- จัดการ โดยกลไกการควบคุม: อิเล็กโทร - คีโมและควบคุมโดยกลไก
- ไม่สามารถควบคุมได้ พวกเขาไม่มีกลไกประตูและเปิดอยู่ตลอดเวลาไอออนจะทำงานอย่างต่อเนื่อง แต่ช้า
ศักยภาพในการพักผ่อน คือความแตกต่างของศักย์ไฟฟ้าระหว่างภายนอกและ สภาพแวดล้อมภายใน เซลล์.
พักผ่อนกลไกการก่อตัวที่อาจเกิดขึ้น สาเหตุทันทีของศักยภาพในการพักคือความเข้มข้นของแอนไอออนและไอออนบวกที่ไม่เท่ากันทั้งภายในและภายนอกเซลล์ ประการแรกการจัดเรียงของไอออนนั้นขึ้นอยู่กับความแตกต่างของการซึมผ่าน ประการที่สองโพแทสเซียมไอออนออกจากเซลล์มากกว่าโซเดียม
ศักยภาพในการดำเนินการ - นี่คือการกระตุ้นของเซลล์ความผันผวนอย่างรวดเร็วในศักยภาพของเมมเบรนเนื่องจากการแพร่กระจายของไอออนเข้าและออกจากเซลล์
เมื่อสิ่งกระตุ้นกระทำต่อเซลล์ของเนื้อเยื่อที่กระตุ้นได้ในตอนแรกช่องโซเดียมจะถูกกระตุ้นและปิดใช้งานอย่างรวดเร็วจากนั้นด้วยความล่าช้าช่องโพแทสเซียมจะเปิดใช้งานและปิด
ด้วยเหตุนี้ไอออนจึงแพร่เข้าหรือออกจากเซลล์อย่างรวดเร็วตามการไล่ระดับสีด้วยไฟฟ้าเคมี นี่คือความตื่นเต้น ตามการเปลี่ยนแปลงขนาดและสัญลักษณ์ของประจุไฟฟ้ามีสามเฟสที่แตกต่างกัน:
- ระยะที่ 1 - การลดขั้ว การลดประจุของเซลล์เป็นศูนย์ โซเดียมเคลื่อนที่เข้าหาเซลล์ตามความเข้มข้นและการไล่ระดับสีทางไฟฟ้า สภาพการเคลื่อนไหว: ประตูของช่องโซเดียมเปิดอยู่
- ระยะที่ 2 - การผกผัน เครื่องหมายการกลับรายการ การผกผันเกี่ยวข้องกับสองส่วน: ขึ้นและลง
จากน้อยไปมาก โซเดียมยังคงเคลื่อนที่เข้าสู่เซลล์ตามการไล่ระดับความเข้มข้น แต่ตรงกันข้ามกับการไล่ระดับสีทางไฟฟ้า (มันรบกวน)
จากมากไปหาน้อย โพแทสเซียมเริ่มออกจากเซลล์ตามความเข้มข้นและการไล่ระดับสีทางไฟฟ้า ประตูของช่องโพแทสเซียมเปิดอยู่
- ระยะที่ 3 - การเปลี่ยนขั้ว โพแทสเซียมยังคงออกจากเซลล์ตามความเข้มข้น แต่ตรงกันข้ามกับการไล่ระดับสีทางไฟฟ้า
เกณฑ์สำหรับความตื่นเต้นเร้าใจ
ด้วยการพัฒนาศักยภาพในการออกฤทธิ์ทำให้เกิดการเปลี่ยนแปลงความตื่นเต้นของเนื้อเยื่อ การเปลี่ยนแปลงนี้เกิดขึ้นในระยะ สถานะของโพลาไรเซชันเมมเบรนเริ่มต้นมีลักษณะเฉพาะสะท้อนให้เห็นถึงศักยภาพของเมมเบรนที่พักซึ่งสอดคล้องกับสถานะเริ่มต้นของความสามารถในการปลุกปั่นและด้วยเหตุนี้สถานะเริ่มต้นของเซลล์ที่กระตุ้นได้ นี่เป็นระดับความตื่นเต้นปกติ ช่วงเวลาก่อนการบัดกรีเป็นช่วงเวลาที่จุดเริ่มต้นของศักยภาพในการดำเนินการ ความตื่นเต้นของเนื้อเยื่อเพิ่มขึ้นเล็กน้อย ความสามารถในการปลุกปั่นระยะนี้เป็นความสูงส่งขั้นต้น (ความสามารถในการปลุกปั่นเหนือธรรมชาติหลัก) ในระหว่างการพัฒนาพรีฟิวชันศักยภาพของเมมเบรนจะเข้าใกล้ระดับวิกฤตของการลดขั้วและเพื่อให้ถึงระดับนี้ความแรงของสิ่งกระตุ้นอาจน้อยกว่าเกณฑ์
ในระหว่างการพัฒนาของ spike (ศักยภาพสูงสุด) การไหลของโซเดียมไอออนเข้าไปในเซลล์คล้ายหิมะถล่มเกิดขึ้นซึ่งเป็นผลมาจากการที่เมมเบรนชาร์จใหม่และสูญเสียความสามารถในการตอบสนองด้วยการกระตุ้นต่อสิ่งเร้าที่มีกำลังเหนือกว่า ระยะนี้เรียกว่าการหักเหของแสงสัมบูรณ์เช่น non-excitability แน่นอนซึ่งคงอยู่จนกว่าจะสิ้นสุดการชาร์จเมมเบรน การหักเหของแสงสัมบูรณ์ของเมมเบรนเกิดขึ้นเนื่องจากช่องโซเดียมถูกเปิดออกอย่างสมบูรณ์และถูกปิดใช้งาน
หลังจากสิ้นสุดระยะการชาร์จไฟความสามารถในการปลุกปั่นจะค่อยๆกลับคืนสู่ระดับเดิม - นี่คือเฟสของการหักเหของแสงสัมพัทธ์นั่นคือ ความไม่ตื่นเต้นสัมพัทธ์ จะดำเนินต่อไปจนกว่าประจุเมมเบรนจะกลับคืนสู่ค่าที่สอดคล้องกับระดับวิกฤตของการลดขั้ว เนื่องจากในช่วงเวลานี้ยังไม่ได้รับการฟื้นฟูศักยภาพของพังผืดความตื่นเต้นของเนื้อเยื่อจะลดลงและการกระตุ้นใหม่สามารถเกิดขึ้นได้ภายใต้การกระทำของสิ่งกระตุ้นที่เหนือกว่าเท่านั้น ความสามารถในการปลุกปั่นที่ลดลงในระยะของการหักเหเชิงสัมพัทธ์มีความสัมพันธ์กับการปิดใช้งานช่องโซเดียมบางส่วนและการกระตุ้นช่องโพแทสเซียม
ช่วงเวลาต่อไปสอดคล้องกับระดับความตื่นเต้นที่เพิ่มขึ้น: ระยะของความสูงส่งรองหรือความตื่นเต้นเหนือธรรมชาติรอง เนื่องจากศักยภาพของเมมเบรนในระยะนี้อยู่ใกล้กับระดับวิกฤตของการลดขั้วเมื่อเปรียบเทียบกับสถานะพักของโพลาไรเซชันเริ่มต้นเกณฑ์การกระตุ้นจึงลดลงเช่น ความตื่นเต้นของเซลล์เพิ่มขึ้น ในระยะนี้ความตื่นเต้นใหม่ ๆ สามารถเกิดขึ้นได้ภายใต้การกระทำของสิ่งเร้าของกองกำลังย่อย ช่องโซเดียมจะไม่ถูกปิดใช้งานอย่างสมบูรณ์ในช่วงนี้ ศักยภาพของเมมเบรนเพิ่มขึ้น - เกิดภาวะเมมเบรน hyperpolarization เมื่อก้าวออกจากระดับวิกฤตของการลดขั้วเกณฑ์ของการกระตุ้นจะเพิ่มขึ้นเล็กน้อยและการกระตุ้นใหม่จะเกิดขึ้นได้ภายใต้การกระทำของสิ่งเร้าที่มีขนาดเหนือระดับเท่านั้น
กลไกของเยื่อหุ้มที่มีศักยภาพ
เซลล์แต่ละเซลล์ที่เหลือมีลักษณะการปรากฏตัวของความต่างศักย์ของเมมเบรน (ศักยภาพในการพัก) โดยทั่วไปความแตกต่างของประจุไฟฟ้าระหว่างพื้นผิวด้านในและด้านนอกของเมมเบรนอยู่ระหว่าง -80 ถึง -100 mV และสามารถวัดได้โดยใช้ไมโครอิเล็กโทรดภายนอกและภายในเซลล์ (รูปที่ 1)
ความต่างศักย์ระหว่างด้านนอกและด้านในของเยื่อหุ้มเซลล์ในสภาวะพักตัวเรียกว่า ศักยภาพของเมมเบรน (ศักยภาพในการพักผ่อน)
การสร้างศักยภาพในการพักผ่อนจัดทำโดยกระบวนการหลัก 2 กระบวนการคือการกระจายไอออนอนินทรีย์ที่ไม่สม่ำเสมอระหว่างช่องว่างภายในและภายนอกเซลล์และการซึมผ่านของเยื่อหุ้มเซลล์ที่ไม่เท่ากัน การวิเคราะห์องค์ประกอบทางเคมีของของเหลวนอกเซลล์และภายในเซลล์บ่งชี้การกระจายของไอออนที่ไม่สม่ำเสมออย่างมาก (ตารางที่ 1)
ในส่วนที่เหลือมีแอนไอออนของกรดอินทรีย์และ K + ไอออนอยู่ภายในเซลล์ซึ่งมีความเข้มข้นสูงกว่าภายนอก 30 เท่า ในทางตรงกันข้าม Na + ไอออนอยู่นอกเซลล์มากกว่าภายใน 10 เท่า CI- มีขนาดใหญ่กว่าด้านนอกด้วย
ในส่วนที่เหลือเยื่อหุ้มเซลล์ประสาทจะซึมผ่านได้มากที่สุดสำหรับ K + น้อยกว่าสำหรับ CI และการซึมผ่านของ Na + น้อยมาก / ความสามารถในการซึมผ่านของเยื่อหุ้มใยประสาทสำหรับ Na + B ขณะพักน้อยกว่า K + ถึง 100 เท่า สำหรับแอนไอออนของกรดอินทรีย์จำนวนมากเมมเบรนที่อยู่นิ่งจะไม่สามารถผ่านได้อย่างสมบูรณ์
รูป: 1. การวัดศักยภาพในการพักผ่อนของเส้นใยกล้ามเนื้อ (A) โดยใช้ไมโครอิเล็กโทรดภายในเซลล์: M - ไมโครอิเล็กโทรด; และ - อิเล็กโทรดที่ไม่แยแส ลำแสงบนหน้าจอออสซิลโลสโคป (B) แสดงให้เห็นว่าก่อนที่ไมโครอิเล็กโทรดจะเจาะเมมเบรนความต่างศักย์ระหว่าง M และ I เป็นศูนย์ ในช่วงเวลาของการเจาะ (แสดงด้วยลูกศร) ตรวจพบความแตกต่างที่อาจเกิดขึ้นซึ่งบ่งชี้ว่าด้านในของเมมเบรนมีประจุลบเมื่อเทียบกับพื้นผิวด้านนอก (อ้างอิงจาก B.I.Khodorov)
ตาราง. ความเข้มข้นของไอออนของเซลล์กล้ามเนื้อภายในและภายนอกเซลล์ในสัตว์เลือดอุ่น mmol / l (อ้างอิงจาก J. Dudel)
|
ความเข้มข้นภายในเซลล์ |
ความเข้มข้นภายนอกเซลล์ |
|
|
A- (แอนไอออนของสารประกอบอินทรีย์) |
เนื่องจากการไล่ระดับความเข้มข้น K + จะเข้าสู่ผิวด้านนอกของเซลล์โดยมีประจุบวก แอนไอออนที่มีน้ำหนักโมเลกุลสูงไม่สามารถติดตาม K + ได้เนื่องจากไม่สามารถซึมผ่านไปยังเมมเบรนได้ Na + ion ไม่สามารถแทนที่โพแทสเซียมไอออนด้านซ้ายได้เนื่องจากความสามารถในการซึมผ่านของเยื่อหุ้มเซลล์นั้นต่ำกว่ามาก СI-ตามการไล่ระดับความเข้มข้นสามารถผสมได้เฉพาะภายในเซลล์ดังนั้นจึงเพิ่มประจุลบของพื้นผิวด้านในของเมมเบรน เนื่องจากการเคลื่อนที่ของไอออนนี้โพลาไรเซชันของเมมเบรนเกิดขึ้นเมื่อผิวด้านนอกมีประจุบวกและพื้นผิวด้านในมีประจุลบ
