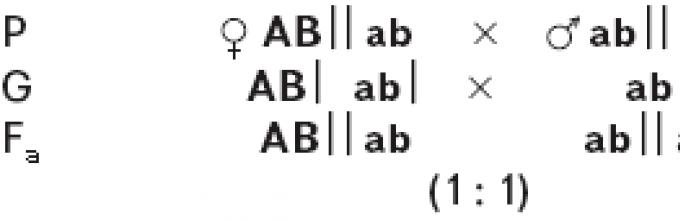
32. Особенности наследования признаком при неполном и полном сцелении
СЦЕПЛЕННОЕ НАСЛЕДОВАНИЕ ПРИЗНАКОВ
Мы познакомились с дигибридным скрещиванием и уяснили, что независимое комбинирование признаков объясняется тем, что расщепление одной пары аллельных генов, определяющих соответствующие признаки, происходит независимо от другой пары. Однако это наблюдается только в том случае, когда гены разных пар находятся в разных парах хромосом и при образовании половых клеток гибрида в мейозе отцовские и материнские хромосомы независимо комбинируются. Но количество хромосом очень ограниченно по сравнению с количеством признаков, каждый из которых развивается под контролем определенного гена. Так, у дрозофилы известно около 7000 генов при четырех парах хромосом. Предполагается, что у человека не менее 50 тыс. генов при 23 парах хромосом, и т. д. Отсюда следует, что в каждой паре хромосом должны быть локализованы сотни аллелей. Естественно; что между генами, которые находятся в одной хромосоме, наблюдается сцепление и при образовании половых клеток они должны передаваться вместе.
Сцепленное наследование открыли в 1906 г. английские генетики У. Бэтсон и Р. Пеннет при изучении наследования признаков у душистого горошка, но они не смогли вскрыть причины этого явления. Природу сцепленного наследования в 1910 г. выяснили ученые Т. Морган и его сотрудники К. Бриджес и А. Стертевант. В качестве объекта исследования они избрали плодовую муху дрозофилу, которая оказалась очень удобной для генетических опытов. В клетках тела дрозофилы находится 4 пары хромосом. Она отличается очень высокой плодовитостью - одна пара дает более ста потомков. У нее большая скорость развития - в течение 12-15 дней после оплодотворения из яйца развиваются личинка, куколка и взрослая особь, которая почти сразу же способна давать потомство. Можно исследовать в течение года более двадцати поколений. Мухи серого цвета, с красными глазами,. имеют маленькие размеры (около 3 мм), легко разводятся в биологических пробирках; для изучения их признаков можно пользоваться лупами. При просмотре сотен тысяч особей Морган обнаружил множество разных мутаций: встреча-лись мухи с черным и желтым телом, с белыми и другого цвета глазами, с измененной формой и положением крыльев и т. д. Иногда попадались особи, имеющие сразу несколько мутаций, например черное тело, зачаточные крылья, киноварные глаза.
Изучая наследование разных пар признаков при дигибридном и полигибридном скрещиваниях, Морган и его сотрудники обнаружили большое число примеров сцепленного (совместного) их наследования. Все изученные признаки распределились на четыре группы сцепления в соответствии с числом и размерами хромосом у дрозофилы. На этом основании Морган сделал вывод о том, что гены, определяющие эти признаки, находятся в хромосомах. Гены, расположенные в одной хромосоме, представляют собой группу сцепления.
Сцепление генов - это совместное наследование генов, расположенных в одной и той же хромосоме. Количество групп сцепления соответствует гаплоидному числу хромосом. Например, у дрозофилы 4 группы сцепления, у человека 23, у крупного рогатого скота 30, у свиней 19 и т. д.
Мысль о расположении генов в хромосомах высказал Сеттон еще в 1902 г. Он обнаружил параллелизм в поведении хромосом в мейозе и наследовании признаков у одного из видов кузнечика. Дальнейшие исследования, проведенные Морганом, показали, что сцепление генов, расположенных в одной хромосоме, может быть полным или неполным.
Наиболее четко разница в поведении сцепленных и независимо наследующихся генов выявляется при проведении анализирующего скрещивания. При независимом наследовании двух пар признаков у гибрида Fi (АаВЬ) с равной вероятностью образуется 4 сорта гамет: АВ, Ab , aB , ab . При скрещивании с полным рецессивом (aabb ) количество сортов гамет у гибрида обусловливает число типов потомков и одинаковую вероятность их появления, так как гаметы рецессивной особи (ab ) не могут изменить проявления доминантных и рецессивных генов гамет гибрида. В результате соотношение фенотипов потомства будет равно 1:1:1:1. Если же обе пары аллельных генов расположены в одной паре хромосом, то при образовании половых клеток гены этих аллелей не смогут свободно комбинироваться. В этом случае наблюдается сцепленное наследование.
