- διαχειρίζεται. Σύμφωνα με τον μηχανισμό ελέγχου: ηλεκτρικό, αιμο- και μηχανοποιημένο.
- Δεν έχει διαχειριστεί. Μην έχετε φορητό μηχανισμό και είναι πάντα ανοιχτά, τα ιόντα είναι συνεχώς, αλλά αργά.
Πιθανή ανάπαυση - Αυτή είναι η διαφορά των ηλεκτρικών δυνατοτήτων μεταξύ του εξωτερικού και του εξωτερικού και Εσωτερικό περιβάλλον Κύτταρα.
Ο μηχανισμός για το σχηματισμό δυναμικών ανάπαυσης. Η άμεση αιτία του δυναμικού ανάπαυσης είναι η άνιση συγκέντρωση ανιόντων και κατιόντων μέσα και έξω από το κελί. Πρώτον, αυτή η θέση των ιόντων δικαιολογείται από τη διαφορά της διαπερατότητας. Δεύτερον, τα ιόντα καλίου αφήνουν το κύτταρο σημαντικά περισσότερο από το νάτριο.
Δυναμικό δράσης - Αυτή είναι η διέγερση του κυττάρου, η ταχεία διακύμανση του δυναμικού μεμβράνης λόγω της διάχυσης ιόντων στο κύτταρο και από το κύτταρο.
Υπό τη δράση του ερεθιστικού σε κύτταρα με ευγεθμιστό ιστό, πρώτα ενεργοποιούνται γρήγορα ενεργοποιημένα και τα κανάλια νατρίου, τότε τα κανάλια καλίου ενεργοποιούνται και απενεργοποιούνται με κάποια καθυστέρηση.
Κατά συνέπεια, τα ιόντα διαχέουν γρήγορα σε ένα κλουβί ή από αυτήν σύμφωνα με μια ηλεκτροχημική κλίση. Αυτός είναι ο ενθουσιασμός. Με την αλλαγή του μεγέθους και του σημείου των κυττάρων φορτίου, διακρίνονται τρεις φάσεις:
- 1η φάση - αποπόλωση. Μειώνοντας το φορτίο του κελιού στο μηδέν. Το νάτριο κινείται στο κύτταρο σύμφωνα με μια συγκέντρωση και ηλεκτρική κλίση. Κατάσταση κίνησης: Άνοιγμα της πύλης του καναλιού νατρίου.
- Η 2η φάση είναι η αναστροφή. Αλλαγή του σημείου φόρτισης στο αντίθετο. Αναστροφή υποθέτει δύο μέρη: αύξουσα και φθίνουσα.
Αύξουσα μέρος. Το νάτριο συνεχίζει να μετακινείται σε ένα κύτταρο σύμφωνα με μια κλίση συγκέντρωσης, αλλά σε αντίθεση με την ηλεκτρική κλίση (εμπλακεί).
Προς τα κάτω μέρος. Το κάλιο αρχίζει να αφήνει το κύτταρο σύμφωνα με τη συγκέντρωση και την ηλεκτρική κλίση. Οι πύλες του καναλιού καλίου είναι ανοιχτοί.
- Η 3η φάση είναι η επαναπόλωση. Το κάλιο συνεχίζει να αφήνει το κύτταρο σύμφωνα με τη συγκέντρωση, αλλά αντίθετα με την ηλεκτρική κλίση.
Κριτήρια διεγερνότητας
Με την ανάπτυξη του δυναμικού δράσης, εμφανίζεται μια αλλαγή στη διεγερσιμότητα των ιστών. Αυτή η αλλαγή προχωρά με φάσεις. Η κατάσταση της αρχικής πόλωσης του χαρακτηριστικού της μεμβράνης αντικατοπτρίζει το δυναμικό της μεμβράνης της ανάπαυσης, το οποίο αντιστοιχεί στην αρχική κατάσταση της διεγερότητας Α, κατά συνέπεια, η αρχική κατάσταση του κυττάρου με ευγενείς. Αυτό είναι το κανονικό επίπεδο διεγερότητας. Προετοιμασία της περιόδου - την περίοδο από την αρχή του δυναμικού δράσης. Η διεγερσιμότητα των υφασμάτων είναι ελαφρώς αυξημένη. Αυτή η φάση της διεγερότητας είναι η πρωταρχική εξύψωση (πρωτογενής υπερφυσίμενη διέγερση). Κατά την ανάπτυξη της προετοιμασίας, το δυναμικό της μεμβράνης προσεγγίζει το κρίσιμο επίπεδο αποπόλωσης και την επίτευξη αυτού του επιπέδου, η αντοχή του ερεθίσματος μπορεί να είναι μικρότερη από το όριο.
Κατά τη διάρκεια της ανάπτυξης της ακίδας (δυναμικό αιχμής) υπάρχει η ροή του χιονοστιβάδας των ιόντων νατρίου στο εσωτερικό του κυττάρου, ως αποτέλεσμα της οποίας εμφανίζεται η επαναφόρτιση της μεμβράνης και χάνει την ικανότητα να ανταποκρίνεται σε ενθουσιασμό για ερεθίσματα μη θέσης. Αυτή η φάση της διεγερότητας έλαβε το όνομα της απόλυτης ανθεκτικότητας, δηλ. Απόλυτη μη επαναφορά, η οποία διαρκεί μέχρι το τέλος της επαναφόρτησης της μεμβράνης. Η απόλυτη διάθλαση της μεμβράνης προκύπτει λόγω του γεγονότος ότι τα κανάλια νατρίου είναι πλήρως ανοιχτά και στη συνέχεια απενεργοποιημένα.
Μετά τη φάση της φάσης επαναφόρτισης, η διεγερότητα αποκαθίσταται σταδιακά στο αρχικό επίπεδο - αυτή είναι η φάση των σχετικών διαφορτών, δηλ. σχετική μη εμπιστοσύνη. Συνεχίζεται μέχρις ότου η χρέωση της μεμβράνης αποκατασταθεί στην τιμή που αντιστοιχεί στο κρίσιμο επίπεδο αποπόλωσης. Δεδομένου ότι κατά τη διάρκεια αυτής της περιόδου, το δυναμικό της μεμβράνης της ανάπαυσης δεν αποκαθίσταται, τότε η διεγερσιμότητα των ιστών μειώνεται και ο νέος ενθουσιασμός μπορεί να συμβεί μόνο υπό τη δράση ενός ερέθισμα σούπερ τοποθέτησης. Η μείωση της διεξαθειμότητας στη φάση των σχετικών διαθρόσωτων συσχετίζεται με τη μερική απενεργοποίηση των καναλιών νατρίου και την ενεργοποίηση των καναλιών καλίου.