สนามไฟฟ้าที่สร้างขึ้นบนเมมเบรนขัดขวางการกระจายตัวของไอออนระหว่างเนื้อหาภายในและภายนอกของเซลล์ เมื่อประจุบวกบนพื้นผิวด้านนอกของเซลล์เพิ่มขึ้นทำให้ K + ไอออนซึ่งเป็นประจุบวกเคลื่อนที่จากด้านในไปด้านนอกได้ยากขึ้นเรื่อย ๆ เขาเคลื่อนไหวราวกับขึ้นเขา ยิ่งค่าของประจุบวกที่ผิวด้านนอกมากเท่าใดปริมาณของ K + ไอออนก็จะไปที่ผิวเซลล์ได้น้อยลง ที่ค่าหนึ่งของศักยภาพบนเมมเบรนจำนวน K + ไอออนที่ข้ามเมมเบรนในทิศทางใดทิศทางหนึ่งจะมีค่าเท่ากันเช่น การไล่ระดับความเข้มข้นของโพแทสเซียมมีความสมดุลโดยศักยภาพบนเมมเบรน ศักยภาพที่ฟลักซ์การแพร่กระจายของไอออนจะเท่ากับฟลักซ์ของไอออนที่เคลื่อนที่ไปในทิศทางตรงกันข้ามเรียกว่าศักย์สมดุลของไอออนที่กำหนด สำหรับ K + ไอออนศักย์สมดุลคือ -90 mV ในเส้นใยประสาทชนิดไมอีลินด์ค่าของศักยภาพสมดุลของ CI- ไอออนจะใกล้เคียงกับค่าของศักยภาพเยื่อหุ้มเซลล์ (-70 mV) ดังนั้นแม้ว่าความเข้มข้นของСI-ไอออนภายนอกเส้นใยจะสูงกว่าภายใน แต่ก็ไม่พบกระแสทางเดียวตามการไล่ระดับความเข้มข้น ในกรณีนี้ความแตกต่างของความเข้มข้นจะสมดุลกับศักยภาพบนเมมเบรน
Na + ion ตามการไล่ระดับความเข้มข้นจะต้องเข้าสู่เซลล์ (ศักยภาพสมดุลคือ +60 mV) และการมีประจุลบภายในเซลล์ไม่ควรขัดขวางการไหลนี้ ในกรณีนี้ Na + ที่เข้ามาจะทำให้ประจุลบภายในเซลล์เป็นกลาง อย่างไรก็ตามสิ่งนี้ไม่ได้เกิดขึ้นจริงเนื่องจากเมมเบรนที่อยู่นิ่งไม่สามารถซึมผ่าน Na + ได้ไม่ดี
กลไกที่สำคัญที่สุดที่สนับสนุนความเข้มข้นของ Na + ไอออนภายในเซลล์ต่ำและ K + ไอออนที่มีความเข้มข้นสูงคือปั๊มโซเดียม - โพแทสเซียม (การขนส่งที่ใช้งานอยู่) เป็นที่ทราบกันดีว่ามีระบบของพาหะในเยื่อหุ้มเซลล์ซึ่งแต่ละส่วนจะจับกับโกลนโดยไอออน Na + ภายในเซลล์และกำจัดออกไปภายนอก จากภายนอกผู้ให้บริการจะจับกับ K + สองไอออนนอกเซลล์ซึ่งจะถูกพาเข้าไปในไซโทพลาสซึม แหล่งจ่ายไฟสำหรับการทำงานของระบบพาหะจัดทำโดย ATP การทำงานของปั๊มในระบบดังกล่าวนำไปสู่ผลลัพธ์ต่อไปนี้:
- k + ไอออนที่มีความเข้มข้นสูงภายในเซลล์จะได้รับการบำรุงรักษาซึ่งทำให้มั่นใจได้ถึงความคงที่ของศักยภาพในการพักผ่อน เนื่องจากความจริงที่ว่าในหนึ่งรอบของการแลกเปลี่ยนไอออนอีกหนึ่งไอออนบวกจะถูกกำจัดออกจากเซลล์มากกว่าที่จะนำมาใช้การขนส่งแบบแอคทีฟจึงมีบทบาทในการสร้างศักยภาพในการพักผ่อน ในกรณีนี้พวกเขาพูดถึงปั๊มไฟฟ้าเนื่องจากตัวมันเองสร้างขนาดเล็ก แต่ กระแสตรง. ประจุบวกจากเซลล์ดังนั้นจึงมีส่วนช่วยโดยตรงในการก่อตัวของศักยภาพเชิงลบภายในเซลล์ อย่างไรก็ตามการมีส่วนร่วมของปั๊มอิเล็กโตรจีนิกต่อมูลค่ารวมของศักยภาพในการพักผ่อนมักจะมีขนาดเล็กและมีปริมาณหลายมิลลิโวลต์
- รักษาความเข้มข้นของ Na + ไอออนไว้ในระดับต่ำภายในเซลล์ซึ่งในอีกด้านหนึ่งจะช่วยให้มั่นใจได้ถึงการทำงานของกลไกในการสร้างศักยภาพในการดำเนินการในอีกด้านหนึ่งทำให้มั่นใจได้ถึงการบำรุงรักษาออสโมลาริตีและปริมาณเซลล์ตามปกติ
- ด้วยการรักษาระดับความเข้มข้นของ Na + ให้คงที่ปั๊มโซเดียม - โพแทสเซียมจะส่งเสริมการขนส่ง K +, Na + ที่เชื่อมต่อกันของกรดอะมิโนและน้ำตาลผ่านเยื่อหุ้มเซลล์
ดังนั้นการเกิดขึ้นของความต่างศักย์ของทรานส์เมมเบรน (ศักยภาพในการพักตัว) เกิดจากการนำเยื่อหุ้มเซลล์ที่มีค่าความนำไฟฟ้าสูงสำหรับ K +, CI- ไอออน, ความไม่สมดุลของไอออนิกของความเข้มข้นของ K + และ CI- ไอออน, การทำงานของระบบขนส่งที่ใช้งานอยู่ (Na + / K + -ATPase), ซึ่งสร้างและรักษาความไม่สมมาตรของไอออนิก
ศักยภาพในการทำงานของใยประสาทแรงกระตุ้นของเส้นประสาท
ศักยภาพในการดำเนินการ - นี่คือความผันผวนในระยะสั้นของความต่างศักย์ของเมมเบรนของเซลล์ที่เคลื่อนไหวได้พร้อมกับการเปลี่ยนแปลงของเครื่องหมายประจุ
ศักยภาพในการดำเนินการเป็นสัญญาณหลักของการปลุกเร้าอารมณ์ การลงทะเบียนระบุว่าเซลล์หรือโครงสร้างของมันตอบสนองต่อการกระตุ้น อย่างไรก็ตามตามที่ระบุไว้แล้ว PD ในเซลล์บางเซลล์สามารถเกิดขึ้นเองได้ (ตามธรรมชาติ) เซลล์ดังกล่าวมีอยู่ในเครื่องกระตุ้นหัวใจผนังหลอดเลือด ระบบประสาท... PD ถูกใช้เป็นตัวพาข้อมูลโดยส่งสัญญาณในรูปแบบของสัญญาณไฟฟ้า (การส่งสัญญาณไฟฟ้า) ไปตามเส้นใยประสาทที่ส่งผลและระบบการทำงานของหัวใจรวมทั้งเริ่มหดตัวของเซลล์กล้ามเนื้อ
ให้เราพิจารณาสาเหตุและกลไกของการสร้าง AP ในเส้นใยประสาทที่สร้างตัวรับประสาทสัมผัสหลัก สาเหตุทันทีของการโจมตี (การสร้าง) ของ AP ในพวกเขาคือศักยภาพของตัวรับ
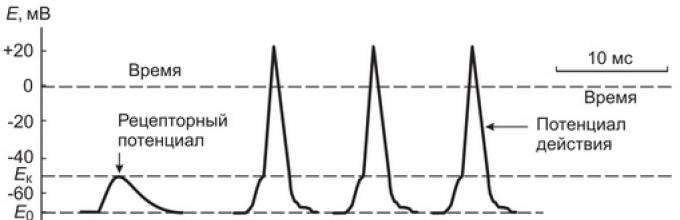
ถ้าเราวัดความแตกต่างที่อาจเกิดขึ้นบนเมมเบรนของการสกัดกั้น Ranvier ที่ใกล้กับจุดสิ้นสุดของเส้นประสาทมากที่สุดในช่วงระหว่างผลกระทบต่อแคปซูลของ Pacini corpuscle จะยังคงไม่เปลี่ยนแปลง (70 mV) และในระหว่างที่เกิดการกระทบนั้นจะทำการแยกขั้วเกือบพร้อมกันกับการลดขั้วของเยื่อรับของปลายประสาท
ด้วยการเพิ่มขึ้นของแรงกดบนร่างกาย Pacini ทำให้ศักยภาพในการรับเพิ่มขึ้นถึง 10 mV ในการสกัดกั้นที่ใกล้ที่สุดของ Ranvier โดยปกติจะมีการบันทึกการสั่นอย่างรวดเร็วของศักยภาพของเมมเบรนพร้อมกับการเติมเมมเบรน - ศักยภาพในการออกฤทธิ์ (AP) หรือแรงกระตุ้นของเส้นประสาท (รูปที่ 2) หากแรงกดดันต่อร่างกายเพิ่มมากขึ้นความกว้างของศักยภาพของตัวรับจะเพิ่มขึ้นและศักยภาพในการกระทำจำนวนมากจะถูกสร้างขึ้นในเส้นประสาทที่ลงท้ายด้วยความถี่ที่แน่นอน

รูป: 2. การแสดงแผนผังของกลไกการเปลี่ยนแปลงของศักยภาพตัวรับเป็นศักยภาพในการกระทำ (กระแสประสาท) และการแพร่กระจายของแรงกระตุ้นไปตามเส้นใยประสาท
สาระสำคัญของกลไกของการสร้าง AP คือศักยภาพของตัวรับทำให้เกิดกระแสหมุนเวียนในท้องถิ่นระหว่างเยื่อหุ้มตัวรับที่ไม่ได้โพลาไรซ์ของส่วนที่ไม่ได้รับการตัดแต่งของปลายประสาทและเยื่อของการสกัดกั้นครั้งแรกของ Ranvier กระแสเหล่านี้ซึ่งถูกพัดพาโดยไอออน Na +, K +, CI- และไอออนของแร่ธาตุอื่น ๆ ไม่เพียง แต่ "ไหล" ไปตาม แต่ยังข้ามเยื่อหุ้มของใยประสาทในพื้นที่ของการสกัดกั้นของ Ranvier ด้วย ในเมมเบรนของการสกัดกั้นของ Ranvier ตรงกันข้ามกับเมมเบรนรับของเส้นประสาทที่สิ้นสุดตัวเองมีความหนาแน่นสูงของช่องโซเดียมและโพแทสเซียมที่ขึ้นกับแรงดันไฟฟ้าไอออนิก
เมื่อถึงค่าดีโพลาไรเซชันประมาณ 10 mV บนเมมเบรนของการสกัดกั้น Ranvier ช่องโซเดียมที่ขึ้นกับแรงดันไฟฟ้าอย่างรวดเร็วจะถูกเปิดออกและผ่านการไหลของไอออนของ Na + จะไหลเข้าสู่แอกโซพลาสซึมตามการไล่ระดับทางเคมีไฟฟ้า ทำให้เกิดการลดขั้วอย่างรวดเร็วและการเติมพลังของเมมเบรนสกัดกั้น Ranvier อย่างไรก็ตามในเวลาเดียวกันกับการเปิดช่องโซเดียมที่ขึ้นกับแรงดันไฟฟ้าอย่างรวดเร็วในเมมเบรนสกัดกั้นของ Ranvier ช่องโพแทสเซียมที่ขึ้นกับแรงดันไฟฟ้าช้าจะเปิดออกและไอออน K + เริ่มออกจาก axoilasm เอาต์พุตของพวกมันจะล่าช้าเมื่อเทียบกับอินพุตของ Na + ไอออน ดังนั้นไอออนของ Na + ที่เข้าสู่แอกโซพลาสซึมในอัตราที่สูงจะทำการดีโพลาไรซ์อย่างรวดเร็วและชาร์จเมมเบรนเป็นเวลาสั้น ๆ (0.3-0.5 มิลลิวินาที) ในขณะที่ไอออน K + ที่ส่งออกจะคืนค่าการกระจายตัวของประจุเดิมบนเมมเบรน (เปลี่ยนขั้วเมมเบรน) เป็นผลให้ในระหว่างการดำเนินการทางกลกับร่างกาย Pacini ด้วยแรงที่เท่ากับหรือเกินเกณฑ์จะสังเกตเห็นความผันผวนระยะสั้นของศักยภาพบนเมมเบรนของการสกัดกั้น Ranvier ที่ใกล้ที่สุดในรูปแบบของการลดขั้วอย่างรวดเร็วและการเปลี่ยนขั้วของเมมเบรนเช่น PD (กระแสประสาท) ถูกสร้างขึ้น
เนื่องจากสาเหตุทันทีของการสร้าง AP คือศักยภาพของตัวรับในกรณีนี้จึงเรียกว่าศักยภาพของเครื่องกำเนิดไฟฟ้า จำนวนกระแสประสาทที่มีแอมพลิจูดและระยะเวลาเท่ากันที่สร้างขึ้นต่อหนึ่งหน่วยเวลานั้นเป็นสัดส่วนกับแอมพลิจูดของศักยภาพของตัวรับดังนั้นแรงกดดันต่อตัวรับ กระบวนการแปลงข้อมูลเกี่ยวกับความแรงของผลกระทบที่ฝังอยู่ในแอมพลิจูดของศักยภาพของตัวรับเป็นจำนวนของกระแสประสาทที่ไม่ต่อเนื่องเรียกว่าการเข้ารหัสข้อมูลแบบไม่ต่อเนื่อง
