
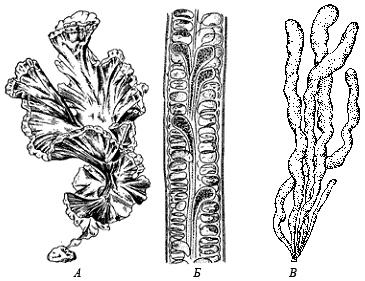
Czerwony - fikoerytryna , Pomarańczowy - karoten I " ksantofil”. Dodatkowe pigmenty wpływają na barwę alg i pełnią ważną funkcję systematyczną. Algi dzielą się na kilka rodzajów. Typowi przedstawiciele mikroskopijnych glonów pokazano na rysunku.
Mikroskopijne algi.
A. Okrzemki B. Algi sinicowe, C. Algi zielone (Chlamydomonas)
Zielone algi
Zielone algi są szeroko rozpowszechnione w wodach powierzchniowych. Wśród nich są formy jednokomórkowe, wielokomórkowe i kolonialne. Pigmenty skoncentrowane są w specjalnych ciałach plazmowych różne kształty– chromatofory. Rozmnażają się poprzez podział cytoplazmy w celu utworzenia komórek potomnych lub drogą płciową. Niektóre gatunki rozmnażają się poprzez wytwarzanie ruchliwych zarodników. Kolonie powstają w wyniku podziału bezpłciowego, w którym komórki potomne pozostają ze sobą powiązane. Komórki zielonych alg mają różne kształty: kulisty, owalny, w kształcie półksiężyca, trójkątny itp. Ich komórki zawierają organelle charakterystyczne dla komórek roślin wyższych. Jądro jest zróżnicowane. Skorupa składa się z celulozy. Cytoplazma może zawierać ziarna skrobi, która jest produktem fotosyntezy. Najczęstsze jednokomórkowe formy chlorelli ( Chlorella zwykła), Chlamydomonas ( Chlamidomony), z kolonialnych – Volvox ( Volvox aureus), gon ( Gonium piersiowy), z wielokomórkowego - ulothrix. Zielone algi spotykany w zbiornikach z czystą i brudną wodą, z wolnymi i szybkimi prądami, w różnych dziurach, kałużach wypełniających się po deszczu, a także na glebie.
Niebiesko-zielone algi
Niebiesko-zielone algi ( cyjanobakteria ) uważany jest za najstarszy z obecnie istniejących zakładów. Czy to ten lub Organizmy wielokomórkowe, najprościej zorganizowane, charakteryzują się specjalną strukturą komórkową. Nie ma typowego jądra i chromatoforów. Protoplazma niebiesko-zielonych alg jest zróżnicowana w obwodowo zabarwioną warstwę - chromatoplazma , I Środkowa część – centroplazma . Asymilujące pigmenty światłoczułe to chlorofil, fikocyna, fikoerytryna i karoten. W zależności od ilościowego stosunku pigmentów zmienia się również kolor komórek. Komórki zawierają specjalne ciała - endoplasty gęsta lub lepka konsystencja. W ściankach plazmatycznych komórek pomiędzy endoplastykami znajduje się „substancja chromatyny”, która barwi barwnikami jądrowymi. Komórkom niebiesko-zielonych alg brakuje wakuoli wypełnionych sokiem komórkowym. Pod tym względem podczas plazmolizy komórka całkowicie się kurczy. Komórki tych organizmów zawierają wakuole gazowe, które ułatwiają im unoszenie się na powierzchnię. Komórki niebiesko-zielonych alg mają błonę. Może być cienki i ledwo zauważalny lub pogrubiony. Błony komórkowe są często pokryte śluzem, co prowadzi do tworzenia się kolonii w wyniku zbrylania się tego śluzu. Skład muszli składa się głównie z pektyn. Z reguły kolonie nie mają określonego kształtu. W komórkach nitkowatych rzędy komórek są zamknięte w wydrążonej cylindrycznej osłonie, która pokrywa cały rząd komórek. Zbiór komórek z osłoną nazywa się włóknem. Komórki w tym samym włóknie mogą mieć taki sam lub różny rozmiar i kształt. Komórki włókna pokryte są na górze wspólną osłoną śluzową. U niektórych gatunków nici mogą się rozgałęziać. Często obserwuje się formację heterocysta znajduje się w wątku przez określoną liczbę komórek. Heterocysty powstają z komórek wegetatywnych, ale są znacznie większe. Mają gęstą skorupę, ale komunikują się z sąsiednimi komórkami przez pory. Uważa się, że heterocysty to wyspecjalizowane komórki, które wiążą azot.
Wiele gatunków cyjanobakterii jest zdolnych do tworzenia zarodników. U niektórych gatunków zarodniki, podobnie jak prawdziwe bakterie, są formą najbardziej odporną na niesprzyjające warunki. W tym przypadku z jednej komórki powstaje tylko jeden zarodnik. W przypadku innych cyjanobakterii zarodniki, podobnie jak grzyby, służą jako metoda rozmnażania. W tym przypadku wewnątrz komórki macierzystej powstaje wiele małych zarodników, które są uwalniane w przypadku pęknięcia błony. Niebiesko-zielone algi bardzo powszechny w przyrodzie: rozwijają się w zbiornikach wodnych o charakterze słonym i świeża woda, w glebie i na skałach, w Arktyce i na pustyni. Sprzyja temu wyjątkowa odporność na niekorzystne warunki i niewymagające wymagania dotyczące składników odżywczych.
Okrzemki
Okrzemki ( Okrzemka). Są to jednokomórkowe mikroskopijne organizmy. Niektóre gatunki tworzą kolonie w postaci nitek, wstążek i krzewów. Komórki mają wielkość od 4 do 1500 mikronów, a kolonie czasami osiągają kilka centymetrów. Komórki okrzemek zawierają utworzone jądro i chloroplasty. Te ostatnie oprócz chlorofilu zawierają brązowe pigmenty, dzięki czemu kolor alg jest żółtawy lub ciemnobrązowy. Komórki mają otoczkę pektynową i otoczkę składającą się z krzemionki. Błona komórkowa składa się z dwóch połówek, które nie rosną razem i mogą się od siebie oddalać. Protoplazma znajduje się w cienkiej warstwie wzdłuż ścian, tworząc u wielu gatunków mostek protoplazmatyczny w środku komórki, pozostała część przestrzeni komórkowej jest wypełniona sokiem komórkowym, jest tylko jedno jądro. Chromatofory różnią się kształtem. Produktami asymilacji są olej, wolutyna, leukozyna. Rozmnażać się prosty podział i za pomocą sporów. Podczas podziału wegetatywnego każda część otrzymuje zastawkę matczyną, a brakująca część odrasta na nowo w trakcie rozwoju komórki. Charakterystyczną cechą gatunku jest struktura otoczki krzemowej. Grupa okrzemek Kary spotykany głównie wśród zanieczyszczeń obiektów dennych i w glebie.
Zielonki są najbardziej rozległym ze wszystkich działów numerycznych glonów różne szacunki od 4 do 13 - 20 tysięcy gatunków. Oni wszyscy mają zielony kolor thalli, co wynika z przewagi chlorofilu w chloroplastach A I B nad innymi pigmentami. Komórki niektórych przedstawicieli zielonych alg ( Chlamydomonas, Trentepolia, Hematococcus) mają barwę czerwoną lub pomarańczową, co jest związane z gromadzeniem się pigmentów karotenoidowych i ich pochodnych poza chloroplastem.
Morfologicznie są bardzo zróżnicowane. Wśród zielonych alg są przedstawiciele jednokomórkowi, kolonialni, wielokomórkowi i niekomórkowi, aktywnie mobilni i nieruchomi, przywiązani i wolno żyjący. Rozpiętość ich rozmiarów jest również niezwykle duża – od kilku mikrometrów (co jest porównywalne wielkością z komórkami bakteryjnymi) do 1–2 metrów.
Komórki są jednojądrzaste lub wielojądrowe, z jednym lub większą liczbą chromatoforów zawierających chlorofil i karotenoidy. Chloroplasty są pokryte dwiema błonami i zwykle mają piętno, czyli ocellus, filtr przewodzący światło niebieskie i zielone do fotoreceptora. Oko składa się z kilku rzędów kuleczek lipidowych. Tylakoidy - struktury, w których zlokalizowane są pigmenty fotosyntetyczne - gromadzą się w stosy (lamele) po 2–6. W strefie przejściowej wici znajduje się formacja gwiaździsta. Najczęściej występują dwie wici. Głównym składnikiem ściany komórkowej jest celuloza.
Chlorofity mają różne rodzaje odżywiania: fototroficzne, miksotroficzne i heterotroficzne. Rezerwowy polisacharyd zielonych alg, skrobia, osadza się wewnątrz chloroplastu. Chlorofity mogą również akumulować lipidy, które odkładają się w postaci kropelek w zrębie chloroplastów i cytoplazmie.
Plechy wielokomórkowe nitkowate, rurkowate, blaszkowate, krzaczaste lub o innej strukturze oraz różne kształty. Ze znanych typów organizacji plechy u zielonych alg nieobecny jest tylko typ ameboidalny.
Są szeroko dystrybuowane w postaci świeżej i wody morskie, w glebie i siedliskach lądowych (na glebie, skałach, korze drzew, ścianach domów itp.). Około 1/10 ogólnej liczby gatunków występuje w morzach, zwykle rosnąc w górnych warstwach wody do 20 m. Wśród nich występują formy planktonowe, peryfitoniczne i bentosowe. Innymi słowy, zielone algi opanowały trzy główne siedliska organizmów żywych: woda – ziemia – powietrze.
Zielone algi wykazują fototaksję dodatnią (ruch w kierunku źródła światła) i ujemną (ruch od jasnego źródła światła). Oprócz intensywności światła temperatura wpływa na fototaksję. Zoospory gatunków z tego rodzaju wykazują pozytywną fototaksję w temperaturze 160°C Hematococcus, Ulothrix, Ulva, a także niektóre rodzaje glonów desmidianowych, w których ruch komórek odbywa się poprzez wydzielanie śluzu przez pory w skorupce.
Reprodukcja. Zielone glony charakteryzują się występowaniem wszystkich znanych metod rozmnażania: wegetatywnego, bezpłciowego i płciowego .
Rozmnażanie wegetatywne w postaciach jednokomórkowych komórka dzieli się na pół. Kolonialne i wielokomórkowe formy chlorofitu rozmnażają się przez części ciała (talus lub plecha).
