{!LANG-e5fb3d2458f3bb2dd7f4d2eacd5306a8!}
{!LANG-bcd96800d38bbe57c8c2852d883448eb!}
{!LANG-e54a0bb1ba122b520e9fbb400d9db7b6!} {!LANG-af205327e20c84e5e0384e9dfb081091!}{!LANG-3b5c8b1650ba45fe1332bc71678117c0!}
{!LANG-70f790c63e7c131ebad176ef651666de!}
{!LANG-097b3f116f5b6e0c3618b73d524785ea!}
- {!LANG-55ee0482750a6e064604098ecfd5382f!}
- {!LANG-fe4b9bffbfe4e679f5442bdac83e17fd!}
{!LANG-82bf2656991b065b24709c4677dc1809!}
- {!LANG-9ed3999efeab1a7a8dfdd15990f9adbc!}
{!LANG-4488457b7de2a4093ddb5676a199b964!}
- {!LANG-0d203479aea601ea1f9fde21b98e51fb!}
- {!LANG-48292cf757a16cd018392a7c81e5f76d!}
{!LANG-caaa4871a0ab156c09feaf193834a8ae!}
{!LANG-46fc90bdce76c99cca594a8d9d934f96!}
{!LANG-4708436814a97e44bb17a9d696068cc1!}{!LANG-6e59e4c4ad46b885b25e71282bca543d!} {!LANG-5cfeaa832f5f4b4d9097b3a129457f6b!}
{!LANG-6043f9e0c253e2825500da2495f3dbf6!} {!LANG-d281a2e951982762dc9c08644aa955ee!}
{!LANG-03f1ebb25da567b7a012d65629792b26!}
{!LANG-641c8cb5f8a81c3a63314d5fd9dd0744!}{!LANG-46a897409006e2e376c9bffbd2211735!} {!LANG-240062525d3e9aa40ea36bf90060fd7d!}
A layer of gray matter 1 5 mm thick covering the cerebral hemispheres of mammals and humans. This part of the brain (see. Brain), which developed at the later stages of the evolution of the animal world, plays exclusively ... ... Great Soviet Encyclopedia
Large brain (cortex cerebri, PNA, LNH; substantia corticalis, BNA, JNA; synonym: K. cerebral hemispheres, K. brain, mantle, cloak) surface layer of the cerebral hemispheres, formed by its gray matter; plays an important role in ... ... Medical encyclopedia
- (cortex cerebri) gray matter located on the surface of the cerebral hemispheres and consisting of nerve cells (neurons), neuroglia, interneuronal connections of the cortex, and blood vessels. K. b. m. contains central (cortical) sections ... ... Medical encyclopedia
NEOCORTEX NEOCORTEX
(from neo ... and lat. cortex- bark, shell), new bark, neopallium, osn. part of the cerebral cortex. N. carries out the highest level of coordination of the brain and the formation of complex forms of behavior. In the course of evolution, N. first appears in reptiles, in which it is insignificant in size and is relatively simple in structure (the so-called lateral cortex). N. receives a typical multilayer structure only in mammals, in which it consists of 6-7 layers of cells (pyramidal, stellate, fusiform) and is subdivided into lobes: frontal, parietal, temporal, occipital, and mediobasal. In turn, the lobes are subdivided into regions, sub-regions and fields, differing in cellular structure and connections with the deep parts of the brain. Along with projection (vertical) fibers, N.'s neurons form associative (horizontal) fibers, to-rye in mammals and especially in humans are collected in anatomically expressed bundles (for example, the occipital-frontal bundle), providing simultaneous coordinated activity of dec. N.'s zones. As part of N., a naib is distinguished, a complexly constructed associative cortex, which in the process of evolution experiences the greatest increase, while N.'s primary sensory fields relatively decrease. (see the CEREAL BRAIN).
.(Source: "Biological Encyclopedic Dictionary." Ed. M. S. Gilyarov; Editorial board: A. A. Babaev, G. G. Vinberg, G. A. Zavarzin et al. - 2nd ed., Revised - M .: Sov.Encyclopedia, 1986.)
See what "NEOCORTEX" is in other dictionaries:
The neocortex ...