กลไกไอออนิกและพลวัตชั่วขณะของกระบวนการสร้าง AP ได้รับการศึกษาในรายละเอียดเพิ่มเติมภายใต้เงื่อนไขการทดลองภายใต้การกระทำเทียมบนเส้นใยประสาทด้วยกระแสไฟฟ้าที่มีความแข็งแรงและระยะเวลาต่างๆกัน
ลักษณะของศักยภาพในการออกฤทธิ์ของใยประสาท (กระแสประสาท)
เมมเบรนของเส้นใยประสาทที่จุดของการแปลอิเล็กโทรดที่ระคายเคืองตอบสนองต่อผลกระทบของกระแสไฟฟ้าที่อ่อนมากซึ่งยังไม่ถึงค่าเกณฑ์ คำตอบนี้เรียกว่าโลคัลและความผันผวนของความต่างศักย์ทั่วเมมเบรนเรียกว่าศักย์ท้องถิ่น
การตอบสนองเฉพาะที่บนเมมเบรนของเซลล์ที่กระตุ้นได้อาจนำหน้าการเกิดขึ้นของการกระทำที่อาจเกิดขึ้นหรือเกิดขึ้นเป็นกระบวนการที่เป็นอิสระ มันเป็นความผันผวนในระยะสั้น (การลดขั้วและการเปลี่ยนขั้ว) ของศักยภาพในการพักซึ่งไม่ได้มาพร้อมกับการชาร์จแบบเมมเบรน Depolarization ของเมมเบรนในระหว่างการพัฒนาศักยภาพในท้องถิ่นเกิดจากการเข้าสู่แอกโซพลาสซึมของ Na + ไอออนขั้นสูงและการแบ่งขั้วเกิดจากการออกจากแกนของ K + ไอออนล่าช้า
หากคุณกระทำกับเมมเบรนด้วยกระแสไฟฟ้าที่มีแรงเพิ่มขึ้นค่านี้เรียกว่าขีด จำกัด การลดขั้วของเมมเบรนอาจถึงระดับวิกฤต - E k ซึ่งช่องโซเดียมที่ขึ้นกับแรงดันไฟฟ้าอย่างรวดเร็วจะเปิดออก ด้วยเหตุนี้จึงมีการไหลของ Na + ไอออนเข้าสู่เซลล์เพิ่มขึ้นเช่นเดียวกับหิมะถล่ม กระบวนการที่เกิดจากการลดขั้วทำให้เกิดลักษณะที่เร่งตัวเองและศักยภาพในท้องถิ่นจะพัฒนาไปสู่ศักยภาพในการดำเนินการ
ได้มีการกล่าวไว้แล้วว่า คุณลักษณะเฉพาะ PD คือการผกผัน (การเปลี่ยนแปลง) ระยะสั้นของเครื่องหมายของประจุบนเมมเบรน ภายนอกจะกลายเป็นประจุลบในช่วงเวลาสั้น ๆ (0.3-2 มิลลิวินาที) และภายใน - เป็นบวก ขนาดของการผกผันอาจสูงถึง 30 mV และขนาดของศักยภาพการกระทำทั้งหมด - 60-130 mV (รูปที่ 3)
ตาราง. ลักษณะเปรียบเทียบ ศักยภาพในท้องถิ่นและศักยภาพในการดำเนินการ
|
ลักษณะเฉพาะ |
ศักยภาพในท้องถิ่น |
ศักยภาพในการดำเนินการ |
|
การนำไฟฟ้า |
กระจายในพื้นที่ 1-2 มม. พร้อมการลดทอน (ลดลง) |
แพร่กระจายโดยไม่มีการลดทอนเป็นระยะทางไกลตลอดความยาวทั้งหมดของใยประสาท |
|
กฎแห่ง "แรง" |
ส่ง |
ไม่เชื่อฟัง |
|
กฎหมายทั้งหมดหรือไม่มีอะไร |
ไม่เชื่อฟัง |
ส่ง |
|
ปรากฏการณ์ Summation |
มันสะสมเพิ่มขึ้นพร้อมกับการระคายเคืองที่เกิดขึ้นบ่อยๆซ้ำ ๆ |
ไม่สะสม |
|
แอมพลิจูด |
||
|
ความสามารถในการปลุกปั่น |
เพิ่มขึ้น |
ลดความสามารถในการไม่ปลุกปั่น (การหักเหของแสง) |
|
ขนาดสิ่งกระตุ้น |
Subthreshold |
เกณฑ์และขีด จำกัด สูงสุด |
ศักยภาพในการดำเนินการขึ้นอยู่กับลักษณะของการเปลี่ยนแปลงของประจุบนพื้นผิวด้านในของเมมเบรนแบ่งออกเป็นขั้นตอนของการดีโพลาไรซ์การแบ่งขั้วและการเกิดไฮโพลาไรเซชันของเมมเบรน Depolarization เรียกส่วนจากน้อยไปหามากทั้งหมดของ AP ซึ่งพื้นที่ที่สอดคล้องกับศักยภาพในท้องถิ่น (จากระดับ E 0 ก่อน E ถึง), Depolarization อย่างรวดเร็ว (จากระดับ E ถึง ถึงระดับ 0 mV) การผกผัน เครื่องหมายของประจุ (จาก 0 mV ถึงค่าสูงสุดหรือจุดเริ่มต้นของการเปลี่ยนขั้ว) Repolarization เรียกว่าส่วนที่ลดลงของ AP ซึ่งสะท้อนถึงกระบวนการฟื้นฟูโพลาไรเซชันของเมมเบรนเริ่มต้น ในตอนแรกการเปลี่ยนขั้วจะดำเนินการอย่างรวดเร็ว แต่เข้าใกล้ระดับ E 0ความเร็วอาจช้าลงและส่วนนี้เรียกว่า ติดตามการปฏิเสธ (หรือติดตามศักยภาพเชิงลบ) ในบางเซลล์หลังจาก repolarization การเกิด hyperpolarization จะพัฒนาขึ้น พวกเขาโทรหาเธอ ติดตามศักยภาพเชิงบวก
เรียกอีกอย่างว่าส่วนที่ไหลเร็วแอมพลิจูดสูงเริ่มต้นของ PD จุดสูงสุด หรือ ขัดขวาง รวมถึงขั้นตอนของการลดขั้วและการเปลี่ยนขั้วอย่างรวดเร็ว
ในกลไกของการพัฒนา PD บทบาทที่สำคัญที่สุดคือช่องไอออนที่ขึ้นกับแรงดันไฟฟ้าและการเพิ่มขึ้นของการซึมผ่านของเยื่อหุ้มเซลล์โดยไม่พร้อมกันสำหรับไอออน Na + และ K + ดังนั้นเมื่อกระแสไฟฟ้ากระทำต่อเซลล์จะทำให้เกิดการลดขั้วของเยื่อหุ้มเซลล์และเมื่อประจุเมมเบรนลดลงถึงระดับวิกฤต (E ถึง) ช่องโซเดียมที่ขึ้นกับแรงดันไฟฟ้าจะเปิดขึ้น ดังที่ได้กล่าวไปแล้วช่องเหล่านี้เกิดขึ้นจากโมเลกุลของโปรตีนที่ฝังอยู่ในเมมเบรนซึ่งภายในมีรูพรุนและกลไกประตูสองอัน หนึ่งในกลไกประตู - ตัวกระตุ้นหนึ่งให้ (ด้วยการมีส่วนร่วมของส่วนที่ 4) การเปิด (การเปิดใช้งาน) ของช่องระหว่างการลดขั้วของเมมเบรนและที่สอง (ด้วยการมีส่วนร่วมของลูปภายในเซลล์ระหว่างโดเมนที่ 3 และ 4) - การปิดใช้งานซึ่งพัฒนาขึ้นระหว่างการชาร์จเมมเบรนใหม่ (รูปที่. 4). เนื่องจากกลไกทั้งสองนี้เปลี่ยนตำแหน่งของประตูช่องอย่างรวดเร็วช่องโซเดียมที่มีแรงดันไฟฟ้าจึงเป็นช่องไอออนที่รวดเร็ว สถานการณ์นี้มีความสำคัญอย่างยิ่งต่อการสร้าง AP ในเนื้อเยื่อที่เคลื่อนไหวได้และการนำผ่านเยื่อของเส้นประสาทและเส้นใยกล้ามเนื้อ

รูป: 3. ศักยภาพของการกระทำเฟสและกระแสไอออนิก (a, o) คำอธิบายเป็นข้อความ

รูป: 4. ตำแหน่งของประตูและสถานะของกิจกรรมของช่องโซเดียมและโพแทสเซียมที่มีแรงดันไฟฟ้าในระดับต่างๆของโพลาไรซ์เมมเบรน
เพื่อให้ช่องโซเดียมที่มีแรงดันไฟฟ้าผ่าน Na + ไอออนเข้าสู่เซลล์จำเป็นต้องเปิดประตูกระตุ้นเท่านั้นเนื่องจากประตูปิดการใช้งานจะเปิดภายใต้สภาวะพัก สิ่งนี้เกิดขึ้นเมื่อการลดขั้วของเมมเบรนถึงระดับ E ถึง (รูปที่ 3, 4)
การเปิดประตูเปิดใช้งานของช่องโซเดียมทำให้โซเดียมเข้าสู่เซลล์เหมือนหิมะถล่มซึ่งได้รับแรงหนุนจากการกระทำของกองกำลังของการไล่ระดับสีไฟฟ้าเคมี เนื่องจากไอออน Na + มีประจุบวกจึงทำให้ประจุลบส่วนเกินที่อยู่บนพื้นผิวด้านในของเมมเบรนเป็นกลางลดความต่างศักย์ของเมมเบรนและลดขั้ว ในไม่ช้าไอออน Na + จะส่งประจุบวกส่วนเกินไปยังพื้นผิวด้านในของเมมเบรนซึ่งมาพร้อมกับการผกผัน (การเปลี่ยนแปลง) ของเครื่องหมายประจุจากลบเป็นบวก
อย่างไรก็ตามช่องโซเดียมจะยังคงเปิดอยู่ประมาณ 0.5 ms เท่านั้นและหลังจากช่วงเวลานี้นับจากจุดเริ่มต้น
AP ปิดประตูปิดการใช้งานช่องโซเดียมจะถูกปิดใช้งานและไม่สามารถซึมผ่านของไอออน Na + ได้ซึ่งการเข้าสู่เซลล์จะถูก จำกัด อย่างรวดเร็ว
จากช่วงเวลาของการลดขั้วของเมมเบรนไปจนถึงระดับ E ถึง นอกจากนี้ยังมีการกระตุ้นช่องโพแทสเซียมและการเปิดประตูของ K + ไอออน K + ไอออนภายใต้การกระทำของกองกำลังของการไล่ระดับความเข้มข้นให้ออกจากเซลล์โดยใช้ประจุบวกจากมัน อย่างไรก็ตามกลไกประตูของช่องโพแทสเซียมทำงานช้าและอัตราการปลดปล่อยประจุบวกด้วย K + ไอออนจากเซลล์สู่ภายนอกล่าช้าเมื่อเทียบกับทางเข้าของไอออน Na + การไหลของ K + ไอออนลบประจุบวกส่วนเกินออกจากเซลล์ทำให้เกิดการฟื้นฟูการกระจายประจุเริ่มต้นบนเมมเบรนหรือการเปลี่ยนขั้วและประจุลบจะถูกเรียกคืนที่ด้านในหลังจากช่วงเวลาหนึ่งจากช่วงเวลาของการชาร์จ
การปรากฏตัวของ AP บนเมมเบรนที่เคลื่อนไหวได้และการฟื้นฟูในภายหลังของศักยภาพการพักเริ่มต้นบนเมมเบรนเป็นไปได้เนื่องจากพลวัตของการเข้าและออกจากเซลล์ของประจุบวกของไอออน Na + และ K + นั้นแตกต่างกัน การเข้ามาของ Na + ion อยู่ก่อนทางออกของ K + ion ในเวลา หากกระบวนการเหล่านี้อยู่ในสภาวะสมดุลความต่างศักย์ของเมมเบรนจะไม่เปลี่ยนแปลง การพัฒนาความสามารถในการกระตุ้นและสร้าง AP โดยกล้ามเนื้อและเซลล์ประสาทที่เคลื่อนไหวได้เกิดจากการก่อตัวในเยื่อหุ้มเซลล์ของช่องไอออนความเร็วต่างกันสองประเภท ได้แก่ โซเดียมเร็วและโพแทสเซียมช้า
การสร้าง AP เดียวต้องการการเข้าสู่เซลล์ของ Na + ไอออนจำนวนค่อนข้างน้อยซึ่งไม่รบกวนการกระจายภายนอกและภายในเซลล์ เมื่อมีการสร้าง AP จำนวนมากการกระจายของไอออนทั้งสองด้านของเยื่อหุ้มเซลล์อาจถูกรบกวน อย่างไรก็ตามภายใต้สภาวะปกติสิ่งนี้ถูกขัดขวางโดยการทำงานของปั๊ม Na +, K +