Rozmnażanie bezpłciowe w zielonych algach jest szeroko reprezentowany. Częściej dokonują tego ruchliwe zoospory, rzadziej nieruchome aplanospory i hipnospory. Komórki, w których tworzą się zarodniki (sporangia), w większości przypadków nie różnią się od pozostałych komórek wegetatywnych plechy, rzadziej mają inny kształt i większe rozmiary. Tworzące się zoospory mogą być nagie lub pokryte sztywną ścianą komórkową. Liczba wici w zoosporach waha się od 2 do 120. Zoospory mają różne kształty: kulisty, elipsoidalny lub gruszkowy, jednojądrowy, bez oddzielnej otoczki, z 2–4 wiciami na przednim, bardziej spiczastym końcu i chloroplastem na rozszerzonym tylny koniec. Zwykle mają pulsujące wakuole i piętno. Zoospory powstają pojedynczo lub częściej z kilku z wewnętrznej zawartości komórki macierzystej, wychodzą przez okrągły lub szczelinowy otwór utworzony w skorupce, rzadziej w wyniku jej ogólnego śluzu. W momencie wyjścia z komórki macierzystej zoospory są czasami otoczone cienkim pęcherzem śluzowym, który wkrótce się rozpuszcza (rodzaj Ulotrix).
U wielu gatunków zamiast zoospor lub wraz z nimi powstają nieruchome zarodniki - aplanospory. Aplanospory to zarodniki rozmnażające się bezpłciowo, które nie mają wici, ale mają kurczliwe wakuole. Aplanospory uważa się za komórki, w których zawieszony jest dalszy rozwój zoospor. Powstają również z protoplastu komórki, jednego lub więcej, ale nie wytwarzają wici, ale po przyjęciu kulistego kształtu są pokryte własną skorupą, w tworzeniu której skorupa komórki macierzystej nie uczestniczy. Aplanospory są uwalniane w wyniku pęknięcia błon śluzowych komórek macierzystych i kiełkują po pewnym okresie spoczynku. Aplanospory o bardzo grubych błonach nazywane są hipnosporami. Przeważnie przejmują funkcję fazy spoczynkowej. Autospory, które są mniejszymi kopiami nieruchliwych komórek wegetatywnych, nie mają kurczliwych wakuoli. Powstawanie autosporów koreluje z pokonaniem warunków lądowych, w których woda nie zawsze może być obecna w wystarczającej ilości.
Rozmnażanie płciowe przeprowadzana przez gamety powstałe w niezmienionych, nieznacznie zmienionych lub znacząco przekształconych komórkach - gametangia. Ruchliwe gamety o strukturze monadycznej, dwuwiciowe. Proces seksualny w zielonych algach jest reprezentowany przez różne formy: hologamię, koniugację, izogamię, heterogamię, oogamię. W przypadku izogamii gamety są morfologicznie całkowicie do siebie podobne, a różnice między nimi są czysto fizjologiczne. Zygota pokryta jest grubą skorupą, często z rzeźbionymi naroślami, zawiera dużą ilość substancji rezerwowych i kiełkuje natychmiast lub po pewnym okresie spoczynku. Podczas kiełkowania zawartość zygoty u większości gatunków dzieli się na cztery części, które wyłaniają się z muszli i wyrastają na nowe osobniki. Znacznie rzadziej gamety rozwijają się w nowy organizm bez fuzji, samodzielnie, bez tworzenia zygoty. Ten rodzaj reprodukcji nazywa się partenogeneza i zarodniki utworzone z poszczególnych gamet partenospory.
W heterogamii obie gamety różnią się od siebie wielkością, a czasem kształtem. Gamety większe, często mniej mobilne, uważa się za żeńskie, mniejsze i bardziej mobilne – męskie. W niektórych przypadkach różnice te są niewielkie, a wtedy mówią po prostu o heterogamii, w innych są bardzo znaczące.
Jeśli gameta żeńska jest nieruchoma i przypomina bardziej jajo, wówczas ruchliwa gameta męska staje się plemnikiem, a proces seksualny nazywa się oogamią. Nazywa się gametangie, w których powstają jaja oogonia, Różnią się od komórek wegetatywnych zarówno kształtem, jak i rozmiarem. Gametangie, w których powstają plemniki, nazywane są gametangiami pylniki. Zygota powstała w wyniku zapłodnienia komórki jajowej przez plemnik tworzy grubą skorupę i nazywa się oospora.
W typowej oogamii jaja są duże, nieruchome i najczęściej rozwijają się pojedynczo w oogonii; plemniki są małe, ruchliwe i powstają w dużych ilościach w antheridium. Na jednym osobniku mogą rozwinąć się oogonia i antheridia, w tym przypadku glony są jednopienne; jeśli rozwiną się na różnych osobnikach, są dwupienne. Zapłodnione jajo pokryte jest grubą brązową skorupą; Często sąsiadujące z nim komórki wytwarzają krótkie gałęzie, które przerastają oosporę, oplatając ją jednowarstwową korą.
Cykle życia. Większość przedstawicieli glonów zielonych ma haplobiontowy cykl życiowy z redukcją zygotyczną. U takich gatunków jedynie zygota jest stadium diploidalnym – komórką powstałą w wyniku zapłodnienia komórki jajowej przez plemnik. Inny typ cyklu życiowego – haplodiplobiont z redukcją sporadyczną – występuje u Ulvoceae, Cladophoraceae i niektórych Trentepoliaceae. Glony te charakteryzują się naprzemiennością diploidalnego sporofitu i haploidalnego gametofitu. Cykl życiowy haplodiplobiontów z redukcją somatyczną jest znany tylko w Prasiole. Kwestionuje się obecność diplobiontowego cyklu życiowego u Bryopsidae i Dasycladiaceae.
U niektórych Ulothrixidae ten sam osobnik może dać początek zarówno zoosporom, jak i gametom. W innych przypadkach zoospory i gamety powstają u różnych osobników, tj. Cykl życiowy glonów obejmuje zarówno formy rozwoju płciowe (gametofit), jak i bezpłciowe (sporofity). Sporofit jest zwykle diploidalny, tj. ma w komórkach podwójny zestaw chromosomów, gametofit jest haploidalny, tj. ma pojedynczy zestaw chromosomów. Obserwuje się to w przypadkach, gdy mejoza zachodzi podczas tworzenia zarodników (redukcja sporyczna), a część cyklu życiowego glonów od zygoty do powstania zarodników odbywa się w diplofazie, a część od zarodnika do tworzenia gamet w haplofaza. Ten cykl rozwojowy jest typowy dla gatunków z rodzaju Ulva.
W glonach Ulothrix powszechna jest redukcja zygotyczna, gdy podczas kiełkowania zygoty następuje mejoza. W tym przypadku tylko zygota jest diploidalna, reszta cyklu życiowego zachodzi w haplofazie. Redukcja gamet występuje znacznie rzadziej, gdy podczas tworzenia gamet zachodzi mejoza. W tym przypadku tylko gamety są haploidalne, a reszta cyklu jest diploidalna.
Taksonomia
Nadal nie ma jednego ustalonego systemu zielonych alg, zwłaszcza jeśli chodzi o grupowanie rzędów w różne proponowane klasy. Przez bardzo długi czas przy rozróżnianiu rzędów u zielenic główne znaczenie przywiązywano do rodzaju zróżnicowania plechy. Jednak ostatnio, w związku z nagromadzeniem danych na temat cech ultrastrukturalnych komórek wiciowych, rodzaju mitozy i cytokinezy itp., Niejednorodność wielu z tych rzędów jest oczywista.
Dział obejmuje 5 klas: Ulvophyceae – Ulvophyceae, Brypsodaceae – Bryopsidophyceae, Chlorophyceae – Chlorophyceae, Trebouxiophyceae, Prasinophyceae – Prasinophyceae.
Klasa Ulvophyceae –Ulvophyceae
Znanych jest około 1 tysiąca gatunków. Nazwa klasy pochodzi od rodzaju typu Ulwa. Obejmuje gatunki z plechą nitkowatą i blaszkowatą. Cykle życiowe są zróżnicowane. Gatunki są przeważnie morskie, rzadziej słodkowodne i lądowe. Niektóre są częścią porostów. U przedstawicieli morskich w ścianach komórkowych może osadzać się wapno.
Zamów Ulotrix –Ulotrichales.
Rodzaj Ulotrix(ryc. 54). Rodzaje Ulotrix Częściej żyją w wodach słodkich, rzadziej w morzach, zbiornikach słonawych i w glebie. Przywiązują się do obiektów podwodnych, tworząc jasnozielone krzewy o wielkości do 10 cm lub większej. Nierozgałęzione wątki Ulotrix, składające się z pojedynczego rzędu cylindrycznych komórek z grubymi błonami celulozowymi, są przymocowane do podłoża za pomocą bezbarwnej stożkowej komórki podstawnej, która pełni funkcję ryzoidu. Charakterystyczna jest budowa chromatoforu, który ma postać płytki ściennej tworzącej otwarty pas lub pierścień (cylinder).
Ryż. 54. UlothricC (by:): 1 – plecha nitkowata, 2 – zoospora, 3 – gameta, 4 – kopulacja gamet
Rozmnażanie bezpłciowe Ulotrix można przeprowadzić na dwa sposoby: poprzez rozpad włókna na krótkie odcinki, z których powstaje nowe włókno, lub poprzez utworzenie w komórkach zoospor czterowiciowych. Zoospory wyłaniają się z komórki macierzystej, zrzucają jedną po drugiej wici, przyczepiają się bokiem do podłoża, pokrywają się cienką błoną celulozową i kiełkują, tworząc nowe włókno. Proces seksualny jest izogamiczny. Po zapłodnieniu zygota najpierw unosi się na wodzie, następnie osiada na dnie, traci wici, tworzy gęstą skorupę i śluzową łodygę, którymi przyczepia się do podłoża. To sporofit spoczynkowy. Po okresie spoczynku następuje podział jądra redukcyjnego i zygota kiełkuje w postaci zoospor. Tak w cyklu życia Ulotrix następuje przemiana pokoleń lub zmiana seksualnych i bezpłciowych form rozwoju: nitkowaty wielokomórkowy gametofit (pokolenie tworzące gamety) zostaje zastąpiony jednokomórkowym sporofitem - pokoleniem reprezentowanym przez rodzaj zygoty na łodydze i jest zdolny do tworzenia zarodników.
Zamawiaj Ulvaceae -Ulvales. Mają blaszkowatą, workowatą, rurkowatą lub rzadko nitkowatą plechę w różnych odcieniach zieleni. Krawędzie płytek mogą być faliste lub złożone, a do mocowania do podłoża są wyposażone w krótką łodygę lub podstawę z małym dyskiem podstawowym. Gatunki morskie i słodkowodne. Najczęstszymi gatunkami rodzajów w wodach przybrzeżnych mórz Dalekiego Wschodu są Ulva, Monostroma, Cornmannia I Ulwaria.
Rodzaj Ulwa(ryc. 55). Plecha to jasnozielona lub jasnozielona, cienka dwuwarstwowa, często perforowana płytka lub jednowarstwowa pusta rurka, przymocowana do podłoża za pomocą podstawy zwężonej w krótki ogonek.
