie czynnych, leków i trucizn. Na przykład trucizna strychnina blokuje działanie synaps hamujących w ośrodkowym układzie nerwowym, wiążąc się z receptorami wrażliwymi na mediator glicynę. Toksyna tężcowa blokuje synapsy hamujące, zakłócając uwalnianie przekaźnika z zakończenia presynaptycznego. W obu przypadkach rozwijają się zjawiska zagrażające życiu. Przykłady wpływu substancji biologicznie czynnych i trucizn na przekazywanie sygnału w synapsach nerwowo-mięśniowych omówiono powyżej.
Właściwości ułatwiające i depresyjne transmisji synoptycznej. Ułatwienie transmisji synaptycznej następuje wtedy, gdy impulsy nerwowe docierają do synapsy po krótkim czasie (10-50 ms) jeden po drugim, tj. wystarczająco często. Ponadto, w pewnym okresie czasu, każde kolejne PD docierające do błony presynaptycznej powoduje wzrost zawartości transmitera w szczelinie synaptycznej, wzrost amplitudy EPSP i wzrost efektywności transmisji synaptycznej.
Jednym z mechanizmów ułatwiających jest akumulacja jonów Ca 2 w terminalu presynaptycznym. Pompa wapniowa potrzebuje kilkudziesięciu milisekund, aby usunąć część wapnia, która dostała się do zakończenia synaptycznego po przybyciu AP. Jeśli w tym czasie pojawi się nowy potencjał czynnościowy, wówczas nowa porcja wapnia dostanie się do terminala, a jej wpływ na uwalnianie neuroprzekaźnika zostanie dodany do pozostałej ilości wapnia, której pompa wapniowa nie zdążyła usunąć z neuroplazmy terminal.
Istnieją inne mechanizmy rozwoju ulgi. Zjawisko to nazywane jest także w klasycznych podręcznikach fizjologii wzmocnienie po tężcowe. Ułatwienie transmisji synaptycznej jest ważne w funkcjonowaniu mechanizmów pamięciowych, dla ich formowania odruchy warunkowe i trening. Ułatwienie przekazywania sygnału leży u podstaw rozwoju plastyczności synaptycznej i poprawy ich funkcji przy częstej aktywacji.
Depresja (hamowanie) przekazywania sygnału w synapsach rozwija się, gdy bardzo częste (w przypadku synapsy nerwowo-mięśniowej powyżej 100 Hz) impulsy nerwowe docierają do błony presynaptycznej. W mechanizmach rozwoju zjawiska depresji, wyczerpaniu się rezerw nadajnika w terminalu presynaptycznym, zmniejszeniu wrażliwości receptorów błony postsynaptycznej na przekaźnik i rozwoju trwałej depolaryzacji błony postsynaptycznej, co komplikuje wytwarzanie AP na błonie komórki postsynaptycznej są ważne.
Synapsy elektryczne
Oprócz synaps z chemicznym przekazywaniem wzbudzenia, w organizmie znajdują się synapsy z przekazywaniem elektrycznym. Synapsy te mają bardzo wąską szczelinę synaptyczną i są zmniejszone opór elektryczny pomiędzy dwiema membranami. Ze względu na obecność kanałów poprzecznych pomiędzy membranami i niski opór, impuls elektryczny z łatwością przechodzi przez membrany. Synapsy elektryczne są zwykle charakterystyczne dla komórek tego samego typu.
W wyniku ekspozycji na bodziec presynaptyczny potencjał czynnościowy pobudza błonę postsynaptyczną, gdzie następuje propagujący potencjał czynnościowy.
Charakteryzują się większą szybkością wzbudzenia w porównaniu do synaps chemicznych i małą wrażliwością na działanie środków chemicznych.
Synapsy elektryczne mają jedno- i dwukierunkową transmisję wzbudzenia.
Elektryczne synapsy hamujące znajdują się również w organizmie. Efekt hamujący rozwija się w wyniku działania prądu, który powoduje hiperpolaryzację błony postsynaptycznej.
W synapsach mieszanych wzbudzenie może być przekazywane zarówno za pomocą impulsów elektrycznych, jak i mediatorów.
Moskwa PsychologicznaInstytut Społeczny (MSSI)
Streszczenie anatomii centralnego układu nerwowego na temat:
SYNAPSY(struktura, struktura, funkcje).
studentka I roku Wydziału Psychologii,
grupa 21/1-01 Logachev A.Yu.
Nauczyciel:
Marina Chołodowa Władimirowna.
rok 2001.
Plan pracy:
1.Prolog.
2. Fizjologia neuronu i jego budowa.
3.Budowa i funkcje synapsy.
4.Synapsa chemiczna.
5. Izolacja mediatora.
6. Mediatory chemiczne i ich rodzaje.
7.Epilog.
8. Lista referencji.
PROLOG:
Nasze ciało to jeden wielki mechanizm zegarowy. Składa się z ogromnej liczby drobnych cząstek, które znajdują się w w ścisłym porządku a każdy z nich pełni określone funkcje i ma swoje własne unikalne właściwości. Ten mechanizm - ciało, składa się z komórek, łączących ich tkanki i układy: wszystko to jako całość stanowi pojedynczy łańcuch, supersystem ciała. Największa różnorodność elementów komórkowych nie mogłaby działać jako pojedyncza całość, gdyby w organizmie nie istniał wyrafinowany mechanizm regulacyjny. Układ nerwowy odgrywa szczególną rolę w regulacji. Cała ciężka praca system nerwowy- regulacja pracy narządy wewnętrzne, kontrola ruchów, czy to prostych i nieświadomych ruchów (na przykład oddychania), czy złożonych ruchów rąk danej osoby - wszystko to w istocie opiera się na interakcji komórek ze sobą. Wszystko to zasadniczo opiera się na transmisji sygnału z jednej komórki do drugiej. Co więcej, każda komórka wykonuje swoją pracę, a czasami ma kilka funkcji. Różnorodność funkcji wynika z dwóch czynników: sposobu, w jaki komórki są ze sobą połączone oraz sposobu rozmieszczenia tych połączeń.
FIZJOLOGIA NEURONA I JEGO STRUKTURA:
Najprostsza reakcja układ nerwowy na bodziec zewnętrzny - to odruch. Przede wszystkim rozważmy strukturę i fizjologię elementarnej jednostki strukturalnej Tkanka nerwowa zwierzęta i ludzie - neuronu. Funkcjonalne i podstawowe właściwości neuronu są określone przez jego zdolność do wzbudzania i samopobudzania. Przekazywanie wzbudzenia odbywa się wzdłuż procesów neuronu - aksony i dendryty.
Aksony to dłuższe i szersze procesy. Mają szereg specyficznych właściwości: izolowane przewodzenie wzbudzenie i przewodzenie obustronne.
Komórki nerwowe potrafią nie tylko odbierać i przetwarzać stymulację zewnętrzną, ale także samoistnie wytwarzać impulsy, które nie są spowodowane stymulacją zewnętrzną (samopobudzenie). W odpowiedzi na stymulację neuron reaguje impuls do działania- potencjał czynnościowy, którego częstotliwość generowania waha się od 50-60 impulsów na sekundę (dla neuronów ruchowych) do 600-800 impulsów na sekundę (dla interneuronów mózgu). Akson kończy się wieloma cienkimi gałęziami, tzw terminale. Z końcówek impuls przechodzi do innych komórek, bezpośrednio do ich ciał lub częściej do ich procesów dendrytycznych. Liczba zakończeń w aksonie może sięgać nawet tysiąca i kończą się różnymi komórkami. Z drugiej strony typowy neuron kręgowców ma od 1000 do 10 000 zakończeń z innych komórek.