New cortex (synonyms: neocortex, isocortex) (lat. Neocortex) new areas of the cerebral cortex, which are only outlined in lower mammals, and in humans they constitute the main part of the cortex. The new crust is located in the upper layer of the hemispheres ... ... Wikipedia
neocortex - 3.1.15 neocortex: The new cerebral cortex that provides the implementation of intellectual mental activity by human thinking. 3.1.16 Source ... Dictionary-reference book of terms of normative and technical documentation
- (neocortex; neo + lat. cortex bark) see new bark ... Large Medical Dictionary
neocortex - at, h. Evolutionarily ninety and the most complex of nerve tissues, from which the foreheads, the yang, the skrone and powerful parts of the brain are stored ... Ukrainian Tlumachny vocabulary
{!LANG-4708436814a97e44bb17a9d696068cc1!}{!LANG-6e59e4c4ad46b885b25e71282bca543d!} {!LANG-5cfeaa832f5f4b4d9097b3a129457f6b!}
Archi, paleo, neocortex ... Spelling dictionary-reference
{!LANG-641c8cb5f8a81c3a63314d5fd9dd0744!}{!LANG-46a897409006e2e376c9bffbd2211735!} {!LANG-240062525d3e9aa40ea36bf90060fd7d!}
The term cortex refers to any outer layer of cells in the brain. The mammalian brain has three types of cortex: the pyriform cortex, which has olfactory functions; old bark (archicortex), constituting the main. part ... ... Psychological encyclopedia
By origin, the cerebral cortex is divided into ancient (pleocortex), old (archecortex), and new (neocortex). The ancient cortex includes structures associated with the analysis of olfactory stimuli; it includes olfactory bulbs, tracts, and tubercles. The old cortex includes the cingulate cortex, the hippocampal cortex, the dentate gyrus, and the amygdala. The ancient and old cortex forms the olfactory brain. In addition to smell, the olfactory brain provides alertness and attention reactions, takes part in the regulation of autonomic functions, plays a role in the formation of sexual, food, defensive instinctive behavior, and the provision of emotions.
All other structures of the cortex belong to the neocortex, which occupies about 96% of the total area of \u200b\u200bthe entire cortex.
The location of nerve cells in the cortex is denoted by the term "cytoarchitectonics". And conductive fibers - "myeloarchitectonics".
The new cortex consists of 6 cell layers that differ in cell composition, neural connections and functions. In the areas of the ancient bark and old bark, only 2-3 layers of cells are revealed. The neurons in the top four layers of the neocortex mainly process information from other parts of the nervous system. The main centrifugal layer is layer 5. The axons of its cells form the main descending pathways of the cerebral cortex, they conduct signals that control the work of the stem structures and the spinal cord.
Layer 1 is the outermost, molecular layer. It contains mainly nerve fibers deeper located neurons. In addition, it contains a small number of small cells. The fibers of the molecular layer form bonds between different areas of the cortex
2nd layer - outer granular. It contains a large number of small multipolar neurons. Part of the ascending dendrites from the third layer ends in this layer.
3rd layer - outer pyramidal. It is the widest, contains mainly medium and less often small and large pyramidal neurons. The dendrites of neurons from this layer are sent to the second layer.
4th layer - inner granular. Consists of a large number of small granular, as well as medium and large stellate cells. They are divided into two sublayers: 4a and 4b.
5th layer - ganglionic, or inner pyramidal. It is characterized by the presence of large pyramidal neurons. Their upward dendrites reach the molecular layer, while the basal and collateral axons are distributed in the fifth layer.
Layer 6 is polymorphic. It contains, along with cells of other forms, fusiform neurons. The shapes of other cells are very diverse: they have triangular, pyramidal, oval and polygonal shapes.
Cortex - the highest part of the central nervous system, which ensures the functioning of the body as a whole in its interaction with the environment.
brain (cerebral cortex, new cortex) is a layer of gray matter, consisting of 10-20 billion and covering the large hemispheres (Fig. 1). The gray matter of the bark makes up more than half of the total gray matter of the central nervous system. The total area of \u200b\u200bthe gray matter of the bark is about 0.2 m 2, which is achieved by the tortuous folding of its surface and the presence of grooves of different depths. The thickness of the cortex in its different parts ranges from 1.3 to 4.5 mm (in the anterior central gyrus). The neurons of the cortex are located in six layers oriented parallel to its surface.
In the areas of the cortex related to, there are zones with a three-layer and five-layer arrangement of neurons in the structure of the gray matter. These areas of the phylogenetically ancient cortex occupy about 10% of the surface of the cerebral hemispheres, the remaining 90% are new cortex.

Figure: 1. Mole of the lateral surface of the cerebral cortex (according to Brodman)
The structure of the cerebral cortex
The cerebral cortex has a six-layer structure
The neurons of different layers differ in cytological characteristics and functional properties.
Molecular layer - the most superficial. It is represented by a small number of neurons and numerous branching dendrites of pyramidal neurons lying in deeper layers.
Outer granular layer formed by densely spaced numerous small neurons of various shapes. The processes of the cells of this layer form corticocortical connections.
Outer pyramidal layer consists of medium-sized pyramidal neurons, the processes of which are also involved in the formation of corticocortical connections between adjacent areas of the cortex.
Inner granular layer similar to the second layer in terms of the type of cells and the arrangement of fibers. In the layer there are bundles of fibers connecting different parts of the cortex.