Ryż. 55. Ulwa: A- wygląd Ulva fenestrowana, B– przekrój wzgórza, W- wygląd Ulwa jelitowa
Zmiana form rozwoju w cyklu życia Ulwa redukuje się do izomorficznego, gdy etap bezpłciowy (sporofit) i etap płciowy (gametofit) są do siebie morfologicznie podobne, i heteromorficzny, gdy są morfologicznie różne. Gametofit jest wielokomórkowy, blaszkowy, sporofit jest jednokomórkowy. Gametofity wytwarzają gamety dwuwiciowe, a sporofity wytwarzają zoospory czterowiciowe.
Gatunki z rodzaju występują w morzach wszystkich stref klimatycznych, chociaż preferują ciepłe wody. Na przykład w płytkich wodach Morza Czarnego i Morza Japońskiego Ulva jest jednym z najliczniejszych rodzajów glonów. Dużo typów Ulwa tolerować odsalanie wody; często można je spotkać w ujściach rzek.
Klasa Bryopsidae–Bryopsidophyceae
Znanych jest około 500 gatunków. Plecha jest bezkomórkowa. Tworzą je proste lub splecione nitki syfonu, tworzące złożone struktury. Plecha w postaci bąbelków, krzewów, gąbczastych, dychotomicznie rozgałęzionych krzewów. Plecha jest podzielona na segmenty, co symuluje wielokomórkowość, składającą się z kilku lub wielu komórek jądrowych. Nici i krzewy wszystkich odcieni zieleni lub brązu.
Zamów Bryopsidae– Bryopsidales
Większość gatunków występuje w wodach słodkich i słonawych. Część z nich rośnie na ziemi, na kamieniach, piasku, czasem na słonych bagnach.
Rodzaj Bryopsis– nitkowate krzewy dorastające do 6-8 cm wys., pierzasto lub nieregularnie rozgałęzione, górne gałęzie ze zwężeniem u nasady. Plecha ma syfoniczną strukturę niekomórkową. Rośnie w pojedynczych krzakach lub małych kępach w strefie przybrzeżnej, żyje w morzach ciepłych i umiarkowanych (Załącznik 7B).
Rodzaj Kod– sznurkowate, dychotomicznie rozgałęzione krzewy o wysokości 10–20 cm, gąbczaste. miękkie, mocowane na podeszwie w kształcie krążka. Wewnętrzną część wzgórza tworzą kompleksowo splecione nici syfonu. Rośnie na glebach miękkich i twardych w strefie sublitoralnej do głębokości 20 m w pojedynczych roślinach lub w małych grupach (załącznik 7A, B).
Rodzaj Caulerpa obejmuje około 60 gatunków wodorostów, pełzające części plechy rozłożone na ziemi mają postać rozgałęzionych cylindrów, osiągających długość kilkudziesięciu centymetrów. W pewnych odstępach w dół od nich wychodzą obficie rozgałęzione ryzoidy, zakotwiczające roślinę w glebie, a w górę - płaskie, liściaste pionowe pędy, w których skupiają się chloroplasty.

Ryż. 56. Caulerpa: A – wygląd plechy; B – przekrój plechy z belkami celulozowymi
Plecha caulerpa, pomimo swoich dużych rozmiarów, nie ma budowy komórkowej – całkowicie pozbawiona jest przegród poprzecznych i formalnie reprezentuje jedną komórkę olbrzymią (ryc. 56). Ta struktura wzgórza nazywa się syfon. Wewnątrz wzgórza caulerpa znajduje się centralna wakuola otoczona warstwą cytoplazmy zawierającą liczne jądra i chloroplasty. Różne części wzgórza wyrastają na końcach, gdzie gromadzi się cytoplazma. Centralną wnękę we wszystkich częściach plechy przecinają cylindryczne pasma szkieletowe - belki celulozowe, które nadają ciału glonów wytrzymałość mechaniczną.
Caulerpa łatwo rozmnaża się wegetatywnie: gdy obumierają starsze części plechy, jej poszczególne odcinki z pionowymi pędami stają się niezależnymi roślinami. Gatunki tego rodzaju żyją głównie w morzach tropikalnych, a tylko nieliczne wkraczają na szerokości subtropikalne, na przykład powszechne w Morzu Śródziemnym Kiełkuje Caulerpa. Glony te preferują płytkie, spokojne wody, na przykład laguny chronione przed działaniem ciągłych fal przez rafy koralowe i osiadają na różnych twardych podłożach - kamieniach, rafach, skałach, glebie piaszczystej i błotnistej.
Klasa Chlorophyceae–Chlorophyceae
Znanych jest około 2,5 tysiąca gatunków. Plecha jest jednokomórkowa lub kolonialna monnadyczna, wolno żyjąca.
Zamów Volvoxidae -Volvocales.
Rodzaj Chlamydomonas(ryc. 57) obejmuje ponad 500 gatunków jednokomórkowych glonów żyjących w świeżych, płytkich, dobrze nagrzanych i zanieczyszczonych zbiornikach wodnych: stawach, kałużach, rowach itp. Kiedy rozmnażają się masowo, woda zmienia kolor na zielony. Chlamydomonasżyje także na ziemi i śniegu. Jego ciało jest owalne, gruszkowate lub kuliste. Komórka pokryta jest gęstą skorupą, często pozostającą w tyle za protoplastem, z dwiema identycznymi wiciami na przednim końcu; z ich pomocą Chlamydomonas aktywnie porusza się w wodzie. Protoplast zawiera 1 jądro, chromatofor w kształcie miseczki, piętno i pulsujące wakuole.

Ryż. 57. Budowa i rozwój Chlamydomonas: A – osobnik wegetatywny; B – stadium palmelle; B – rozmnażanie (młode osobniki wewnątrz komórki macierzystej)
Chlamydomonas rozmnażają się głównie bezpłciowo. Kiedy zbiornik wyschnie, rozmnażają się, dzieląc komórkę na pół. Komórki zatrzymują się, tracą wici, ich ściany komórkowe stają się śluzem i w tym nieruchomym stanie komórki zaczynają się dzielić. Ściany powstałych komórek potomnych zawierają także śluz, dzięki czemu ostatecznie tworzy się system zagnieżdżonych w sobie błon śluzowych, w których grupami rozmieszczone są nieruchome komórki. To jest palmelle stan glonów. Kiedy dostaną się do wody, komórki ponownie tworzą wici, opuszczają komórkę macierzystą w postaci zoospor i przechodzą w samotny stan monadyczny.
W sprzyjających warunkach Chlamydomonas rozmnaża się intensywnie w inny sposób - komórka zatrzymuje się, a jej protoplast nieco za ścianą dzieli się sukcesywnie wzdłużnie na dwie, cztery lub osiem części. Te komórki potomne tworzą wici i pojawiają się jako zoospory, które wkrótce zaczynają się ponownie rozmnażać.
Proces rozrodczy u Chlamydomonas jest izogamiczny lub oogamiczny. Mniejsze gamety powstają wewnątrz komórki macierzystej w taki sam sposób jak zoospory, ale w większej liczbie (16, 32 lub 64). Zapłodnienie następuje w wodzie. Zapłodnione jajo zostaje przykryte wielowarstwową membraną i osiada na dnie zbiornika. Po okresie spoczynku zygota dzieli się mejotycznie, tworząc 4 osobniki haploidalnej córki Chlamydomonas.
Rodzaj Volvox- najbardziej zorganizowani przedstawiciele zakonu tworzą gigantyczne kolonie składające się z setek i tysięcy komórek. Kolonie mają postać kulek śluzowych o średnicy do 2 mm, w warstwie obwodowej których znajduje się do 50 tysięcy komórek z wiciami, zrośniętych ze sobą bocznymi ścianami śluzowymi i połączonych plazmodesmami (ryc. 58). Wnęka wewnętrzna

Ryż. 58. Wygląd Kolonie Volvox
Kulka wypełniona jest płynnym śluzem. W kolonii istnieje specjalizacja komórek: jej obwodową część tworzą komórki wegetatywne, a pomiędzy nimi rozproszone są większe komórki rozrodcze.
Około tuzina komórek kolonii to gonidia, komórki rozmnażania bezpłciowego. W wyniku powtarzających się podziałów dają początek młodym koloniom potomnym, które wpadają do kuli matki i uwalniają się dopiero po jej zniszczeniu. Proces seksualny to oogamia. Oogonia i antheridia powstają również z komórek rozrodczych. Kolonie są jednopienne i dwupienne. Gatunki z rodzaju spotykane są w stawach i starorzeczach rzek, gdzie w okresie intensywnego rozmnażania powodują „zakwitanie” wody.
Klasa Trebuxiaceae –Trebouxiophyceae
Klasa nazwana na cześć rodzaju typu Trebuksja. Obejmuje głównie jednokomórkowe formy kokosowe. Są przedstawiciele sarcinoidów i nitkowatych. Formy słodkowodne i lądowe, rzadziej morskie, wiele z nich tworzy symbiozy. Około 170 gatunków.
Zamów chlorellę -Chlorele. Jednoczy przedstawicieli autospor kokoidowych.
Rodzaj Chlorella- algi jednokomórkowe w postaci stacjonarnej kuli. Komórka pokryta jest gładką skorupą; zawiera jedno jądro i ścianę, cały, wycięty lub płatkowany chromatofor z pirenoidem. Ściana komórkowa wielu gatunków, wraz z celulozą, zawiera sporopoleninę, substancję niezwykle odporną na działanie różnych enzymów, która występuje także w ziarnach pyłku i zarodnikach roślin wyższych. Chlorella rozmnaża się bezpłciowo, tworząc do 64 nieruchomych autospor. Nie ma rozmnażania płciowego. Chlorella szeroko rozpowszechniony w różnych zbiornikach wodnych, spotykany na wilgotnej glebie, korze drzew i części porostów.
Zamawiaj Trebuxiaceae - Trebouxiales . Obejmuje rodzaje i gatunki zaliczane do porostów.
Rodzaj Trebuksja- algi jednokomórkowe. Komórki kuliste mają pojedynczy osiowy chloroplast gwiaździsty z pojedynczym pyrenoidem. Rozmnażanie bezpłciowe odbywa się za pomocą nagich zoospor. Występuje w postaci wolno żyjącej w siedliskach lądowych (na korze drzew) lub jako fotobiont porostów.
Zajęcia Prazyne –Prasinophyceae
Nazwa klasy pochodzi z języka greckiego. prasino - zielony. Wiciowce lub rzadziej jednokomórkowe organizmy kokosowe lub palmellooidalne.
Zamów Pyramidonidae - Piramimonady. Komórki mają 4 lub więcej wici i trzy warstwy łusek. Mitoza jest otwarta, wrzeciono pozostaje w telofazie, cytokineza zachodzi w wyniku tworzenia się bruzdy szczelinowej.