Dendryty - krótsze i liczniejsze procesyneurony. Odczuwają wzbudzenie z sąsiednich neuronów i kierują je do ciała komórki. Istnieją papkowate i niemiazgowe komórki nerwowe i włókna.
Włókna celulozowe są częścią wrażliwych inerwy ruchowe mięśni szkieletowych i narządy zmysłówSą pokryte lipidową osłonką mielinową. Włókna celulozowe są bardziej „szybkie”: w takich włóknach o średnicy 1-3,5 mikromilimetra wzbudzenie rozprzestrzenia się z prędkością 3-18 m/s. Wyjaśnia to fakt, że przewodzenie impulsów wzdłuż mielinowanego nerwu zachodzi spazmatycznie. W tym przypadku potencjał czynnościowy „przeskakuje” przez obszar nerwu pokryty mieliną i w węźle Ranviera (odsłonięty obszar nerwu) przechodzi do osłony osiowego cylindra nerwu błonnik. Osłonka mielinowa jest dobrym izolatorem i zapobiega przenoszeniu wzbudzenia na połączenie równoległych włókien nerwowych.
Włókna inne niż mięśniowe stanowią większość nerwów współczulnych. Nie mają osłonki mielinowej i są oddzielone od siebie komórkami neuroglejowymi.
We włóknach bezmiazgowych komórki pełnią rolę izolatorów. neuroglej(nerwowa tkanka podporowa). Komórki Schwanna - jeden z typów komórek glejowych. Oprócz neuronów wewnętrznych, które odbierają i przetwarzają impulsy pochodzące od innych neuronów, istnieją neurony, które postrzegają wpływy bezpośrednio od innych neuronów. środowisko- Ten receptory, a także neurony, które bezpośrednio wpływają na narządy wykonawcze - efektory, na przykład na mięśniach lub gruczołach. Jeśli neuron działa na mięsień, nazywa się go neuronem ruchowym lub neuron ruchowy. Wśród neuroreceptorów wyróżnia się 5 typów komórek, w zależności od rodzaju patogenu:
- fotoreceptory, które wzbudzają się pod wpływem światła i zapewniają funkcjonowanie narządów wzroku,
- mechanoreceptory, te receptory, które reagują na wpływy mechaniczne. Znajdują się w narządach słuchu i równowagi. Komórki dotykowe są również mechanoreceptorami. Niektóre mechanoreceptory znajdują się w mięśniach i mierzą stopień ich rozciągnięcia.
- chemoreceptory - selektywnie reagują na obecność lub zmianę stężenia różnych substancji chemicznych, na których opiera się praca narządów węchu i smaku,
- termoreceptory, reagują na zmiany temperatury lub jej poziomu – receptory zimna i ciepła,
- elektroreceptory reagują na impulsy prądu i występują u niektórych ryb, płazów i ssaków, na przykład dziobaka.
Na podstawie powyższego pragnę zauważyć, że wśród biologów zajmujących się badaniem układu nerwowego przez długi czas panowała opinia, że komórki nerwowe tworzą długie, złożone sieci, które w sposób ciągły przekształcają się w siebie.
Jednak w 1875 roku włoski naukowiec, profesor histologii na Uniwersytecie w Pawii, wymyślił nowy sposób barwienie komórek - srebrzenie. Kiedy jedna z tysięcy pobliskich komórek zmieni kolor na srebrny, tylko ona zostanie zabarwiona - jedyna, ale całkowicie, ze wszystkimi swoimi procesami. Metoda Golgiego znacznie pomogło w badaniu struktury komórek nerwowych. Jego zastosowanie pokazało, że pomimo tego, że komórki w mózgu znajdują się niezwykle blisko siebie, a ich procesy są zdezorientowane, to jednak każda komórka jest nadal wyraźnie od siebie oddzielona. Oznacza to, że mózg, podobnie jak inne tkanki, składa się z pojedynczych komórek, które nie są połączone we wspólną sieć. Do takiego wniosku doszedł hiszpański histolog S. Ramon y Cahalem, który w ten sposób rozszerzył teorię komórkową na układ nerwowy. Odrzucenie koncepcji połączonej sieci oznaczało, że w układzie nerwowym puls przechodzi z komórki do komórki nie poprzez bezpośredni kontakt elektryczny, ale przez nią luka
Kiedy stosowano go w biologii? mikroskop elektronowy, który został wynaleziony w 1931 roku M. Knollem I E. Ruska, te pomysły dotyczące obecności luki otrzymały bezpośrednie potwierdzenie.
STRUKTURA I FUNKCJA SYNAPSY:
Każdy organizm wielokomórkowy, każda tkanka składająca się z komórek potrzebuje mechanizmów zapewniających interakcje międzykomórkowe. Przyjrzyjmy się, jak są przeprowadzane międzyneuronowyinterakcje. Informacje wędrują wzdłuż komórki nerwowej w formie potencjały czynnościowe. Przeniesienie wzbudzenia z zakończeń aksonów do unerwionego narządu lub innej komórki nerwowej następuje poprzez międzykomórkowe formacje strukturalne - synapsy(z greckiego „Synapsa”- połączenie, komunikacja). Pojęcie synapsy wprowadził angielski fizjolog C. Sherringtona w 1897 r., aby oznaczyć funkcjonalny kontakt między neuronami. Warto dodać, że już w latach 60. ubiegłego wieku ICH. Sieczenow podkreślił, że bez komunikacji międzykomórkowej nie da się wyjaśnić metod powstawania nawet najbardziej elementarnego procesu nerwowego. Im bardziej złożony jest układ nerwowy i większa liczba składników elementów nerwowych mózgu, tym ważniejsze staje się znaczenie kontaktów synaptycznych.
Różne kontakty synaptyczne różnią się od siebie. Jednak przy całej różnorodności synaps są pewne właściwości ogólne ich struktury i funkcje. Dlatego najpierw opisujemy ogólne zasady ich funkcjonowania.
Synapsa - jest złożoną strukturą formacja składająca się z błony presynaptycznej (najczęściej jest to odgałęzienie końcowe aksonu), błony postsynaptycznej (najczęściej jest to odcinek błony ciała lub dendryt innego neuronu), a także szczeliny synaptycznej.
Mechanizm transmisji przez synapsy pozostawał przez długi czas niejasny, choć było oczywiste, że transmisja sygnału w obszarze synaptycznym różni się znacznie od procesu przewodzenia potencjału czynnościowego wzdłuż aksonu. Jednak na początku XX wieku postawiono hipotezę, że zachodzi także transmisja synaptyczna elektryczny Lub chemicznie. Elektryczna teoria transmisji synaptycznej w ośrodkowym układzie nerwowym była znana aż do wczesnych lat 50. XX wieku, ale znacznie straciła na popularności po wykazaniu w wielu przypadkach synapsy chemicznej. synapsy obwodowe. Na przykład, AV Kibiakow, po przeprowadzeniu eksperymentu na zwoju nerwowym oraz zastosowaniu technologii mikroelektrod do wewnątrzkomórkowej rejestracji potencjałów synaptycznych
neurony ośrodkowego układu nerwowego pozwoliły wyciągnąć wnioski na temat chemicznej natury transmisji w synapsach międzyneuronalnych rdzeń kręgowy.
Badania mikroelektrod ostatnie lata wykazało, że w niektórych synapsach interneuronów istnieje mechanizm transmisji elektrycznej. Obecnie stało się oczywiste, że istnieją synapsy posiadające zarówno mechanizm transmisji chemicznej, jak i elektryczną. Co więcej, w niektórych strukturach synaptycznych zarówno elektrycznych, jak i mechanizmy chemiczne przekładnie to tzw synapsy mieszane.
Nazywa się obszar kontaktu między dwoma neuronami synapsa.