ПОЛНОЕ СЦЕПЛЕНИЕ
Т. Морган скрещивал черных длиннокрылых самок с серыми с зачаточными крыльями самцами. У дрозофилы серая окраска тела доминирует над черной, длиннокрылость - над зачаточными крыльями. Обозначим ген серой окраски тела В, аллельный ему ген черной окраски тела Ь; ген длиннокрылое™ V , аллельный ему ген зачаточных крыльев v. Обе пары этих генов нахо-дятся в одной и той же второй паре хромосом. По обеим парам Признаков родительские формы были гомозиготны: самка по рецессивному признаку черного тела (bb ) и доминантному признаку длиннокрылое™ (VV ), самец по доминантному признаку серой окраски. (ВВ) и рецессивному признаку зачаточных крыльев (w>. Гаметы родителей при редукционном делении получают у материнской формы хромосому с генами b и V , у отцовской"-с генами В и v. Все потомство первого поколения (Fi) имело серое тело и длинные крылья (рис. 13) и было гетерозиготно по обеим парам признаков (bV / Bv ). Затем из Fi были отобраны самцы, которых скрестили с гомозиготными по обоим рецессивным генам самками, черными зачаточнокрылыми (bv / bv ), т. е. было проведено анализирующее скрещивание, в результате которого при независимом комбинировании признаков должны были бы получить потомство четырех фенотипов в равных соотношениях: серых длиннокрылых, серых с зачаточными крыльями, черных длиннокрылых, черных с зачаточными крыльями, но были получены потомки только двух фенотипов, похожих на исходные родительские формы: черные длиннокрылые и серые короткокрылые. В этом случае наблюдается полное сцепление признаков. Это связано с тем, что у гетерозиготного самца в одной и той же хромосоме из гомологичной пары расположены и ген черной окраски, и ген длинных крыльев, в другой - ген серой окраски и ген зачаточнокрылости.
При спермиогенезе в период мейоза гомологичные хромосомы расходятся в разные половые клетки. Образуется только два сорта гамет: один с хромосомой, которая несет гены Ъ и V , другой с хромосомой, в которой расположены гены В и v. При сочетании указанных гамет с гаметами особи с рецессивными признаками и образуется потомство только двух типов. При полном сцеплении гены, расположенные в одной хромосоме, всегда передаются вместе. Полное сцепление пока установлено только у са>щов дрозофилы и самок тутового шелкопряда.
НЕПОЛНОЕ СЦЕПЛЕНИЕ
В следующем опыте, так же как и в предыдущем, Морган скрещивал черных длиннокрылых самок с серыми зачаточно-крылыми самцами. В первом поколении получил все потомство серое длиннокрылое. Затем снова произвел анализирующее скрещивание, но из первого поколения отобрал не самца, а самку и скрестил ее с черным с зачаточными крыльями самцом (рис. 14). В этом случае появилось потомство не двух типов, как при полном сцеплении, а четырех: серое о-зачаточными крыльями, черное длиннокрылое, серое длиннокрылое и черное с зачаточными крыльями, но не в равных соотношениях, как при независимом комбинировании признаков, а со значительным преобладанием фенотипов, сходных с родительскими формами. 41,5 % мух было серых с зачаточными крыльями, как у одного исходного родителя, и 41,5 % особей черных длиннокрылых, как у другого исходного родителя. Только 17 % потомков родилось с новым сочетанием признаков: 8,5 % черных с зачаточными крыльями и 8,5 % серых длиннокрылых. Таким образом, 83 % потомков имели сочетание признаков, как у исходных родительских форм, но появились особи и с новым сочетанием признаков. Следовательно, сцепление является неполным. / Встал вопрос: почему появились особи с новой комбинацией родительских признаков? Для объяснения этого явления Морган использовал и развил теорию хиазмотипии бельгийского цитолога Янсенса. В 1909 г. Янсенс наблюдал, что при спермиогенезе у саламандры в профазе мейоза гомологичные хромосомы конъ-югируют, а затем, при начале расхождения, образуют фигуры в хромосомы обмениваются участками. Если сцепленные гены лежат в одной хромосоме и у гетерозигот при образовании гамет происходит рекомбинация этих генов, значит, гомологичные хромосомы во время мейоза обменялись своими частями. Обмен гомологичных хромосом своими частями называется перекрестом или крос-синговером (английское слово crossingover означает образование перекреста). Особей с новыми сочетаниями признаков, образовавшимися в результате кроссинговера, называют кроссоверами.
Вернемся к рисунку 14. У самки Fi, гетерозиготной по обеим парам признаков, в одной из гомологичных хромосом расположены гены Ъ и V , в другой - аллельные им гены В и v. В профазе редукционного деления, когда две гомологичные хромосомы соединились в один бивалент, каждая из хромосом удвоена и состоит из двух хроматид. Всего будет 4 хроматиды. Между двумя хроматидами гомологичных хромосом и происходит обмен их частями. В результате ген Ъ, расположенный в хроматиде одной гомологичной хромосомы, может соединиться с геном v, расположенным в хроматиде другой гомологичной хромосомы, и как результат одного события образуется вторая хроматида, где соединятся гены В и V . В дальнейшем хроматиды разойдутся и образуются кроссоверные гаметы с хромосомами с новым сочетанием генов (bv и BV ).