Signals from specific nuclei of the thalamus are transmitted to the neurons of this layer. The layer is very well represented in the sensory areas of the cortex.
Inner pyramids formed by medium and large pyramidal neurons. In the motor area of \u200b\u200bthe cortex, these neurons are especially large (50-100 microns) and are called giant, Betz pyramidal cells. The axons of these cells form fast-conducting (up to 120 m / s) fibers of the pyramidal tract.
Layer of polymorphic cells represented mainly by cells, the axons of which form corticothalamic pathways.

The neurons of the 2nd and 4th layers of the cortex are involved in the perception, processing of signals coming to them from the neurons of the associative areas of the cortex. Sensory signals from the switching nuclei of the thalamus come mainly to the neurons of the 4th layer, the severity of which is greatest in the primary sensory areas of the cortex. The neurons of the 1st and other layers of the cortex receive signals from other nuclei of the thalamus, basal ganglia, and the brain stem. The neurons of the 3rd, 5th and 6th layers form efferent signals that are sent to other areas of the cortex and along descending pathways to the lower parts of the central nervous system. In particular, layer 6 neurons form fibers that follow to the thalamus.
There are significant differences in the neuronal composition and cytological features of different parts of the cortex. Based on these differences, Brodmann divided the cortex into 53 cytoarchitectonic fields (see Fig. 1).
The location of many of these zeros, identified on the basis of histological data, coincides in topography with the location of the cortical centers, identified on the basis of their functions. Other approaches to dividing the cortex into regions are also used, for example, based on the content of certain markers in neurons, the nature of neuronal activity, and other criteria.
The white matter of the cerebral hemispheres is formed by nerve fibers. Allocate associative fibers,subdivided into arcuate fibers, but which signals are transmitted between neurons of adjacent convolutions and long longitudinal bundles of fibers that deliver signals to neurons in more distant sections of the hemisphere of the same name.
Commissural fibers - transverse fibers that transmit signals between neurons of the left and right hemispheres.
Projection fibers - conduct signals between neurons of the cortex and other parts of the brain.
The listed types of fibers are involved in the creation of neural circuits and networks, the neurons of which are located at considerable distances from each other. The cortex also contains a special kind of local neural circuits formed by adjacent neurons. These neural structures are called functional cortical columns. Neural columns are formed by groups of neurons located one above the other perpendicular to the surface of the cortex. The belonging of neurons to the same column can be determined by the increase in their electrical activity to stimulate the same receptive field. Such activity is recorded with a slow movement of the recording electrode in the cortex in the perpendicular direction. If the electrical activity of neurons located in the horizontal plane of the cortex is recorded, an increase in their activity is noted when various receptive fields are stimulated.
The diameter of the functional column is up to 1 mm. The neurons of one functional column receive signals from the same afferent thalamocortical fiber. Neurons of adjacent columns are connected to each other by processes, with the help of which they exchange information. The presence of such interconnected functional columns in the cortex increases the reliability of the perception and analysis of information coming to the cortex.
The efficiency of perception, processing and use of information by the cortex for the regulation of physiological processes is also ensured somatotopic principle of organization sensory and motor fields of the cortex. The essence of such an organization is that in a certain (projection) area of \u200b\u200bthe cortex, not any, but topographically outlined areas of the receptive field of the body surface, muscles, joints or internal organs are represented. So, for example, in the somatosensory cortex, the surface of the human body is projected in the form of a diagram, when at a certain point of the cortex the receptive fields of a specific area of \u200b\u200bthe body surface are presented. Efferent neurons are represented in a strict topographic manner in the primary motor cortex, the activation of which causes contraction of certain muscles of the body.
Bark fields also have screen principle of operation. In this case, the receptor neuron sends a signal not to a single neuron or to a single point of the cortical center, but to a network or zero of neurons connected by processes. The functional cells of this field (screen) are the columns of neurons.
The cerebral cortex, forming at the later stages of the evolutionary development of higher organisms, to a certain extent subordinated to itself all the lower parts of the central nervous system and is able to correct their functions. At the same time, the functional activity of the cerebral cortex is determined by the influx of signals from neurons of the reticular formation of the brain stem and signals from the receptive fields of the body's sensory systems.
Functional areas of the cerebral cortex
On a functional basis, sensory, associative and motor areas are distinguished in the cortex.
Sensory (sensitive, projection) areas of the cortex
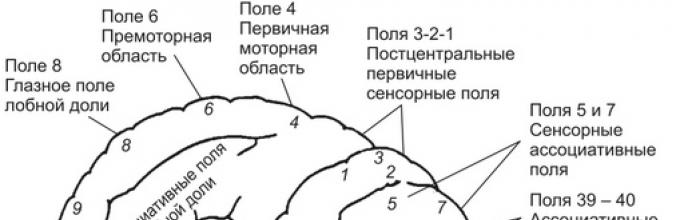
They consist of zones containing neurons, the activation of which by afferent impulses from sensory receptors or by direct exposure to stimuli causes the appearance of specific sensations. These zones are found in the occipital (fields 17-19), parietal (zero 1-3) and temporal (fields 21-22, 41-42) areas of the cortex.