Rodzaj Piramimony– organizmy jednokomórkowe (ryc. 59). Z przedniego końca komórki znajduje się 4–16 wici, które mogą być pięciokrotnie dłuższe od komórki. Chloroplast jest zwykle pojedynczy, z jednym pirenoidem i jednym lub większą liczbą oczek. Komórki i wici pokryte są kilkoma warstwami łusek. Szeroko rozpowszechniony w wodach słodkich, słonawych i morskich. Występują w planktonie i bentosie i mogą powodować zakwity wody.

Ryż. 59. Wygląd glonów Piramimony
Zamów Chlorodendraceae– Chlorodendrale. Komórki są skompresowane, z czterema wiciami, pokryte osłonką, mitoza jest zamknięta, zachodzi cytokineza w wyniku utworzenia bruzdy podziału.
Rodzaj Tetraselmis mogą występować jako ruchliwe komórki czterowiciowe lub jako komórki nieruchliwe przyczepione łodygami śluzowymi. Komórki pokryte są osłonką. Kiedy komórki dzielą się, wokół każdej komórki potomnej tworzy się nowa osłonka w osłonce komórki macierzystej. Na przednim końcu komórki wici wychodzą przez otwór w osłonce, który jest pokryty włoskami i łuskami. Jest jeden chloroplast z podstawowym pirenodem. Komórki są zwykle zielone, ale czasami zmieniają kolor na czerwony w wyniku nagromadzenia karotenoidów. Przedstawiciele morscy mogą żyć w płazińcach morskich.
Ekologia i znaczenie
Zielone algi są szeroko rozpowszechnione na całym świecie. Większość z nich można znaleźć w zbiornikach słodkowodnych, ale istnieje wiele form słonawych i morskich. Nitkowate zielone algi, przyczepione lub niezwiązane, wraz z okrzemkami i niebieskozieleniną, są dominującymi glonami bentosowymi w kontynentalnych zbiornikach wodnych. Występują w zbiornikach wodnych o różnym poziomie troficznym (od dystroficznych do eutroficznych) i o różnej zawartości materia organiczna(od kseno- do polisaprobowych), jony wodoru (od zasadowego do kwaśnego), w różnych temperaturach (termo-, mezo- i kriofile).
Wśród zielonych glonów występują formy planktonowe, peryfitoniczne i bentosowe. W grupie pikoplanktonu morskiego, algi prasinowe Ostreokok uważana za najmniejszą wolno żyjącą komórkę eukariotyczną. Istnieją gatunki zielonych alg, które przystosowały się do życia w glebie i siedliskach lądowych. Można je znaleźć na korze drzew, skałach, różnych budynkach, na powierzchni gleby i w powietrzu. Przedstawiciele rodzajów są szczególnie pospoliti w tych siedliskach Trentepoly I Trebuksja. Zielone algi rosną w gorących źródłach o temperaturze 35–52°C, a w niektórych przypadkach nawet do 84°C i wyższej, często ze zwiększoną zawartością soli mineralnych lub substancji organicznych (silnie zanieczyszczone gorące ścieki z fabryk, fabryk, elektrowni lub elektrownie jądrowe). Przeważają także wśród gatunków glonów kriofilnych. Mogą powodować zielone, żółte, niebieskie, czerwone, brązowe, brązowe lub czarne „zakwity” śniegu lub lodu. Glony te występują w powierzchniowych warstwach śniegu lub lodu i intensywnie namnażają się w stopionej wodzie o temperaturze około 0°C. Tylko kilka gatunków przechodzi stadia spoczynku, podczas gdy większości brakuje żadnych specjalnych adaptacji morfologicznych do niskich temperatur.
W zbiornikach zasolonych dominują jednokomórkowe, mobilne algi zielone - hiperhaloby, których komórki nie mają błony i są otoczone jedynie plazmalemą. Glony te charakteryzują się zwiększoną zawartością chlorku sodu w protoplazmie, wysokim wewnątrzkomórkowym ciśnieniem osmotycznym, akumulacją karotenoidów i gliceryny w komórkach oraz dużą labilnością układów enzymatycznych i procesów metabolicznych. W słonych zbiornikach wodnych często rozwijają się w dużych ilościach, powodując czerwone lub zielone „kwitnienie” słonych zbiorników wodnych.
Mikroskopijne jednokomórkowe, kolonialne i nitkowate formy zielonych alg przystosowały się do niesprzyjających warunków życia w powietrzu. W zależności od stopnia zawilgocenia dzieli się je na 2 grupy: glony powietrzne, które żyją w warunkach wyłącznie wilgoci atmosferycznej i dlatego podlegają ciągłej zmianie wilgotności i suszeniu; algi wodne narażone na ciągłe nawadnianie wodą (pod strumieniem wodospadu, falami itp.). Warunki istnienia glonów w zbiorowiskach aerofilnych są bardzo wyjątkowe i charakteryzują się przede wszystkim częstymi i gwałtownymi zmianami dwóch czynników - wilgotności i temperatury.
W warstwie gleby żyją setki gatunków zielonych alg. Gleba jako biotop przypomina siedliska wodne i powietrzne: zawiera powietrze, ale jest nasycona parą wodną, co zapewnia oddychanie powietrzem atmosferycznym bez zagrożenia wysychaniem. Intensywny rozwój glonów jako organizmów fototroficznych możliwy jest jedynie w granicach przenikania światła. Na glebach dziewiczych jest to wierzchnia warstwa gleby o grubości do 1 cm, na glebach uprawnych jest ona nieco grubsza. Jednak w grubości gleby, gdzie światło nie przenika, żywe glony znajdują się na głębokości do 2 m na glebach dziewiczych i do 3 m na glebach ornych. Wyjaśnia to zdolność niektórych glonów do przejścia na odżywianie heterotroficzne w ciemności. Wiele glonów pozostaje w glebie w stanie uśpionym.
Aby utrzymać swoje funkcje życiowe, algi glebowe mają pewne cechy morfologiczne i fizjologiczne. Należą do nich stosunkowo niewielki rozmiar gatunków glebowych, a także zdolność do wytwarzania obfitego śluzu – śluzowatych kolonii, osłon i zawijasów. Dzięki obecności śluzu glony szybko wchłaniają wodę po zwilżeniu i magazynują ją, spowalniając wysychanie. Charakterystyczną cechą glonów glebowych jest „efemeryczność” ich okresu wegetacyjnego - zdolność szybkiego przejścia ze stanu spoczynku do aktywnego życia i odwrotnie. Są również w stanie tolerować różne wahania temperatury gleby. Zakres przeżycia wielu gatunków mieści się w zakresie od -200 do +84°C i więcej. Glony lądowe stanowią ważną część roślinności Antarktydy. Są prawie czarne, więc ich temperatura ciała jest wyższa niż środowisko. Glony glebowe są także ważnym składnikiem biocenoz strefy suchej (suchej), gdzie latem gleba nagrzewa się do 60–80°C. Ciemne osłonki śluzowe wokół komórek stanowią ochronę przed nadmiernym nasłonecznieniem.
Wyjątkową grupę stanowią algi endolitofilne związane z podłożem wapiennym. Po pierwsze, są to nudne glony. Na przykład glony z rodzaju Gomontia Wwiercają się w skorupy jęczmienia perłowego i bezzębnych chrząszczy oraz penetrują wapienne podłoże zbiorników słodkowodnych. Sprawiają, że podłoże wapienne staje się luźne, podatne na działanie różnych czynników chemicznych i fizycznych. Po drugie, wiele glonów występujących w wodach słodkich i morskich ma zdolność przekształcania soli wapnia rozpuszczonych w wodzie w nierozpuszczalne i osadzania ich na plechach. Szereg tropikalnych zielonych alg, m.in. Galimeda, odkłada węglan wapnia w plechy. Biorą czynny udział w budowie raf. Gigantyczne złoża szczątków Halimedy, czasami osiągające 50 m wysokości, występują w wodach szelfu kontynentalnego związanych z Wielką Rafą Koralową w Australii i innych regionach, na głębokościach od 12 do 100 m.
Zielone algi Trebuxia, wchodzące w symbiotyczny związek z grzybami, są częścią porostów. Około 85% porostów zawiera jednokomórkowe i nitkowate zielone algi jako fotobionty, 10% zawiera sinice, a 4% (lub więcej) zawiera zarówno niebiesko-zielone, jak i zielone algi. Występują jako endosymbionty w komórkach pierwotniaków, alg kryptofitowych, hydr, gąbek i niektórych płazińce. Nawet chloroplasty poszczególnych glonów syfonowych, m.in. Kod, stają się symbiontami ślimaków nagoskrzelnych. Zwierzęta te żywią się glonami, których chloroplasty pozostają żywotne w komórkach jamy oddechowej, a przy świetle bardzo skutecznie fotosyntetyzują. Na sierści ssaków rozwijają się liczne glony zielone. Endosymbionty podlegające zmianom morfologicznym w porównaniu z przedstawicielami wolno żyjącymi nie tracą zdolności do fotosyntezy i rozmnażania się wewnątrz komórek gospodarza.
Znaczenie gospodarcze. Powszechne występowanie glonów zielonych decyduje o ich ogromnym znaczeniu w biosferze i działalności gospodarczej człowieka. Ze względu na zdolność do fotosyntezy tak jest główni producenci ogromna ilość materia organiczna w zbiornikach wodnych, które są szeroko stosowane przez zwierzęta i ludzi. Pochłaniając z wody dwutlenek węgla, zielone algi nasycają ją tlenem, niezbędnym wszystkim organizmom żywym. Ich rola w biologicznym cyklu substancji jest ogromna. Szybkie rozmnażanie i bardzo wysokie tempo asymilacji (około 3-5 razy wyższe niż u roślin lądowych) powodują, że masa glonów zwiększa się ponad 10 razy dziennie. Jednocześnie w komórkach chlorelli gromadzą się węglowodany (w szczepach selekcji ich zawartość sięga 60%), lipidy (do 85%), witaminy B, C i K. Białko chlorelli, które może stanowić nawet 50% suchej masy masa komórki, zawiera wszystkie niezbędne aminokwasy. Unikalna zdolność gatunku Chlorella Przyswojenie od 10 do 18% energii świetlnej (w porównaniu z 1–2% w roślinach lądowych) pozwala na wykorzystanie tej zielonej algi do regeneracji powietrza w zamkniętych biologicznych systemach podtrzymywania życia człowieka podczas długotrwałych lotów kosmicznych i nurkowania.
Wiele gatunków zielonych alg wykorzystuje się jako organizmy wskaźnikowe w systemie monitorowania ekosystemów wodnych. Oprócz fototroficznego sposobu odżywiania wiele jednokomórkowych zielonych alg (chlamydomonas) jest w stanie wchłaniać przez skorupę substancje organiczne rozpuszczone w wodzie, co przyczynia się do aktywnego oczyszczania zanieczyszczonych wód, w których rozwijają się te gatunki. Dlatego się je wykorzystuje Dla czyszczenie i obróbka końcowa zanieczyszczonej wody , a także jak karmić w zbiornikach rybackich.