Две другие хроматиды из пары гомологичных хромосом не участвуют в перекресте и сохраняют в первоначальном сочетании материнские (bV ) и отцовские (Bv ) гены. Образование новых кроссоверных гамет обеспечило появление дрозофил -с новым сочетанием признаков: черных с зачаточными крыльями и серых длиннокрылых. Однако большая часть потомков будет сходна с исходными родителями (черные длиннокрылые и серые коротко-крылые). Морган приходит к выводу, что количество появления новых форм зависит от частоты перекреста, которая определяется по следующей формуле:
Если, например, общее число потомков 900, а новых кроссоверных форм 180, то частота перекреста будет составлять 20 %. Морган установил, что частота перекреста между определенной парой генов - относительно постоянная величина, но различная для разных пар генов. На основании этого был сделан вывод о том, что по частоте перекреста можно судить о расстояниях между генами. За единицу измерения перекреста принята его величина, равная 1 %. Иногда ее называют морганидой. Величина перекреста зависит от расстояния между изучаемыми генами. Чем больше отдалены гены друг от друга, тем чаще происходит перекрест; чем ближе они расположены, тем вероятность перекреста меньше. Установлено, что количество кроссоверных особей к общему числу потомков никогда не превышает 50 %, так как при очень больших расстояниях между генами чаще происходит двойной кроссинговер и часть кроссоверных особей остается неучтенной. Их можно учесть при изучении не двух пар сцепленных признаков, а трех или четырех. В этом случае, учитывая двойные и тройные перекресты, можно точнее судить о расстояниях и частоте перекреста между генами.
Блок информации.
Сцепление генов.
В начале 20-го века (1902-1907гг.) американским учёным У.Сеттоном и немецким эмбриологом Т.Бовери был обнаружен параллелизм в наследовании признаков и поведении хромосом клеточного ядра в процессе гаметогенеза и при оплодотворении. Это подтвердило локализацию наследственной информации в хромосомах. Установлено, что число генов значительно превышает число хромосом. Так у человека - 46 хромосом, а генов от 70 000 до 100 000. Следовательно, в каждой хромосоме локализовано большое количество генов. Гены одной хромосомы наследуются совместно (сцеплено). Экспериментальное исследование этого явления проведено американским генетиком Т.Морганом и его сотрудниками: А.Стертевантом, А.Бриджесом и Г.Мёллером в 1910-1916гг. Эти исследования подтвердили хромосомную локализацию генов и легли в основу хромосомной теории наследственности.
Основные положения хромосомной теории Наследственности.
1. Каждый ген занимает в хромосоме определённое место - локус.
2. Гены в хромосоме расположены линейно в определенной последовательности.
3. Различные хромосомы содержат неодинаковое число генов. Набор генов каждой негомологичной хромосомы уникален.
4. Гены одной хромосомы образуют " группу сцепления и наследуются вместе, т.е. сцеплено.
5. Число групп сцепления равно числу хромосом в гаплоидном наборе, (у дрозофилы их четыре, у кукурузы - 10, у мыши - 20, у человека -23).
6. Между гомологичными хромосомами может происходить обмен аллельными генами, т.е кроссинговер.
7. Частота кроссинговера прямо пропорциональна расстоянию между генами в группе сцепления.
8. За единицу расстояния между генами в группе сцепления принята особая единица - морганида (М). 1М=1% кроссинговера.
Различают полное и неполное сцепление генов.
Полное сцепление.
В экспериментах на дрозофиле было установлено, что развитие
признаков, которые наследуются сцеплено контролируется генами одной
хромосомы. Гены окраски тела (b - серой и B - черной) и длины крыльев
(v - нормальные и V - короткие, рудиментарные крылья) локализованы в одной паре гомологичных хромосом.
Скрещивание серых мух с нормальными крыльями и серых мух с рудиментарными крыльями дает в первом поколении серых гибридов с нормальными
При проведении анализирующего скрещивания, из р, были выбраны самцы, т. к. известно, что у самцов дрозофил ахиазматический сперматогенез (т. е. кроссинговер не происходит и полнота сцепления генов ничем не нарушается).В результате такого скрещивания на свет появляются особи двух фенотипов, аналогичные исходным родительским формам, причем в равных количествах: расщепление по фенотипу 1:1.
Рассматриваемые совместно результаты обоих скрещивании убеждают в том что развитие анализируемых признаков контролируется Разными генами, и сцепленное наследование объясняется локализацией генов в одной хромосоме. Полнота сцепления в данном случае ничем не нарушается. Такое сцепление генов является полным.
Для изучения неполного сцепления из Р, были выбраны самки (генотип В| |b) у С амок во время гаметогенеза происходит кроссинговер. Поэтому дигетерозиготная особь образует дополнительные, т.е. кроссоверные сорта гамет. Вероятность их образования обусловлена вероятностью кроссинговера, т.е. зависит от расстояния между генами в группе сцепления.
Не рекомбинантные особи; у Рекомбинантные особи; при
них наблюдаются такие же образовании их генотипов
сочетания, что и у исходных участвовали кроссоверные
родительских форм гаметы.