ภายใต้สภาวะธรรมชาติในเซลล์ประสาท CNS ศักยภาพในการกระทำส่วนใหญ่เกิดขึ้นในพื้นที่ของเนินเขาแอกโซนัลในเซลล์ประสาทที่ได้รับผลกระทบ - ในการสกัดกั้นของ Ranvier ของเส้นประสาทที่สิ้นสุดใกล้กับตัวรับความรู้สึกเช่น ในพื้นที่เหล่านั้นของเมมเบรนที่มีช่องโซเดียมขึ้นอยู่กับแรงดันไฟฟ้าที่เลือกอย่างรวดเร็วและช่องโพแทสเซียมช้า ในเซลล์ประเภทอื่น ๆ (เช่นเครื่องกระตุ้นหัวใจ, myocytes แบบเรียบ) ไม่เพียง แต่โซเดียมและโพแทสเซียมเท่านั้น แต่ยังมีช่องแคลเซียมที่มีบทบาทในการพัฒนา AP ด้วย
กลไกการรับรู้และการแปลงสัญญาณเป็น AP ในตัวรับประสาทสัมผัสทุติยภูมิแตกต่างจากกลไกที่วิเคราะห์สำหรับตัวรับประสาทสัมผัสหลัก ในตัวรับเหล่านี้การรับรู้สัญญาณจะดำเนินการโดยเซลล์ประสาทสัมผัสพิเศษ (เซลล์รับแสง, การดมกลิ่น) หรือเซลล์ประสาทสัมผัส (รับรส, หู, ขนถ่าย) เซลล์ที่บอบบางเหล่านี้แต่ละเซลล์มีกลไกพิเศษในการรับรู้สัญญาณของตัวเอง อย่างไรก็ตามในเซลล์ทั้งหมดพลังงานของสัญญาณการรับรู้ (สิ่งกระตุ้น) จะถูกแปลงเป็นความผันผวนของความต่างศักย์ของพลาสมาเมมเบรนนั่นคือ เข้าไปในศักยภาพของตัวรับ
ดังนั้นช่วงเวลาสำคัญในกลไกของการเปลี่ยนแปลงของสัญญาณที่รับรู้เป็นศักยภาพของตัวรับโดยเซลล์ประสาทสัมผัสคือการเปลี่ยนแปลงความสามารถในการซึมผ่านของช่องไอออนเพื่อตอบสนองต่อการสัมผัส การเปิดช่อง Na +, Ca 2+, K + -ion ในระหว่างการรับรู้สัญญาณและการเปลี่ยนแปลงเกิดขึ้นได้ในเซลล์เหล่านี้ด้วยการมีส่วนร่วมของโปรตีน G, ตัวไกล่เกลี่ยภายในเซลล์ที่สอง, การจับกับลิแกนด์และฟอสโฟรีเลชันของช่องไอออน ตามกฎแล้วศักยภาพของตัวรับที่เกิดขึ้นในเซลล์ประสาทสัมผัสทำให้เกิดการปล่อยสารสื่อประสาทจากพวกมันไปยังช่องว่างระหว่างซินแนปติกซึ่งช่วยให้มั่นใจได้ว่าการส่งสัญญาณไปยังเมมเบรนโพสซิแนปติกของเส้นประสาทที่สิ้นสุดลงและการสร้างกระแสประสาทบนเยื่อหุ้มเซลล์ กระบวนการเหล่านี้มีรายละเอียดอยู่ในบทที่เกี่ยวกับระบบประสาทสัมผัส
ศักยภาพในการออกฤทธิ์สามารถจำแนกได้ตามแอมพลิจูดและระยะเวลาซึ่งสำหรับใยประสาทเดียวกันจะยังคงเหมือนเดิมเมื่อ AP แพร่กระจายไปตามเส้นใย ดังนั้นศักยภาพในการกระทำจึงเรียกว่าศักยภาพที่ไม่ต่อเนื่อง
มีการเชื่อมต่อที่ชัดเจนระหว่างลักษณะของผลกระทบต่อตัวรับประสาทสัมผัสและจำนวน AP ที่เกิดขึ้นในเส้นใยประสาทสัมผัสเพื่อตอบสนองต่อผลกระทบ มันอยู่ในความจริงที่ว่ามีขนาดใหญ่ แต่มีความแข็งแรงหรือระยะเวลาในการสัมผัสกับเส้นใยประสาท มากกว่า กระแสประสาทเช่น ด้วยการเพิ่มขึ้นของผลกระทบแรงกระตุ้นของความถี่ที่สูงขึ้นจะถูกส่งไปยังระบบประสาทจากตัวรับ กระบวนการแปลงข้อมูลเกี่ยวกับลักษณะของผลกระทบเป็นความถี่และพารามิเตอร์อื่น ๆ ของกระแสประสาทที่ส่งไปยังระบบประสาทส่วนกลางเรียกว่าการเข้ารหัสข้อมูลแบบไม่ต่อเนื่อง
สำหรับคำอธิบาย ต้นกำเนิดของศักยภาพในการพักผ่อน ได้รับการแนะนำ ทฤษฎีต่างๆ... ต้นกำเนิดของความเข้าใจสมัยใหม่เกี่ยวกับปัญหานี้คือผลงานของ V. ต่อมาในปี 1902 ยูริเบิร์นสไตน์ได้พัฒนาทฤษฎีเมมเบรน - ไอออนซึ่งได้รับการดัดแปลงและพิสูจน์โดยการทดลองโดย A. Hodgkin และ A. Huxley (1952) และปัจจุบันได้รับการยอมรับอย่างกว้างขวาง ตามทฤษฎีนี้ศักยภาพทางชีวภาพเกิดจากความเข้มข้นของไอออน K ионов, Na ֹ, Cl "ที่ไม่เท่ากันทั้งภายในและภายนอกเซลล์และความสามารถในการซึมผ่านของเยื่อผิวที่แตกต่างกัน
โพรโทพลาสซึมของเซลล์ประสาทและกล้ามเนื้อมีโพแทสเซียมไอออนมากกว่า 30-50 เท่าโซเดียมไอออน 8-10 เท่าและคลอรีนไอออนน้อยกว่าของเหลวนอกเซลล์ 50 เท่า
อุปสรรคในการทำให้เท่ากันอย่างรวดเร็วของความแตกต่างของความเข้มข้นนี้คือเยื่อหุ้มพลาสมาที่บางที่สุด (ประมาณ 100 Å) ที่ปกคลุมเซลล์ของสิ่งมีชีวิต
|
โครงสร้างของเมมเบรนนี้ขึ้นอยู่กับข้อมูลที่ได้รับจากกล้องจุลทรรศน์อิเล็กตรอนกล้องจุลทรรศน์แบบออปติคัลการเลี้ยวเบนของรังสีเอกซ์และการวิเคราะห์ทางเคมี สันนิษฐานว่าเมมเบรนประกอบด้วยโมเลกุลฟอสโฟลิปิดสองชั้นปกคลุมจากด้านในด้วยชั้นของโมเลกุลโปรตีนและจากภายนอกด้วยชั้นของโมเลกุลคาร์โบไฮเดรตเชิงซ้อน - มิวโคโพลีแซ็กคาไรด์ โครงสร้างเมมเบรนสามชั้นแสดงในแผนผัง รูปที่. 116. รูป: 116. แผนภาพโครงสร้างโมเลกุลของเมมเบรน. แสดงให้เห็นว่าเป็นชั้นไขมัน bimolecular Z (วงกลมบ่งบอกถึงกลุ่มขั้วของฟอสโฟลิปิด) และโมโนเลย์ที่ไม่ใช่ไขมันสองตัว: นอก - มิวโคโพลีแซคคาไรด์ - X, โปรตีนชั้นใน - Y (โดย Robertson) |
เยื่อหุ้มเซลล์ประกอบด้วยท่อที่ดีที่สุด - "รูพรุน" ที่มีเส้นผ่านศูนย์กลางหลายอังสตรอม ผ่านท่อโมเลกุลของน้ำและสารอื่น ๆ รวมทั้งไอออนที่มีเส้นผ่านศูนย์กลางตรงกับขนาดของรูพรุนเข้าไปในเซลล์และปล่อยทิ้งไว้
ไอออนส่วนบุคคลได้รับการแก้ไขบนองค์ประกอบโครงสร้างของเมมเบรนซึ่งทำให้ผนังของรูขุมขนมีประจุหนึ่งหรือประจุอื่นและทำให้เกิดความซับซ้อนหรืออำนวยความสะดวกในการเคลื่อนผ่านของไอออนผ่านพวกมัน ดังนั้นจึงสันนิษฐานได้ว่าการปรากฏตัวของหมู่ฟอสเฟตและคาร์บอกซิลที่แยกออกจากกันในเมมเบรนเป็นสาเหตุที่เมมเบรนของเส้นใยประสาทดูดซึมแอนไอออนได้น้อยกว่าไอออนบวก
ความสามารถในการซึมผ่านของเมมเบรนสำหรับไอออนบวกที่แตกต่างกันนั้นไม่เหมือนกันและจะเปลี่ยนแปลงตามธรรมชาติตามสถานะการทำงานที่แตกต่างกันของเนื้อเยื่อ ในส่วนที่เหลือเมมเบรนของเส้นใยประสาทจะซึมผ่านไปยังไอออนของ K ֹได้มากกว่าไอออน Na ประมาณ 20-100 เท่าและเมื่อมีการกระตุ้นความสามารถในการซึมผ่านของโซเดียมจะสูงเกินความสามารถในการซึมผ่านของโพแทสเซียมอย่างมีนัยสำคัญ
เพื่อให้เข้าใจกลไกของการปรากฏตัวของศักยภาพของเยื่อหุ้มจากมุมมองของทฤษฎีเบิร์นสไตน์ - ฮอดจ์กินให้เราพิจารณาการทดลองแบบจำลอง ครึ่งแรกของเรือ ( รูปที่. 117) คั่นด้วยเมมเบรนกึ่งซึมผ่านเทียมรูขุมขนซึ่งผ่าน K ֹไอออนที่มีประจุบวกได้อย่างอิสระและไม่ผ่าน SO "4 ไอออนที่มีประจุลบจะเต็มไปด้วยสารละลาย K2SO4 เข้มข้นและครึ่งซ้ายเต็มไปด้วยสารละลาย K2SO4 แต่มีความเข้มข้นต่ำกว่า
 |
เนื่องจากการมีอยู่ของการไล่ระดับความเข้มข้น K ֹไอออนจะเริ่มกระจายผ่านเมมเบรนส่วนใหญ่จากครึ่งขวาของเรือ (โดยที่ความเข้มข้นคือ C1) ไปทางซ้าย (โดยมีความเข้มข้น C2) ในทำนองเดียวกันแอนไอออน SO "4 ที่มีประจุลบซึ่งเมมเบรนไม่สามารถซึมผ่านได้จะรวมตัวอยู่ที่ครึ่งขวาของเรือที่ผิวเมมเบรน ด้วยประจุลบพวกมันจะจับไอออน K ที่ด้านซ้ายของผิวเมมเบรนด้วยไฟฟ้าสถิต เป็นผลให้เมมเบรนกลายเป็นโพลาไรซ์: ความต่างศักย์เกิดขึ้นระหว่างพื้นผิวทั้งสอง รูป: 117. การเกิดขึ้นของความต่างศักย์บนเยื่อเทียมที่แยกสารละลายของ K2SO4 ที่มีความเข้มข้นต่างกัน (C1 และ C2) เมมเบรนสามารถซึมผ่านได้ถึง K ֹไอออนบวก (วงกลมเล็ก ๆ ) และไม่อนุญาตให้แอนไอออน SO "4 (วงกลมใหญ่) ผ่าน 1 และ 2 - อิเล็กโทรดที่แช่อยู่ในสารละลาย 3 - อุปกรณ์วัดไฟฟ้า |
หากตอนนี้อิเล็กโทรดลดระดับลงในซีกขวาและซีกซ้ายของภาชนะแล้วอุปกรณ์วัดไฟฟ้าจะตรวจจับความต่างศักย์ในขณะที่สารละลายที่มีความเข้มข้นต่ำกว่าของไอออน K2SO4 ซึ่งการแพร่กระจายของไอออน K ֹที่มีประจุบวกเกิดขึ้นเป็นส่วนใหญ่จะได้รับประจุบวกเมื่อเทียบกับสารละลายที่มีความเข้มข้นของ K2SO4 สูงกว่า
ความต่างศักย์ (E) ในกรณีที่พิจารณาสามารถคำนวณได้โดยใช้สูตร Nernst:

มีหลายเหตุผลที่เชื่อว่าความสัมพันธ์ที่คล้ายคลึงกันเกิดขึ้นในเส้นใยประสาทของสิ่งมีชีวิตเนื่องจากความเข้มข้นของ K ֹไอออนในโปรโตพลาสซึมนั้นสูงกว่าความเข้มข้นของไอออนเหล่านี้ในสารละลายภายนอกมากกว่า 30 ดอกและแอนไอออนอินทรีย์ (โปรตีน ฯลฯ ) ของโปรโตพลาสซึมผ่านเมมเบรนในทางปฏิบัติไม่ได้ เจาะ.