Struktura wewnętrzna synapsa aksodendrytyczna.A) Synapsy elektryczne. Synapsy elektryczne są rzadkie w układzie nerwowym ssaków. Tworzą je połączenia szczelinowe (nexusy) pomiędzy dendrytami lub somatami sąsiadujących neuronów, które są połączone kanałami cytoplazmatycznymi o średnicy 1,5 nm. Proces przekazywania sygnału odbywa się bez opóźnienia synaptycznego i bez udziału mediatorów.
Poprzez synapsy elektryczne potencjały elektrotoniczne mogą rozprzestrzeniać się z jednego neuronu na drugi. Ze względu na bliski kontakt synaptyczny modulacja transmisji sygnału jest niemożliwa. Zadaniem tych synaps jest jednoczesne pobudzenie neuronów pełniących tę samą funkcję. Przykładem są neurony ośrodka oddechowego rdzenia przedłużonego, które synchronicznie generują impulsy podczas wdechu. Ponadto przykładem są obwody nerwowe kontrolujące sakady, w których punkt fiksacji wzroku przesuwa się z jednego obiektu uwagi na drugi.
B) Synapsy chemiczne. Większość synaps w układzie nerwowym ma charakter chemiczny. Funkcjonowanie takich synaps zależy od uwolnienia przekaźników. Klasyczna synapsa chemiczna jest reprezentowana przez błonę presynaptyczną, szczelinę synaptyczną i błonę postsynaptyczną. Błona presynaptyczna jest częścią maczugowatego przedłużenia zakończenia nerwowego komórki, która przekazuje sygnał, a błona postsynaptyczna jest częścią komórki, która odbiera sygnał.
Nadajnik jest uwalniany z przedłużenia obojczyka w wyniku egzocytozy, przechodzi przez szczelinę synaptyczną i wiąże się z receptorami na błonie postsynaptycznej. Pod błoną postsynaptyczną znajduje się subsynaptyczna strefa aktywna, w której po aktywacji receptorów błony postsynaptycznej zachodzą różne procesy biochemiczne.
Przedłużenie w kształcie maczugi zawiera pęcherzyki synaptyczne zawierające mediatory, a także dużą liczbę mitochondriów i zbiorników gładkiej siateczki śródplazmatycznej. Zastosowanie tradycyjnych technik fiksacji w badaniu komórek umożliwia wyróżnienie presynaptycznych uszczelnień na błonie presynaptycznej, ograniczających aktywne strefy synapsy, do których kierowane są pęcherzyki synaptyczne za pomocą mikrotubul.
 Synapsa aksodendrytyczna.
Synapsa aksodendrytyczna. Fragment rdzenia kręgowego: synapsa pomiędzy końcową częścią dendrytu i prawdopodobnie neuronem ruchowym.
Charakterystyczną cechą synaps pobudzających jest obecność okrągłych pęcherzyków synaptycznych i zagęszczenie postsynaptyczne.
Dendryt przecięto w kierunku poprzecznym, o czym świadczy obecność wielu mikrotubul.
Ponadto widoczne są niektóre neurofilamenty. Miejsce synapsy jest otoczone przez astrocyt protoplazmatyczny.
 Procesy zachodzące w dwóch rodzajach zakończeń nerwowych.
Procesy zachodzące w dwóch rodzajach zakończeń nerwowych. (A) Transmisja synaptyczna małe cząsteczki(np. glutaminian).
(1) Pęcherzyki transportowe zawierające białka błonowe pęcherzyków synaptycznych kierowane są wzdłuż mikrotubul do błony komórkowej zgrubienia maczugowatego.
Jednocześnie cząsteczki enzymów i glutaminianu są przenoszone w drodze powolnego transportu.
(2) Białka błonowe pęcherzyków opuszczają błonę komórkową i tworzą pęcherzyki synaptyczne.
(3) Glutaminian jest ładowany do pęcherzyków synaptycznych; następuje akumulacja mediatorów.
(4) Pęcherzyki zawierające glutaminian zbliżają się do błony presynaptycznej.
(5) W wyniku depolaryzacji dochodzi do egzocytozy mediatora z częściowo zniszczonych pęcherzyków.
(6) Uwolniony przekaźnik rozprzestrzenia się rozproszonie w obszarze szczeliny synaptycznej i aktywuje specyficzne receptory na błonie postsynaptycznej.
(7) Błony pęcherzyków synaptycznych są transportowane z powrotem do komórki na drodze endocytozy.
(8) Następuje częściowy wychwyt zwrotny glutaminianu do komórki w celu jego ponownego wykorzystania.
(B) Transmisja neuropeptydów (np. substancji P) zachodząca jednocześnie z transmisją synaptyczną (np. glutaminianu).
Wspólna transmisja tych substancji zachodzi w ośrodkowych zakończeniach nerwowych neuronów jednobiegunowych, które zapewniają wrażliwość na ból.
(1) Pęcherzyki i prekursory peptydów (propeptydy) syntetyzowane w kompleksie Golgiego (w obszarze perykarionu) są transportowane do maczugowatego przedłużenia poprzez szybki transport.
(2) Kiedy dostaną się w obszar maczugowatego zgrubienia, proces tworzenia cząsteczki peptydu zostaje zakończony, a pęcherzyki są transportowane do błony komórkowej.
(3) Depolaryzacja błony i przeniesienie zawartości pęcherzyków do przestrzeni międzykomórkowej na drodze egzocytozy.
(4) Jednocześnie uwalniany jest glutaminian.
1. Aktywacja receptora. Cząsteczki przekaźnikowe przechodzą przez szczelinę synaptyczną i aktywują białka receptorowe zlokalizowane parami na błonie postsynaptycznej. Aktywacja receptorów wyzwala procesy jonowe, które prowadzą do depolaryzacji błony postsynaptycznej (pobudzające działanie postsynaptyczne) lub hiperpolaryzacji błony postsynaptycznej (hamujące działanie postsynaptyczne). Zmiana elektrotoniczności przekazywana jest do somy w postaci potencjału elektrotonicznego, który zanika w miarę rozprzestrzeniania się, przez co zmienia się potencjał spoczynkowy w początkowym odcinku aksonu.
Procesy jonowe zostały szczegółowo opisane w osobnym artykule na stronie. Kiedy przeważają pobudzające potencjały postsynaptyczne, początkowy odcinek aksonu ulega depolaryzacji do poziomu progowego i generuje potencjał czynnościowy.
Najczęstszym neuroprzekaźnikiem pobudzającym ośrodkowy układ nerwowy jest glutaminian, a hamującym kwas gamma-aminomasłowy (GABA). W obwodowym układzie nerwowym acetylocholina pełni funkcję przekaźnika dla neuronów ruchowych mięśni poprzecznie prążkowanych, a glutaminian dla neuronów czuciowych.
Kolejność procesów zachodzących w synapsach glutaminergicznych przedstawiono na poniższym rysunku. Kiedy glutaminian jest przenoszony razem z innymi peptydami, uwalnianie peptydów następuje poprzez szlaki pozasynaptyczne.
Większość neuronów czuciowych, oprócz glutaminianu, wydziela także inne peptydy (jeden lub więcej), uwalniane w różnych częściach neuronu; jednakże główną funkcją tych peptydów jest modulowanie (zwiększanie lub zmniejszanie) wydajności synaptycznej transmisji glutaminianu.
Ponadto neurotransmisja może zachodzić poprzez rozproszoną pozasynaptyczną transmisję sygnału, charakterystyczną dla neuronów monoaminergicznych (neuronów, które wykorzystują aminy biogenne do pośredniczenia w neurotransmisji). Istnieją dwa typy neuronów monoaminergicznych. W niektórych neuronach katecholaminy (norepinefryna lub dopamina) są syntetyzowane z aminokwasu tyrozyny, a w innych serotonina jest syntetyzowana z aminokwasu tryptofanu. Przykładowo dopamina uwalniana jest zarówno w obszarze synaptycznym, jak i z żylaków aksonalnych, w których zachodzi także synteza tego neuroprzekaźnika.