Niektóre rodzaje zielonych alg są wykorzystywane przez ludność kilku krajów dla jedzenia. Na przykład do celów spożywczych w Japonii gatunki z tego rodzaju są specjalnie uprawiane Ulwa. Wodorosty te są szeroko stosowane, zwłaszcza w krajach Azji Południowo-Wschodniej, pod nazwą Sea Salad. Ulvaceae mają zauważalnie wyższą zawartość białka (do 20%) niż inne rodzaje glonów. Niektóre rodzaje zielonych alg są stosowane w jako producenci substancji fizjologicznie czynnych. Glony zielone są dobrym obiektem modelowym do różnorodnych badań biologicznych. Gatunki Hematococcus hoduje się w celu uzyskania astaksantyny, Botryococcus - w celu uzyskania lipidów. Jednocześnie śmierć ryb wiąże się z „rozkwitem” wody jednego z jezior na Tajwanie, spowodowanym przez Botryococcus.
Rodzaje porodu Chlorella I Chlamydomonas - obiekty modelowe do badania fotosyntezy w komórkach roślinnych. Chlorella, ze względu na bardzo wysokie wskaźniki reprodukcji, jest przedmiotem masowej uprawy z przeznaczeniem na różne pola
Warstwy powierzchniowe zielonych alg mają duże rozmiary wartość przeciwerozyjna. Niektóre jednokomórkowe gatunki zielonych alg, które wydzielają obfite ilości śluzu, mają działanie wiążące. Substancje śluzowe błon komórkowych sklejają ze sobą cząsteczki gleby. Rozwój glonów wpływa na strukturę drobnej ziemi, nadając ją wodoodporność i zapobieganie usuwaniu z warstwy powierzchniowej. Wilgotność gleby pod błonami glonów jest zwykle wyższa niż tam, gdzie ich nie ma. Ponadto folie zmniejszają przepuszczalność gleby i spowalniają parowanie wody, co również wpływa na reżim zasolenia gleby. Zmniejsza się wymywanie łatwo rozpuszczalnych soli z gleby; ich zawartość pod makroporostami glonów jest wyższa niż na innych obszarach. Jednocześnie spowalnia się wypływ soli z głębokich warstw gleby.
Glony glebowe wpływają także na wzrost i rozwój roślin wyższych. Uwalniając fizjologicznie substancje czynne przyspieszają wzrost sadzonek, zwłaszcza ich korzeni.
Wśród zielonych alg żyjących w zanieczyszczonych wodach dominują glony chlorokokowe, odporne na długotrwałe narażenie na wiele substancji toksycznych.
Komórki glonów są zdolne do gromadzenia różnych pierwiastki chemiczne, a ich stopa akumulacji jest dość wysoka. Słodkowodne algi zielone, zwłaszcza algi nitkowate, są silnymi koncentratorami. Jednocześnie intensywność akumulacji metali jest w nich znacznie wyższa niż w innych słodkowodnych organizmach wodnych. Bardzo interesująca jest zdolność glonów do koncentracji pierwiastków radioaktywnych. Martwe komórki glonów zatrzymują nagromadzone pierwiastki nie mniej mocno niż żywe, a w niektórych przypadkach desorpcja z martwych komórek jest mniejsza niż z żywych. Zdolność wielu rodzajów ( Chlorella, Scenedesmus itp.) koncentrują i mocno zatrzymują w swoich komórkach pierwiastki chemiczne i radionuklidy, co pozwala na ich wykorzystanie w wyspecjalizowanych systemach oczyszczania odkażenie przemysłowy Ścieki na przykład do dodatkowego oczyszczania niskoaktywnych ścieków z elektrowni jądrowych.
Niektóre zielone glony są antagoniści wirusa grypy, wirusa polio itp. Ważną rolę odgrywają substancje biologicznie czynne uwalniane przez glony dezynfekcja wody i tłumienie aktywności patogennej mikroflory.
W specjalnych stawach biologicznych korzystają zbiorowiska glonów i bakterii do rozkładu i detoksykacji herbicydów. Udowodniono zdolność wielu zielonych alg do hydrolizy herbicydu propanilu, który jest szybciej niszczony przez bakterie.
Pytania kontrolne
Nazwa cechy charakteru struktura komórkowa zielonych alg.
Jakie pigmenty i rodzaje składników odżywczych są znane w zielonych algach?
Jak rozmnażają się zielone algi? Co to są zoospory, aplanospory i autospory?
Jakie są klasy glonów zielonych?
Wymień charakterystyczne cechy zielonych alg z klasy Ulvophyceae.
Wymień charakterystyczne cechy zielonych alg z klasy Bryopsidae.
Wymień charakterystyczne cechy zielonych alg z klasy Chlorophyceae.
Wymień charakterystyczne cechy zielonych alg z klasy Trebuxiaceae.
Wymień charakterystyczne cechy zielonych alg z klasy Prasin.
W jakich siedliskach występują zielone algi? Opisz ich główne grupy ekologiczne.
Rola i znaczenie zielonych glonów w przyrodzie.
Jakie to jest znaczenie gospodarcze zielone algi?
Co to jest „rozkwit wody”? Udział zielonych glonów w biologicznym oczyszczaniu wody.
Zielone algi jako nietradycyjne źródła energii.
Wyjątkowość struktury wewnątrzkomórkowej krasnorostów polega zarówno na cechach zwykłych składników komórkowych, jak i na obecności specyficznych wtrętów wewnątrzkomórkowych.
Błony komórkowe. W ścianach komórkowych krasnorostów wyraźnie wyróżniają się dwie warstwy: wewnętrzna, zbudowana z celulozy i zewnętrzna, zbudowana ze związków pektynowych. W formach z szorstką, twardą plechą na zewnątrz ścian komórkowych rozwija się cienka warstwa naskórka. Skład i struktura naskórka roślin szkarłatnych różni się od naskórka roślin wyższych. Na przykład w porfirze powstaje w wyniku kondensacji cząsteczek mannozy monosacharydowej. W miękkich postaciach śluzu naskórek jest nieobecny.
Substancje pektynowe zawarte w algach czerwonych to sole wapniowe i magnezowe specjalnych kwasów pektynowych. Mają zdolność rozpuszczania się we wrzącej wodzie, tworząc roztwory śluzowe. Do grupy substancji pektynowych zaliczają się także specjalne substancje koloidalne zawarte w błonach komórkowych i przestrzeniach międzykomórkowych wielu szkarłatnych grzybów. Stanowią złożoną mieszaninę polisacharydów zawierających siarkę i są zbiorczo nazywane fikokoloidami. Fikokoloidy nie rozpuszczają się w zimnej wodzie, ale dobrze rozpuszczają się we wrzącej wodzie, tworząc układy koloidalne. Obecnie najważniejszymi produktami pozyskiwanymi z wodorostów są fikokoloidy z alg czerwonych, a także algi brunatne. Ponieważ substancje te są szeroko stosowane w życiu gospodarczym człowieka, ich skład chemiczny został dość dobrze zbadany. Fikokoloidy otrzymuje się z wielu gatunków, w wyniku czego powstało kilka odmian. Najbardziej znane to agar, karagen, nori i agaroidy. Substancje te różnią się między sobą składem i właściwościami, ale mają wspólną zdolność żelowania.
Obecność substancji koloidalnych determinuje zdolność błon komórkowych do silnego pęcznienia po śmierci, co utrudnia ich zabarwienie do badania.
Wśród krasnorostów występują formy, których muszle są zwapnione. Najpierw wapno osadza się w płycie środkowej, pomiędzy zewnętrzną i wewnętrzną warstwą łupiny, stopniowo wnikając w warstwę celulozy i mniej lub bardziej silnie ją impregnując. Ale nawet przy silnym zwapnieniu w środku zawsze pozostaje cienka membrana pozbawiona wapna, oddzielająca plazmę od warstwy wapiennej.
Skład związków wapna nie jest taki sam. Kalcyt odkłada się u koralowców, a aragonit u niektórych gatunków innych niż maliaceae. Ponadto znajdują się węglany wapnia i magnezu, a także żelazo.
Wzrost muszli następuje w następujący sposób. Jego nowe warstwy układane są na szczycie protoplastu komórki wierzchołkowej, natomiast warstwy zewnętrzne stopniowo rozpadają się w miarę wzrostu komórki. W rezultacie cała skorupa ma strukturę warstwową, a ponieważ nowe warstwy są nakładane pod pewnym kątem, wygląda jak lejek.
Cytoplazma krasnorostów charakteryzuje się dużą lepkością i ściśle przylega do ścian komórkowych. Bardzo łatwo ulega plazmolizie, dlatego krasnorosty są bardzo wrażliwe na zmiany warunków środowiskowych, w szczególności na odsalanie.
Rdzeń. Wśród czerwonych alg jest duża liczba tworzy się z komórkami jednojądrzastymi. Z reguły są to najprościej zorganizowane formy. W wysoce zorganizowanych komórkach szkarłatnych komórki są zwykle wielojądrowe, z wyjątkiem młodych komórek wzgórza - wierzchołka, rdzenia itp. Jednojądrzaste komórki gałęzi o ograniczonym wzroście w pewnych warunkach stają się wielojądrowe. Można jednak znaleźć również glony, w których stare komórki włókien centralnych są jednojądrzaste, a otaczające je młodsze komórki, wręcz przeciwnie, zawierają kilka jąder. Komórki rozrodcze - plemniki, karpospory, tetraspory - zawsze mają jedno jądro, ale jaja często są otoczone komórkami wielojądrzastymi, nawet w formach, których cała plecha składa się z komórek jednojądrzastych. Jądro krasnorostów jest małe, ma przezroczystą błonę jądrową i jąderko.
Chloroplasty. Komórki czerwonych alg zawierają jeden lub więcej chloroplastów. Rośliny z klasy Bangiaceae i prymitywni przedstawiciele najniższego zorganizowanego rzędu Nemaliaceae mają pojedynczy chloroplast w kształcie gwiazdy z jednym pyrenoidem. Zwykle zajmuje w komórce położenie osiowe i składa się wówczas z korpusu centralnego oraz wyrostków odchodzących od niego we wszystkich kierunkach (ryc. 149, 2). W chloroplastach osiowych pyrenoid znajduje się w jego środku. Procesy zachodzące w centralnym chloroplastie mogą rozszerzać się na obrzeżach i zamykać, tworząc chloroplast o nieregularnej lub wstęgowej ścianie. Prawdopodobnie chloroplast ścienny większości fioletowych chloroplastów pochodzi z chloroplastu osiowego w wyniku utraty części środkowej.