Η επόμενη περίοδος αντιστοιχεί σε αυξημένο επίπεδο διεγερότητας: φάση δευτερογενούς εξύψωσης ή δευτερογενή υπερφυσική διέγερση. Επειδή το δυναμικό της μεμβράνης σε αυτή τη φάση είναι πιο κοντά στο κρίσιμο επίπεδο αποπόλωσης, σε σύγκριση με την κατάσταση της υπόλοιπης αρχικής πόλωσης, το όριο του ερεθισμού μειώνεται, δηλ. Η διεγερσιμότητα των κυττάρων βελτιώνεται. Σε αυτή τη φάση, ο νέος ενθουσιασμός μπορεί να συμβεί υπό τη δράση των ερεθισμάτων της δύναμης της βεράντας. Τα κανάλια νατρίου σε αυτή τη φάση δεν απενεργοποιούνται εντελώς. Το δυναμικό της μεμβράνης αυξάνεται - εμφανίζεται η κατάσταση της υπερ-φορτισμού της μεμβράνης. Αφαίρεση από το κρίσιμο επίπεδο αποπόλωσης, το όριο του ερεθισμού είναι ελαφρώς αυξάνεται και ο νέος ενθουσιασμός μπορεί να συμβεί μόνο υπό τη δράση των ερεθιστικών του συνολικού μεγέθους.
Ο μηχανισμός της εμφάνισης του δυναμικού μεμβράνης ανάπαυσης
Κάθε κύτταρο σε ηρεμία χαρακτηρίζεται από την παρουσία διαμεμβρανικής διαφοράς δυναμικού (δυναμικό ανάπαυσης). Τυπικά, η διαφορά των επιβαρύνσεων μεταξύ των εσωτερικών και εξωτερικών επιφανειών της μεμβράνης είναι από -80 έως -100 mV και μπορεί να μετρηθεί χρησιμοποιώντας εξωτερικά και ενδοκυτταρικά μικροηλεκτώδια (Εικ. 1).
Η πιθανή διαφορά μεταξύ του εξωτερικού και της εσωτερικής πλευράς της μεμβράνης του κελιού στην κατάσταση της πολιτείας της ονομάζεται Δυναμικό μεμβράνης (δυναμικό ανάπαυσης).
Η δημιουργία ειρηνικών δυνατοτήτων παρέχεται από δύο βασικές διαδικασίες - ανομοιογενής διανομή Ανόργανα ιόντα Μεταξύ του ενδοκυτταρικού χώρου και της διαπερατότητας μη-διύλισης γι 'αυτούς της κυτταρικής μεμβράνης. Ανάλυση Χημική σύνθεση Η έξοδος και το ενδοκυτταρικό υγρό υποδεικνύουν την εξαιρετικά ανομοιογενή κατανομή των ιόντων (Πίνακας 1).
Σε κατάσταση ηρεμίας μέσα στο κελί, πολλά ανιόντα οργανικών οξέων και ιόντων Κ +, η συγκέντρωση του οποίου είναι 30 φορές περισσότερο από το εξωτερικό. Na + ιόντα, αντίθετα, έξω από το κελί είναι 10 φορές περισσότερο από το εσωτερικό. Ci και περισσότερο έξω.
Στην υπόλοιπη μεμβράνη των νευρικών κυττάρων, το πιο διαπερατό για το Κ +, λιγότερο - για το Ci- και πολύ λίγο διαπερατό για το Na + / τη διαπερατότητα της μεμβράνης των νευρικών ινών για το Na + B της ανάπαυσης είναι 100 φορές μικρότερο από το K +. Για πολλά ανιόντα, τα οργανικά οξέα της μεμβράνης μόνο είναι εντελώς αδιαπέραστα.
Σύκο. 1. Μέτρηση του δυναμικού συντήρησης των μυϊκών ινών (α) χρησιμοποιώντας ενδοκυτταρικό μικροηλεκτρόδρομο: m - μικροηλεκτρόδρομος. Και - το αδιάφορο ηλεκτρόδιο. Η δέσμη στην οχήστρα του παλμογράφου (Β) δείχνει ότι ο ρυθμός διαφοράς των δυνατοτήτων μεταξύ Μ και ήταν μηδέν στην διάτρηση της μικροηλεκτικής μεμβράνης. Τη στιγμή της διάτρησης (που φαίνεται από το βέλος), ανιχνεύθηκε η διαφορά των δυνατοτήτων, υποδεικνύοντας ότι η εσωτερική πλευρά της μεμβράνης φορτίζεται αρνητικά σε σχέση με την εξωτερική του επιφάνεια (σύμφωνα με τον Β.Ι. Khodorov)
Τραπέζι. Εσωτερικές και εξωκυτταρικές συγκεντρώσεις των ιόντων μυϊκής κυττάρων του θερμού-αιματηρέσου ζώου, mmol / l (από j. Doodle)
|
Ενδοκυτταρική συγκέντρωση |
Εξωκυτταρική συγκέντρωση |
|
|
Α- (ανιόντα οργανικών ενώσεων) |
Λόγω της κλίσης συγκέντρωσης Κ + έρχεται στην εξωτερική επιφάνεια του κελιού, καθιστώντας τη θετική του χρέωση. Τα ανιόντα υψηλού μοριακού βάρους δεν μπορούν να ακολουθήσουν το K + λόγω της αδιαπέρασης για τις μεμβράνες τους. Το NA + ION επίσης δεν μπορεί επίσης να αντισταθμίσει τα ιόντα καλίου, επειδή η διαπερατότητα της μεμβράνης είναι σημαντικά μικρότερη για αυτό. Το Ci-By A Brackation μπορεί να κινηθεί μόνο μέσα στο κύτταρο, αυξάνοντας έτσι το αρνητικό φορτίο της εσωτερικής επιφάνειας της μεμβράνης. Λόγω μιας τέτοιας κίνησης ιόντων, η πόλωση της μεμβράνης συμβαίνει όταν η εξωτερική του επιφάνεια φορτίζεται θετικά και εσωτερικά - αρνητικά.