Dopamina przenika do płynu międzykomórkowego ośrodkowego układu nerwowego i przed degradacją jest w stanie aktywować określone receptory w odległości do 100 mikronów. Neurony monoaminergiczne są obecne w wielu strukturach ośrodkowego układu nerwowego; zakłócenie przekazywania impulsów przez te neurony prowadzi do różnych chorób, w tym choroby Parkinsona, schizofrenii i ciężkiej depresji.
Tlenek azotu (cząsteczka gazowa) bierze także udział w rozproszonej neurotransmisji w glutaminergicznym układzie neuronowym. Nadmiar tlenku azotu działa cytotoksycznie, szczególnie w tych obszarach, gdzie ukrwienie jest upośledzone na skutek zakrzepicy tętniczej. Glutaminian jest także potencjalnie cytotoksycznym neuroprzekaźnikiem.
W przeciwieństwie do rozproszonej neurotransmisji, tradycyjna transmisja sygnału synaptycznego nazywana jest „przewodnikiem” ze względu na jej względną stabilność.
V) Streszczenie. Neurony wielobiegunowe OUN składają się z somy, dendrytów i aksonu; akson tworzy gałęzie boczne i końcowe. Soma zawiera gładką i szorstką siateczkę śródplazmatyczną, kompleksy Golgiego, neurofilamenty i mikrotubule. Mikrotubule przenikają cały neuron, biorą udział w procesie postępującego transportu pęcherzyków synaptycznych, mitochondriów i substancji budujących błony, a także zapewniają wsteczny transport cząsteczek „markerowych” i zniszczonych organelli.
Istnieją trzy typy chemicznych interakcji międzyneuronalnych: synaptyczne (np. glutaminergiczne), pozasynaptyczne (peptydergiczne) i rozproszone (np. monoaminergiczne, serotoninergiczne).
Ze względu na budowę anatomiczną synapsy chemiczne dzielą się na aksodendrytyczne, aksosomatyczne, aksoaksonalne i dendrodendrytyczne. Synapsa jest reprezentowana przez błony pre- i postsynaptyczne, szczelinę synaptyczną i subsynaptyczną strefę aktywną.
Synapsy elektryczne zapewniają jednoczesną aktywację całych grup, tworząc między nimi połączenia elektryczne dzięki stykom szczelinowym (nexusom).
Aksony neuronów glutaminergicznych (1) i dopaminergicznych (2) tworzą ścisłe kontakty synaptyczne z wyrostkiem neuronu gwiaździstego (3) prążkowia.
Dopamina jest uwalniana nie tylko z obszaru presynaptycznego, ale także z żylakowatego zgrubienia aksonu, skąd dyfunduje do przestrzeni międzykomórkowej i aktywuje receptory dopaminowe pnia dendrytycznego i ścian perycytów naczyń włosowatych.
 Odhamowanie.
Odhamowanie. (A) Neuron pobudzający 1 aktywuje neuron hamujący 2, który z kolei hamuje neuron 3.
(B) Pojawienie się drugiego neuronu hamującego (2b) ma odwrotny wpływ na neuron 3, ponieważ neuron 2b jest hamowany.
Spontanicznie aktywny neuron 3 generuje sygnały przy braku wpływów hamujących.
2. Leki - „klucze” i „zamki”. Receptor można porównać do zamka, a mediator do pasującego do niego klucza. Jeżeli proces uwalniania mediatora zostanie zakłócony z wiekiem lub na skutek jakiejkolwiek choroby, lek może pełnić rolę „zapasowego klucza”, który pełni funkcję zbliżoną do mediatora. Lek ten nazywany jest agonistą. Jednocześnie w przypadku nadmiernej produkcji mediator może zostać „przechwycony” przez bloker receptora – „fałszywy klucz”, który zetknie się z receptorem „zamka”, ale nie spowoduje jego aktywacji.
3. Hamowanie i odhamowanie. Funkcjonowanie spontanicznie aktywnych neuronów jest hamowane przez wpływ neuronów hamujących (zwykle GABAergicznych). Z kolei aktywność neuronów hamujących może zostać zahamowana przez działające na nie inne neurony hamujące, co skutkuje odhamowaniem komórki docelowej. Proces odhamowania jest ważną cechą aktywności neuronalnej w zwojach podstawy.
4. Rzadkie typy synaps chemicznych. Istnieją dwa typy synaps aksonalnych. W obu przypadkach zgrubienie w kształcie maczugi tworzy neuron hamujący. Synapsy pierwszego typu powstają w obszarze początkowego odcinka aksonu i przekazują silny efekt hamujący neuronu hamującego. Synapsy drugiego typu powstają pomiędzy maczugowatym pogrubieniem neuronu hamującego a maczugowatym pogrubieniem neuronów pobudzających, co prowadzi do zahamowania uwalniania przekaźników. Proces ten nazywany jest hamowaniem presynaptycznym. Pod tym względem tradycyjna synapsa zapewnia hamowanie postsynaptyczne.
Synapsy dendrodendrytyczne (D-D) powstają pomiędzy kolcami dendrytycznymi dendrytów sąsiadujących neuronów kolczastych. Ich zadaniem nie jest wygenerowanie impulsu nerwowego, ale zmiana napięcia elektrycznego komórki docelowej. W kolejnych synapsach D-D pęcherzyki synaptyczne znajdują się tylko w jednym kolcu dendrytycznym, a w synapsach wzajemnych D-D w obu. Pobudzające synapsy D-D pokazano na poniższym rysunku. Hamujące synapsy D-D są szeroko reprezentowane w jądrach przełączających wzgórza.
Ponadto istnieje kilka synaps somato-dendrytycznych i somato-somatycznych.
 Synapsy aksoaksonalne kory mózgowej.
Synapsy aksoaksonalne kory mózgowej. Strzałki wskazują kierunek impulsów.
 (1) Presynaptyczne i (2) postsynaptyczne hamowanie neuronu rdzeniowego przemieszczającego się do mózgu.
(1) Presynaptyczne i (2) postsynaptyczne hamowanie neuronu rdzeniowego przemieszczającego się do mózgu. Strzałki wskazują kierunek przewodzenia impulsu (możliwe jest zahamowanie neuronu przełączającego pod wpływem wpływów hamujących).
 Pobudzające synapsy dendro-dendrytyczne. Przedstawiono dendryty trzech neuronów.
Pobudzające synapsy dendro-dendrytyczne. Przedstawiono dendryty trzech neuronów. Synapsa wzajemna (po prawej). Strzałki wskazują kierunek propagacji fal elektrotonicznych.
Film edukacyjny - budowa synapsy
Synapsa to pewna strefa kontaktu procesów komórek nerwowych z innymi komórkami niepobudliwymi i pobudliwymi, które zapewniają przekazywanie sygnału informacyjnego. Synapsa jest morfologicznie utworzona przez stykające się błony 2 komórek. Błona związana z tym procesem nazywana jest błoną presynaptyczną komórki, do której odbierany jest sygnał, a jej druga nazwa to postsynaptyczna. Wraz z błoną postsynaptyczną synapsa może być międzyneuronalna, nerwowo-mięśniowa i neurosekrecyjna. Słowo synapsa zostało wprowadzone w 1897 roku przez Charlesa Sherringtona (angielski fizjolog).
Co to jest synapsa?
Synapsa to specjalna struktura, która zapewnia przekazywanie impulsu nerwowego z włókna nerwowego do innego włókna nerwowego lub komórki nerwowej i aby komórka receptorowa (obszar kontaktu między komórkami nerwowymi) oddziaływała na włókno nerwowe i kolejne włókno nerwowe), wymagane są dwie komórki nerwowe.