In the sensory zones of the cortex, central projection fields are distinguished, which provide a slushy, clear perception of sensations of certain modalities (light, sound, touch, heat, cold) and secondary projection zeros. The function of the latter is to provide an understanding of the connection of the primary sensation with other objects and phenomena of the surrounding world.
The zones of representation of receptive fields in the sensory zones of the cortex largely overlap. A feature of the nerve centers in the area of \u200b\u200bthe secondary projection fields of the cortex is their plasticity, which is manifested by the possibility of restructuring specialization and restoring functions after damage to any of the centers. These compensatory capabilities of the nerve centers are especially pronounced in childhood. At the same time, damage to the central projection fields after suffering a disease is accompanied by a gross dysfunction of sensitivity and often the impossibility of its recovery.
Visual cortex
The primary visual cortex (VI, field 17) is located on both sides of the spur sulcus on the medial surface of the occipital lobe of the brain. In accordance with the identification of alternating white and dark stripes on unstained sections of the visual cortex, it is also called the striate (striped) cortex. The neurons of the lateral geniculate body send visual signals to the neurons of the primary visual cortex, which receive signals from the ganglion cells of the retina. The visual cortex of each hemisphere receives visual signals from the ipsilateral and contralateral halves of the retina of both eyes and their delivery to the neurons of the cortex is organized according to the somatotopic principle. The neurons that receive visual signals from photoreceptors are topographically located in the visual cortex, similar to receptors in the retina. Moreover, the area of \u200b\u200bthe macular retina has a relatively larger area of \u200b\u200brepresentation in the cortex than other areas of the retina.
The neurons of the primary visual cortex are responsible for visual perception, which, based on the analysis of input signals, is manifested by their ability to detect a visual stimulus, to determine its specific shape and orientation in space. In a simplified way, one can represent the sensory function of the visual cortex in solving a problem and answering the question of what a visual object is.
In the analysis of other qualities of visual signals (for example, location in space, movement, connections with other events, etc.), neurons of fields 18 and 19 of the extrastriatal cortex, located but adjacent to zero 17, take part. Information about signals received by sensory visual areas of the cortex, will be transferred for further analysis and use of vision to perform other brain functions in the associative areas of the cortex and other parts of the brain.
Auditory cortex
Located in the lateral groove of the temporal lobe in the area of \u200b\u200bthe Heschl gyrus (AI, fields 41-42). The neurons of the primary auditory cortex receive signals from the neurons of the medial geniculate bodies. The fibers of the auditory tract, which conduct sound signals to the auditory cortex, are organized tonotopically, and this allows the neurons of the cortex to receive signals from certain auditory receptor cells of the organ of Corti. The auditory cortex regulates the sensitivity of the auditory cells.
In the primary auditory cortex, sound sensations are formed and an analysis of the individual qualities of sounds is carried out, which makes it possible to answer the question of what the perceived sound is. The primary auditory cortex plays an important role in the analysis of short sounds, intervals between sound signals, rhythm, sound sequence. A more complex analysis of sounds is carried out in the associative areas of the cortex adjacent to the primary auditory. Based on the interaction of neurons in these areas of the cortex, binaural hearing is carried out, the characteristics of pitch, timbre, sound volume, the belonging of the sound are determined, and an idea of \u200b\u200ba three-dimensional sound space is formed.
Vestibular cortex
Located in the superior and middle temporal gyri (fields 21-22). Its neurons receive signals from neurons of the vestibular nuclei of the brainstem, connected by afferent connections with the receptors of the semicircular canals of the vestibular apparatus. In the vestibular cortex, a feeling is formed about the position of the body in space and the acceleration of movements. The vestibular cortex interacts with the cerebellum (through the temporocerebellar pathway), participates in the regulation of body balance, the adaptation of posture to the implementation of targeted movements. Based on the interaction of this area with the somatosensory and associative areas of the cortex, awareness of the body scheme occurs.
Olfactory cortex
Located in the region of the upper part of the temporal lobe (hook, zero 34, 28). The cortex includes a number of nuclei and belongs to the structures of the limbic system. Its neurons are located in three layers and receive afferent signals from the mitral cells of the olfactory bulb, connected by afferent connections with the olfactory receptor neurons. In the olfactory cortex, a primary qualitative analysis of smells is carried out and a subjective sense of smell, its intensity, and belonging is formed. Damage to the cortex leads to a decrease in the sense of smell or to the development of anosmia - loss of smell. When this area is artificially irritated, sensations of various odors appear, such as hallucinations.