Το ηλεκτρικό πεδίο, το οποίο θα δημιουργηθεί στη μεμβράνη, θα παρεμβαίνει ενεργά με την κατανομή των ιόντων μεταξύ της εσωτερικής και εξωτερικής περιεκτικότητας του κυττάρου. Καθώς η θετική φόρτιση αυξάνεται στην εξωτερική επιφάνεια του ιόντος κυττάρων έως + καθώς ένα θετικά φορτισμένο γίνεται όλο και πιο δύσκολο να μετακινηθεί από το εσωτερικό. Κινείται σαν ένα βουνό. Όσο μεγαλύτερο είναι το μέγεθος του θετικού φορτίου στην εξωτερική επιφάνεια, όσο μικρότερο είναι ο αριθμός των ιόντων στο + μπορεί να μεταβεί στην επιφάνεια του κελιού. Σε μια ορισμένη ποσότητα δυναμικού στη μεμβράνη, ο αριθμός των ιόντων Κ + που διασχίζει τη μεμβράνη στην άλλη κατεύθυνση είναι ίση, δηλ. Η βαθμίδα συγκέντρωσης του καλίου ισορροπείται από το δυναμικό στη μεμβράνη. Το δυναμικό στο οποίο η ροή διάχυσης των ιόντων καθίσταται ίση με το ρεύμα των ίδιων ονομάτων που πηγαίνουν αντίστροφη κατεύθυνση, που ονομάζεται δυναμικό ισορροπίας για αυτό το ιόν. Για το δυναμικό ισορροπίας IONS K + IS -90 MV. Στις ίνες μυελιδωμένου νεύρου, το μέγεθος του δυναμικού ισορροπίας για ιόντα είναι κοντά στην τιμή του δυναμικού πρόσληψης ανάπαυσης (-70 mV). Επομένως, παρά το γεγονός ότι η συγκέντρωση ιόντων Si-εξωτερικού της ίνας είναι μεγαλύτερη από το εσωτερικό του, δεν υπάρχει μονόπλευρο ρεύμα σύμφωνα με τη βαθμίδα συγκέντρωσης. Στην περίπτωση αυτή, η διαφορά στις συγκεντρώσεις εξισορροπείται από τις δυνατότητες που διατίθεται στη μεμβράνη.
Το NA + ION σύμφωνα με τη βαθμίδα συγκέντρωσης πρέπει να περιλαμβάνεται στο κύτταρο (το δυναμικό ισορροπίας του είναι +60 mV) και η παρουσία αρνητικού φορτίου μέσα στο κελί δεν πρέπει να παρεμποδίζεται σε αυτό το ρεύμα. Στην περίπτωση αυτή, η εισερχόμενη NA + θα εξουδετερώσει τις αρνητικές χρεώσεις μέσα στο κελί. Ωστόσο, αυτό στην πραγματικότητα δεν συμβαίνει, καθώς η μεμβράνη με ειρήνη μειώνεται για Na +.
Ο σημαντικότερος μηχανισμός που υποστηρίζει τη χαμηλή ενδοκυτταρική συγκέντρωση των ιόντων Na + και η υψηλή συγκέντρωση ιόντων Κ + είναι η αντλία του καλίου νατρίου (ενεργή μεταφορά). Είναι γνωστό ότι στην κυτταρική μεμβράνη υπάρχει ένα σύστημα φορέων, καθένα από τα οποία είναι δεσμευτικό με τον γέροντα στα ιόντα Na + μέσα στο κελί και τις εμφανίζει έξω. Από το εξωτερικό, ο φορέας συνδέεται με δύο ιόντα που είναι έξω από το κύτταρο που μεταφέρονται στο κυτταρόπλασμα. Η ενεργειακή παροχή των λειτουργικών συστημάτων παρέχεται από την ΑΤΡ. Η λειτουργία της αντλίας σε ένα τέτοιο σύστημα οδηγεί στα ακόλουθα αποτελέσματα:
- Η υψηλή συγκέντρωση ιόντων στο + μέσα στο κύτταρο διατηρείται, γεγονός που εξασφαλίζει την σταθερότητα της ποσότητας ανάπαυσης. Λόγω του γεγονότος ότι σε κύκλο ανταλλαγής ιόντων από το κελί απεκκρίνεται σε ένα θετικό ιόν περισσότερο από το εισάγεται, η ενεργή μεταφορά διαδραματίζει ρόλο στη δημιουργία ειρηνικού δυναμικού. Σε αυτή την περίπτωση, λένε την ηλεκτρική αντλία, αφού ο ίδιος δημιουργεί ένα μικρό, αλλά Δ.Κ. Θετικές χρεώσεις από το κελί και συνεπώς συμβάλλουν άμεσα συμβολή στο σχηματισμό αρνητικού δυναμικού μέσα σε αυτό. Ωστόσο, το μέγεθος της συμβολής της ηλεκτρικής αντλίας στο Γενική αξία Το δυναμικό των ανθρώπων είναι συνήθως μικρό και κάνει λίγα milvolt.
- Η χαμηλή συγκέντρωση ιόντων Na + διατηρείται μέσα στο κελί, ο οποίος, αφενός, εξασφαλίζει τη λειτουργία του μηχανισμού για τη δημιουργία της ικανότητας της δράσης, από την άλλη - εξασφαλίζει τη διατήρηση της κανονικής οσμωτικής και τον όγκο του κελιού ;
- Διατηρώντας μια σταθερή κλίση συγκέντρωσης Na +, η αντλία καλίου νατρίου συμβάλλει στο συζυγές K +, Na + -Transport αμινοξέα και σάκχαρα μέσω της κυτταρικής μεμβράνης.
Έτσι, η εμφάνιση της διαφοράς διαμεμβρανικής πιθανότητας (δυναμικό ανάπαυσης) οφείλεται στην υψηλή αγωγιμότητα της κυτταρικής μεμβράνης σε κατάσταση ηρεμίας για τα ιόντα Κ +, ιόν, ασυμμετρία ιόντων των συγκεντρώσεων του K + ιόντων και ιόντων Ci-, λειτουργία της ενεργού μεταφοράς Συστήματα (Na + / K + -atfaz), τα οποία δημιουργούν και διατηρούν ασυμμετρία ιόντων.
Δυναμικό νευρικών ινών, νευρική ώθηση
Δυναμικό δράσης - Αυτή η βραχυπρόθεσμη διακύμανση της δυναμικής διαφοράς της μεμβράνης ενός μεγέθους κυττάρων, συνοδευόμενη από μια αλλαγή στην υπογραφή του.
Το δυναμικό δράσης είναι το κύριο ιδιαίτερο χαρακτηριστικό της διέγερσης. Η εγγραφή του υποδηλώνει ότι το κελί ή οι δομές της ανταποκρίθηκαν στις επιπτώσεις της διέγερσης. Ωστόσο, όπως ήδη σημειώθηκε, το PD σε μερικά κύτταρα μπορεί να εμφανιστεί αυθόρμητα (αυθόρμητα). Τέτοια κύτταρα περιέχονται στους οδηγούς του καρδιακού ρυθμού, των τοίχων των αγγείων, νευρικό σύστημα. Το PD χρησιμοποιείται ως φορέας πληροφοριών που το μεταδίδει με τη μορφή ηλεκτρικών σημάτων (ηλεκτρικός συναγερμός) σύμφωνα με τις προσαγωγικές και δραστικές νευρικές ίνες που διεξάγονται από το καρδιακό σύστημα, καθώς και να ξεκινήσουν τη μείωση των μυϊκών κυττάρων.