Synapsa to niewielka część znajdująca się na końcu neuronu. Za jego pomocą informacja jest przekazywana z pierwszego neuronu do drugiego. Synapsa znajduje się w trzech obszarach komórek nerwowych. Synapsy znajdują się również w miejscu, w którym komórka nerwowa łączy się z różnymi gruczołami lub mięśniami ciała.
Z czego składa się synapsa?
Struktura synapsy ma prosty schemat. Składa się z 3 części, z których każda pełni określone funkcje podczas przesyłania informacji. Zatem tę strukturę synapsy można nazwać odpowiednią do transmisji.Na proces ten mają bezpośredni wpływ dwie główne komórki: odbierająca i nadawcza. Na końcu aksonu komórki nadawczej znajduje się zakończenie presynaptyczne (początkowa część synapsy). Może wpływać na uruchomienie neuroprzekaźników w komórce (słowo to ma kilka znaczeń: mediatory, pośredniki lub neuroprzekaźniki) – definiowane za pomocą których realizowana jest transmisja sygnału elektrycznego pomiędzy 2 neuronami.

Szczelina synaptyczna to środkowa część synapsy – jest to przerwa pomiędzy 2 oddziałującymi ze sobą komórkami nerwowymi. Przez tę szczelinę impuls elektryczny pochodzi z komórki nadawczej. Za część recepcyjną komórki uważa się końcową część synapsy, czyli zakończenie postsynaptyczne (fragment komórki stykający się w jej strukturze z różnymi wrażliwymi receptorami).
Mediatory synaps
Mediator (z języka łacińskiego Media - nadawca, pośrednik lub środek). Takie mediatory synaptyczne są bardzo ważne w procesie transmisji
Różnica morfologiczna między synapsami hamującymi i pobudzającymi polega na tym, że nie mają one mechanizmu uwalniania przekaźnika. Za przekaźnik w synapsie hamującej, neuronie ruchowym i innych synapsach hamujących uważa się aminokwas glicynę. Ale o hamującym lub pobudzającym charakterze synapsy decydują nie ich mediatory, ale właściwość błony postsynaptycznej. Na przykład acetylocholina działa stymulująco na zakończenia synaps nerwowo-mięśniowych (nerwy błędne w mięśniu sercowym).
Acetylocholina służy jako przekaźnik pobudzający w synapsach cholinergicznych (błona presynaptyczna w niej odgrywa zakończenie rdzenia kręgowego neuronu ruchowego), w synapsie na komórkach Renshawa, w presynaptycznym zakończeniu gruczołów potowych, rdzeniu nadnerczy, w synapsie jelitowej i zwojach współczulnego układu nerwowego. Acetylocholinoesterazę i acetylocholinę znaleziono także we frakcjach różnych części mózgu, czasami w duże ilości, ale poza synapsą cholinergiczną na komórkach Renshawa nie udało im się jeszcze zidentyfikować innych synaps cholinergicznych. Według naukowców bardzo prawdopodobna jest mediatorowa funkcja pobudzająca acetylocholiny w ośrodkowym układzie nerwowym.

Catelchominy (dopamina, norepinefryna i epinefryna) są uważane za mediatory adrenergiczne. Adrenalina i noradrenalina są syntetyzowane na końcu nerwu współczulnego, w komórkach mózgowych nadnercza, rdzenia kręgowego i mózgu. Za materiał wyjściowy uważa się aminokwasy (tyrozyna i L-fenyloalanina), a końcowym produktem syntezy jest adrenalina. Substancja pośrednia, do której zalicza się norepinefrynę i dopaminę, pełni także funkcję mediatorów w synapsie utworzonej na zakończeniach nerwów współczulnych. Funkcja ta może być hamująca (gruczoły wydzielnicze jelit, kilka zwieraczy i mięśnie gładkie oskrzeli i jelit) lub pobudzająca (mięśnie gładkie niektórych zwieraczy i naczyń krwionośnych, w synapsie mięśnia sercowego - noradrenalina, w podskórnych jądrach mózgu - dopamina).
Kiedy mediatory synaps zakończą swoją funkcję, katecholamina zostaje wchłonięta przez presynaptyczne zakończenie nerwowe i aktywowany jest transport przezbłonowy. Podczas wchłaniania przekaźników synapsy są chronione przed przedwczesnym wyczerpaniem się podaży podczas długiej i rytmicznej pracy.
Synapsa: główne typy i funkcje
Langley w 1892 zasugerował, że transmisja synaptyczna w zwoju autonomicznym ssaków nie zachodzi charakter elektryczny, ale chemiczny. Dziesięć lat później Elliott odkrył, że adrenalina jest wytwarzana w nadnerczach w wyniku tego samego działania, co stymulacja nerwów współczulnych.

Następnie zasugerowano, że adrenalina może być wydzielana przez neurony, a po wzbudzeniu uwalniana przez zakończenia nerwowe. Jednak w 1921 roku Levy przeprowadził eksperyment, w którym ustalił chemiczną naturę transmisji w synapsie autonomicznej między sercem a nerwami błędnymi. Wypełnił naczynia solą fizjologiczną i pobudził nerw błędny, powodując spowolnienie akcji serca. Kiedy płyn został przeniesiony z serca o zahamowanej stymulacji do serca, które nie miało stymulacji, biło ono wolniej. Jest oczywiste, że pobudzenie nerwu błędnego spowodowało uwolnienie do roztworu substancji hamującej. Acetylocholina całkowicie odtworzyła działanie tej substancji. W 1930 roku Feldberg i jego współpracownik ostatecznie ustalili rolę acetylocholiny w transmisji synaptycznej w zwoju.
Synapsa chemiczna
Synapsa chemiczna zasadniczo różni się pod względem przenoszenia podrażnienia za pomocą przekaźnika z presynapsy do postsynapsy. Dlatego powstają różnice w morfologii synapsy chemicznej. Synapsa chemiczna występuje częściej w OUN kręgowym. Obecnie wiadomo, że neuron jest zdolny do uwalniania i syntezy pary przekaźników (współistniejących nadajników). Neurony charakteryzują się także plastycznością neuroprzekaźnika – możliwością zmiany głównego przekaźnika w trakcie rozwoju.

Złącze nerwowo-mięśniowe
Ta synapsa przekazuje pobudzenie, ale to połączenie może zostać zniszczone przez różne czynniki. Transmisja kończy się podczas blokady uwalniania acetylocholiny do szczeliny synaptycznej, a także podczas nadmiaru jej zawartości w obszarze błon postsynaptycznych. Wiele trucizn i leków wpływa na wychwyt, wyjście, które jest związane z receptorami cholinergicznymi błony postsynaptycznej, następnie synapsa mięśniowa blokuje przekazywanie wzbudzenia. Ciało umiera podczas uduszenia i zatrzymania skurczu mięśni oddechowych.

Botulinus jest toksyną drobnoustrojową występującą w synapsie; blokuje przekazywanie wzbudzenia poprzez niszczenie białka syntaksyny na końcu presynaptycznym, co jest kontrolowane przez uwalnianie acetylocholiny do szczeliny synaptycznej. Kilka toksycznych środków bojowych, leków farmakologicznych (neostygmina i proseryna), a także insektycydów blokują przewodzenie wzbudzenia w synapsie nerwowo-mięśniowej poprzez inaktywację acetylocholinoesterazy, enzymu niszczącego acetylocholinę. Dlatego acetylocholina gromadzi się w obszarze błony postsynaptycznej, wrażliwość na mediator maleje, a blok receptorowy zostaje uwolniony z błony postsynaptycznej i zanurzony w cytozolu. Acetylocholina będzie nieskuteczna, a synapsa zostanie zablokowana.