Taste bark
Located in the lower part of the somatosensory gyrus, immediately anterior to the projection area of \u200b\u200bthe face (field 43). Its neurons receive afferent signals from relay neurons in the thalamus, which are associated with neurons in the nucleus of the solitary tract of the medulla oblongata. The neurons of this nucleus receive signals directly from sensory neurons that form synapses on the cells of taste buds. In the gustatory cortex, a primary analysis of the taste qualities of bitter, salty, sour, sweet is carried out, and on the basis of their summation, a subjective sensation of taste, its intensity, and belonging is formed.
The signals of smell and taste reach the neurons in the anterior part of the insular cortex, where, based on their integration, a new, more complex quality of sensations is formed, which determines our attitude to the sources of smell or taste (for example, to food).
Somatosensory cortex
Occupies the area of \u200b\u200bthe postcentral gyrus (SI, fields 1-3), including the paracentral lobule on the medial side of the hemispheres (Fig. 9.14). The somatosensory region receives sensory signals from thalamic neurons connected by spinothalamic pathways with skin receptors (tactile, temperature, pain sensitivity), proprioceptors (muscle spindles, bursae, tendons) and interoreceptors (internal organs).

Figure: 9.14. The most important centers and areas of the cerebral cortex
Due to the intersection of afferent pathways, signaling from the right side of the body arrives in the somatosensory zone of the left hemisphere, respectively, to the right hemisphere - from the left side of the body. In this sensory area of \u200b\u200bthe cortex, all parts of the body are somatotopically represented, but the most important receptive areas of the fingers, lips, skin of the face, tongue, larynx occupy relatively larger areas than the projections of such body surfaces as the back, front of the body, and legs.
The location of the representation of the sensitivity of body parts along the postcentral gyrus is often referred to as the "inverted homunculus", since the projection of the head and neck is in the lower part of the postcentral gyrus, and the projection of the caudal trunk and legs is in the upper part. In this case, the sensitivity of the legs and feet is projected onto the cortex of the paracentral lobule of the medial surface of the hemispheres. Within the primary somatosensory cortex, there is a certain specialization of neurons. For example, neurons of field 3 receive mainly signals from muscle spindles and mechanoreceptors of the skin, field 2 - from receptors of joints.
The cortex of the postcentral gyrus is referred to as the primary somatosensory region (SI). Its neurons send processed signals to neurons in the secondary somatosensory cortex (SII). It is located posterior to the postcentral gyrus in the parietal cortex (fields 5 and 7) and belongs to the associative cortex. SII neurons do not receive direct afferent signals from thalamic neurons. They are associated with SI neurons and neurons in other areas of the cerebral cortex. This makes it possible to carry out here an integral assessment of the signals entering the cortex along the spinothalamic pathway with signals coming from other (visual, auditory, vestibular, etc.) sensory systems. The most important function of these fields of the parietal cortex is the perception of space and the transformation of sensory signals into motor coordinates. In the parietal cortex, the desire (intention, urge) to carry out a motor action is formed, which is the basis for the beginning of planning the upcoming motor activity in it.

The integration of different sensory signals is associated with the formation of different sensations addressed to different parts of the body. These sensations are used both for the formation of mental and other responses, examples of which can be movements with the simultaneous participation of the muscles of both sides of the body (for example, moving, feeling with both hands, grabbing, unidirectional movement with both hands). The functioning of this area is necessary for recognizing objects by touch and determining the spatial location of these objects.
The normal function of the somatosensory areas of the cortex is an important condition for the formation of sensations such as heat, cold, pain and their addressing to a specific part of the body.
Damage to neurons in the primary somatosensory cortex leads to a decrease in various types of sensitivity on the opposite side of the body, and local damage to a loss of sensitivity in a certain part of the body. The discriminatory sensitivity of the skin is especially vulnerable when the neurons of the primary somatosensory cortex are damaged, and the least painful one. Damage to neurons in the secondary somatosensory area of \u200b\u200bthe cortex may be accompanied by impaired ability to recognize objects by touch (tactile agnosia) and skills in using objects (apraxia).
Motor areas of the cortex
About 130 years ago, researchers, applying point stimuli to the cerebral cortex with an electric current, found that exposure to the surface of the anterior central gyrus causes muscle contraction on the opposite side of the body. So the presence of one of the motor areas of the cerebral cortex was discovered. Later it turned out that several areas of the cerebral cortex and its other structures are related to the organization of movements, and in the areas of the motor cortex there are not only motor neurons, but also neurons performing other functions.