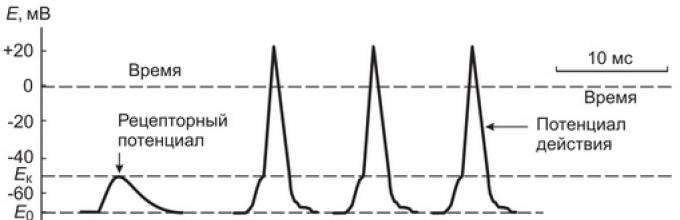
Εξετάστε τις αιτίες και τον μηχανισμό παραγωγής PD σε προσαγωγικές νευρικές ίνες που σχηματίζουν πρωτογενείς αισθητικούς υποδοχείς. Η άμεση αιτία (γενιά) PD σε αυτά είναι ένα δυναμικό υποδοχέα.
Εάν μετρήσετε τη διαφορά στις δυνατότητες στη μεμβράνη που βρίσκεται πλησιέστερα στο νευρικό άκρο της παρακολούθησης του Ranvier, στη συνέχεια στα διαστήματα μεταξύ των επιπτώσεων στην κάψουλα του Ταύρου, παραμένει αμετάβλητη (70 mV) και κατά τη διάρκεια της έκθεσης, είναι Σχεδόν ταυτόχρονα με αποπόλωση της μεμβράνης του υποδοχέα του νευρικού άκρου.
Με αύξηση της δύναμης πίεσης στον Ταύρο του Pacin, ο οποίος προκαλεί αύξηση του δυναμικού υποδοχέα σε 10 MV, στην κοντινή παρακολούθηση του Ranvier, η ταχεία διακύμανση του δυναμικού μεμβράνης συνήθως καταγράφεται, συνοδευόμενη από επαναφόρτωση της μεμβράνης - η ενέργεια πιθανότητες (PD) ή νευρική ώθηση (Εικ. 2). Εάν η δύναμη πίεσης στον καλούντα θα αυξηθεί ακόμη περισσότερο, το πλάτος του δυναμικού υποδοχέα αυξάνεται και ένας αριθμός δυναμικών με μια συγκεκριμένη συχνότητα δημιουργείται στο νευρικό άκρο.

Σύκο. 2. Σχηματική αναπαράσταση του μετασχηματισμού του δυναμικού υποδοχέα στο δυναμικό δράσης (νευρική ώθηση) και της διάδοσης του παλμού στις νευρικές ίνες
Η ουσία του μηχανισμού παραγωγής του PD είναι ότι το δυναμικό υποδοχέα προκαλεί την εμφάνιση τοπικών κυκλικών ρευμάτων μεταξύ της αποπολωμένης μεμβράνης υποδοχέα του μη μικτού τμήματος του νευρικού άκρου και της μεμβράνης της πρώτης παρακολούθησης του Ranvier. Αυτά τα ρεύματα, οι φορείς των οποίων είναι Na +, K +, Cique ιόντα ιόντων ορυκτών, "ροή" όχι μόνο μαζί, αλλά και σε όλη τη μεμβράνη της νευρικής ίνας στον τομέα της αναζωογόνησης παρακολούθησης. Στη μεμβράνη της παρακολούθησης, ο Ranvier, σε αντίθεση με τη μεμβράνη του υποδοχέα, το ίδιο το νευρικό άκρο υπάρχει μεγάλη πυκνότητα ιονικών δυνητικά εξαρτώμενων διαύλων νατρίου και καλίου.
Κατά την επίτευξη της παρακολούθησης του Ranvier της αποπόλωσης της αποπόλωσης περίπου 10 mV, ανακαλύπτεται η ανακάλυψη των ταχειών δυνητικών διαύλων νατρίου και η ροή των ιόντων Na + στερεώνεται στο Axoplasm για μια ηλεκτροχημική κλίση. Καθορίζει την ταχεία αποπόλωση και επαναφόρτιση της μεμβράνης της σύλληψης του Ranvier. Εντούτοις, ταυτόχρονα με την ανακάλυψη των ταχειών που εξαρτώνται από τα ραγδαία δυνητικά κανάλια νατρίου στη συλλογή των Ranviers, αργά δυνητικά εξαρτώμενα δυνητικά κανάλια ανοιχτά και τα ιόντα των ιόντων αρχίζουν να εξέρχονται, τα ιόντα των ιόντων Na + αρχίζουν να εισέρχονται στην είσοδο. Έτσι, σε υψηλή ταχύτητα στο Αξιπλασμό των ιόντων Na + γρήγορα αποπόλωση και επαναφόρτιση για βραχυπρόθεσμα (0,3-0,5 ms) μεμβράνη και τα εξερχόμενα ιόντα Κ + αποκαθιστούν την αρχική κατανομή των χρεώσεων στη μεμβράνη (η μεμβράνη επαναπολλήθηκε) . Ως αποτέλεσμα, κατά τη μηχανική επίδραση στον Ταύρο, η ζωγραφική μιας δύναμης ίση ή υπέρβαση του κατωφλίου, παρατηρείται βραχυπρόθεσμη διακύμανση του δυναμικού υπό μορφή ταχείας αποπόλωσης και επαναπολυγμένης της μεμβράνης στη μεμβράνη του πλησιέστερη παρακολούθηση του Ranvier, δηλαδή PD (νευρική ώθηση) δημιουργείται.
Δεδομένου ότι η άμεση αιτία της δημιουργίας PD είναι ένα δυναμικό υποδοχέα, στη συνέχεια στην περίπτωση αυτή ονομάζεται επίσης δυναμικό γεννήτριας. Ο αριθμός του χρόνου που παράγεται ανά μονάδα του ίδιου πλάτους και η διάρκεια των πάλων των νεύρων είναι ανάλογη προς το πλάτος του δυναμικού υποδοχέα και επομένως, η πίεση στον υποδοχέα. Η διαδικασία μετασχηματισμού των πληροφοριών σχετικά με τη δύναμη της έκθεσης ενσωματωμένη στο πλάτος του δυναμικού υποδοχέα, ο αριθμός των διακριτών νεύρων παλμών έλαβε το όνομα της διακριτής κωδικοποίησης των πληροφοριών.