Synapsa nerwowa: cechy i elementy
Synapsa to połączenie pomiędzy punktem styku dwóch komórek. Co więcej, każdy z nich jest zamknięty we własnej elektrogenicznej membranie. Synapsa nerwowa składa się z trzech głównych elementów: błony postsynaptycznej, szczeliny synaptycznej i błony presynaptycznej. Błona postsynaptyczna to zakończenie nerwowe, które przechodzi do mięśnia i schodzi do tkanki mięśniowej. W obszarze presynaptycznym znajdują się pęcherzyki - są to zamknięte wnęki zawierające przekaźnik. Są zawsze w ruchu.

Zbliżając się do błony zakończeń nerwowych, pęcherzyki łączą się z nią, a przekaźnik wchodzi do szczeliny synaptycznej. Jeden pęcherzyk zawiera kwant mediatora oraz mitochondria (są one potrzebne do syntezy mediatora – głównego źródła energii), następnie z choliny syntetyzowana jest acetylocholina i pod wpływem enzymu transferazy acetylocholinowej przetwarzana jest do acetyloCoA) .
Szczelina synaptyczna pomiędzy błonami post- i presynaptycznymi
Rozmiar szczeliny jest różny w różnych synapsach. wypełnione płynem międzykomórkowym, który zawiera mediator. Błona postsynaptyczna pokrywa miejsce kontaktu zakończenia nerwowego z unerwioną komórką w synapsie mięśniowo-nerwowej. W niektórych synapsach błona postsynaptyczna fałduje się i zwiększa się powierzchnia kontaktu.
Dodatkowe substancje tworzące błonę postsynaptyczną
W strefie błony postsynaptycznej występują następujące substancje:
Receptor (receptor cholinergiczny w synapsie mięśniowo-nerwowej).
Lipoproteina (bardzo podobna do acetylocholiny). Białko to ma koniec elektrofilowy i głowę jonową. Głowa wchodzi do szczeliny synaptycznej i oddziałuje z kationową głową acetylocholiny. W wyniku tej interakcji następuje zmiana błony postsynaptycznej, następnie następuje depolaryzacja i otwierają się kanały Na bramkowane potencjałem. Depolaryzacji błony nie uważa się za proces samowzmacniający;
Jest stopniowy, jego potencjał na błonie postsynaptycznej zależy od liczby mediatorów, to znaczy potencjał charakteryzuje się właściwością lokalnych wzbudzeń.
Cholinoesteraza jest uważana za białko posiadające funkcję enzymatyczną. Ma podobną budowę do receptora cholinergicznego i ma podobne właściwości do acetylocholiny. Cholinoesteraza niszczy acetylocholinę, w pierwszej kolejności tę związaną z receptorem cholinergicznym. Pod wpływem cholinoesterazy receptor cholinergiczny usuwa acetylocholinę, co powoduje repolaryzację błony postsynaptycznej. Acetylocholina rozkłada się do kwas octowy i cholina, niezbędna do trofizmu tkanki mięśniowej.
Za pomocą transportu aktywnego cholina usuwana jest do błony presynaptycznej, gdzie wykorzystywana jest do syntezy nowego przekaźnika. Pod wpływem mediatora zmienia się przepuszczalność błony postsynaptycznej, a pod wpływem cholinoesterazy czułość i przepuszczalność wraca do wartości początkowej. Chemoreceptory są w stanie oddziaływać z nowymi mediatorami.
Wykład 2. Fizjologia synaps: budowa, klasyfikacja i mechanizmy działania. Mediatory, neurochemiczne podstawy zachowania.
Pod koniec XIX wieku istniały dwie równoległe teorie organizacji układu nerwowego (NS). Teoria siatkowa wierzył, że NS jest funkcjonalnym syncytium: neurony są połączone procesami podobnymi do naczyń włosowatych układu krążenia. Według Teoria komórkowa Waldeyera(1981) NS składa się z pojedynczych neuronów oddzielonych błonami. Aby rozwiązać problem interakcji pomiędzy poszczególnymi neuronami, Sherringtona w 1987 roku zasugerował obecność specjalnej formacji membranowej - synapsa. Za pomocą mikroskopu elektronowego jednoznacznie potwierdzono obecność synaps. Jednakże komórkowa teoria budowy układu nerwowego została ogólnie przyjęta; jak na ironię, w 1959 roku Fershpan i Potter odkryli synapsę ze złączami szczelinowymi (synapsę elektryczną) w układzie nerwowym skorupiaków.
Synapsa jest formacją błonową dwóch (lub więcej) komórek, w której wzbudzenie (informacja) jest przenoszone z jednej komórki do drugiej.
Istnieje następujący podział synaps:
1) przez mechanizm przenoszenia wzbudzenia (i strukturę):
Chemiczny;
Elektryczne (ephaps);
Mieszany.
2) według uwolnionego neuroprzekaźnika:
Adrenergiczny – neuroprzekaźnik noradrenalina;
Cholinergiczne – neuroprzekaźnik acetylocholina;
Dopaminergiczny – neuroprzekaźnik dopamina;
Serotonergiczny – neuroprzekaźnik serotonina;
GABAergiczny – neuroprzekaźnik kwas gamma-aminomasłowy (GABA)
3) pod wpływem:
Ekscytujący;
Hamulec.
4) według lokalizacji:
nerwowo-mięśniowy;
Neuro-neuralny:
a) aksosomatyczny;
b) aksoaksonalny;
c) akso-dendrytyczny;
d) dendrosomatyczny.
Rozważmy trzy typy synaps: chemiczne, elektryczne i mieszane(łączenie właściwości synaps chemicznych i elektrycznych).
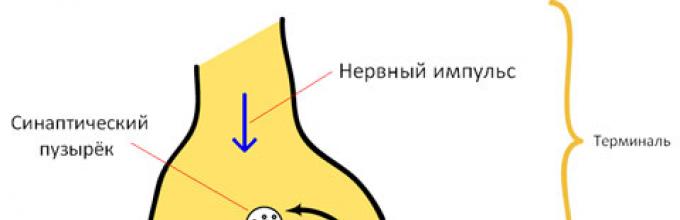
Niezależnie od rodzaju synapsy mają wspólne cechy strukturalne: wyrostek nerwowy na końcu tworzy przedłużenie ( płytka synaptyczna, SB); błona końcowa SB różni się od innych części błony neuronowej i nazywa się błona presynaptyczna(PreSM); wyspecjalizowana błona drugiej komórki nazywana jest błoną postsynaptyczną (PostSM); znajduje się pomiędzy błonami synapsy szczelina synaptyczna(SCH, ryc. 1, 2).
Ryż. 1. Schemat budowy synapsy chemicznej
Synapsy elektryczne(ephapses, ES) są dziś spotykane w północnej części kraju nie tylko skorupiaków, ale także mięczaków, stawonogów i ssaków. ES mają wiele unikalnych właściwości. Mają wąską szczelinę synaptyczną (około 2-4 nm), dzięki czemu wzbudzenie może być przekazywane elektrochemicznie (jak przez włókno nerwowe z powodu pola elektromagnetycznego) przy dużej prędkości i w obu kierunkach: zarówno z membrany PreSM do PostSM, jak i z PostSM do PreSM. Pomiędzy komórkami znajdują się połączenia szczelinowe (koneksony lub koneksony), utworzone przez dwa białka zwane koneksynami. Sześć podjednostek każdej koneksyny tworzy kanały PreSM i PostSM, przez które komórki mogą wymieniać substancje niskocząsteczkowe o masie cząsteczkowej 1000-2000 daltonów. Pracę koneksonów można regulować za pomocą jonów Ca 2+ (ryc. 2).