В данном примере расщепление по фенотипу в потомстве получено следующее: серых мух с длинными крыльями - 41,5%; черных с короткими крыльями - 41,5%; серых короткокрылых - 8,5%; черных длиннокрылых - 8,5 %. Таким образом, вероятность появления в потомстве рекомбинантных особей составляет 17%. Следовательно, расстояние между генами В и V в группе сцепления равно 17 морганиидам.
Преобладание в потомстве серых длиннокрылых и черных короткокрылых мух, указывает на то, что гены В и V; Ь и V действительно сцеплены. С другой стороны, появление рекомбинантных особей говорит о том, что в определенном числе случаев происходит разрыв сцепления между генами В и V и генами в и V. Это результат кроссинговера.
Примером полного сцепления генов у человека может служить наследование резус-фактора. Оно обусловлено тремя парами С, Д, К, тесно сцепленных между собой, поэтому наследование резус - принадлежности происходит по типу моногибридного скрещивания. Другим примером тесного сцепления генов у человека является наследование катаракты и полидактилии. Гены гемофилии и дальтонизма локализованы в Х - хромосоме на расстоянии 9,8 морганид (М), т.е. подвергаются кроссинговеру, поэтому наследуются как неполностью сцепленные. Аутососмные гены резус-фактора и формы эритроцитов, расположенные друг от друга на расстоянии 3 М и так же является примером неполного сцепления.
В 1909 Ф. Янссенс при изучении мейоза у земноводных оонаружил хиазмы (перекресты) хромосом, которые являются цитологическим свидетельством кроссинговера. С этого времени было предпринято множество попыток объяснить механизм этого явления. Существует несколько теорий кроссинговера. Наиболее распространенными являются две гипотезы.
Поурочное планирование 10 класс
Тема: «Полное и неполное сцепление генов. Генетические карты хромосом».
Цель урока: ознакомить учащихся с процессом полного и неполного сцепления генов; и сформировать представление о генетических картах хромосом.
Учебно – воспитательные задачи:
Раскрыть сущность явления сцепленного наследования генов.
Сформировать знания об основных положениях закона Т. Моргана.
Познакомить с принципом составления генетических карт
Развивать логическое мышления учащихся.
Оборудование, наглядные пособия : таблицы по общей биологии, иллюстрирующие сцепленное наследование генов и признаков, презентация к уроку, задачи на закрепление нового материала.
Тип урока: Урок изучения нового материала.
Методы: объяснительно - иллюстративный.
Ход урока:
I Организационный момент
Проверка личного состава учащихся и визуальной готовности класса к уроку.
II Проверка знаний учащихся
Фронтальный опрос:
1. Назовите три закона Г. Менделя?
2. Каких правил придерживался Г. Мендель при проведении своих опытов?
3. Сформулируйте закон чистоты гамет. Кому принадлежит открытие этого закона?
4. Всегда ли признаки можно чётко разделить на доминантные и рецессивные?
5. Какое название получило это явление?
6. Всегда ли по фенотипу можно определить, какие гены содержит данная особь? Приведите пример.
7. Можно ли установить генотип особей, которые не различаются по фенотипу? Какой метод используют для этого?
8. Какими особенностями характеризуется дигибридное скрещивание?
Молодцы! С этим этапом работы Вы справились
III Изучение нового материала:
Сцепленное наследование генов
Г.Мендель проследил наследование семи пар признаков у гороха. Многие исследователи, повторяя опыты Менделя, подтвердили открытые им законы. Было признано, что эти законы носят всеобщий характер. Однако в 1906 г. английские генетики В.Бэтсон и Р.Пеннет, проводя скрещивание растений душистого горошка и анализируя наследование формы пыльцы и окраски цветков, обнаружили, что эти признаки не дают независимого распределения в потомстве. Потомки всегда повторяли признаки родительских форм. Постепенно факты исключений из третьего закона Менделя накапливались. Стало ясно, что не для всех генов характерно независимое распределение в потомстве и свободное комбинирование.
Любой организм обладает многообразием морфологических, физиологических, биохимических и прочих признаков и свойств, причем каждый признак или свойство контролируется одним или несколькими генами, локализованными в хромосомах.
Однако если число генов организма огромно и может исчисляться десятками тысяч, то число хромосом сравнительно невелико и, как правило, измеряется несколькими десятками. Поэтому в каждой паре хромосом локализованы сотни и тысячи аллельных генов, образующих группы сцепления.
Установлено полное соответствие между числом групп сцепления и числом пар хромосом. Например, у кукурузы набор хромосом 2n = 20 и 10 групп сцепления, а у дрозофилы 2n = 8 и 4 группы сцепления, то есть число групп сцепления равно гаплоидному набору хромосом.
Закон Томаса Моргана
Гены, локализованные в одной хромосоме, передаются совместно, и способ их наследования отличается от наследования генов, локализованных в разных парах гомологичных хромосом.
Так, например, при независимом распределении хромосом дигибрид АаВb образует четыре типа гамет (АВ , аВ , Аb , аb ), а при условии полного сцепления такой же дигибрид даст только два типа гамет (АВ и аb ), так как эти гены расположены в одной хромосоме.