ในสภาพที่เหลือทางสรีรวิทยาการแพร่กระจายของ K ไอออนที่มีประจุบวกจากโปรโตพลาสซึมไปสู่ของเหลวภายนอกทำให้ผิวด้านนอกของเมมเบรนมีประจุบวกและด้านในเป็นประจุลบ
ข้อโต้แย้งที่สำคัญที่สนับสนุนความถูกต้องของแนวคิดนี้คือความจริงที่ว่าความต่างศักย์ที่คำนวณโดยสูตร Nernst ระหว่างด้านนอกและด้านในของเยื่อหุ้มเส้นใยกล้ามเนื้อ (ประมาณ 90 mV) นั้นใกล้เคียงกับที่วัดได้ในการทดลองโดยใช้ไมโครอิเล็กโทรดภายในเซลล์
นอกจากนี้ยังพบว่าการเพิ่มขึ้นของความเข้มข้นของ K ֹไอออนในสภาพแวดล้อมภายนอกของเซลล์และด้วยเหตุนี้การลดลงของความแตกต่างของความเข้มข้นของไอออนเหล่านี้ทั้งสองด้านของเมมเบรนทำให้ศักยภาพในการพักลดลงและในช่วงความเข้มข้นหนึ่งการเปลี่ยนแปลงเหล่านี้ในเชิงปริมาณจะสอดคล้องกับที่คำนวณโดยสูตร Nernst
อย่างไรก็ตามผลการพิสูจน์ที่สำคัญที่สุดโดยตรงและตรงถึงความถูกต้องของแนวคิดเหล่านี้ได้มาจาก A. Hodzhkin และเพื่อนร่วมงาน (1962) ในการทดลองด้วยการเปลี่ยนโปรโตพลาสซึมในเส้นใยประสาทขนาดยักษ์ของหอยปลาหมึกด้วยน้ำเกลือ โปรโตพลาสซึมถูกบีบออกอย่างระมัดระวังจากเส้นใยที่มีเส้นผ่านศูนย์กลางประมาณ 1 มล. และเปลือกที่ยุบจะเต็มไปด้วยน้ำเกลือเทียม
ในกรณีที่ความเข้มข้นของโพแทสเซียมไอออนในสารละลายนี้ใกล้เคียงกับภายในเซลล์จะมีการสร้างความต่างศักย์ระหว่างด้านในและด้านนอกของเมมเบรนโดยประมาณเท่ากับศักยภาพในการพักของเส้นใยปกติ (50-80 mV) การลดลงของความเข้มข้นของ K ֹไอออนในสารละลายภายในทำให้ศักยภาพในการพักลดลงตามธรรมชาติหรือแม้กระทั่งการบิดเบือน
การทดลองดังกล่าวแสดงให้เห็นว่าการไล่ระดับความเข้มข้นของ K ไอออนเป็นปัจจัยหลักที่กำหนดมูลค่าของศักยภาพในการพักผ่อนของเส้นใยประสาท
นอกจาก K ไอออนแล้ว Na ไอออนซึ่งแพร่เข้าสู่โปรโตพลาสซึมจากของเหลวนอกเซลล์ซึ่งมีความเข้มข้นสูงก็มีส่วนร่วมในการปรากฏตัวของศักยภาพที่เหลือ การแพร่กระจายนี้ถูกขัดขวางอย่างมากเนื่องจากการซึมผ่านของโซเดียมต่ำของเมมเบรนในขณะพัก อย่างไรก็ตามการแพร่กระจายผ่านเมมเบรนเข้าสู่โปรโตพลาสซึม Na ไอออนจะถ่ายเทประจุบวกของพวกมันที่นี่ซึ่งจะช่วยลดค่าของศักยภาพในการพักผ่อนที่เกิดจากการแพร่ของ K ไอออนจากเซลล์ สิ่งนี้อธิบายถึงความจริงที่ว่าศักยภาพในการพักผ่อนของเซลล์ประสาทและเส้นใยส่วนใหญ่ไม่ใช่ 90 mV อย่างที่คาดไว้หากศักยภาพนี้สร้างขึ้นโดย K ไอออนเท่านั้น แต่เป็น 60-70 mV
ดังนั้นค่าของศักยภาพในการพักตัวของเส้นใยประสาทและเซลล์จะถูกกำหนดโดยอัตราส่วนของจำนวนไอออน K ֺที่มีประจุบวกการกระจายต่อหน่วยเวลาจากเซลล์ออกไปด้านนอกและ Na ไอออนที่มีประจุบวกซึ่งกระจายผ่านเมมเบรนในทิศทางตรงกันข้าม ยิ่งอัตราส่วนนี้สูงเท่าไหร่มูลค่าของศักยภาพในการพักก็จะยิ่งมากขึ้นและในทางกลับกัน
text_fields
text_fields
arrow_upward
ศักยภาพในการพักผ่อนของเมมเบรน (MPP) หรือ ศักยภาพในการพักผ่อน (PP) คือความต่างศักย์ของเซลล์พักผ่อนระหว่างด้านในและด้านนอกของเมมเบรน ด้านในของเยื่อหุ้มเซลล์มีประจุลบเมื่อเทียบกับด้านนอก การใช้ศักยภาพของโซลูชันภายนอกเป็นศูนย์ MPP จะเขียนด้วยเครื่องหมายลบ ปริมาณ WFPขึ้นอยู่กับชนิดของผ้าและแตกต่างกันไปตั้งแต่ -9 ถึง -100 mV ดังนั้นในส่วนที่เหลือเยื่อหุ้มเซลล์ โพลาไรซ์เรียกว่าค่า MPP ลดลง depolarizationเพิ่มขึ้น - hyperpolarizationคืนค่าเดิม WFP- อีกครั้งโพลาไรซ์เยื่อ
บทบัญญัติพื้นฐานของทฤษฎีการกำเนิดเมมเบรน WFPต้มต่อไปนี้ ในส่วนที่เหลือเยื่อหุ้มเซลล์สามารถซึมผ่าน K + ไอออนได้ดี (ในบางเซลล์และถึง SG) ซึมผ่าน Na + ได้น้อยลงและไม่สามารถซึมผ่านไปยังโปรตีนภายในเซลล์และไอออนอินทรีย์อื่น ๆ ได้ K + ไอออนจะแพร่กระจายออกจากเซลล์ตามการไล่ระดับความเข้มข้นในขณะที่แอนไอออนที่ไม่ทะลุทะลวงยังคงอยู่ในไซโทพลาสซึมทำให้เกิดความแตกต่างที่อาจเกิดขึ้นบนเยื่อ
ความต่างศักย์ที่เกิดขึ้นจะป้องกันไม่ให้ K + ออกจากเซลล์และที่ค่าหนึ่งของมันความสมดุลจะเกิดขึ้นระหว่างทางออกของ K + ตามการไล่ระดับความเข้มข้นและการเข้าของไอออนบวกเหล่านี้ตามการไล่ระดับทางไฟฟ้าที่เกิดขึ้น ศักยภาพของเมมเบรนที่เกิดความสมดุลนี้เรียกว่า ศักยภาพสมดุลสีแดงค่าของมันสามารถคำนวณได้จากสมการ Nernst:
ที่ไหน E ถึง- ศักยภาพสมดุลสำหรับ ถึง + ; ร - ค่าคงที่ของก๊าซ ต- อุณหภูมิสัมบูรณ์ ฉ - หมายเลขฟาราเดย์; ป- วาเลนซ์ K + (+1), [K n +] - [K + ต่อ] -ความเข้มข้นภายนอกและภายใน K + -
ถ้าเราเปลี่ยนจากลอการิทึมธรรมชาติไปเป็นทศนิยมและแทนที่ค่าตัวเลขของค่าคงที่ลงในสมการสมการจะอยู่ในรูปแบบ: 
ในเซลล์ประสาทไขสันหลัง (ตารางที่ 1.1) E k \u003d -90 mV ค่า MPP ที่วัดด้วยไมโครอิเล็กโทรดต่ำกว่าอย่างเห็นได้ชัด - 70 mV
ตารางที่ 1.1... ความเข้มข้นของไอออนบางส่วนภายในและภายนอกของ motoneurons กระดูกสันหลังของสัตว์เลี้ยงลูกด้วยนม
| และเขา |
ความเข้มข้น |
(mmol / ลิตร H 2 O) |
ศักยภาพสมดุล (mv) |
|
ภายในเซลล์ |
นอกกรง |
||
| นา + | 15,0 | 150,0 | |
| K + | 150,0 | 5,5 | |
| Сl - | 125,0 | ||
|
ศักยภาพของเมมเบรนพัก \u003d -70 mV |
|||
หากศักยภาพของเมมเบรนของเซลล์มีลักษณะโพแทสเซียมตามสมการ Nernst ค่าของมันควรลดลงเป็นเชิงเส้นเมื่อการไล่ระดับความเข้มข้นของไอออนเหล่านี้ลดลงตัวอย่างเช่นเมื่อความเข้มข้นของ K + เพิ่มขึ้นในของเหลวนอกเซลล์ อย่างไรก็ตามการพึ่งพาเชิงเส้นของ BMP (Resting Membrane Potential) บนการไล่ระดับความเข้มข้นของ K + จะมีอยู่ก็ต่อเมื่อความเข้มข้นของ K + ในของเหลวนอกเซลล์สูงกว่า 20 mM ที่ความเข้มข้นต่ำกว่าของ K + ภายนอกเซลล์เส้นโค้งของการพึ่งพา E m บนลอการิทึมของอัตราส่วนของความเข้มข้นของโพแทสเซียมภายนอกและภายในเซลล์จะแตกต่างจากทางทฤษฎี เป็นไปได้ที่จะอธิบายความเบี่ยงเบนที่กำหนดขึ้นของการพึ่งพาการทดลองของขนาดของ MPP และการไล่ระดับความเข้มข้น K + ที่คำนวณตามทฤษฎีโดยใช้สมการ Nernst โดยสมมติว่า MPP ของเซลล์ที่กระตุ้นได้นั้นไม่เพียง แต่กำหนดโดยโพแทสเซียมเท่านั้น แต่ยังรวมถึงศักยภาพสมดุลของโซเดียมและคลอรีนด้วย การใช้เหตุผลคล้ายกับข้อก่อนหน้าคุณสามารถเขียน:
ค่าของศักยภาพสมดุลโซเดียมและคลอไรด์สำหรับเซลล์ประสาทไขสันหลัง (ตารางที่ 1.1) คือ +60 และ -70 mV ตามลำดับ ค่า E Cl เท่ากับค่า MPP สิ่งนี้บ่งบอกถึงการกระจายตัวของคลอรีนไอออนแบบพาสซีฟทั่วเมมเบรนตามการไล่ระดับทางเคมีและทางไฟฟ้า สำหรับโซเดียมไอออนการไล่ระดับทางเคมีและไฟฟ้าจะถูกส่งเข้าไปในเซลล์
การมีส่วนร่วมของศักยภาพสมดุลแต่ละค่าต่อค่า MPP จะถูกกำหนดโดยอัตราส่วนระหว่างความสามารถในการซึมผ่านของเยื่อหุ้มเซลล์สำหรับแต่ละไอออนเหล่านี้ การคำนวณค่าของศักยภาพของเมมเบรนทำได้โดยใช้สมการ Goldmann:

อีม- ศักยภาพของเมมเบรน ร- ค่าคงที่ของก๊าซ ต- อุณหภูมิสัมบูรณ์ ฉ- หมายเลขฟาราเดย์; ร K, P Naและ ร Cl -ค่าคงที่การซึมผ่านของเมมเบรนสำหรับ K + Na + และ Cl ตามลำดับ [ถึง + n ], [ เค + ต่อ, [ นา + n [ นา + ต่อ], [Сl - n] และ [Сl - int] - ความเข้มข้นของ K +, Na + และ Cl นอก (n) และภายใน (ภายนอก) เซลล์
การแทนที่ความเข้มข้นของไอออนและค่า MPP ที่ได้จากการศึกษาทดลองในสมการนี้จะแสดงให้เห็นว่าสำหรับแอกซอนยักษ์ของปลาหมึกควรมีอัตราส่วนของค่าคงที่การซึมผ่านต่อไปนี้ P ถึง: P Na: P C1 \u003d I: 0.04: 0.45 เห็นได้ชัดว่าเนื่องจากเมมเบรนสามารถซึมผ่านโซเดียมไอออนได้ (P N a =/ 0) และศักย์สมดุลของไอออนเหล่านี้มีเครื่องหมาย "บวก" จากนั้นการเข้าสู่เซลล์ตามการไล่ระดับทางเคมีและทางไฟฟ้าจะลดอิเล็กโทรเนกาติวิตีของไซโตพลาสซึมเช่น เพิ่ม MPP (Resting Membrane Potential)
ด้วยการเพิ่มความเข้มข้นของโพแทสเซียมไอออนในสารละลายภายนอกที่สูงกว่า 15 mM MPP จะเพิ่มขึ้นและอัตราส่วนของค่าคงที่การซึมผ่านจะเปลี่ยนไปเป็นส่วนเกินที่มีนัยสำคัญมากขึ้นจาก» P ถึงมากกว่า P Na และ P C1 P ถึง: P Na: P C1 \u003d 1: 0.025: 0.4 ภายใต้เงื่อนไขดังกล่าว MPP จะถูกกำหนดโดยการไล่ระดับของโพแทสเซียมไอออนเกือบทั้งหมดดังนั้นการทดลองและการพึ่งพาทางทฤษฎีของค่า MPP บนลอการิทึมของอัตราส่วนของความเข้มข้นโพแทสเซียมภายนอกและภายในเซลล์จึงเริ่มตรงกัน
ดังนั้นการปรากฏตัวของความต่างศักย์ที่อยู่นิ่งระหว่างไซโทพลาสซึมและสภาพแวดล้อมภายนอกในเซลล์พักผ่อนเกิดจากการไล่ระดับความเข้มข้นที่มีอยู่สำหรับ K +, Na + และ Cl และความสามารถในการซึมผ่านของเยื่อหุ้มที่แตกต่างกันสำหรับไอออนเหล่านี้ บทบาทหลักในการสร้าง MPP คือการแพร่กระจายของโพแทสเซียมไอออนจากเซลล์ไปสู่สารละลายภายนอก นอกจากนี้ MPP ยังถูกกำหนดโดยศักยภาพสมดุลของโซเดียมและคลอรีนและการมีส่วนร่วมของแต่ละตัวจะถูกกำหนดโดยความสัมพันธ์ระหว่างการซึมผ่านของเยื่อหุ้มเซลล์พลาสมาสำหรับไอออนเหล่านี้