Primary motor cortexPrimary motor cortex located in the anterior central gyrus (MI, field 4). Its neurons receive the main afferent signals from neurons of the somatosensory cortex - fields 1, 2, 5, premotor cortex and thalamus. In addition, cerebellar neurons send signals to the MI through the ventrolateral thalamus.
The efferent fibers of the pyramidal pathway begin from the pyramidal neurons Ml. Some of the fibers of this pathway follow to the motor neurons of the cranial nerve nuclei of the brain stem (corticobulbar tract), some - to the neurons of the stem motor nuclei (red nucleus, nuclei of the reticular formation, stem nuclei associated with the cerebellum) and some - to inter- and motor neurons of the spinal cord brain (corticospinal tract).
There is a somatotopic organization of the arrangement of neurons in MI that control the contraction of different muscle groups of the body. The neurons that control the muscles of the legs and trunk are located in the upper parts of the gyrus and occupy a relatively small area, while the control muscles of the hands, especially the fingers, face, tongue and pharynx, are located in the lower areas and occupy a large area. Thus, in the primary motor cortex, a relatively large area is occupied by those neural groups that control muscles that carry out a variety of precise, small, finely regulated movements.
Since many Ml neurons increase electrical activity immediately before the onset of voluntary contractions, the primary motor cortex is assigned a leading role in controlling the activity of the motor nuclei of the trunk and motor neurons of the spinal cord and initiating voluntary, purposeful movements. Damage to the Ml field leads to muscle paresis and the impossibility of performing fine voluntary movements.
Secondary motor cortexIncludes areas of the premotor and accessory motor cortex (MII, field 6). Premotor cortex located in field 6, on the lateral surface of the brain, in front of the primary motor cortex. Its neurons receive afferent signals through the thalamus from the occipital, somatosensory, parietal associative, prefrontal regions of the cortex and cerebellum. Signals processed in it are sent by neurons of the cortex along efferent fibers to the motor cortex MI, a small number to the spinal cord and more to the red nuclei, nuclei of the reticular formation, basal ganglia and cerebellum. The premotor cortex plays a major role in the programming and organization of vision-controlled movements. The bark is involved in the organization of posture and auxiliary movements for actions carried out by the distal muscles of the limbs. Damage to the proximal cortex often causes a tendency to re-execute the initiated movement (perseveration), even if the movement performed has reached the goal.
In the lower part of the premotor cortex of the left frontal lobe, immediately anterior to the area of \u200b\u200bthe primary motor cortex, which contains the neurons that control the muscles of the face, is located speech area, or the motor center of Broca's speech. Violation of its function is accompanied by impaired speech articulation, or motor aphasia.
Additional motor cortex located in the upper part of field 6. Its neurons receive afferent signals from the somatosensory, parietal and prefrontal regions of the cerebral cortex. Signals processed in it are sent by neurons of the cortex along efferent fibers to the primary motor cortex MI, spinal cord, and stem motor nuclei. The activity of neurons in the additional motor cortex increases earlier than neurons in the MI cortex, mainly due to the implementation of complex movements. At the same time, the increase in neural activity in the additional motor cortex is not associated with movements as such; for this, it is enough to mentally imagine a model of the upcoming complex movements. The additional motor cortex takes part in the formation of the program of upcoming complex movements and in the organization of motor responses to the specificity of sensory stimuli.
Since neurons in the secondary motor cortex send many axons to the MI field, it is considered a higher structure in the hierarchy of motor centers of motor organization, standing above the motor centers of the MI motor cortex. The nerve centers of the secondary motor cortex can influence the activity of motor neurons in the spinal cord in two ways: directly through the corticospinal pathway and through the MI field. Therefore, they are sometimes referred to as supra-motor fields, the function of which is to instruct the centers of the MI field.
It is known from clinical observations that the maintenance of the normal function of the secondary motor cortex is important for the implementation of precise hand movements, and especially for the performance of rhythmic movements. So, for example, if they are damaged, the pianist ceases to feel the rhythm and maintain the interval. The ability to carry out opposite hand movements is impaired (manipulation with both hands).
With simultaneous damage to the MI and MII motor zones of the cortex, the ability to fine coordinated movements is lost. Point irritations in these areas of the motor zone are accompanied by the activation not of individual muscles, but of a whole group of muscles that cause directional movement in the joints. These observations gave rise to the conclusion that the motor cortex contains not so much muscles as movements.
Prefrontal cortexIt is located in the area of \u200b\u200bfield 8. Its neurons receive the main afferent signals from the occipital visual, parietal associative cortex, upper hillocks of the quadruple. The processed signals are transmitted along efferent fibers to the premotor cortex, the upper hillocks of the quadruple, and the brainstem motor centers. The cortex plays a decisive role in the organization of movements under the control of vision and is directly involved in the initiation and control of eye and head movements.