Разработка проблемы сцепленного наследования генов принадлежит школе Т.Моргана (1866–1945). Если Мендель проводил свои опыты на горохе, то для Моргана основным объектом стала плодовая мушка дрозофила. Мушка каждые две недели при температуре 25 °С дает многочисленное потомство. Самец и самка хорошо различимы – у самца брюшко меньше и темнее. Кроме того, они имеют различия по многочисленным признакам и могут размножаться в пробирках на дешевой питательной среде.
Изучая закономерности наследования генов, локализанных в одной и той же хромосоме , Морган пришел к выводу, что они наследуются сцепленно . Это и есть закон Т.Моргана.
Кроссинговер - процесс обмена участками во время в профазе I . Помимо мейотического, описан также . Хромосома разделяется на эти участки в определённых точках, одних и тех же для одного вида, что может быть определением вида на генетическом уровне, месторасположение этих точек задаётся единственным геном.
Поскольку кроссинговер вносит возмущения в картину , его удалось использовать для картирования «групп сцепления» (хромосом). Возможность картирования была основана на предположении о том, что, чем чаще наблюдается кроссинговер между двумя , тем дальше друг от друга расположены эти гены в группе сцепления и тем чаще будут наблюдаться отклонения от сцепленного наследования.
Полное и неполное сцепление
Для определения типа наследования двух пар генов (сцепленное или независимое) необходимо провести анализирующее скрещивание и по его результатам сделать вывод о характере наследования генов. Рассмотрим три возможных варианта результатов анализирующего скрещивания.
1) Независимое наследование .
Если в результате анализирующего скрещивания среди гибридов образуется четыре класса фенотипов, значит, гены наследуются независимо.
2) Полное сцепление генов .

При полном сцеплении генов
А
и
В
по результатам анализирующего скрещивания обнаруживают-
ся два фенотипических класса гибридов, полностью копирующих родителей.
3) Неполное сцепление генов .

В случае неполного сцепления генов А и В при анализирующем скрещивании появляются четыре фенотипа, два из которых имеют новое сочетание генов: Аb ‖ аb ; аВ ‖ аb . Появление подобных форм свидетельствует о том, что дигибрид с гаметами АВ │ и аb │ образует кроссоверные гаметы Аb │ и аВ │. Появление таких гамет возможно только в результате обмена участками гомологичных хромосом, то есть в процессе кроссинговера. Количество кроссоверных гамет значительно меньшее, чем некроссоверных.
Частота перекреста пропорциональна расстоянию между генами. Чем ближе расположены гены в хромосоме, тем теснее сцепление между ними и тем реже они разделяются при перекресте. И наоборот, чем дальше гены отстоят друг от друга, тем слабее сцепление между ними и чаще перекрест. Следовательно, о расстоянии между генами в хромосомах можно судить по частоте перекреста.
Генетические карты
Под генетическим картированием обычно понимают определение положения какого-либо гена по отношению к другим генам.
Рассмотрим порядок составления генетических карт.
1. Установление группы сцепления (то есть определение хромосомы, в которой локализован данный ген). Для этого необходимо иметь хотя бы по одному гену-маркеру в каждой группе сцепления.
2. Нахождение места локализации исследуемого гена в хромосоме. Для этого проводится скрещивание мутантной формы с нормальной и учитывается результат кроссинговера.
3. Определение расстояния между сцепленными генами, что позволяет составлять генетические карты хромосом, на которых указаны порядок расположения генов в хромосомах и относительные расстояния их друг от друга. Чем частота кроссинговера выше, тем на большем расстоянии друг от друга располагаются гены. Если установлено, что между сцепленными генами А и В частота кроссинговера 10%, а между генами В и С – 20%, то очевидно, что расстояние ВС в 2 раза больше, чем АВ . Расстояние между генами выражается в единицах, соответствующих 1% кроссинговера. Эти единицы называют морганидами.
Таким образом, на основе данных о частоте кроссинговера составляются генетические карты.
IV Закрепление знаний
Решение генетической задачи
Самку дрозофилы, гетерозиготную по рецессивным генам темной окраски тела и миниатюрных крыльев, скрестили с самцом, имевшим темное тело и миниатюрные крылья. От этого скрещивания было получено:
– 244 мухи с темным телом и миниатюрными крыльями;
– 20 мух с серой окраской тела и миниатюрными крыльями;
– 15 мух с темной окраской тела и нормальными крыльями;
– 216 мух с серой окраской тела и нормальными крыльями.
Исходя из приведенных данных определите, являются две эти пары генов сцепленными или нет. Как гены сцеплены?
Дано :
А
– серое тело
а
– темное тело
В
– нормальные крылья
b
– миниатюрные крылья
Характер наследования генов А и В – ?
Решение
Результаты расщепления среди гибридов (два фенотипических класса являются господствующими и повторяют фенотипически и генотипически родительские формы, а два других класса фенотипов представлены небольшим количеством особей) свидетельствуют о неполном сцепление генов §41, стр.161-164. Ответить на вопросы стр.165.