ปัจจัยทั้งหมดที่ระบุไว้ข้างต้นถือเป็นสิ่งที่เรียกว่า ส่วนประกอบไอออนิกMPP (ศักยภาพในการพักผ่อนของเมมเบรน) เนื่องจากศักยภาพสมดุลของโพแทสเซียมและโซเดียมไม่เท่ากับ MPP เซลล์ต้องดูดซับ Na + และสูญเสีย K + ความคงที่ของความเข้มข้นของไอออนเหล่านี้ในเซลล์จะยังคงอยู่เนื่องจากการทำงานของ Na + K + -ATPase
อย่างไรก็ตามบทบาทของปั๊มไอออนนี้ไม่ได้ จำกัด อยู่ที่การรักษาการไล่ระดับโซเดียมและโพแทสเซียม เป็นที่ทราบกันดีว่าปั๊มโซเดียมเป็นอิเล็กโทรจีนิกและในระหว่างการทำงานจะมีการไหลของประจุบวกบริสุทธิ์จากเซลล์ไปยังของเหลวนอกเซลล์ซึ่งทำให้อิเล็กโทรเนกาติวิตีของไซโตพลาสซึมเพิ่มขึ้นเมื่อเทียบกับตัวกลาง ความเป็นไฟฟ้าของโซเดียมปั๊มถูกเปิดเผยในการทดลองกับเซลล์ประสาทขนาดยักษ์ของมอลลัสซี การฉีดอิเล็กโตรโฟเรติกของไอออน Na + เข้าไปในร่างกายของเซลล์ประสาทเดี่ยวทำให้เกิดเยื่อหุ้มโพลาไรเซชันมากเกินไปในระหว่างที่ IPP ต่ำกว่าค่าของศักยภาพสมดุลโพแทสเซียมอย่างมีนัยสำคัญ hyperpolarization นี้อ่อนลงเมื่ออุณหภูมิของสารละลายที่เซลล์อยู่ลดลงและถูกยับยั้งโดยตัวยับยั้งเฉพาะของ Na +, K + -ATPase ouabain
จากข้างต้น MPP สามารถแบ่งออกเป็นสองส่วน - "ไอออนิก"และ “ เมตาบอลิก”.ส่วนประกอบแรกขึ้นอยู่กับการไล่ระดับความเข้มข้นของไอออนและความสามารถในการซึมผ่านของเมมเบรน ประการที่สอง "การเผาผลาญ" เกิดจากการขนส่งโซเดียมและโพแทสเซียมที่ใช้งานอยู่และมีผลสองเท่า MPP.ในแง่หนึ่งปั๊มโซเดียมจะรักษาระดับความเข้มข้นระหว่างไซโทพลาสซึมและสิ่งแวดล้อม ในทางกลับกันการเป็น Electrogenic ปั๊มโซเดียมมีผลโดยตรงต่อ MPP การมีส่วนร่วมของค่า MPP ขึ้นอยู่กับความหนาแน่นของกระแส "สูบน้ำ" (กระแสต่อหน่วยพื้นที่ของผิวเยื่อหุ้มเซลล์) และความต้านทานของเมมเบรน
ศักยภาพในการทำงานของเมมเบรน
text_fields
text_fields
arrow_upward
หากมีการระคายเคืองต่อเส้นประสาทหรือกล้ามเนื้อสูงกว่าเกณฑ์กระตุ้น MPP ของเส้นประสาทหรือกล้ามเนื้อจะลดลงอย่างรวดเร็วและในช่วงเวลาสั้น ๆ (มิลลิวินาที) เมมเบรนจะถูกชาร์จใหม่: ด้านในของมันจะกลายเป็นประจุบวกเมื่อเทียบกับด้านนอก มัน การเปลี่ยนแปลงระยะสั้นใน MPP ที่เกิดขึ้นเมื่อเซลล์รู้สึกตื่นเต้นซึ่งบนหน้าจอออสซิลโลสโคปมีรูปแบบของจุดสูงสุดเดียวเรียกว่า ศักยภาพในการทำงานของเมมเบรน (MTD).
IVD ในเนื้อเยื่อของเส้นประสาทและกล้ามเนื้อเกิดขึ้นเมื่อค่าสัมบูรณ์ของ MPP (เมมเบรนดีโพลาไรเซชัน) ลดลงเป็นค่าวิกฤตที่เรียกว่า เกณฑ์การสร้างMTD. ในเส้นใยประสาทขนาดยักษ์ของปลาหมึก IVD คือ - 60 mV เมื่อเมมเบรนถูกเปลี่ยนขั้วเป็น -45 mV (เกณฑ์ของการสร้าง IVD) IVD จะปรากฏขึ้น (รูปที่ 1.15)
รูป: 1.15 ศักยภาพในการดำเนินการของใยประสาท (A) และการเปลี่ยนแปลงการนำเมมเบรนของโซเดียมและโพแทสเซียมไอออน (B)ในช่วงเริ่มมีอาการของ IVD ในซอนปลาหมึกความต้านทานของเมมเบรนจะลดลง 25 เท่าจาก 1,000 ถึง 40 โอห์มซม. 2 ในขณะที่ความจุไฟฟ้าไม่เปลี่ยนแปลง การลดลงของความต้านทานของเมมเบรนที่ระบุนั้นเกิดจากการเพิ่มขึ้นของความสามารถในการซึมผ่านไอออนิกของเมมเบรนเมื่อมีการกระตุ้น
ในแง่ของแอมพลิจูด (100-120 mV) MTD (Membrane Action Potential) จะสูงกว่า MPP (ศักยภาพในการพักตัว) 20-50 mV กล่าวอีกนัยหนึ่งด้านในของเมมเบรนในช่วงเวลาสั้น ๆ จะกลายเป็นประจุบวกเมื่อเทียบกับด้านนอก - "แหก" หรือ การกลับรายการ
จากสมการของโกลด์มันน์พบว่าการเพิ่มขึ้นของความสามารถในการซึมผ่านของเมมเบรนของโซเดียมไอออนเท่านั้นที่สามารถนำไปสู่การเปลี่ยนแปลงศักยภาพของเมมเบรนได้ ค่า E k จะน้อยกว่าค่า MPP เสมอดังนั้นการเพิ่มความสามารถในการซึมผ่านของเมมเบรนสำหรับ K + จะเพิ่มค่าสัมบูรณ์ของ MPP ศักย์สมดุลโซเดียมมีเครื่องหมายบวกดังนั้นการเพิ่มขึ้นอย่างรวดเร็วของความสามารถในการซึมผ่านของเมมเบรนสำหรับไอออนบวกเหล่านี้จะนำไปสู่การชาร์จใหม่ของเมมเบรน
ในระหว่างการทำ IVD ความสามารถในการซึมผ่านของเมมเบรนของโซเดียมไอออนจะเพิ่มขึ้น จากการคำนวณแสดงให้เห็นว่าหากพักอัตราส่วนของค่าคงที่การซึมผ่านของเมมเบรนสำหรับ K +, Na + และ SG คือ 1: 0.04: 0.45 แล้วที่ MPD - P k: P Na: P \u003d 1: 20: 0.45 ... ดังนั้นในสภาวะที่ถูกกระตุ้นเยื่อหุ้มของเส้นใยประสาทจึงไม่เพียง แต่สูญเสียความสามารถในการซึมผ่านของไอออนิกที่เลือกได้เท่านั้น แต่ในทางกลับกันการซึมผ่านของโพแทสเซียมอิออนจะถูกเลือกให้ซึมผ่านได้ การเพิ่มขึ้นของความสามารถในการซึมผ่านของโซเดียมของเมมเบรนมีความสัมพันธ์กับการเปิดช่องโซเดียมขึ้นอยู่กับแรงดันไฟฟ้า
กลไกที่ให้การเปิดและปิดช่องไอออนเรียกว่า ประตูช่องเป็นเรื่องปกติที่จะแยกแยะ การเปิดใช้งาน(m) และ กำลังปิดใช้งาน(h) ประตู ช่องไอออนสามารถอยู่ในสถานะพื้นฐานสามสถานะ: ปิด (m-gate ถูกปิด h-open) เปิด (m- และ h-gate เปิดอยู่) และปิดใช้งาน (m-gate เปิดอยู่ h-gate ปิด) (รูปที่ 1.16)
รูป: 1.16 รูปแบบของตำแหน่งของการเปิดใช้งาน (m) และการปิดใช้งาน (h) ประตูของช่องโซเดียมที่สอดคล้องกับสถานะปิด (ส่วนที่เหลือ A) เปิด (การเปิดใช้งาน B) และสถานะปิดใช้งาน (C)
Depolarization ของเมมเบรนซึ่งเกิดจากสิ่งกระตุ้นที่ทำให้ระคายเคืองตัวอย่างเช่นกระแสไฟฟ้าเปิดประตู m ของช่องโซเดียม (เปลี่ยนจากสถานะ A เป็น B) และให้ประจุบวกไหลเข้าด้านใน - โซเดียมไอออน สิ่งนี้นำไปสู่การลดขั้วของเมมเบรนต่อไปซึ่งในทางกลับกันจะเพิ่มจำนวนช่องโซเดียมที่เปิดอยู่และทำให้การซึมผ่านของโซเดียมเพิ่มขึ้น มีการเปลี่ยนขั้วของเมมเบรน "regenerative" ซึ่งเป็นผลมาจากการที่ศักยภาพของด้านในของเมมเบรนมีแนวโน้มที่จะถึงค่าของศักย์สมดุลโซเดียม
สาเหตุของการยุติการเติบโตของ IVD (Membrane Action Potential) และการเปลี่ยนขั้วของเยื่อหุ้มเซลล์คือ:
ก) Depolarization เมมเบรนที่เพิ่มขึ้นเช่น เมื่อ E m - » E Na ซึ่งเป็นผลมาจากการที่การไล่ระดับสีทางเคมีไฟฟ้าสำหรับไอออนโซเดียมลดลงเท่ากับ E m -\u003e E Na กล่าวอีกนัยหนึ่งแรงที่ "ผลัก" โซเดียมเข้าสู่เซลล์จะลดลง
ข) Depolarization ของเมมเบรนก่อให้เกิดกระบวนการปิดใช้งานช่องโซเดียม (ปิดประตู h สถานะของช่อง B) ซึ่งยับยั้งการเติบโตของการซึมผ่านของโซเดียมของเมมเบรนและนำไปสู่การลดลง
ใน) Depolarization ของเมมเบรนเพิ่มความสามารถในการซึมผ่านของโพแทสเซียมไอออน กระแสโพแทสเซียมที่ส่งออกมีแนวโน้มที่จะเปลี่ยนศักยภาพของเมมเบรนไปสู่ศักยภาพสมดุลโพแทสเซียม
การลดลงของศักย์ไฟฟ้าเคมีสำหรับโซเดียมไอออนและการปิดใช้งานช่องโซเดียมจะลดปริมาณโซเดียมที่เข้ามาในปัจจุบัน ในช่วงเวลาหนึ่งค่าของกระแสโซเดียมอินพุตจะถูกเปรียบเทียบกับกระแสเอาต์พุตที่เพิ่มขึ้น - การเติบโตของ MPD จะหยุดลง เมื่อกระแสไฟฟ้าขาออกทั้งหมดเกินกว่าขาเข้าการเปลี่ยนขั้วของเมมเบรนจะเริ่มขึ้นซึ่งมีลักษณะการสร้างใหม่ด้วย การเปลี่ยนขั้วของ incipient นำไปสู่การปิดประตูกระตุ้น (m) ซึ่งจะลดการซึมผ่านของโซเดียมของเมมเบรนเร่งการเปลี่ยนขั้วและการเพิ่มจำนวนช่องปิดเป็นต้น
ขั้นตอนการเปลี่ยนโพลาไรเซชันของ IVD ในบางเซลล์ (เช่นในคาร์ดิโอไมโอไซต์และเซลล์กล้ามเนื้อเรียบจำนวนหนึ่ง) สามารถชะลอตัว ที่ราบสูงAP เนื่องจากการเปลี่ยนแปลงที่ซับซ้อนในช่วงเวลาของกระแสขาเข้าและขาออกผ่านเมมเบรน Hyperpolarization และ / หรือ Depolarization ของเมมเบรนอาจเกิดขึ้นจากผลของ IVD สิ่งเหล่านี้เรียกว่า ศักยภาพในการติดตามTrace hyperpolarization มีลักษณะสองเท่า: ไอออนิกและ การเผาผลาญปลอม.ประการแรกเกี่ยวข้องกับความจริงที่ว่าการซึมผ่านของโพแทสเซียมในเส้นใยประสาทของเมมเบรนยังคงอยู่เป็นระยะเวลาหนึ่ง (หลายสิบและหลายร้อยมิลลิวินาที) เพิ่มขึ้นหลังจากการสร้าง IVD และเปลี่ยนศักยภาพของเมมเบรนไปสู่ศักยภาพสมดุลโพแทสเซียม การติดตาม hyperpolarization หลังจากการกระตุ้นเซลล์เป็นจังหวะส่วนใหญ่เกี่ยวข้องกับการกระตุ้นปั๊มอิเล็กโตรจีนิกโซเดียมเนื่องจากการสะสมของโซเดียมไอออนในเซลล์
สาเหตุของการดีโพลาไรเซชันที่พัฒนาขึ้นหลังจากการสร้าง IVD (Membrane Action Potential) คือการสะสมของโพแทสเซียมไอออนที่ผิวด้านนอกของเมมเบรน ประการหลังดังต่อไปนี้จากสมการ Goldmann นำไปสู่การเพิ่มขึ้นของ MPP (ศักยภาพในการพักผ่อนของเมมเบรน)
คุณสมบัติที่สำคัญของเส้นใยประสาทเกี่ยวข้องกับการปิดใช้งานช่องโซเดียมที่เรียกว่าการหักเหของแสง .