The mechanisms that implement the transformation of the concept of movement into a specific motor program, into bursts of impulses sent to specific muscle groups, remain insufficiently understood. It is believed that the intention of movement is formed due to the functions of the associative and other areas of the cortex that interact with many structures of the brain.
Information about the intention of movement is transmitted to the motor areas of the frontal cortex. The motor cortex through the descending pathways activates systems that ensure the development and use of new motor programs or the use of old ones, already worked out in practice and stored in memory. The basal ganglia and cerebellum are part of these systems (see their functions above). Movement programs developed with the participation of the cerebellum and basal ganglia are transmitted through the thalamus to the motor zones and, above all, to the primary motor cortex. This area directly initiates the execution of movements, connecting certain muscles to it and providing a sequence of alternating their contraction and relaxation. The commands of the cortex are transmitted to the motor centers of the brainstem, spinal motoneurons and motoneurons of the cranial nerve nuclei. In the implementation of movements, motor neurons play the role of the final path through which motor commands are transmitted directly to the muscles. Features of signal transmission from the cortex to the motor centers of the trunk and spinal cord are described in the chapter on the central nervous system (brain stem, spinal cord).
Associative areas of the cortex
In humans, the associative areas of the cortex occupy about 50% of the area of \u200b\u200bthe entire cerebral cortex. They are located in the areas between the sensory and motor areas of the cortex. Associative areas do not have clear boundaries with secondary sensory areas, both in morphological and functional features. The parietal, temporal and frontal associative areas of the cerebral cortex are distinguished.
Parietal associative area of \u200b\u200bthe cortex. Located in fields 5 and 7 of the superior and inferior parietal lobes of the brain. The area is bordered in front by the somatosensory cortex, and behind - by the visual and auditory cortex. The neurons of the parietal associative region can receive and activate their visual, sound, tactile, proprioceptive, painful, signals from the memory apparatus and other signals. Some neurons are polysensory and can increase their activity upon receipt of somatosensory and visual signals. However, the degree of increase in the activity of neurons in the associative cortex to the receipt of afferent signals depends on the current motivation, attention of the subject, and information retrieved from memory. It remains insignificant if the signal coming from the sensory regions of the brain is indifferent to the subject, and it increases significantly if it coincided with the existing motivation and attracted his attention. For example, when a monkey is presented with a banana, the activity of neurons in the associative parietal cortex remains low if the animal is full, and vice versa, the activity increases sharply in hungry animals that like bananas.
The neurons of the parietal associative cortex are connected by efferent connections with the neurons of the prefrontal, premotor, motor regions of the frontal lobe and cingulate gyrus. Based on experimental and clinical observations, it is generally accepted that one of the functions of the cortex of field 5 is the use of somatosensory information for the implementation of purposeful voluntary movements and manipulation of objects. The function of the cortex of field 7 is the integration of visual and somatosensory signals to coordinate eye movements and visually guided hand movements.
Disruption of these functions of the parietal associative cortex when its connections with the frontal lobe cortex are damaged or a disease of the frontal lobe itself explains the symptoms of the consequences of diseases localized in the parietal associative cortex. They can be manifested by difficulty in understanding the semantic content of signals (agnosia), an example of which is the loss of the ability to recognize the shape and spatial location of an object. The processes of transformation of sensory signals into adequate motor actions may be disrupted. In the latter case, the patient loses the skills of practical use of well-known tools and objects (apraxia), and he may develop the inability to carry out visually guided movements (for example, moving the hand in the direction of the object).
Frontal associative area of \u200b\u200bthe cortex. It is located in the prefrontal cortex, which is part of the frontal lobe cortex located anterior to fields 6 and 8. Neurons in the frontal associative cortex receive processed sensory signals via afferent connections from neurons in the cortex of the occipital, parietal, temporal lobes of the brain and from neurons in the cingulate gyrus. The frontal associative cortex receives signals about the current motivational and emotional states from the nuclei of the thalamus, limbic and other brain structures. In addition, the frontal cortex can operate with abstract, virtual signals. The associative frontal cortex sends efferent signals back to the brain structures from which they were received, to the motor areas of the frontal cortex, the caudate nucleus of the basal ganglia and the hypothalamus.
This area of \u200b\u200bthe cortex plays a primary role in the formation of the higher mental functions of a person. It provides the formation of target attitudes and programs of conscious behavioral reactions, recognition and semantic assessment of objects and phenomena, understanding of speech, logical thinking. After extensive damage to the frontal cortex, patients may develop apathy, a decrease in the emotional background, a critical attitude towards their own actions and the actions of others, complacency, a violation of the ability to use past experience to change behavior. Patient behavior can become unpredictable and inadequate.
The temporal associative area of \u200b\u200bthe cortex. It is located in fields 20, 21, 22. Cortex neurons receive sensory signals from neurons of the auditory, extrastriatal visual and prefrontal cortex, hippocampus and amygdala.