Rola pyrenoidu w krasnorostach nie jest zbyt jasna. W niektórych przypadkach jego obecność wiąże się z odkładaniem się ziaren skrobi; w innych pyrenoid występuje w komórkach, które nie biorą udziału w procesach asymilacji. W bardziej zorganizowanych formach pyrenoid znika; proces ten można prześledzić już w kolejności gatunków innych niż maliaceae.

U krasnorostów pozbawionych pirenoidów chloroplasty dzielą się na dwa główne typy - w kształcie wstążki i soczewki (lub w kształcie dysku) z licznymi przejściami między nimi (ryc. 154). Gatunki na niższym etapie ewolucji mają zwykle chloroplast przypominający wstążkę; przeciwnie, w przypadku form wysoce zorganizowanych bardziej charakterystyczne są chloroplasty w kształcie soczewki. To samo można powiedzieć o liczbie chloroplastów - ich liczba wzrasta wraz ze złożonością organizacji. Kształt chloroplastu nie jest czymś stałym, może zmieniać się wraz z wiekiem, warunkami oświetleniowymi, zmianami wielkości komórek, chociaż duże grupy szkarłatnych roślin często charakteryzują się chloroplastem o określonym kształcie. W ceramiu, w dużych komórkach międzywęźli, chloroplasty są wydłużone, przypominające wstążki, a w krótkich komórkach rdzeniowych na węzłach są to krótkie, nieregularnie płatkowane płytki. Powielanie chloroplastów następuje poprzez proste przeplatanie, jak u roślin wyższych.
Ze względu na subtelną strukturę, widoczną jedynie pod spodem mikroskop elektronowy, chloroplasty czerwonych alg prawie nie różnią się od chloroplastów innych alg.
Pigmenty. Szkarłatne kwiaty wyróżniają się złożonym zestawem pigmentów. Oprócz rozpuszczalnego w alkoholu chlorofilu, karotenu i ksantofilu powszechnych w roślinach zielonych, chloroplasty czerwonych alg zawierają dodatkowe pigmenty rozpuszczalne w wodzie, biliproteiny. Są to fikoerytryna i fikocyjanina. Rośliny zielone, zarówno algi, jak i lądowe, zawierają dwie modyfikacje chlorofilu - niebiesko-zielony chlorofil a i żółto-zielony chlorofil b. W krasnorostach występuje wyłącznie chlorofil a – uniwersalny pigment charakterystyczny dla wszystkich roślin. Ponadto w niektórych szkarłatnych roślinach znaleziono chlorofil d, którego charakter pozostaje jednak niejasny. Rośliny fioletowe zawierają niewiele zielonych pigmentów w porównaniu z roślinami wyższymi i zwykle są maskowane przez dodatkowe biliproteiny. Zaobserwowano pewną prawidłowość w zmianie ilości chlorofilu w zależności od ilości światła. Glony przystosowane do życia w warunkach słabego oświetlenia w morzach polarnych są zwykle bogatsze w chlorofil niż glony z mórz południowych. Podobnie glony głębinowe są bogatsze w chlorofil niż te rosnące w pobliżu powierzchni wody.
Do karotenoidów krasnorostów zalicza się α- i β-karoten oraz ksantofile, luteinę, zeaksantynę i prawdopodobnie taraksantynę. Biliproteinami szkarłatnych grzybów są czerwona fikoerytryna i niebieska fikocyjanina. Są zbliżone do pigmentów niebiesko-zielonych alg, ale nie są z nimi identyczne, ponieważ różnią się skład chemiczny. Jak wykazano w licznych eksperymentach, liczba pigmentów w szkarłatnych grzybach wzrasta wraz z głębokością; w tym przypadku ilość fikoerytryny wzrasta w większym stopniu niż ilość chlorofilu. Każdy, kto zebrał te glony w naturze, wie, że glony czerwono zabarwione rosną na głębokości, a w płytkiej wodzie zmieniają kolor. W miarę wzrostu ilości światła stają się bladoczerwone, następnie żółtozielone, słomkowe, a na koniec całkowicie wybielone.
Istnieje teoria tzw. adaptacji chromatycznej, według której wnikanie glonów na określone głębokości wiąże się z jakością światła przechodzącego przez słup wody. Jak wiadomo, promienie z zielonej i niebieskiej części widma wnikają najgłębiej. Czerwone pigmenty szkarłatnic umożliwiają im fotosyntezę w świetle niebieskim, dlatego zgodnie z tą teorią wnikają w głębiny niedostępne dla innych glonów. Jednak w praktyce ten schemat nie zawsze jest przestrzegany. Jaka jest rola biliprotein w fotosyntezie krasnorostów? W eksperymentach stwierdzono, że przy słabym oświetleniu uczestniczą w zwiększonej absorpcji światła. Dlatego można je uznać za sensybilizatory optyczne. Zatem przenikanie krasnorostów na znaczne głębokości można lepiej wyjaśnić ich zdolnością do pochłaniania niewielkich ilości światła. Ogólnie rzecz biorąc, algi fioletowe są organizmami tolerującymi cień: potrafią lepiej wykorzystywać słabe światło niż inne algi. Jeśli czerwony kolor alg w słabym świetle przynosi korzyści, to w silniejszym świetle, wręcz przeciwnie, intensywność fotosyntezy szkarłatnych alg jest niższa niż w przypadku innych alg, właśnie ze względu na obecność czerwonych pigmentów. Aby chronić się przed silnym światłem, szkarłatne ryby żyjące na płytkich głębokościach, zwłaszcza w morzach tropikalnych i subtropikalnych, mają specjalne opalizujące ciała. Te matowe żółte nieregularny kształt Ciała powstają w wakuolach komórek powierzchniowych wzgórza i składają się z małych ziaren o charakterze białkowym. Mają zdolność rozpraszania i odbijania padających na nie promieni słonecznych. Przy bardzo silnym oświetleniu ciała drażniące znajdują się pod zewnętrzną ścianą komórki, natomiast chloroplast na wewnętrznej lub bocznej ścianie i służą jako rodzaj kurtyny dla chloroplastu. Kiedy roślina wchodzi w warunki rozproszonego światła, następuje wzajemny ruch i chloroplast trafia na zewnętrzną ścianę.
Glony o opalizujących ciałach mają zwykle w padającym świetle niebieskawo-stalowy połysk. U niektórych gatunków w komórkach pojawiają się duże ciała w kształcie soczewek, które znikają wraz ze zmniejszającym się oświetleniem.
Substancje zapasowe. W wyniku asymilacji algi czerwone odkładają polisacharyd zwany skrobią szkarłatną. Z natury chemicznej jest najbliżej amylopektyny i glikogenu i najwyraźniej zajmuje pozycję pośrednią między zwykłą skrobią a glikogenem. Skrobia fioletowa osadza się w postaci małych półstałych ciał o różnych kształtach i kolorach. Bryły te mogą mieć kształt stożków lub płaskich owalnych płytek z wgłębieniem na szerokiej powierzchni. Często widać na nich koncentryczne strefy. Fioletowe ziarna skrobi powstają częściowo w cytoplazmie, częściowo na powierzchni chloroplastów, ale nigdy nie tworzą się wewnątrz plastydów, w przeciwieństwie do zwykłej skrobi roślin zielonych. W formach zawierających pirenoid ten ostatni jest w pewnym stopniu zaangażowany w syntezę skrobi.
Oprócz skrobi szkarłatnej, w czerwonych algach jako substancje rezerwowe odkładają się cukry: trehaloza, florydozyd, sacharoza itp. W niektórych postaciach występują w dużych ilościach alkohole wielowodorotlenowe. Wśród tłuszczów znane są cholesterol, silosterol i fukosterol. Zawartość tłuszczu różni się w zależności od warunków środowiskowych.

Komórki gruczołowe. Osobliwością krasnorostów jest obecność u niektórych przedstawicieli klasy florydyjskiej specjalnych komórek o bezbarwnej zawartości, które silnie załamują światło (ryc. 155). W literaturze nazywane są komórkami pęcherzykowymi lub gruczołowymi. Zawartość tych komórek jest różna u różnych glonów; są wypełnione związkami jodku, rzadziej bromu. Najczęściej komórki gruczołowe znajdują się w kolejności ceramidów. W nitkowatych rozgałęzionych glonach antytamnion siedzą na górnej stronie bocznych gałęzi (ryc. 155, 2). Podczas ich rozwoju najpierw oddziela się małą komórkę w kształcie soczewki, zawierającą niewielką ilość osocza i małe czerwone chloroplasty. Jądro można prześledzić jedynie na najwcześniejszym etapie rozwoju. Wkrótce na dnie tej komórki tworzy się bezbarwna, załamująca światło bańka. Rośnie, a wraz z nim powiększa się cała komórka. W utworzonej komórce większość z nich zajmuje pęcherzyk, a tylko w górnej części pozostaje wąska warstwa plazmy z małymi chloroplastami (ryc. 155, 2-5). Rola komórek pęcherzykowych nie została wyjaśniona, chociaż istnieje wiele różnych założeń na ten temat. Uważa się je za słabo rozwinięte zarodnie, magazynujące substancje rezerwowe, „pęcherze pławne” itp.
Komórki gruczołowe są charakterystyczne dla niektórych grup glonów i dlatego pełnią ważną funkcję taksonomiczną.

Owłosienie. Tworzenie się włosów jest zjawiskiem powszechnym w klasie Florididae. Prawdziwe szkarłatne włosy należy odróżnić od włosowatych gałęzi lub sztucznych włosów. U gatunków z jednorzędową plechą nitkowatą można zobaczyć, jak komórki końcowe gałęzi bocznych wydłużają się i odbarwiają, uzyskując strukturę przypominającą włos (ryc. 151, 7). To są sztuczne włosy. Prawdziwe włosy czerwonych alg dzielą się na dwa typy: jednokomórkowe i wielokomórkowe. Włosy jednokomórkowe nigdy się nie rozgałęziają. W jednorzędowych postaciach nitkowatych powstają z wierzchołkowych komórek gałęzi, w wielorzędowych - z powierzchniowych komórek kory. Komórka przyszłego włosa jest oddzielona od komórki macierzystej poprzeczną przegrodą i jest znacznie wydłużona, często osiągając długość milimetra lub więcej (ryc. 156). Zawiera jądro i niewielką ilość cytoplazmy. Chloroplast znika w miarę wzrostu włosów, a włosy stają się bezbarwne. Zwykle komórka macierzysta włosa nie różni się niczym od sąsiednich komórek wegetatywnych, ale czasami jest znacznie większa i pozostaje wyraźnie widoczna po wypadaniu włosa. W przypadku włosów koralowych włosy nie są niezależnymi komórkami, lecz jedynie wyrostkiem specjalnych komórek, od których nie są oddzielone przegrodą. Komórki te są znacznie większe od pozostałych i nazywane są trichocytami lub heterocystami (ryc. 172, 2, 3). Jeśli w klasie Floridaeidae włosy jednokomórkowe występują dość często, to w Bangiaceae są one całkowicie nieobecne.