ระหว่าง absoดุร้ายระยะเวลาทนไฟ เส้นใยประสาทสูญเสียความสามารถในการตื่นเต้นอย่างสิ้นเชิงจากการกระทำของสิ่งที่ทำให้ระคายเคืองต่อความแข็งแรงใด ๆ
ญาติการหักเหของแสงตามค่าสัมบูรณ์มีลักษณะตามเกณฑ์ที่สูงกว่าของ IVD (Membrane Action Potential)
แนวคิดของกระบวนการพังผืดที่เกิดขึ้นระหว่างการกระตุ้นเส้นใยประสาททำหน้าที่เป็นพื้นฐานในการทำความเข้าใจปรากฏการณ์ ที่พัก.หัวใจสำคัญของที่พักเนื้อเยื่อที่ความสูงชันต่ำของการเพิ่มขึ้นของกระแสที่ระคายเคืองคือการเพิ่มขึ้นของเกณฑ์การกระตุ้นซึ่งอยู่ก่อนการทำให้พังผืดช้าลง การเพิ่มขึ้นของเกณฑ์การกระตุ้นนั้นเกือบทั้งหมดถูกกำหนดโดยการปิดใช้งานช่องโซเดียม บทบาทของการเพิ่มขึ้นของความสามารถในการซึมผ่านของโพแทสเซียมของเมมเบรนในการพัฒนาที่พักคือทำให้ความต้านทานของเมมเบรนลดลง เนื่องจากความต้านทานลดลงอัตราการลดขั้วของเมมเบรนจะช้าลง อัตราที่พักจะสูงขึ้นจำนวนช่องโซเดียมที่ศักยภาพในการพักอยู่ในสถานะปิดใช้งานมากขึ้นอัตราการพัฒนาของการหยุดใช้งานจะสูงขึ้นและความสามารถในการซึมผ่านของโพแทสเซียมสูงขึ้นของเมมเบรน
ดำเนินการปลุกเร้า
text_fields
text_fields
arrow_upward
การนำกระแสกระตุ้นไปตามเส้นใยประสาทเกิดขึ้นเนื่องจากกระแสในท้องถิ่นระหว่างส่วนที่ตื่นเต้นและพักผ่อนของเมมเบรน ลำดับเหตุการณ์ในกรณีนี้ถูกนำเสนอดังนี้
เมื่อใช้การกระตุ้นจุดกับเส้นใยประสาทศักยภาพในการกระทำจะเกิดขึ้นในส่วนที่เกี่ยวข้องของเยื่อหุ้มเซลล์ ด้านในของเมมเบรน ณ จุดหนึ่งจะมีประจุบวกเมื่อเทียบกับบริเวณใกล้เคียงโดยวางตัวอยู่ กระแสเกิดขึ้นระหว่างจุดไฟเบอร์ที่มีศักยภาพต่างกัน (กระแสไฟฟ้าในท้องถิ่น),นำจากตื่นเต้น (เครื่องหมาย (+) ที่ด้านในของเมมเบรน) ไปยังไม่ตื่นเต้น (เครื่องหมาย (-) ที่ด้านในของเมมเบรน) ไปยังส่วนเส้นใย กระแสไฟฟ้านี้มีผลต่อการลดขั้วต่อเมมเบรนไฟเบอร์ในพื้นที่พักผ่อนและเมื่อถึงระดับวิกฤตของการลดขั้วของเมมเบรนในบริเวณนี้ IVD (Membrane Action Potential) จะปรากฏขึ้น กระบวนการนี้จะแพร่กระจายไปยังทุกส่วนของเส้นใยประสาทตามลำดับ
ในบางเซลล์ (เซลล์ประสาทกล้ามเนื้อเรียบ) IVD ไม่ได้มีลักษณะของโซเดียม แต่เกิดจากการป้อนข้อมูลของ Ca 2+ ไอออนผ่านช่องแคลเซียมที่ขึ้นกับแรงดันไฟฟ้า ในคาร์ดิโอไมโอไซต์การสร้าง IVD เกี่ยวข้องกับกระแสโซเดียมและโซเดียม - แคลเซียมที่เข้ามา
ในหัวข้อนี้จะพิจารณาสองไอออนบวก - โซเดียม (Na) และโพแทสเซียม (K) เมื่อพูดถึงแอนไอออนให้เราพิจารณาว่าแอนไอออนจำนวนหนึ่งอยู่ที่ด้านนอกและด้านในของเยื่อหุ้มเซลล์
รูปร่างของเซลล์ขึ้นอยู่กับเนื้อเยื่อที่อยู่ ตาม แบบฟอร์ม เซลล์สามารถ:
·ทรงกระบอกและลูกบาศก์ (เซลล์ผิวหนัง);
ดิสนิฟอร์ม (เม็ดเลือดแดง);
ทรงกลม (ไข่);
Fusiform (กล้ามเนื้อเรียบ);
สเตลเลตและเสี้ยม (เซลล์ประสาท);
·ไม่มีรูปแบบถาวร - อะมีบา (เม็ดเลือดขาว)
เซลล์มีจำนวน คุณสมบัติ: มันกินอาหารเติบโตเพิ่มจำนวนฟื้นฟูปรับให้เข้ากับสภาพแวดล้อมแลกเปลี่ยนพลังงานและสารด้วย สิ่งแวดล้อมทำหน้าที่โดยธรรมชาติของมัน (ขึ้นอยู่กับเนื้อเยื่อของเซลล์ที่กำหนด) นอกจากนี้เซลล์ยังมี ความตื่นเต้น
ความตื่นเต้น – มันเป็นความสามารถของเซลล์ในการเคลื่อนที่จากสถานะที่เหลือไปสู่สถานะของกิจกรรมเพื่อตอบสนองต่อสิ่งเร้า
การระคายเคืองอาจมาจากสภาพแวดล้อมภายนอกหรือเกิดขึ้นภายในเซลล์ สิ่งเร้าที่ทำให้เกิดความตื่นเต้น ได้แก่ ไฟฟ้าเคมีเครื่องกลอุณหภูมิและสิ่งเร้าอื่น ๆ
เซลล์สามารถอยู่ในสถานะพื้นฐานได้สองสถานะคือในช่วงพักและอยู่ในอารมณ์ ส่วนที่เหลือและความตื่นเต้นของเซลล์เรียกอีกอย่างว่า - ศักยภาพในการพักตัวของเมมเบรนและศักยภาพในการทำงานของเมมเบรน
เมื่อเซลล์ไม่ได้รับการกระตุ้นใด ๆ แสดงว่าอยู่ในช่วงพัก เซลล์ที่เหลือเรียกว่าแตกต่างกัน ศักยภาพของเมมเบรนพัก (MPP)
ในส่วนที่เหลือพื้นผิวด้านในของเมมเบรนจะมีประจุเป็นลบและพื้นผิวด้านนอกจะมีประจุบวก สิ่งนี้อธิบายได้จากข้อเท็จจริงที่ว่าภายในเซลล์มีแอนไอออนจำนวนมากและไอออนบวกน้อยในขณะที่ด้านหลังเซลล์ตรงกันข้ามไอออนบวกมีชัย
เนื่องจากเซลล์ประกอบด้วย ประจุไฟฟ้าจากนั้นก็วัดกระแสไฟฟ้าที่สร้างขึ้นได้ ค่าของความเป็นไปได้ของเยื่อหุ้มเซลล์คือ: - 70 mV (ลบ 70 เนื่องจากมีประจุลบอยู่ภายในเซลล์) ค่านี้เป็นไปตามอำเภอใจเนื่องจากแต่ละเซลล์อาจมีค่าของศักยภาพในการพัก
ในส่วนที่เหลือรูพรุนของเมมเบรนจะเปิดสำหรับไอออนโพแทสเซียมและปิดสำหรับโซเดียมไอออน นั่นหมายความว่าโพแทสเซียมไอออนสามารถเข้าและออกจากเซลล์ได้ง่าย โซเดียมไอออนไม่สามารถเข้าสู่เซลล์ได้เนื่องจากรูขุมขนของเมมเบรนปิดอยู่ แต่โซเดียมไอออนจำนวนเล็กน้อยจะแทรกซึมเข้าไปในเซลล์เนื่องจากถูกดึงดูดโดยแอนไอออนจำนวนมากที่อยู่บนผิวด้านในของเมมเบรน (มีการดึงดูดประจุตรงกันข้าม) การเคลื่อนที่ของไอออนนี้คือ เฉยๆ , เนื่องจากไม่ต้องใช้พลังงาน
สำหรับกิจกรรมของเซลล์ปกติค่าของ MPP จะต้องอยู่ในระดับคงที่ อย่างไรก็ตามการเคลื่อนที่ของโซเดียมและโพแทสเซียมไอออนบนเมมเบรนทำให้เกิดความผันผวนของค่านี้ซึ่งอาจทำให้ค่าลดลงหรือเพิ่มขึ้น: - 70 mV
เพื่อให้ค่า MPP ค่อนข้างคงที่เรียกว่า โซเดียม - ปั๊มโพแทสเซียม . หน้าที่ของมันคือการกำจัดโซเดียมไอออนออกจากเซลล์และปั๊มโพแทสเซียมไอออนเข้าสู่เซลล์ เป็นอัตราส่วนของโซเดียมและโพแทสเซียมไอออนในเซลล์และหลังเซลล์ที่สร้างค่า MPP ที่ต้องการ การทำงานของปั๊มคือ กลไกที่ใช้งานอยู่ , เพราะต้องใช้พลังงาน
แหล่งพลังงานในเซลล์คือ ATP ATP ให้พลังงานเฉพาะเมื่อแยกออกเป็นกรดที่ง่ายกว่า - ADP โดยมีส่วนร่วมบังคับของเอนไซม์ ATP-ase ในปฏิกิริยา:
ATP + เอนไซม์ ATP-ase ADP + พลังงาน
เซลล์ของสิ่งมีชีวิตทั้งหมดมีความสามารถภายใต้อิทธิพลของสิ่งเร้าที่จะผ่านจากสภาวะที่เหลือทางสรีระไปสู่สภาวะของกิจกรรมหรือความตื่นเต้น
การกระตุ้น เป็นความซับซ้อนของการเปลี่ยนแปลงทางไฟฟ้าเคมีและการทำงานในเนื้อเยื่อที่เคลื่อนไหวได้ (ประสาทกล้ามเนื้อหรือต่อม) ซึ่งเนื้อเยื่อตอบสนองต่ออิทธิพลภายนอก มีบทบาทสำคัญในการกระตุ้นโดยกระบวนการทางไฟฟ้าที่ช่วยให้มั่นใจได้ว่าการกระตุ้นผ่านเส้นใยประสาทและนำเนื้อเยื่อเข้าสู่สถานะ (กำลังทำงาน)
ศักยภาพของเมมเบรน
เซลล์ที่มีชีวิตมีคุณสมบัติที่สำคัญ: พื้นผิวด้านในของเซลล์จะมีประจุลบเสมอเมื่อเทียบกับด้านนอก มีความต่างศักย์ระหว่างพื้นผิวด้านนอกของเซลล์ซึ่งมีประจุไฟฟ้าในแง่ของโปรโตพลาสซึมและด้านในของเยื่อหุ้มเซลล์ซึ่งอยู่ในช่วง 60 ถึง 70 mV ตามข้อมูลของ PG Kostyuk (2001) ในเซลล์ประสาทความแตกต่างนี้อยู่ในช่วง 30 ถึง 70 mV ความต่างศักย์ระหว่างด้านนอกและด้านในของเยื่อหุ้มเซลล์เรียกว่า ศักยภาพของเมมเบรนหรือ ศักยภาพในการพักผ่อน (รูปที่ 2.1)
ศักยภาพของเยื่อหุ้มเซลล์มีอยู่บนเมมเบรนตราบเท่าที่เซลล์ยังมีชีวิตอยู่และหายไปพร้อมกับการตายของเซลล์ L. Galvani ย้อนกลับไปในปี 1794 แสดงให้เห็นว่าหากคุณทำลายเส้นประสาทหรือกล้ามเนื้อ ข้ามส่วน และด้วยการใช้อิเล็กโทรดที่เชื่อมต่อกับกัลวาโนมิเตอร์ไปยังส่วนที่เสียหายและไปยังจุดที่เกิดความเสียหายกัลวาโนมิเตอร์จะแสดงกระแสไฟฟ้าที่ไหลจากส่วนที่ไม่เสียหายไปยังบริเวณที่เกิดรอยบากเสมอ เขาเรียกกระแสนี้ว่ากระแสนิ่ง โดยสาระสำคัญทางสรีรวิทยากระแสไฟฟ้าที่เหลือและศักยภาพของเยื่อหุ้มเซลล์เป็นหนึ่งเดียวกัน ความต่างศักย์ที่วัดได้ในการทดลองนี้คือ 30-50 mV เนื่องจากเมื่อเนื้อเยื่อได้รับความเสียหายกระแสไฟฟ้าส่วนหนึ่งจะถูกตัดออกไปในช่องว่างระหว่างเซลล์และของเหลวที่อยู่รอบ ๆ โครงสร้างที่อยู่ระหว่างการศึกษา ความต่างศักย์สามารถคำนวณได้โดยใช้สูตร Nernst:
โดยที่ R คือค่าคงที่ของก๊าซ T คืออุณหภูมิสัมบูรณ์ F คือหมายเลขฟาราเดย์ [K] ต่อ และ [K] สองชั้น - ความเข้มข้นของโพแทสเซียมภายในและภายนอกเซลล์

รูป: 2.1.