Ryż. 2. Schemat synapsy elektrycznej
ES mają większą specjalizację w porównaniu z synapsami chemicznymi i zapewniają wysoką prędkość transmisji wzbudzenia. Wydaje się jednak, że jest pozbawiony możliwości bardziej subtelnej analizy (regulacji) przesyłanej informacji.
Synapsy chemiczne dominują w NS. Historia ich badań rozpoczyna się od prac Claude'a Bernarda, który w 1850 roku opublikował artykuł „Badania nad kurare”. Oto co napisał: „Kurara to silna trucizna przygotowana przez niektóre ludy (głównie kanibali) zamieszkujące lasy… Amazonii”. I dalej: „Kurara jest podobna do jadu węża w tym sensie, że można ją bezkarnie wprowadzić do przewodu pokarmowego człowieka lub zwierzęcia, natomiast wstrzyknięcie pod skórę lub w jakąkolwiek część ciała szybko prowadzi do śmierci. ...po kilku chwilach zwierzęta kładą się jak zmęczone. Potem oddech ustaje, ich wrażliwość i życie znikają, a zwierzęta nie wydają krzyku ani nie okazują żadnych oznak bólu. Choć na pomysł chemicznego przekazywania impulsów nerwowych nie wpadł C. Bernard, to jego klasyczne eksperymenty z kurarą pozwoliły na powstanie tego pomysłu. Minęło ponad pół wieku, kiedy J. Langley ustalił (1906), że paraliżujące działanie kurary jest związane ze specjalną częścią mięśnia, którą nazwał substancją receptywną. Pierwszą sugestię dotyczącą przeniesienia wzbudzenia z nerwu na narząd efektorowy za pomocą substancji chemicznej przedstawił T. Eliot (1904).
Jednak dopiero prace G. Dale'a i O. Löwy'ego ostatecznie potwierdziły hipotezę o synapsie chemicznej. Dale w 1914 roku ustalił, że acetylocholina imituje podrażnienie nerwu przywspółczulnego. Löwy udowodnił w 1921 r., że acetylocholina jest uwalniana z zakończenia nerwowego nerwu błędnego, a w 1926 r. odkrył acetylocholinoesterazę, enzym niszczący acetylocholinę.
Wzbudzenie w synapsie chemicznej jest przekazywane za pomocą mediator. Proces ten obejmuje kilka etapów. Rozważmy te cechy na przykładzie synapsy acetylocholiny, która jest szeroko rozpowszechniona w ośrodkowym układzie nerwowym, autonomicznym i obwodowym układzie nerwowym (ryc. 3).

Ryż. 3. Schemat funkcjonowania synapsy chemicznej
1. Mediator acetylocholina (ACh) jest syntetyzowana w płytce synaptycznej z acetylo-CoA (acetylo-koenzym A powstaje w mitochondriach) i choliny (syntetyzowanej w wątrobie) za pomocą transferazy acetylocholinowej (ryc. 3, 1).
2. Pick jest zapakowany pęcherzyki synaptyczne ( Castillo, Katz; 1955). Ilość mediatora w jednym pęcherzyku wynosi kilka tysięcy cząsteczek ( kwant mediatora). Część pęcherzyków znajduje się na PreSM i jest gotowa do uwolnienia mediatora (ryc. 3, 2).
3. Mediator zostaje zwolniony przez egzocytoza po wzbudzeniu PreSM. Przychodzący prąd odgrywa ważną rolę w pęknięciu membrany i uwolnieniu kwantowym nadajnika. Ok 2+ (ryc. 3, 3).
4. Wydany wybór wiąże się ze specyficznym białkiem receptorowym PostSM (ryc. 3, 4).
5. W wyniku interakcji między mediatorem a receptorem zmiany przewodnictwa jonowego PostSM: kiedy kanały Na+ są otwarte, depolaryzacja; prowadzi do otwarcia kanałów K + lub Cl - hiperpolaryzacja(ryc. 3, 5).
6 . W wyniku depolaryzacji w cytoplazmie postsynaptycznej uruchamiają się procesy biochemiczne (ryc. 3, 6).
7. Receptor zostaje uwolniony od mediatora: ACh ulega zniszczeniu przez acetylocholinoesterazę (AChE, ryc. 3.7).
Początek formularza
Proszę to zanotować mediator zwykle oddziałuje z określonym receptorem z określoną siłą i czasem trwania. Dlaczego kurara jest trucizną? Miejscem działania kurary jest właśnie synapsa ACh. Kurara mocniej wiąże się z receptorem acetylocholiny i pozbawia go interakcji z neuroprzekaźnikiem (ACh). Pobudzenie z nerwów somatycznych do mięśni szkieletowych, w tym od nerwu przeponowego do głównego mięśnia oddechowego (przepony) przekazywane jest za pomocą ACh, więc kurara powoduje rozluźnienie mięśni i ustanie oddychania (co w rzeczywistości powoduje śmierć).
Zwróćmy uwagę na główne Cechy transmisji wzbudzenia w synapsie chemicznej.
1. Wzbudzenie przekazywane jest za pomocą pośrednika chemicznego - mediatora.
2. Wzbudzenie przekazywane jest w jednym kierunku: z PreSm do PostSm.
3. W synapsie chemicznej występuje tymczasowe opóźnienie w przewodzeniu wzbudzenia, dlatego ma to synapsa niska labilność.
4. Synapsa chemiczna jest bardzo wrażliwa na działanie nie tylko mediatorów, ale także innych substancji biologicznie czynnych, leków i trucizn.
5. W synapsie chemicznej następuje transformacja wzbudzeń: elektrochemiczny charakter wzbudzenia na PreSM przechodzi w biochemiczny proces egzocytozy pęcherzyków synaptycznych i wiązania mediatora z określonym receptorem. Następuje zmiana przewodności jonowej PostSM (również proces elektrochemiczny), która kontynuowana jest przez reakcje biochemiczne w cytoplazmie postsynaptycznej.
W zasadzie takie wielostopniowe przeniesienie wzbudzenia powinno mieć znaczne znaczenie znaczenie biologiczne. Należy pamiętać, że na każdym etapie istnieje możliwość regulacji procesu przekazywania wzbudzenia. Pomimo ograniczonej liczby mediatorów (nieco kilkanaście) w synapsie chemicznej panują warunki umożliwiające różnorodne decydowanie o losach pobudzenia nerwu dochodzącego do synapsy. Połączenie cech synaps chemicznych wyjaśnia indywidualne zróżnicowanie biochemiczne procesów nerwowych i umysłowych.
Zatrzymajmy się teraz na dwóch ważnych procesach zachodzących w przestrzeni postsynaptycznej. Zauważyliśmy, że w wyniku interakcji ACh z receptorem na PostSM może rozwinąć się zarówno depolaryzacja, jak i hiperpolaryzacja. Co decyduje o tym, czy mediator będzie pobudzający, czy hamujący? Wynik interakcji pomiędzy mediatorem a receptorem zdeterminowane właściwościami białka receptorowego(inną ważną właściwością synapsy chemicznej jest to, że PostSM jest aktywny w stosunku do docierającego do niej pobudzenia). W zasadzie synapsa chemiczna jest formacją dynamiczną, zmieniając receptor, komórka otrzymująca wzbudzenie może na niego wpłynąć przyszły los. Jeśli właściwości receptora są takie, że jego interakcja z nadajnikiem otwiera kanały Na +, to kiedy poprzez izolację jednego kwantu mediatora na PostSM rozwija się potencjał lokalny(dla złącza nerwowo-mięśniowego nazywa się to miniaturowym potencjałem płytki końcowej – MEPP).