Задача №1. Дигетерозиготное растение гороха с гладкими семенами и усиками скрестили с растением с морщинистыми семенами без усиков. Известно, что оба доминантных гена (гладкие семена и наличие усиков) локализованы в одной хромосоме, кроссинговера не происходит. Составьте схему решения задачи. Определите генотипы родителей, фенотипы и генотипы потомства, соотношение особей с разными генотипами и фенотипами. Какой закон при этом проявляется?
А - гладкие семена, а - морщинистые семена
B - наличие усиков, b - без усиков
AB/ab
ab/ab
AB/ab
ab/ab
гладкие
семена,
усы
морщинист.
семена,
без усов
50%
50%
Если кроссинговер не происходит, то у дигетерозиготного родителя образуется только два вида гамет (полное сцепление).
Гены, расположенные в одной хромосоме, представляют собой группу сцепления . Сцепление генов - это совместное наследование генов, расположенных в одной и той же хромосоме. Количество групп сцепления соответствует гаплоидному числу хромосом. Сцепление генов, расположенных в одной хромосоме, может быть полным или неполным. Полное сцепление : Морган скрещивал черных длиннокрылых самок с серыми с зачаточными крыльями самцами. У дрозофилы серая окраска тела доминирует над черной, длиннокрылость - над зачаточными крыльями. Серое тело - А, черное тело а; длиннокрылые - В, зачаточные крылья - в. При спермиогенезе в период мейоза гомологичные хромосомы расходятся в разные половые клетки. 1) АА//АВ*ав//ав=4АВ//ав; 2) АВ//ав*АВ//ав=АВ//АВ, АВ//ав, ав//АВ, ав//ав. Если гены наход в аутосомах, то при полном сцеплении в F1 будет единообразие по фенотипу, а в F2 – 3:1, по скольким бы признакам не различались родители, т.к. изучается одна пара хромосом.
Явление неполного сцепления в наследовании признаков
В результате скрещивания потомки имели сочетание признаков, как у исходных родительских форм, но появились особи и с новым сочетанием признаков - сцепление неполное . В – серое, в – чёрное, V – нормальные, v - зачаточные. Bv||Bv*bV||bV=Bv||bV; самок из первого поколения скрестили с самцами анализаторами: BV//bV*bv//bv=Bv//bv,bV//bv – не кроссоверное. Bv//bV*bv//bv=2bv//bv, 2BV//bv – кросоверное. Обмен гомологичных хромосом своими частями называется перекрестом или кроссинговером. Особей с новыми сочетаниями признаков, образовавшимися в результате кроссинговера, называют кроссоверами. Количество появления новых форм зависит от частоты перекреста, которая определяется по следующей формуле: Частота перекрёста = (Число кроссоверных форм)·100/ Общее число потомков. За единицу измерения перекреста принята его величина, равная 1 %. Ее называют морганидой. Величина перекреста зависит от расстояния между изучаемыми генами. Чем больше отдалены гены друг от друга, тем чаще происходит перекрест; чем ближе они расположены, тем вероятность перекреста меньше.
Карты хромосом. Пример их построения
Карта хромосом - план расположения генов в хромосоме. Гены расположены в хромосомах в линейной последовательности на определенных расстояниях друг от друга. Явление торможения кроссинговера на одном участке кроссинговером на другом получило название интерференции. Чем меньше будет расстояние, разделяющее три гена, тем больше интерференция. Принимая во внимание линейное расположение генов в хромосоме, взяв за единицу расстояния частоту кроссинговера, Морган составили первую карту расположения генов в одной из хромосом дрозофилы: сh___13.6___ y___28.2___b. При построении карт указывают не расстояние между генами, а расстояние до каждого гена от нулевой точки начала хромосомы. Доминантный аллель обозначается прописной буквой, рецессивный - строчной. После построения генетических карт встал вопрос о том, отвечает ли расположение генов в хромосоме, построенное на основании частоты кроссинговера, истинному расположению. Каждая хромосома по длине имеет специфические рисунки дисков, что позволяет отличать разные ее участки друг от друга. Материалом для проверки служили хромосомы, у которых вследствие мутации возникали различные хромосомные перестройки: не хватало отдельных дисков, или они были перевернуты, или удвоены. Физические расстояния между генами на генетической карте не вполне соответствуют установленным цитологическим. Однако это не снижает ценности генетических карт хромосом для предсказания вероятности появления особей с новыми сочетаниями признаков. На основании анализа результатов многочисленных экспериментов с дрозофилой Т. Морган сформулировал хромосомную теорию наследственности, сущность которой заключается в следующем: 1) гены находятся в хромосомах, располагаются в них линейно на определенном расстоянии друг от друга; 2) гены, расположенные в одной хромосоме, относятся к одной группе сцепления. Число групп сцепления соответствует гаплоидному числу хромосом; З) признаки, гены которых находятся в одной хромосоме, наследуются сцеплено; 4) в потомстве гетерозиготных родителей новые сочетания генов, расположенных в одной паре хромосом, могут возникать в результате кроссинговера в процессе мейоза. Частота кроссинговера зависит от расстояния между генами; 5) на основании линейного расположения генов в хромосоме и частоты кроссинговера как показателя расстояния между генами можно построить карты хромосом.