สาเหตุของการเกิดขึ้นของศักยภาพในการพักผ่อนเป็นเรื่องปกติสำหรับทุกเซลล์ มีการกระจายตัวของไอออนที่ไม่สม่ำเสมอ (ความไม่สมดุลของไอออนิก) ระหว่างโปรโตพลาสซึมของเซลล์และสภาพแวดล้อมนอกเซลล์ องค์ประกอบของเลือดมนุษย์ในแง่ของความสมดุลของเกลือคล้ายกับองค์ประกอบของน้ำทะเล สภาพแวดล้อมนอกเซลล์ในระบบประสาทส่วนกลางยังมีโซเดียมคลอไรด์จำนวนมาก องค์ประกอบไอออนิกของไซโทพลาสซึมของเซลล์ด้อยลง ภายในเซลล์มี Na + ไอออนน้อยกว่า 8-10 เท่าและ C ไอออนน้อยกว่า 50 เท่า! "ไอออนบวกหลักของไซโทพลาสซึมคือ K + ความเข้มข้นภายในเซลล์สูงกว่าในสภาพแวดล้อมนอกเซลล์ 30 เท่าและมีค่าประมาณเท่ากับความเข้มข้นภายนอกเซลล์ของ Na ซึ่งเป็นเคาน์เตอร์หลักสำหรับ K + ในไซโทพลาสซึมเป็นแอนไอออนอินทรีย์โดยเฉพาะแอนไอออนของแอสพาร์ติกฮีสตามีนและกรดอะมิโนอื่น ๆ ความไม่สมดุลดังกล่าวเป็นการละเมิดสมดุลทางอุณหพลศาสตร์ในการฟื้นฟูโพแทสเซียมไอออนจะต้องค่อยๆออกจากเซลล์และโซเดียมไอออน - มีแนวโน้มที่จะเป็นเช่นนั้นอย่างไรก็ตามนี่ไม่ใช่ กำลังเกิดขึ้น.
อุปสรรคประการแรกในการทำให้ความแตกต่างของความเข้มข้นของไอออนเท่ากันคือเยื่อหุ้มพลาสมาของเซลล์ ประกอบด้วยโมเลกุลฟอสโฟลิปิดสองชั้นที่ปกคลุมจากด้านในด้วยชั้นของโมเลกุลโปรตีนและจากภายนอกด้วยชั้นของคาร์โบไฮเดรต (mucopolysaccharides) โปรตีนในเซลล์บางส่วนถูกฝังลงใน lipid bilayer โดยตรง สิ่งเหล่านี้เป็นโปรตีนภายใน
โปรตีนเมมเบรนของเซลล์ทั้งหมดแบ่งออกเป็นห้าชั้น: ปั๊มช่องตัวรับเอนไซม์ และ โปรตีนโครงสร้าง ปั๊ม ทำหน้าที่ในการเคลื่อนย้ายไอออนและโมเลกุลกับการไล่ระดับความเข้มข้นโดยใช้พลังงานจากการเผาผลาญ ช่องโปรตีน หรือ รูขุมขน ให้ความสามารถในการซึมผ่านที่เลือก (การแพร่กระจาย) ผ่านเมมเบรนที่มีขนาดไอออนและโมเลกุลที่สอดคล้องกัน โปรตีนตัวรับ มีความจำเพาะสูงรับรู้และผูกติดกับเมมเบรนโมเลกุลหลายประเภทที่จำเป็นสำหรับกิจกรรมที่สำคัญของเซลล์ในช่วงเวลาใดเวลาหนึ่ง เอนไซม์ เร่งการไหล ปฏิกริยาเคมี ที่ผิวเมมเบรน โปรตีนโครงสร้าง ตรวจสอบการเชื่อมต่อของเซลล์เข้ากับอวัยวะและการบำรุงรักษาโครงสร้างใต้เซลล์
โปรตีนทั้งหมดนี้มีความเฉพาะเจาะจง แต่ไม่เคร่งครัด ภายใต้เงื่อนไขบางประการโปรตีนชนิดหนึ่งอาจเป็นได้ทั้งปั๊มเอนไซม์และตัวรับ ผ่านช่องของเมมเบรนโมเลกุลของน้ำและไอออนที่สอดคล้องกับขนาดรูพรุนเข้าและออกจากเซลล์ ความสามารถในการซึมผ่านของเมมเบรนสำหรับไอออนบวกที่แตกต่างกันไม่เหมือนกันและการเปลี่ยนแปลงภายใต้สภาวะการทำงานที่แตกต่างกันของเนื้อเยื่อ ในส่วนที่เหลือเมมเบรนสามารถซึมผ่านโพแทสเซียมไอออนได้มากกว่าไอออนของโซเดียม 25 เท่าและเมื่อรู้สึกตื่นเต้นความสามารถในการซึมผ่านของโซเดียมจะสูงกว่าโพแทสเซียมประมาณ 20 เท่า ในขณะพักความเข้มข้นของโพแทสเซียมในไซโทพลาสซึมและโซเดียมที่เท่ากันในสภาพแวดล้อมนอกเซลล์ควรให้ประจุบวกที่เท่ากันทั้งสองด้านของเมมเบรน แต่เนื่องจากความสามารถในการซึมผ่านของโพแทสเซียมไอออนสูงกว่า 25 เท่าดังนั้นโพแทสเซียมจึงออกจากเซลล์จึงทำให้พื้นผิวของมันมีประจุบวกมากขึ้นเมื่อเทียบกับด้านในของเมมเบรนซึ่งโมเลกุลที่มีประจุลบของแอสพาร์ติกฮีสตามีนและโมเลกุลอื่น ๆ ที่มีขนาดใหญ่เกินไปสำหรับรูพรุนของเมมเบรนจะสะสม กรดอะมิโนที่ "ปล่อย" โพแทสเซียมออกนอกเซลล์ แต่ "ไม่อนุญาต" ให้ไปไกลเนื่องจากมีประจุลบ ประจุลบสะสมที่ด้านในของเมมเบรนและประจุบวกอยู่ด้านนอก ความต่างศักย์เกิดขึ้น กระแสการแพร่กระจายของโซเดียมไอออนไปยังโปรโตพลาสซึมจากของเหลวนอกเซลล์จะรักษาความแตกต่างนี้ไว้ที่ระดับ 60-70 mV ป้องกันไม่ให้เพิ่มขึ้น กระแสการแพร่กระจายของโซเดียมไอออนที่เหลืออยู่จะอ่อนกว่ากระแสโพแทสเซียมไอออน 25 เท่า โซเดียมไอออน, แทรกซึมเข้าไปในเซลล์, ลดค่าของศักยภาพในการพักผ่อนทำให้สามารถเก็บรักษาไว้ได้ในระดับหนึ่ง ดังนั้นค่าของศักยภาพในการพักผ่อนของกล้ามเนื้อและเซลล์ประสาทรวมทั้งเส้นใยประสาทจึงถูกกำหนดโดยอัตราส่วนของจำนวนโพแทสเซียมไอออนที่มีประจุบวกซึ่งกระจายต่อหน่วยเวลาจากเซลล์ไปสู่ภายนอกและโซเดียมไอออนที่มีประจุบวกจะกระจายผ่านเมมเบรนในทิศทางตรงกันข้าม ยิ่งอัตราส่วนนี้สูงเท่าไหร่มูลค่าของศักยภาพในการพักก็จะยิ่งมากขึ้นและในทางกลับกัน
อุปสรรคที่สองที่ทำให้ความต่างศักย์อยู่ในระดับหนึ่งคือปั๊มโซเดียม - โพแทสเซียม (รูปที่ 2.2) เรียกว่าโซเดียม - โพแทสเซียมหรือไอออนิกเนื่องจากมันจะกำจัด (ปั๊มออก) โซเดียมไอออนที่เจาะเข้าไปในโปรโตพลาสซึมและแนะนำ (ฉีด) โพแทสเซียมไอออนเข้าไป แหล่งที่มาของพลังงานสำหรับการทำงานของปั๊มไอออนคือความแตกแยกของ ATP (adenosine triphosphate) ซึ่งเกิดขึ้นภายใต้อิทธิพลของเอนไซม์ adenosine triphosphatase ซึ่งแปลเป็นภาษาท้องถิ่นในเยื่อหุ้มเซลล์และกระตุ้นโดยไอออนเดียวกันเช่นโพแทสเซียมและโซเดียม (ATP-ase ที่ขึ้นกับโซเดียมโพแทสเซียม)

รูป: 2.2.
เป็นโปรตีนขนาดใหญ่ที่มีขนาดใหญ่กว่าความหนาของเยื่อหุ้มเซลล์ โมเลกุลของโปรตีนนี้เจาะเมมเบรนผ่านและทะลุจับโซเดียมและ ATP ที่ด้านในเป็นหลักและโพแทสเซียมและสารยับยั้งประเภทไกลโคไซด์ต่างๆที่ด้านนอก สิ่งนี้ก่อให้เกิดกระแสเมมเบรน เนื่องจากกระแสนี้ทำให้มั่นใจได้ว่ามีทิศทางการขนส่งไอออนที่เหมาะสม การถ่ายเทไอออนเกิดขึ้นในสามขั้นตอน ประการแรกไอออนรวมตัวกับโมเลกุลของผู้ให้บริการเพื่อสร้างไอออนเชิงซ้อนของผู้ให้บริการ จากนั้นคอมเพล็กซ์นี้จะผ่านเมมเบรนหรือถ่ายเทประจุผ่านมัน ในที่สุดไอออนจะถูกปลดปล่อยจากตัวพาที่อยู่ด้านตรงข้ามของเมมเบรน ในเวลาเดียวกันกระบวนการที่คล้ายกันก็เกิดขึ้นโดยถ่ายโอนไอออนไปในทิศทางตรงกันข้าม หากปั๊มถ่ายโอนโซเดียมไอออนหนึ่งไปยังโพแทสเซียมไอออนหนึ่งตัวก็จะรักษาระดับความเข้มข้นของเมมเบรนทั้งสองด้าน แต่ไม่ได้มีส่วนช่วยในการสร้างศักยภาพของเมมเบรน เพื่อให้การสนับสนุนนี้ปั๊มไอออนต้องถ่ายเทโซเดียมและโพแทสเซียมในอัตราส่วน 3: 2 นั่นคือสำหรับโพแทสเซียมไอออน 2 ตัวที่เข้าสู่เซลล์จะต้องกำจัดโซเดียมไอออน 3 ตัวออกจากเซลล์ การทำงานที่โหลดสูงสุดปั๊มแต่ละตัวสามารถสูบโพแทสเซียมไอออนได้ประมาณ 130 และโซเดียมไอออน 200 ต่อวินาทีผ่านเมมเบรน นี่คือสุดยอดความเร็ว ในชีวิตจริงปั๊มแต่ละตัวจะปรับตามความต้องการของกรง เซลล์ประสาทส่วนใหญ่มีปั๊มไอออนระหว่าง 100 ถึง 200 ปั๊มต่อพื้นผิวเมมเบรนสี่เหลี่ยมไมครอน ดังนั้นเมมเบรนของเซลล์ประสาทใด ๆ จึงมีปั๊มไอออน 1 ล้านตัวที่สามารถเคลื่อนที่ได้ถึง 200 ล้านโซเดียมไอออนต่อวินาที
ดังนั้นศักยภาพของเมมเบรน (ศักยภาพในการพักผ่อน) จึงถูกสร้างขึ้นจากกลไกทั้งแบบพาสซีฟและแอคทีฟ ระดับของการมีส่วนร่วมของกลไกบางอย่างในเซลล์ต่าง ๆ ไม่เหมือนกันซึ่งเป็นไปตามที่ศักยภาพของเมมเบรนอาจแตกต่างกันในโครงสร้างที่แตกต่างกัน กิจกรรมของปั๊มอาจขึ้นอยู่กับเส้นผ่านศูนย์กลางของเส้นใยประสาท: ยิ่งเส้นใยบางลงเท่าใดอัตราส่วนของขนาดพื้นผิวต่อปริมาตรของไซโทพลาสซึมจะสูงขึ้นตามลำดับและกิจกรรมของปั๊มที่จำเป็นในการรักษาความแตกต่างของความเข้มข้นของไอออนบนพื้นผิวและภายในเส้นใยจะต้องมากกว่า กล่าวอีกนัยหนึ่งศักยภาพของพังผืดอาจขึ้นอยู่กับโครงสร้างของเนื้อเยื่อประสาทและด้วยเหตุนี้จึงขึ้นอยู่กับวัตถุประสงค์ในการทำงาน โพลาไรเซชันไฟฟ้าของเมมเบรนเป็นเงื่อนไขหลักสำหรับความตื่นเต้นเร้าใจของเซลล์ นี่คือความพร้อมสำหรับการดำเนินการอย่างต่อเนื่องของเธอ นี่คือแหล่งเก็บพลังงานศักย์ของเซลล์ซึ่งสามารถใช้ได้ในกรณีที่ระบบประสาทต้องการปฏิกิริยาทันที