Kiedy pojawia się PD? Wzbudzenie PostSM (pobudzający potencjał postsynaptyczny – EPSP) powstaje w wyniku sumowania potencjałów lokalnych. Możesz wybrać dwa rodzaje procesów sumowania. Na sekwencyjne uwalnianie kilku kwantów mediatora w tej samej synapsie(woda niszczy kamień) powstaje tymczasowyA Jestem podsumowaniem. Jeśli Mediatory kwantowe są uwalniane jednocześnie w różnych synapsach(na błonie neuronu może ich być kilka tysięcy). podsumowanie przestrzenne. Repolaryzacja błony PostSM zachodzi powoli i po uwolnieniu poszczególnych kwantów mediatora PostSM przez pewien czas znajduje się w stanie egzaltacji (tzw. wzmocnienie synaptyczne, ryc. 4). Być może w ten sposób dochodzi do treningu synaps (uwolnienie kwantów przekaźnikowych w niektórych synapsach może „przygotować” błonę do zdecydowanej interakcji z przekaźnikiem).
Kiedy kanały K + lub Cl - otwierają się na PostSM, pojawia się hamujący potencjał postsynaptyczny (IPSP, ryc. 4).

Ryż. 4. Postsynaptyczne potencjały błonowe
Naturalnie, jeśli rozwinie się IPSP, można zatrzymać dalszą propagację wzbudzenia. Inną opcją zatrzymania procesu wzbudzenia jest hamowanie presynaptyczne. Jeśli na błonie płytki synaptycznej utworzy się synapsa hamująca, wówczas w wyniku hiperpolaryzacji PreSM można zablokować egzocytozę pęcherzyków synaptycznych.
Drugim ważnym procesem jest rozwój reakcji biochemicznych w cytoplazmie postsynaptycznej. Zmiana przewodności jonowej PostSM aktywuje tzw wtórni posłańcy (pośrednicy): cAMP, cGMP, kinaza białkowa zależna od Ca 2+, które z kolei aktywują różne kinazy białkowe poprzez ich fosforylację. Te reakcje biochemiczne mogą „zejść” w głąb cytoplazmy aż do jądra neuronu, regulując procesy syntezy białek. Zatem komórka nerwowa może odpowiedzieć na nadchodzące pobudzenie nie tylko decydując o swoim dalszym losie (odpowiedzieć EPSP lub IPSP, tj. przeprowadzić lub nie kontynuować dalej), ale zmienić liczbę receptorów lub syntetyzować białko receptorowe z nowymi właściwości w odniesieniu do pewnego mediatora. W związku z tym kolejna ważna właściwość synapsy chemicznej: dzięki procesom biochemicznym zachodzącym w cytoplazmie postsynaptycznej komórka przygotowuje się (uczy się) do przyszłych interakcji.
W układzie nerwowym działają różne synapsy, które różnią się mediatorami i receptorami. Nazwę synapsy określa mediator, a dokładniej nazwa receptora dla konkretnego mediatora. Zastanówmy się więc nad klasyfikacją głównych mediatorów i receptorów układu nerwowego (patrz także materiał rozpowszechniany na wykładzie!!).
Zauważyliśmy już, że o efekcie interakcji mediatora z receptorem decydują właściwości receptora. Zatem znane mediatory, z wyjątkiem kwasu g-aminomasłowego, mogą pełnić funkcje zarówno mediatorów pobudzających, jak i hamujących. struktura chemiczna Wyróżnia się następujące grupy mediatorów.
Acetylocholina, szeroko rozpowszechniony w ośrodkowym układzie nerwowym, jest mediatorem w synapsach cholinergicznych autonomicznego układu nerwowego, a także w somatycznych synapsach nerwowo-mięśniowych (ryc. 5).

Ryż. 5. Cząsteczka acetylocholiny
Znany dwa typy receptorów cholinergicznych: nikotyna ( Receptory H-cholinergiczne) i muskarynowe ( Receptory M-cholinergiczne). Nazwę nadano substancjom, które powodują w synapsach działanie podobne do acetylocholiny: N-cholinomimetyk Jest nikotyna, A M-cholinomimetyk- toksyna muchomora Amanita muscaria ( muskaryna). Bloker receptora H-cholinergicznego (antycholinergiczny) Jest d-tubokuraryna(główny składnik trucizny kurary) i M-antycholinergiczne jest toksyną belladonny Atropa belladonna – atropina. Co ciekawe, właściwości atropiny są znane od dawna i był czas, kiedy kobiety stosowały atropinę z wilczej jagody, aby wywołać rozszerzenie źrenic wzrokowych (aby oczy były ciemne i „piękne”).
Poniższe cztery główne mediatory mają podobieństwa w budowie chemicznej, dlatego są klasyfikowane jako monoaminy. Ten serotonina lub 5-hydroksytryptaminy (5-HT), odgrywa ważną rolę w mechanizmach wzmocnienia (hormon radości). Jest syntetyzowany z niezbędnego dla człowieka aminokwasu – tryptofanu (ryc. 6).

Ryż. 6. Cząsteczka serotoniny (5-hydroksytryptaminy).
Trzy inne mediatory są syntetyzowane z niezbędnego aminokwasu fenyloalaniny i dlatego są zjednoczone pod wspólną nazwą katecholaminy- Ten dopamina (dopamina), noradrenalina (noradrenalina) i adrenalina (epinefryna, ryc. 7).

Ryż. 7. Katecholaminy
Wśród aminokwasy mediatorami są m.in kwas gamma-aminomasłowy(g-AMK lub GABA – znany jako jedyny neuroprzekaźnik hamujący), glicyna, kwas glutaminowy, kwas asparaginowy.
Mediatorzy obejmują szereg peptydy. W 1931 roku Euler odkrył substancję w ekstraktach mózgu i jelit, która powoduje skurcz mięśni gładkich jelit i rozszerzenie naczyń krwionośnych. Przekaźnik ten został wyizolowany w czystej postaci z podwzgórza i otrzymał nazwę substancja P(od angielskiego proszek - proszek, składa się z 11 aminokwasów). Później ustalono, że substancja P odgrywa ważną rolę w przewodzeniu bolesnych pobudzeń (nazwy nie trzeba było zmieniać, ponieważ ból w języku angielskim to ból).
Peptyd snu Delta otrzymał swoją nazwę ze względu na zdolność do wywoływania powolnych rytmów o wysokiej amplitudzie (rytmy delta) w elektroencefalogramie.
W mózgu syntetyzowanych jest wiele mediatorów białkowych o charakterze narkotycznym (opium). Są to pentapeptydy Met-enkefalina I Leu-enkefalina, I endorfiny. Są to najważniejsze blokery pobudzeń bólowych i mediatory wzmocnienia (radości i przyjemności). Innymi słowy, nasz mózg jest wielką fabryką endogenny narkotyki. Najważniejsze jest nauczenie mózgu ich wytwarzania. "Jak?" - ty pytasz. To proste – endogenne opiaty powstają, gdy doświadczamy przyjemności. Rób wszystko z przyjemnością, zmuś swoją endogenną fabrykę do syntezy opiatów! Tę możliwość mamy naturalnie od urodzenia – zdecydowana większość neuronów reaguje na pozytywne wzmocnienie.
Badania ostatnich dziesięcioleci pozwoliły odkryć kolejnego bardzo interesującego mediatora – tlenek azotu (NO). Okazało się, że NO nie tylko odgrywa ważną rolę w regulacji napięcia naczyń krwionośnych (nitrogliceryna, którą znasz, jest źródłem NO i rozszerza naczynia wieńcowe), ale jest także syntetyzowany w neuronach centralnego układu nerwowego.
W zasadzie historia mediatorów jeszcze się nie skończyła, istnieje szereg substancji biorących udział w regulacji pobudzenia nerwowego. Tyle, że nie ustalono jeszcze dokładnie faktu ich syntezy w neuronach, nie odnaleziono ich w pęcherzykach synaptycznych, nie odnaleziono też specyficznych dla nich receptorów.