Οι μηχανισμοί σύνδεσης και η χρονική δυναμική των διαδικασιών παραγωγής PD μελετήθηκαν υπό πειραματικές συνθήκες με τεχνητή πρόσκρουση στις νευρικές ίνες ηλεκτροπληξία Διάφορες αντοχές και διάρκεια.
Τη φύση του δυναμικού της νευρικής ίνας (νευρική ώθηση)
Η μεμβράνη νευρικού ινών στο σημείο τοποθεσίας του ερεθιστικού ηλεκτροδίου ανταποκρίνεται στην επίδραση του πολύ αδύναμου ρεύματος που δεν έχει ακόμη φτάσει στο κατώτατο όριο. Αυτή η απάντηση ονομάστηκε τοπική και η διακύμανση της πιθανής διαφοράς στη μεμβράνη - τοπικό δυναμικό.
Η τοπική ανταπόκριση στη μεμβράνη του κυττάρου με ευγενείς μπορεί να προηγείται της εμφάνισης του δυναμικού της δράσης ή να προκύψει ως ανεξάρτητη διαδικασία. Πρόκειται για μια βραχυπρόθεσμη ταλάντωση (αποπόλωση και επαναπολίωση) δυναμικού ανάπαυσης, που δεν συνοδεύεται από μεμβράνη επαναφόρτισης. Η αποπόλωση της μεμβράνης στην ανάπτυξη του τοπικού δυναμικού οφείλεται σε μια προχωρημένη είσοδο στο Axoplasm των ιόντων Na + και η επαναπόλωση είναι μια καθυστερημένη έξοδος από το Axoplasm των ιόντων σε +.
Εάν επηρεάζετε τη μεμβράνη με ηλεκτρικό ρεύμα αυξανόμενης δύναμης, τότε στην ίδια τιμή που ονομάζεται κατώτατο όριο, η αποπόλωση της μεμβράνης μπορεί να φτάσει στο κρίσιμο επίπεδο - στο οποίο συμβαίνει το άνοιγμα ταχείας δυνητικά εξαρτώμενων καναλιών νατρίου. Ως αποτέλεσμα, εμφανίζεται η avalanche-που μοιάζει με την αύξηση της εισόδου στο κύτταρο Na +. Η προκληθείσα διαδικασία αποπόλωσης αποκτά την αυτοεκτίμηση και το τοπικό δυναμικό αναπτύσσεται στο δυναμικό δράσης.
Το αναφέρουν ήδη Χαρακτηριστικό σημάδι Το PD είναι βραχυπρόθεσμη αντιστροφή (αλλαγή) σημάδι φόρτισης στη μεμβράνη. Έξω από αυτό είναι για ένα μικρό χρονικό διάστημα (0,3-2 ms) φορτίζεται αρνητικά και το εσωτερικό είναι θετικό. Το μέγεθος της αντιστροφής μπορεί να έχει έως και 30 mV και η τιμή του συνολικού δυναμικού δράσης είναι 60-130 mV (Εικ. 3).
Τραπέζι. Συγκριτικά χαρακτηριστικά Τοπικό δυναμικό και δυναμικό δράσης
|
Χαρακτηριστικό γνώρισμα |
Τοπικό δυναμικό |
Δυναμικό δράσης |
|
Αγώγιμο |
Διανέμονται τοπικά, 1-2 mm με εξασθένηση (μείωση) |
Εκτείνεται χωρίς εξασθένηση σε μεγάλες αποστάσεις κατά μήκος ολόκληρου του μήκους της νευρικής ίνας |
|
Νόμος "δυνάμεις" |
Υπακούει |
Δεν υπακούει |
|
Ο νόμος "όλα ή τίποτα" |
Δεν υπακούει |
Υπακούει |
|
Φαινόμενο άθροισης |
Αθροίζει, αυξάνεται με επαναλαμβανόμενες συχνές διαδρομές |
Δεν αθροίζω |
|
Το μέγεθος του πλάτους |
||
|
ΕΓΓΡΑΦΗ ΓΙΑ ΔΙΕΥΘΥΝΣΗ |
Αυξάνεται |
Μειώστε την πλήρη μη εμπιστοσύνη (ανθεκτικό) |
|
Το μέγεθος του ερεθίσματος |
Podchornaya |
Όριο και στο εξωτερικό |
Το δυναμικό δράσης, ανάλογα με τη φύση της μεταβολής των φορτίων στην εσωτερική επιφάνεια της μεμβράνης, χωρίζεται σε φάσεις αποπολίθευσης, επαναπόλωση και υπερυκλώντας της μεμβράνης. Αποπόλωση Καλέστε ολόκληρο το αύξουσα μέρος του PD, σε ποιες περιοχές που αντιστοιχούν στο τοπικό δυναμικό (από το επίπεδο E πριν ΕΔ.), γρήγορη αποπόλωση (από το επίπεδο ΕΔ. σε επίπεδο 0 mV), Αναστροφή Σημάδι φόρτισης (από 0 mV έως μέγιστη τιμή ή ξεκινώντας μια επαναπόλωση). Σαπίλα Αντικατοπτρίζουν το φθίνον τμήμα του PD, το οποίο αντανακλά τη διαδικασία αποκατάστασης της αρχικής πόλωσης της μεμβράνης. Αρχικά, η συμπολιοποίηση πραγματοποιείται γρήγορα, αλλά πλησιάζει το επίπεδο E, Η ταχύτητα SE μπορεί να επιβραδύνει και να καλέσει αυτόν τον ιστότοπο Παρακολούθηση αρνητικότητας (ή ίχνος αρνητικού δυναμικού). Σε ορισμένα κύτταρα, η υπερδιαγίτιση (αυξανόμενη πόλωση της μεμβράνης) αναπτύσσεται μετά την επαναπόλωση. Ονομάζεται εντοπίστε θετικό δυναμικό.
Το αρχικό υψηλό πλάτος ταχεία μέρος του PD καλείται επίσης κορυφή, ή Ακίδα. Περιλαμβάνει φάσεις αποπόλωσης και γρήγορη επαναπόλωση.