Бисексуальность, интерсексуальность, гиандроморфизм, химеризм по половым хром-мам. Роль гормонов и условий среды в развит признаков пола
Любая зигота имеет х-хром-мы и аутосомы, т.е. имеет гены и женского и мужского пола, т.е. генетически любой организм бисексуальный (двуполый). Интерсексы – гермофродиты – особи с развитыми и женскими и мужскими признаками. 2 типа: истинные – имеют женские и мужские половые железы из-за нарушения баланса генов; условные – имеют железы одного пола, а наружный половой признак другого пола из-за нарушения баланса гормонов. Иногда у насекомых и животных встречается гиандроморфы – одна часть тела имеет женские признаки, а другая – мужские. Причины: зигота женского пола разделяется на 2 бластомера. Один из них потерял одну х-хром-му. Из этого бластомера будет развиваться мужская половина тела. Химеризм по половине хромосом хх/ху встречается у многоплодных животных, у бычков – когда в одном и том же организме содержатся хх- хромосомы, а воспроизводство ху- хромосом нарушено. При обычном кормлении вырастают самцы, а если в корм добавлять женские половые гормоны, то вырастают самки (рыбки мальки). Если личинка морского червя прикрепится ко дну моря – самка, если к хоботку самки – самец.
Типы детерминации пола у животных. Первичное и вторичное соотношение полов. Проблема регулирования пола
Детерминация обеспечивает образование равного кол-ва самцов и самок, что необходимо для нормального самовоспроизведения вида. Типы : 1) эпигамный – пол особи определяется в процессе онтогенеза, зависит от внешней среды. 2) прогамный – пол определяется в ходе гаметогенеза у родителей особи. 3) сингамный – пол определяется в момент слияния гамет. Первичное и вторичное соотношение полов: соотношение полов, кот определяется в момент слияния гамет, наз-ся первичным , всегда 1:1. Любое изменение в соотношении полов, как до, так и после рождения, наз-ся вторичным. Обычно после рождения оно смещается в пользу женского пола, поэтому у многих видов животных и у чел-ка мужских особей рождается больше, чем женских: кролики – 57%, человек – 51%, птицы – 59%. Проблема регулирования пола: имеет важное хозяйственное значение. Н-р: в молочном скотоводстве, в яичном птицеводстве желательны самки, а там, где основной продукт – мясо, лучше самцы. Проблема в том, чтобы разделить сперму на х- и у- фракции. Способы: 1) электрофорез – х – спермии имеют отрицательный заряд – движутся к катоду, а у – спермии – к аноду. Гарантия 80%. 2) Метод осаждения – х – сперма более плотная и осядает вниз, а у – остаётся сверху. 3) Использование набора кислот для изменения рН женских половых путей для создания условий только для х – или только для у-. 4) Партеногенез: геногенез – получение самок – рентгеновскими лучами облучают овоцит. первого порядка, тем самым задерживают расхождение хром-м, образ-ся яйцеклетка с диплоидным набором хром-м, в кот без оплодотворения развивается самка. Андрогенез – получение самцов – ядро яйцеклетки убивают лучами рентгена, затем в неё проникают два спермия, ядра сливаются, давая диплоидный набор, будет самец. 5) Метод разделения спермы на фракции по кол-ву ДНК в спермиях. 6) Чем моложе родители, тем вероятность рождения у них мужского пола больше. 7) Чем больше спермы в половых путях самки, тем вероятнее рождение мужского пола. 8) Чем больше хранится сперма – самка. 9) У птицы кормление: если петуху в корм добавлять Са, то самка, а если К – самцы. 10) В любой популяции действует закон равновесия, т.е. соотношение полов стремится 1:1.
Сцепленное наследование признаков. Анализ наследования одновременно нескольких признаков у дрозофилы, проведенный Т. Морганом, показал, что результаты анализирующего скрещивания гибридов Fi иногда отличаются от ожидаемых в случае их независимого наследования. У потомков такого скрещивания вместо свободного комбинирования признаков разных пар наблюдали тенденцию к наследованию преимущественно родительских сочетаний признаков. Такое наследование признаков было названо сцепленным. Сцепленное наследование объясняется расположением соответствующих генов в одной и той же хромосоме. В составе последней они передаются из поколения в поколение клеток и организмов, сохраняя сочетание аллелей родителей.
Зависимость сцепленного наследования признаков от локализации генов в одной хромосоме дает основание рассматривать хромосомы как отдельные группы сцепления.
При анализирующем скрещивании самцов из F 1 появлялось всего два вида потомков, сходных с родителями по сочетанию вариантов анализируемых признаков (серая окраска тела и нормальные крылья или черная окраска тела и короткие крылья) в соотношении 1:1. Это указывает на образование самцами F 1 всего двух типов гамет с одинаковой вероятностью, в которые попадают исходные родительские сочетания аллелей генов, контролирующих названные признаки (BV или bv).