After a bilateral disease of the temporal associative areas with the involvement of the hippocampus in the pathological process or connections with it, patients may develop pronounced memory impairments, emotional behavior, inability to concentrate (distraction). In some people, if the lower temporal region is damaged, where the center of face recognition is presumably located, visual agnosia may develop - the inability to recognize the faces of familiar people, objects, while maintaining vision.
On the border of the temporal, visual and parietal areas of the cortex in the lower parietal and posterior parts of the temporal lobe, there is an associative area of \u200b\u200bthe cortex, called the sensory center of speech, or Wernicke's center. After its damage, a violation of the speech understanding function develops while the speech motor function is preserved.
Neocortex - evolutionarily the youngest part of the cortex, occupying most of the surface of the hemispheres. Its thickness in humans is approximately 3 mm.
The cellular composition of the neocorhex is very diverse, but about three quarters of the neurons of the cortex are pyramidal neurons (pyramids), in connection with which one of the main classifications of neurons of the cortex divides them into pyramidal and non-iramidal (fusiform, stellate, granular, candelabra cells, Martinotti cells, etc. .). Another classification is related to the length of the axon (see paragraph 2.4). Longaxon Golgi I cells are mainly pyramids and spindles, their axons can exit the cortex, the rest of the cells are shortaxon Golgi II cells.
Cortical neurons also differ in the size of the cell body: the size of ultrasmall neurons is 6x5 µm, the size of giant ones is more than 40 x 18. The largest neurons are Betz pyramids, their size is 120 x 30-60 µm.
Pyramidal neurons (see Fig. 2.6, d) have the shape of a body in the form of a pyramid, the top of which is directed upwards. An apical dendrite departs from this apex and rises into the overlying cortical layers. Basal dendrites extend from the rest of the soma. All dendrites have spines. A long axon departs from the base of the cell, forming numerous collaterals, including recurrent ones, which bend and rise up. Stellate cells have no apical dendrite; spines on dendrites are absent in most cases. In spindle-shaped cells, two large dendrites extend from opposite poles of the body, there are also small dendrites extending from the rest of the body. Dendrites have spines. The axon is long, with little branching.
During embryonic development, the new cortex necessarily goes through the stage of a six-layer structure; during maturation in some areas, the number of layers may decrease. The deeper layers are phylogenetically older, the outer layers are younger. Each layer of the cortex is characterized by its own neuronal composition and thickness, which in different areas of the cortex may differ from each other.
We list new crust layers (Figure 9.8).
I layer - molecular - the outermost, contains a small number of neurons and mainly consists of fibers running parallel to the surface. Dendrites of neurons located in the underlying layers also rise here.
Layer II - outer granular, or outer granular, - consists mainly of small pyramidal neurons and a small number of medium-sized stellate cells.
III layer - outer pyramidal - the widest and thickest layer, contains mainly small and medium-sized pyramidal and stellate neurons. Large and giant pyramids are located deep in the layer.
IV layer - internal granular, or internal granular, - consists mainly of small neurons of all varieties, there are also a few large pyramids.
V layer - inner pyramidal, or ganglionic, a characteristic feature of which is the presence of large and in some areas (mainly in fields 4 and 6; Fig. 9.9; subparagraph 9.3.4) - giant pyramidal neurons (Betz pyramids). The apical dendrites of the pyramids, as a rule, reach layer I.
VI layer - polymorphic, or multiforme, - contains mainly fusiform neurons, as well as cells of all other forms. This layer is divided into two sublayers, which some researchers consider as independent layers, speaking in this case about a seven-layer crust.
Figure: 9.8.
and - neurons are colored entirely; b - only the bodies of neurons are painted; in - painted
only outgrowths of neurons
Main functions each layer is also different. Layers I and II carry out connections between neurons of different layers of the cortex. Callosal and associative fibers mainly come from the pyramids of layer III and come to layer II. The main afferent fibers entering the cortex from the thalamus terminate in layer IV neurons. Layer V is mainly associated with the system of descending projection fibers. The axons of the pyramids of this layer form the main efferent pathways of the cerebral cortex.
In most cortical areas, all six layers are equally well expressed. Such a crust is called homotypic. However, in some fields, the severity of the layers may change during development. This bark is called heterotypic. It is of two types:
granular (zero 3, 17, 41; Fig. 9.9), in which the number of neurons in the outer (II) and especially in the inner (IV) granular layers is greatly increased, as a result of which the IV layer is divided into three sublayers. This cortex is characteristic of the primary sensory zones (see below);
Agranular (fields 4 and 6, or motor and premotor cortex; Fig. 9.9), in which, on the contrary, there is a very narrow layer II and practically no IV, but very wide pyramidal layers, especially the inner one (V).