Στον μηχανισμό ανάπτυξης PD, ο σημαντικότερος ρόλος ανήκει σε δυνητικά εξαρτώμενα κανάλια ιόντων και μια έγχρωμη αύξηση της διαπερατότητας της κυτταρικής μεμβράνης για ιόντα Na + και K +. Έτσι, υπό δράση σε ένα ηλεκτρικό ρεύμα κυττάρων, προκαλεί αποπόλωση μεμβράνης και, όταν το φορτίο της μεμβράνης μειώνεται σε ένα κρίσιμο επίπεδο (ΕΚ), ανοίγουν πιθανά κανάλια νατρίου. Όπως ήδη αναφέρθηκε, αυτά τα κανάλια σχηματίζονται με πρωτεϊνικά μόρια που ενσωματώνονται στη μεμβράνη, εντός της οποίας υπάρχουν χρόνος και δύο μηχανισμοί φορέα. Ένας από τους μηχανισμούς φορέα είναι η ενεργοποίηση εξασφαλίζει (με τη συμμετοχή του τμήματος 4), το άνοιγμα του καναλιού κατά την αποπόλωση της μεμβράνης και το δεύτερο (με τη συμμετοχή του ενδοκυτταρικού βρόχου μεταξύ του 3ου και του 4ου περιοχές) είναι η αδρανοποίηση, αναπτύσσεται κατά την επαναφόρτιση της μεμβράνης (Εικ. Τέσσερα). Δεδομένου ότι και οι δύο αυτοί μηχανισμοί αλλάζουν γρήγορα τη θέση της πύλης του καναλιού, τότε τα πιθανά εξαρτώμενα κανάλια νατρίου είναι τα ταχεία κανάλια ιόντων. Αυτή η περίσταση προσδιορίζει τη σημασία για τη δημιουργία PD σε ευγεθμισμένους ιστούς και για τις μεμβράνες των νευρικών και μυϊκών ινών.

Σύκο. 3. Το δυναμικό δράσης, οι φάσεις και τα ιωνικά ρεύματα (Α, O). Περιγραφή στο κείμενο

Σύκο. 4. Η θέση της πύλης και η κατάσταση δραστηριότητας δυνητικά εξαρτώμενων διαύλων νατρίου και πιθανών διαύλων σε διάφορα επίπεδα πόλωσης της μεμβράνης
Προκειμένου το πιθανό-εξαρτώμενο κανάλι νατρίου να περάσει στα κύτταρα Na +, είναι απαραίτητο να ανοίξει μόνο μια πύλη ενεργοποίησης, καθώς η απενεργοποίηση της ειρήνης είναι ανοιχτή. Αυτό συμβαίνει όταν η αποπόλωση της μεμβράνης φτάσει σε επίπεδο ΕΔ. (Εικ. 3, 4).
Το άνοιγμα της πύλης ενεργοποίησης των καναλιών νατρίου οδηγεί σε ένα νάτριο που εισέρχεται σε χιονοστιβάδες μέσα στο κύτταρο, κινητά την επίδραση των ηλεκτροχημικών δυνάμεων κλίσης του. Δεδομένου ότι τα ιόντα Na + φέρουν θετική χρέωση, εξουδετερώνουν την περίσσεια αρνητικών φορτίων στην εσωτερική επιφάνεια της μεμβράνης, μειώνουν τη δυναμική διαφορά στη μεμβράνη και την αποπολέστερο. Σύντομα τα ιόντα Na + δίνουν την εσωτερική επιφάνεια της περίσσειας θετικών χρεώσεων μεμβράνης, η οποία συνοδεύεται από μια αναστροφή (μετατόπιση) του σημείου φόρτισης με αρνητικό σε ένα θετικό.
Ωστόσο, τα κανάλια νατρίου παραμένουν ανοιχτά μόνο περίπου 0,5 ms και μετά από αυτή την περίοδο από τη στιγμή
Το PD είναι κλειστές αδρανοποιητικές πύλες, τα κανάλια νατρίου απενεργοποιούνται και αδιαπέραστα για τα ιόντα Na +, η παραλαβή της οποίας είναι δραματικά περιορισμένη μέσα στο κύτταρο.
Από την αποπόλωση της μεμβράνης στο επίπεδο ΕΔ. Η ενεργοποίηση των καναλιών καλίου παρατηρείται επίσης και το άνοιγμα των πύργων τους για ιόντα σε +. Τα ιόντα Κ + υπό τη δράση των δυνάμεων κλίσης συγκέντρωσης βγαίνουν από το κελί, καθιστώντας θετικές χρεώσεις από αυτό. Ωστόσο, ο μηχανισμός φορέα των καναλιών καλίου λειτουργεί αργά και ο ρυθμός παραγωγής θετικών χρεώσεων με ιόντα σε + από τις εκροές των κυψελών προς τα έξω σε σχέση με την είσοδο των ιόντων Na +. Η ροή των ιόντων προς +, αφαιρώντας μια περίσσεια θετικών χρεώσεων από το κύτταρο, καθορίζει την αποκατάσταση της αρχικής κατανομής των χρεώσεων στη μεμβράνη ή την επαναπόλωση και στην εσωτερική πλευρά μετά από μια στιγμή επαναφόρτισης της αρνητικής επιβάρυνσης.
Η εμφάνιση PD σε μεμβράνες με ευγενείς και η επακόλουθη αποκατάσταση του δυναμικού εκκίνησης της ανάπαυσης στη μεμβράνη είναι δυνατή επειδή η δυναμική της εισόδου του κελιού και η έξοδος από το κελί των θετικών φορτίων των ιόντων Na + και K + είναι διαφορετική . Η είσοδος ιόντων Na + είναι μπροστά από την έξοδο του ιόντος έως +. Εάν αυτές οι διαδικασίες ήταν ισορροπία, η πιθανή διαφορά στη μεμβράνη δεν θα αλλάξει. Η ανάπτυξη της ικανότητας να διεγείρει και να δημιουργεί PD ευγεθμισμένους μυς και νευρικά κύτταρα οφείλεται στον σχηματισμό δύο τύπων διαύλων ιόντων πολλαπλών ταχυτήτων στη μεμβράνη τους - γρήγορο νάτριο και αργό κάλιο.
Για να δημιουργηθεί ένα μόνο PD, απαιτείται να εισέλθει σε ένα κύτταρο με σχετικά μικρό αριθμό ιόντων Na +, το οποίο δεν διαταράσσει την κατανομή της και μέσα στο κελί. Κατά τη δημιουργία ενός μεγάλου αριθμού PD, η κατανομή των ιόντων και στις δύο πλευρές της κυτταρικής μεμβράνης θα μπορούσε να διαταραχθεί. Ωστόσο, υπό κανονικές συνθήκες, αυτό εμποδίζεται από το έργο του Na +, K + - Racing.
Στις φυσικές συνθήκες στους νευρώνες του κεντρικού νευρικού συστήματος, το δυναμικό της δράσης αρχικά συμβαίνει στην περιοχή του άξονα του Axonny, στους προσαγωγούς νευρώνες - στον δίπλα στον αισθητήριο υποδοχέα, η παρακολούθηση του Ravvier νευρικού τερματισμού, δηλ. Σε εκείνες τις περιοχές της μεμβράνης, όπου υπάρχουν ταχεία επιλεκτικά δυνητικά δυνητικά κανάλια νατρίου και αργά κανάλια καλίου. Σε άλλους τύπους κυττάρων (για παράδειγμα, pasemecre, λεία μυοκύτταρα), όχι μόνο το νάτριο και το κάλιο, αλλά και τα κανάλια ασβεστίου παίζουν στην εμφάνιση PD.