 ,
, 
Włosy wielokomórkowe są zwykle mniej lub bardziej silnie rozgałęzione. Występują jedynie w niektórych glonach z rzędu Ceramiaceae. Jak wykazały eksperymenty, główną rolą włosów jest ułatwianie wchłaniania składników odżywczych z otoczenia.
Pory. Jeden z najbardziej ciekawe funkcje krasnorostów polega na tym, że komórki tworzące plechę są połączone ze sobą za pomocą specjalnych formacji zwanych norami. Pomiędzy komórkami potomnymi, czyli komórkami pochodzącymi od jednej matki, połączenie odbywa się poprzez pory pierwotne (ryc. 157, 2-3). Powstają w wyniku niepełnej przegrody pomiędzy dwiema nowo powstałymi komórkami. Pory pierwotne znajdują się pośrodku przegrody, w miejscu, przez które można poprowadzić linię łączącą jądra komórek potomnych i stanowią cienką płytkę. Nici przechodzą przez tę płytkę i łączą cytoplazmę sąsiednich komórek. Do niedawna uważano, że pory pierwotne są charakterystyczne tylko dla Floridaeidae i nie występują w klasie Bangiaceae – była to jedna z podstawowych cech, na podstawie których wyróżniono obie te klasy. Ale ostatnio pory pierwotne zostały otwarte także u przedstawicieli klasy Bangiaceae.

Życie roślin: w 6 tomach. - M.: Oświecenie. Pod redakcją A. L. Takhtadzhyana, Redaktor naczelny członek-kor. Akademia Nauk ZSRR, prof. AA Fiodorow. 1974 . Encyklopedia biologiczna
Zewnętrznie plechy krasnorostów są bardzo różnorodne, często piękne i dziwaczne (tabele 20-23). Można tu znaleźć formy nitkowate i blaszkowate, cylindryczne i skorupiaste, bąbelkowe i koralowe, w różnym stopniu... ... Encyklopedia biologiczna
Komórka jest podstawową jednostką strukturalną organizmu glonów, reprezentowaną przez formy jednokomórkowe lub wielokomórkowe. Całkowicie unikalną grupę stanowią algi syfonowe: ich plechy nie dzielą się na komórki, ale w cyklu... ... Encyklopedia biologiczna
Rozmnażanie własnego rodzaju w algach następuje poprzez rozmnażanie wegetatywne, bezpłciowe i płciowe. Rozmnażanie wegetatywne glonów jednokomórkowych polega na podziale osobników na dwie części. W algach wielokomórkowych... Encyklopedia biologiczna
W odróżnieniu od roślin wyższych, które w całości charakteryzują się jedną budową typu liściowo-łodygowego (inna budowa wynika z wtórnego uproszczenia), glony w obrębie typu warstwowego wykazują wyjątkową... ... Encyklopedia biologiczna
Rhodomonas salina… Wikipedia
- (nośniki kolorów) tą nazwą można nazwać wszystkie kolorowe ciała zawarte w komórkach roślinnych, ale konkretnie odnosi się ona do tych zawartych w komórkach alg (patrz), w przeciwieństwie do ziaren chlorofilu (patrz) i chromoplasty (patrz), .. .... słownik encyklopedyczny F. Brockhausa i I.A. Efron
Klasa Bangiaceae łączy jednokomórkowe, kolonialne i wielokomórkowe formy struktury miąższowej. Ich wzrost jest rozproszony, w wyniku podziału wszystkich komórek plechy. Komórki Banguiaceae są jednojądrzaste, z jednym chloroplastem gwiaździstym i jednym... ... Encyklopedia biologiczna
Całkowita powierzchnia planety Ziemia wynosi 510 milionów km2. Powierzchnia lądu wynosi 149 mln km2, Ocean Światowy zajmuje 361 mln km2. Zarówno ląd, jak i ocean zamieszkują rośliny i zwierzęta. Różnorodność obu jest bardzo duża. Teraz ustalono... Encyklopedia biologiczna
Wszystkie glony różnią się znacznie zestawem pigmentów fotosyntetycznych. Takie grupy w taksonomii roślin mają status działów.
Głównym pigmentem wszystkich alg jest zielony pigment chlorofil. Znane są cztery rodzaje chlorofilu, różniące się budową: chlorofil a– występuje we wszystkich algach i roślinach wyższych; chlorofil b– występuje w algach zielonych, charofitach, euglenoidach i roślinach wyższych: rośliny zawierające ten chlorofil zawsze mają jasnozieloną barwę; chlorofil c– występuje w algach heterokontowych; chlorofil d– rzadka forma, występująca u alg czerwonych i niebieskozielonych. Większość roślin fotosyntetyzujących zawiera dwa różne chlorofile, z których jeden jest zawsze chlorofilem A. W niektórych przypadkach zamiast drugiego chlorofilu są biliproteiny. Istnieją dwa rodzaje biliprotein występujących w niebiesko-zielonych i czerwonych algach: fikocyjanina– niebieski pigment, fikoerytryna- czerwony pigment.
Obowiązkowymi pigmentami zawartymi w błonach fotosyntetycznych są pigmenty żółte - karotenoidy. Różnią się od chlorofilów widmem pochłanianego światła i uważa się, że pełnią funkcję ochronną, chroniąc cząsteczki chlorofilu przed destrukcyjnym działaniem tlenu cząsteczkowego.
Oprócz wymienionych pigmentów algi zawierają również: fukoksantyna– złoty pigment; ksantofil- brązowy pigment.
Koniec pracy -
Ten temat należy do działu:
Wodorost
Uniwersytet Rybacki.. Instytut Biologii Morza im. A. w Żyrmuńskim Dworze Ran.. l l Arbuzov..
Jeśli potrzebujesz dodatkowy materiał na ten temat lub nie znalazłeś tego, czego szukałeś, polecamy skorzystać z wyszukiwarki w naszej bazie dzieł:
Co zrobimy z otrzymanym materiałem:
Jeśli ten materiał był dla Ciebie przydatny, możesz zapisać go na swojej stronie w sieciach społecznościowych:
| Ćwierkać |
Wszystkie tematy w tym dziale:
Osłony komórek
Osłony komórek zapewniają odporność wewnętrznej zawartości komórek na wpływy zewnętrzne i nadają komórkom określony kształt. Osłony są przepuszczalne dla wody i małych cząsteczek w niej rozpuszczonych
Wici
Monadyczne komórki wegetatywne i monadyczne etapy cyklu życiowego (zoospory i gamety) glonów są wyposażone w wici - długie i dość grube wyrostki komórkowe, pokryte zewnętrznie plazmalemmą. I
Mitochondria
Mitochondria występują w komórkach alg eukariotycznych. Kształt i struktura mitochondriów w komórkach glonów jest bardziej zróżnicowana w porównaniu do mitochondriów roślin wyższych. Mogą być okrągłe
Plastydy
Pigmenty w komórkach alg eukariotycznych znajdują się w plastydach, jak we wszystkich roślinach. W algach występują dwa rodzaje plastydów: kolorowe chloroplasty (chromatofory) i bezbarwne leukoplasty (ami
Jądro i aparat mitotyczny
Jądro glonów ma strukturę typową dla eukariontów. Liczba jąder w komórce może wahać się od jednego do kilku. Na zewnątrz rdzeń pokryty jest powłoką składającą się z dwóch membran, czyli membrany zewnętrznej
Monadyczny (wiciowy) typ struktury plechy
Bardzo cecha charakterystyczna, który determinuje ten typ struktury, to obecność wici, za pomocą których organizmy monadyczne aktywnie poruszają się środowisko wodne(ryc. 9, A). Ruchome z
Struktura typu ryzopodialnego (ameboidalnego).
Najważniejszymi cechami struktury typu ameboidalnego jest brak silnych osłon komórkowych i zdolność do poruszania się ameboidalnego za pomocą qi tymczasowo utworzonego na powierzchni komórki
Struktura typu palmelloidowego (hemimonadalnego).
Charakterystyczne dla tego typu struktury jest połączenie stacjonarnego trybu życia roślin z obecnością organelli komórkowych charakterystycznych dla organizmów monadycznych: wakuoli kurczliwych, piętna, opaski uciskowej
Struktura typu kokosowego
Ten typ łączy glony jednokomórkowe i kolonialne, nieruchome w stanie wegetatywnym. Komórki typu kokosowego są pokryte błoną i mają protoplast typu roślinnego (tonoplast bez sokratesa
Struktura typu trichal (nitkowatego).
Charakterystyczną cechą struktury typu nitkowatego jest nitkowaty układ nieruchomych komórek, które powstają wegetatywnie w wyniku podziału komórek, który zachodzi głównie
Struktura typu heterotrychalnego (niewłóknistego).
Typ heterofilamentowy powstał na bazie typu nitkowatego. Plecha heterofilamentowa składa się głównie z nici poziomych pełzających po podłożu, pełniących funkcję mocowania, oraz nitek pionowych, wzdłuż
Struktura miąższowa (tkankowa).
Jeden z kierunków ewolucji plechy heterofilamentowej wiązał się z pojawieniem się plechy miąższowej. Zdolność do nieograniczonego wzrostu i podziału komórek w różnych kierunkach doprowadziła do ewolucji
Konstrukcja typu syfonowego
Struktura typu syfonalnego (niekomórkowego) charakteryzuje się brakiem komórek wewnątrz wzgórza, które osiąga stosunkowo duże, zwykle makroskopowe rozmiary i pewien stopień zróżnicowania
Konstrukcja typu syfonokladalnego
Główną cechą struktury typu syfonokladalnego jest zdolność do tworzenia kompleksowo ułożonych wzgórz, składających się głównie z segmentów wielojądrzastych, z pierwotnego wzgórza niekomórkowego. W
Rozmnażanie bezpłciowe
Rozmnażanie bezpłciowe glonów odbywa się za pomocą wyspecjalizowanych komórek - zarodników. Sporulacji zwykle towarzyszy podział protoplastu na części i uwolnienie z nich produktów rozszczepienia
Prosty podział
Ta metoda rozmnażania występuje tylko w jednokomórkowych formach glonów. Podział zachodzi najłatwiej w komórkach o budowie ciała typu ameboidalnego. Podział form ameboidalnych
Podział
Fragmentacja jest nieodłączna dla wszystkich grup glonów wielokomórkowych i objawia się w różnych postaciach: tworzeniu hormogoniów, regeneracji oderwanych części plechy, spontanicznej utraty gałęzi, odrastania
Rozmnażanie przez pędy, rozłogi, pąki czerwowe, guzki, akinety
W postaciach tkankowych zielonych, brunatnych i czerwonych alg rozmnażanie wegetatywne przybiera pełną formę, która niewiele różni się od rozmnażania wegetatywnego roślin wyższych. Trzymanie drogi
Rozmnażanie płciowe
Rozmnażanie płciowe u alg jest związane z procesem płciowym, który polega na połączeniu dwóch komórek, w wyniku czego powstaje zygota, z której wyrasta nowy osobnik lub wytwarza zoospory.