При анализирующем скрещивании самок F 1 появлялось четыре вида потомков со всеми возможными сочетаниями признаков. При этом потомки с родительскими сочетаниями признаков встречались в 83%. У 17% потомков имелись новые комбинации признаков (серая окраска тела и короткие крылья или черная окраска тела и нормальные крылья). Видно, что и в этих скрещиваниях проявляется склонность к сцепленному наследованию либо доминантных признаков, либо рецессивных (83%). Частичное нарушение сцепления (17% потомков) было объяснено процессом кроссинговера - обменом соответствующими участками гомологичных хромосом в профазе I мейоза (см. рис. 3.72).
Из результатов скрещивания следует, что самки дрозофилы образуют четыре типа гамет, большинство из которых (83%) некроссоверные ((BV) и (bv)), 17% образуемых ими гамет появляются в результате кроссинговера и несут новые комбинации аллелей анализируемых генов ((Bv) и (bV)). Различия, наблюдаемые при скрещивании самцов и самок из F 1 с рецессивными гомозиготными партнерами объясняются тем, что по малопонятным причинам у самцов дрозофилы не происходит кроссинговера. В итоге самцы-дигетерозиготы по генам, расположенным в одной хромосоме, образуют два типа гамет. У самок кроссинговер имеет место и приводит к образованию некроссоверных и кроссоверных гамет, по два типа каждых. Поэтому в потомстве от анализирующего скрещивания появляется четыре фенотипа, два из которых обладают новыми по сравнению с родителями сочетаниями признаков.
Рис. 6.14. Сцепленное наследование признаков
(цвета тела и длины крыльев у дрозофилы):
I - скрещивание чистых линий, II, III - анализирующее скрещиваний самцов и самок из F 1 .
Изучение наследования других сочетаний признаков показало, что процент кроссоверного потомства для каждой пары признаков всегда один и тот же, но он различается для разных пар. Это наблюдение стало основанием для заключения, что гены в хромосомах расположены в линейном порядке. Выше отмечалось, что хромосома является группой сцепления определенных генов. Гомологичные хромосомы - это одинаковые группы сцепления, которые отличаются друг от друга лишь аллелями отдельных генов. При конъюгации гомологи сближаются своими аллельными генами, а при кроссинговере они обмениваются соответствующими участками. В результате появляются кроссоверные хромосомы с новым набором аллелей. Частота, с которой происходит обмен на участке между двумя данными генами, зависит от расстояния между ними(правило Т. Моргана). Процент кроссоверных гамет, косвенно отражает расстояние между генами. Это расстояние принято выражать в сантшморганидах. За одну сантиморганиду принимают расстояние между генами, при котором образуется 1% кроссоверного потомства (кроссоверных гамет).
При увеличении расстояния между генами увеличивается вероятность кроссинговера на участке между ними в клетках-предшественницах гамет. Так как в акте кроссинговера участвуют две хроматиды из четырех, присутствующих в биваленте, то даже в случае осуществления обмена между генами данной пары во всех клетках-предшественницах гамет процент кроссоверных половых клеток не может превысить 50 . Однако такая ситуация возможна лишь теоретически. Практически с увеличением расстояния между генами возрастает возможность прохождения одновременно нескольких кроссинговеров на данном участке (см. рис. 5.9). Так как каждый второй перекрест приводит к восстановлению прежнего сочетания аллелей в хромосоме, с увеличением расстояния число кроссоверных гамет может не увеличиваться, а уменьшаться. Из этого следует, что процент кроссоверных гамет является показателем истинного расстояния между генами лишь при достаточно близком их расположении, когда возможность второго кроссинговера исключается.
Нарушение сцепленного наследования родительских аллелей в результате кроссинговера позволяет говорить о неполном сцеплении в отличие от полного сцепления, наблюдаемого, например, у самцов дрозофилы.
Использование анализирующего скрещивания в опытах Т. Моргана показало, что с его помощью можно выяснять не только состав пар неаллельных генов, но и характер их совместного наследования. В случае сцепленного наследования признаков по результатам анализирующего скрещивания можно установить также расстояние между генами в хромосоме.

Рис. 6.15. Схема, поясняющая низкий процент кроссоверных гамет
(по отношению к двум данным генам)
Плюсом обозначены клетки-предшественницы гамет, в которых кроссинговер прошел на участке между двумя данными генами; зачернены кроссоверные гаметы
Генетические карты хромосом - это схема взаимного расположения и относительных расстояний между генами определенных хромосом, находящихся в одной группе сцепления.
Впервые на возможность построения генетических карт хромосом указывают Т. Морган и его сотрудники. Они экспериментально показали, что основываясь на явлениях сцепления генов и кроссинговера можно построить генетические карты хромосом . Возможность картирования основана на постоянстве процента кроссинговера между определенными генами. Генетические карты хромосом составлены для многих видов организмов: насекомых (дрозофила, комар, таракан и др.), грибов (дрожжи, аспергилл), для бактерий и вирусов.
Генетические карты человека используются в медицине при диагностике ряда тяжелых наследственных заболеваний человека. В исследованиях эволюционного процесса сравнивают генетические карты разных видов живых организмов.