Οι μηχανισμοί της αντίληψης και της μετατροπής στα σήματα PD σε δευτερεύοντες αισθητικούς αισθητήριους υποδοχείς διαφέρουν από τους μηχανισμούς που αποσυναρμολογούνται για πρωτογενείς αισθητικούς υποδοχείς. Σε αυτούς τους υποδοχείς, η αντίληψη των σημάτων διεξάγεται από εξειδικευμένο νευροσκοπικό (φωτοϋποδοχέα, οσφρητικό) ή ευαίσθητα (γεύση, ακουστικά, αιθουσαία) κύτταρα. Σε κάθε ένα από αυτά τα ευαίσθητα κύτταρα, υπάρχει δικός του, ένας ειδικός μηχανισμός αντίληψης των σημάτων. Ωστόσο, σε όλα τα κύτταρα, η ενέργεια του αντιληπτού σήματος (ερέθισμα) μετατρέπεται σε διακύμανση της δυνητικής διαφοράς της μεμβράνης πλάσματος, δηλ. σε δυναμικό υποδοχέα.
Έτσι, το σημείο κλειδιού στους μηχανισμούς μετασχηματισμού με αισθητήρια κύτταρα των αντιληπτών σημάτων στο δυναμικό του υποδοχέα είναι η μεταβολή της διαπερατότητας των διαύλων ιόντων ως απόκριση στον αντίκτυπο. Το άνοιγμα των καναλιών Na +, Ca2 +, K + -ion κατά την αντίληψη και τον μετασχηματισμό του σήματος επιτυγχάνεται σε αυτά τα κύτταρα με τη συμμετοχή G-πρωτεϊνών, δεύτερων ενδοκυτταρικών διαμεσολαβητών, δεσμεύοντας σε προσδέματα, φωσφορυλίωση των διαύλων ιόντων. Κατά κανόνα, το δυναμικό υποδοχέα εμφανίστηκε σε αισθητήρια κύτταρα προκαλεί την απελευθέρωση τους στην συναπτική σχισμή του νευρώτη, η οποία παρέχει τη μετάδοση του σήματος στη μετασυναπτική μεμβράνη του συναισθηματικού νευρικού άκρου και της παραγωγής στην νευρική της μεμβράνη παλμού. Αυτές οι διαδικασίες περιγράφονται λεπτομερώς στο κεφάλαιο αφιερωμένο σε αισθητήρια συστήματα.
Το δυναμικό δράσης μπορεί να χαρακτηριστεί από πλάτος και διάρκεια, η οποία για την ίδια νευρική ίνα παραμένουν τα ίδια στην διάδοση PD στην ίνα. Ως εκ τούτου, το δυναμικό δράσης ονομάζεται διακριτό δυναμικό.
Υπάρχει μια συγκεκριμένη σχέση μεταξύ της φύσης της πρόσκρουσης στους αισθητικούς υποδοχείς και τον αριθμό των PD που εμφανίζονται στην προσαγωγική νευρική ίνα σε απόκριση στον αντίκτυπο, υπάρχει μια συγκεκριμένη σύνδεση. Βρίσκεται στο γεγονός ότι η μεγάλη αλλά η δύναμη ή η διάρκεια της έκθεσης στη νευρική ίνα σχηματίζεται Περισσότερο νευρικές παρορμήσεις, δηλ. Όταν η έκθεση στο νευρικό σύστημα, οι παλμοί της μεγαλύτερης συχνότητας θα αποστέλλονται από τον υποδοχέα. Οι διαδικασίες για τη μετατροπή των πληροφοριών σχετικά με τη φύση της πρόσκρουσης στη συχνότητα και άλλες παραμέτρους των νεύρων παλμών που μεταδίδονται στο κεντρικό νευρικό σύστημα ονομάστηκαν διακριτική κωδικοποίηση πληροφοριών.
Για επεξήγηση Την προέλευση του δυναμικού ανάπαυσης προτάθηκαν Διάφορες θεωρίες. Στις πηγές της σύγχρονης κατανόησης αυτού του προβλήματος, το έργο του V. Yu. Chavets, ο οποίος το 1896, ο οποίος ήταν ένας φοιτητής ιατρικής, εξέφρασε την ιδέα της ιονικής φύσης των βιοηλεκτρικών διαδικασιών και έκανε μια προσπάθεια εφαρμογής της θεωρίας του Η ηλεκτρολυτική διάσταση του Arrhenius για να εξηγήσει την προέλευση αυτών των δυνατοτήτων. Στο μέλλον, το 1902, ο Υ. Bernstein ανέπτυξε μια θεωρία μεμβράνης ιόντων, η οποία τροποποιήθηκε και τεκμηριώθηκε πειραματικά ο Α. Hodgkin και Α. Huxley (1952) και αυτή τη στιγμή απολαμβάνει ευρεία αναγνώριση. Σύμφωνα με αυτή τη θεωρία, τα βιοηλεκτρικά δυναμικά οφείλονται στην άνιση συγκέντρωση ιόντων σε ֹ, na ֹ, сl "μέσα και έξω από το κύτταρο και διαφορετική διαπερατότητα γι 'αυτούς, η επιφανειακή μεμβράνη.
Το πρωτοπλάσμα των νευρικών και μυϊκών κυττάρων περιέχει 30-50 φορές περισσότερα ιόντα καλίου, 8-10 φορές λιγότερα ιόντα νατρίου και 50 φορές λιγότερα ιόντα χλωρίου, τα οποία εξωκυτταρικό υγρό.