Zmiana faz jądrowych
Podczas procesu seksualnego, w wyniku połączenia gamet i ich jąder, liczba chromosomów w jądrze podwaja się. Na pewnym etapie cyklu rozwojowego, podczas mejozy, następuje zmniejszenie liczby chromosomów
Endofity/endozoity lub endosymbionty
Endosymbionty, czyli symbionty wewnątrzkomórkowe, to glony żyjące w tkankach lub komórkach innych organizmów (bezkręgowców lub glonów). Tworzą rodzaj grupy ekologicznej
Dział sinic (cyjanobakterii) – cyanophyta
Nazwa działu (od greckiego cyanos - niebieski) odzwierciedla cecha charakterystyczna tych alg - kolor plechy wiąże się ze stosunkowo dużą zawartością fikocyjaniny, niebieskiego pigmentu. Cyjan
Zamówienie – Chroococcales
Występują jako jednokomórkowe „proste” osobniki lub częściej tworzą kolonie śluzowe. Kiedy komórki dzielą się na dwie płaszczyzny, pojawiają się jednowarstwowe kolonie blaszkowate. Podział w trzech punktach
Dział krasnorostów – rhodophyta
Nazwa wydziału pochodzi od greckie słowo rodon („rodon”) – różowy. Kolor czerwonych alg wynika z różnych kombinacji pigmentów. Występuje w kolorze szarym i fioletowym
Zamawiaj Banguiaceae – Bangiales
Rodzaj Porphyra ma plechę w postaci cienkiej błyszczącej płytki o gładkich lub złożonych krawędziach, składającej się z jednej lub dwóch warstw ściśle połączonych komórek. Podstawa płyty zwykle wchodzi
Zamów Rhodymeniales
Rodzaj Sparlingia (Rodimenia) - płaskie płytki do 45 cm wysokości, w kształcie liścia i klina, rozszerzone i palmatelnie rozcięte u góry, od jasnoróżowego lub jasnopomarańczowego do
Zamów Coralline - Corallinales
Rodzaj Coralline to podzielony na segmenty, wachlarzowaty, rozgałęziony krzew do 10 cm wysokości, rozgałęziony, wapienny, od różowo-liliowego do prawie białego. Rozmnaża się bezpłciowo i płciowo. Spo
Zamów Gigartinales – Gigartinales
Rodzaj Hondrus - gęste, skórzaste krzewy chrzęstne do 20 cm wysokości, 3-4 razy rozgałęzione, jasnożółte, jasnoróżowe, fioletowo-ciemnoczerwone. Rośnie w dolnej części strefy przybrzeżnej i
Zamów Ceramiaceae – Ceramiales
Rodzaj Ceramium to delikatny, puszysty, segmentowany krzew dorastający do 10 cm wysokości, rozgałęziony dychotomicznie lub naprzemiennie, ciemnożółty z różowawym odcieniem. Oczywiście rozgałęzienie od dwóch do czterech rzędów
Podział okrzemek - Bacillariophyta
Dział nazywa się Okrzemki (od greckiego di - dwa, tom - cięcie, sekcja) lub Bacillaria (bacillum - kij). Obejmuje jednokomórkową organizację samotną lub kolonialną.
Podział glonów heterokontowych (heterokontophyta)
Wszystkie heterokonty mają podobną strukturę aparatu wiciowego. Są 2 wici, a jedna z nich ma bardzo charakterystyczne rurkowate, trójczłonowe pierzaste narośla, czyli włosy - mastigonemy. Dokładnie gotówką
Taksonomia
Kokolity kopalne znane są z osadów mezozoiku i występowały obficie przez większość okresów jurajskiego i kredowego. Prymnesiofity osiągnęły największą różnorodność w późnej kredzie,
Dział glonów kryptofitowych (kryptomonad) – cryptophyta
Nazwa działu pochodzi od rodzaju Cryptomonas (od greckiego kryptos – ukryty, monas – indywidualny). Obejmuje organizmy jednokomórkowe, ruchliwe i monadyczne. Komórki kryptofitów
A B C D E
Ryż. 53. Wygląd glonów kryptofitowych (wg: G.A. Belyakova i in., 2006): A – Rodomonas, B – Chroomonas, C – Cryptomonas, D – Chilomonas, E – Goniomonas can for
Dział zielonych alg – chlorophyta
Glony zielone są najliczniejszą ze wszystkich grup glonów, licząc według różnych szacunków od 4 do 13 - 20 tysięcy gatunków. Wszystkie mają zielony kolor thalli, co wynika z przewagi chloru
Zamów Ulothrixales – Ulotrichales
Rodzaj Ulothrix (ryc. 54). Gatunki Ulotrix częściej żyją w wodach słodkich, rzadziej w morzach, zbiornikach słonawych i w glebie. Przywiązują się do podwodnych obiektów, tworząc jasnozielone krzewy.
Zamów Bryopsidae – Bryopsidales
Większość gatunków występuje w wodach słodkich i słonawych. Część z nich rośnie na ziemi, na kamieniach, piasku, czasem na słonych bagnach. Rodzaj Bryopsis - krzewy nitkowate do 6-8 s
Zamów Volvocales - Volvocales
Rodzaj Chlamydomonas (ryc. 57) obejmuje ponad 500 gatunków jednokomórkowych glonów żyjących w świeżych, małych, dobrze ogrzanych i zanieczyszczonych zbiornikach wodnych: stawach, kałużach, rowach itp. Itp
Oddział Charophyta (Characeae) – Charophyta
Charofity to linia słodkowodnych zielonych alg, z których wywodzą się rośliny wyższe. Są to formy przeważnie z plechą nitkowatą. Często plecha jest pionowa, rozcięta i niesiona
Podział dinofitów (dinofagellates) – dinophyta
1. Nazwa katedry pochodzi z języka greckiego. dineo – obracać się. Łączy przeważnie jednokomórkowe monadyczne, rzadziej kokosowe, ameboidalne lub palmelloidowe, czasem kolonialne
Podział euglenozoa - euglenovae
Nazwa działu pochodzi od rodzaju typu - Euglena (od greckiego eu - dobrze rozwinięty, glene - źrenica, oko). Łączy pojedynczych przedstawicieli monadycznych lub ameboidalnych. Czasami się spotykamy
Słownik terminów
Autogamia – rozmnażanie płciowe, w którym dwa siostrzane jądra haploidalne łączą się we wspólnej cytoplazmie. Autospora to struktura rozmnażania bezpłciowego, która jest
Glony, jednokomórkowe i wielokomórkowe formy glonów bentosowych. Występują tu wszystkie typy morfologiczne plechy, z wyjątkiem jednokomórkowych ryzopodialnych i dużych form wielokomórkowych z złożona struktura. Wiele nitkowatych zielonych glonów przyczepia się do podłoża dopiero we wczesnych stadiach rozwoju, następnie stają się wolno żyjące, tworząc maty lub kule.
| Zielone algi | ||||||||||||
| Różnorodność glonów syfonowych. Ilustracja z książki Ernsta Haeckela Kunstformen der Natur, 1904 |
||||||||||||
| Klasyfikacja naukowa | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Międzynarodowa nazwa naukowa | ||||||||||||
|
Chlorofit Paschera, 1914 |
||||||||||||
|
||||||||||||
Najbardziej rozległe są glony zielone dany czas dział glonów: według przybliżonych szacunków obejmuje od 13 000 do 20 000 gatunków. Wszystkie wyróżniają się przede wszystkim czysto zielonym kolorem plech, zbliżonym do koloru roślin wyższych i spowodowanym przewagą chlorofilu nad innymi pigmentami.
Struktura
Komórki wici zielonych alg są izokontami - wici mają podobną strukturę, chociaż mogą różnić się długością. Zwykle są dwie wici, ale może być ich również cztery lub wiele. Wici zielonych alg nie mają mastigonemów (w przeciwieństwie do heterokontów), ale mogą mieć wdzięczne włosy lub łuski.
Cykle życia
Cykle życiowe zielonych alg są bardzo zróżnicowane. Jest tu cała gama typów.
Haplobiont z redukcją zygotyczną ( Hydrodictyon reticulatum, Eudorina). Gamety dwuwiciowe są uwalniane z komórki rodzicielskiej przez pory w jej błonie, a fuzja gamet odbywa się za pomocą rurki. Następnie zygota zamienia się w zygosporę spoczynkową, która po okresie fizjologicznego spoczynku kiełkuje, tworząc 4 zoospory (w wyniku podziału mejotycznego). Każda zoospora tworzy wielościan i kiełkuje, tworząc małe kuliste sieci przylegających zoospor.
Haplo-diplobiont z redukcją sporadyczną ( Ulwa, Ulothrix, niektóre typy Kladofora). Izogamety biflagellatowe wyłaniają się z komórki macierzystej, po czym gamety utworzone przez różne włókna łączą się w wodzie. Tworzy się zygota z czterema wiciowcami, która aktywnie unosi się w wodzie. Następnie opada na jakieś podłoże i pokrywa się gęstą skorupą, przekształcając się w komórkę maczugowatą (codiolum), po czym następuje etap fizjologicznego odpoczynku. W sprzyjających warunkach kiełkuje w 4-16 zoospor lub aplanospor, które po krótkim okresie pływania przyczepiają się do podłoża i wyrastają w nowe włókna. Różne czynniki aktywują wyjście ze stanu uśpienia: wzrost temperatury, zmiana otoczenia itp.
Diplobiont z redukcją gamet ( Bryopsis). Planozygota osiada i wyrasta w nitkowatą plechę z dużym jądrem; jądro dzieli się, tworząc zoospory stephanokont, które wyrastają na plechę wegetatywną.
Szczególnie dużo zielonych glonów rozwija się wiosną, kiedy wszystkie kamienie w strefie przybrzeżnej pokryte są ciągłą szmaragdową powłoką zielonych alg, ostro kontrastującą z białym śniegiem zalegającym na przybrzeżnych kamieniach. W wyniku rozwijających się włókienek - ulotrix ( Ulothrix) i urospora ( Urospora). Latem często rozwija się dużo Egagropyli ( Aegagropila linnaei) (syn. Cladophora aegagropila), która często wygląda jak zielona masa śluzowa. Na otwartym skalistym wybrzeżu jasnozielone, rozgałęzione krzewy tworzą akrosyfonię ( Akrosyfonia).
Rola w przyrodzie i użytkowaniu
Niektóre zielone algi (takie jak ulva) są powszechnie spożywane. Chlorella służy jako wskaźnik poziomu zanieczyszczenia wody i jest zawarta w: statki kosmiczne, łodzie podwodne do oczyszczania powietrza z dwutlenku węgla.