Ένα εμπόδιο στην ταχεία ευθυγράμμιση αυτής της διαφοράς συγκέντρωσης είναι η λεπτότερη (περίπου 100 Α) μεμβράνη πλάσματος, που καλύπτει ζωντανά κύτταρα.
|
Οι παρουσιάσεις στη δομή αυτής της μεμβράνης βασίζονται με βάση τα δεδομένα που λαμβάνονται με μεθόδους ηλεκτρονικής μικροσκοπίας, οπτική μικροσκοπία, περίθλαση ακτίνων Χ και χημική ανάλυση. Θεωρείται ότι η μεμβράνη αποτελείται από ένα διπλό στρώμα μορίων φωσφολιπιδίων επικαλυμμένα από το εσωτερικό με ένα στρώμα πρωτεϊνικών μορίων και έξω από το στρώμα μορίων σύνθετων υδατανθράκων - βλεννοπολυσακχαρίτες. Η δομή των τριών στρωμάτων της μεμβράνης απεικονίζεται σχηματικά Σύκο. 116.. Σύκο. 116. Το διάγραμμα της μοριακής δομής της μεμβράνης. Το στρώμα επισημασμένο με τη δομαλιακή λιπιδική στρώση Ζ εμφανίζεται (οι πολικές ομάδες φωσφολιπιδίων υποδεικνύονται από τους κύκλους) και δύο μη νιπίδια μονοστιβάδες: εξωτερικό-MukopolysacchariD - X, εσωτερική - πρωτεΐνη - Y (από robertson). |
Στην κυτταρική μεμβράνη υπάρχουν οι λεπτότερο σωλήνες - "Πόροι" με διάμετρο σε διάφορα Angstroms. Μέσω αυτών των σωληνωτών μορίων νερού και άλλων ουσιών και τα ιόντα αντιμετώπισης που έχουν το αντίστοιχο μέγεθος πόρων της διαμέτρου περιλαμβάνονται στο κελί από την παραβίαση.
Στα δομικά στοιχεία της μεμβράνης, καταγράφονται προσωπικά ιόντα, τα οποία δίνουν στους τοίχους των πόρων του, αυτό ή αυτό το φορτίο και έτσι καθιστά δύσκολη ή διευκολύνει τα ιόντα μέσω αυτών. Έτσι, υποτίθεται ότι η παρουσία διαχωρισμένου φωσφορικού φωσφορικού και καρβοξυλικών ομάδων στη διαχωρισμένη και καρβοξυλομάδα μεμβράνη είναι ο λόγος που η μεμβράνη των νευρικών ινών είναι σημαντικά λιγότερο διαπερατό για τα κατιόντα.
Η διαπερατότητα της μεμβράνης για διάφορα κατιόντα δεν είναι επίσης η ίδια και φυσικά αλλάζει με διαφορετικές λειτουργικές καταστάσεις του ιστού. Σε μια υπόλοιπη μεμβράνη των νευρικών ινών, περίπου 20-100 φορές πιο διαπερατό για ιόντα σε ֹ από ό, τι για τα ιόντα Na, και όταν η διαπερατότητα του νατρίου είναι ενθουσιασμένη, η διαπερατότητα του νατρίου αρχίζει να υπερβαίνει σημαντικά την διαπερατότητα του καλίου της μεμβράνης.
Προκειμένου να κατανοηθεί ο μηχανισμός της εμφάνισης του δυναμικού ανάπαυσης της μεμβράνης από την άποψη της θεωρίας του Bernstein - Hodgkin, εξετάστε την εμπειρία του μοντέλου. Το πρώτο μισό του σκάφους ( Σύκο. 117.), μια διαχωρισμένη τεχνητή ημι-διαπερατή μεμβράνη, οι πόροι των οποίων διέρχονται ελεύθερα θετικά φορτισμένα ιόντα σε ֹ και δεν περνούν αρνητικά φορτισμένα ιόντα έτσι "4, γεμίζουν με συμπυκνωμένο διάλυμα K2SO4 και το αριστερό μισό γεμίζει επίσης με ένα Διάλυμα K2SO4, αλλά μικρότερη συγκέντρωση.
 |
Λόγω της ύπαρξης ιόντων ιόντων κλίσης συγκέντρωσης σε ֹ, θα αρχίσουν να διαχέουν διαμέσου της μεμβράνης κυρίως από το δεξιό μισό του δοχείου (όπου η συγκέντρωσή τους είναι C1) προς τα αριστερά (με τη συγκέντρωση C2). Συνεπώς, αρνητικά φορτισμένα ανιόντα έτσι "4, για τα οποία η αδιαπέραστη μεμβράνη θα συγκεντρωθεί στο δεξιό μισό του δοχείου στην επιφάνεια της μεμβράνης. Με την αρνητική τους επιβάρυνση, διατηρούν ηλεκτροστατικά τα ιόντα σε ֹ στην επιφάνεια της μεμβράνης στα αριστερά. Ως αποτέλεσμα, η μεμβράνη πολώνει: μεταξύ των δύο επιφανειών υπάρχει μια διαφορά στις δυνατότητες. Σύκο. 117. Η εμφάνιση της δυνητικής διαφοράς στην τεχνητή μεμβράνη που διαχωρίζει τα διαλύματα του K2S04 διαφορετικών συγκεντρώσεων (C1 και C2). Η μεμβράνη είναι επιλεκτικά διαπερατή για κατιόντα σε ֹ (μικρούς κύκλους) και δεν παραλείπει ανιόντα έτσι "4 (μεγάλους κύκλους). 1 και 2 - ηλεκτρόδια μειωμένα στο διάλυμα, 3 - ηλεκτρική συσκευή μέτρησης. |
Εάν τώρα βρίσκεται στο δεξί και το αριστερό μισό του σκάφους για να μειώσει τα ηλεκτρόδια, η ηλεκτρική συσκευή μέτρησης θα ανιχνεύσει τη διαφορά πιθανής πιθανότητας, ενώ το διάλυμα με μικρότερη συγκέντρωση ιόντων K2S04, η οποία λαμβάνει χώρα κατά κύριο λόγο τη διάχυση των θετικά φορτισμένων ιόντων σε ֹ, αποκτά θετική φόρτιση σε σχέση με το διάλυμα με μεγαλύτερη συγκέντρωση K2SO4.
Η διαφορά πιθανής πιθανότητας (ε) στην εξεταζόμενη περίπτωση μπορεί να υπολογιστεί από τον τύπο του Nernst:

Υπάρχουν πολλοί λόγοι να πιστεύουμε ότι παρόμοιες σχέσεις πραγματοποιούνται σε ζωντανές νευρικές ίνες, καθώς η συγκέντρωση ιόντων σε ֹ στο πρωτόπλασμα περισσότερο από σε 30 τριαντάφυλλα υπερβαίνει τη συγκέντρωση αυτών των ιόντων στο εξωτερικό διάλυμα και οργανική (πρωτεΐνη κ.λπ.) των πρωτοπλασμάτων ανιόντων μέσω της μεμβράνης είναι πρακτικά δεν διεισδύουν.
Στην κατάσταση φυσιολογικής ανάπαυσης, η διάχυση θετικά φορτισμένων ιόντων σε ֹ από το πρωτοπλάσμα στο εξωτερικό υγρό δίνει την εξωτερική επιφάνεια της μεμβράνης, μια θετική χρέωση και εσωτερική - αρνητική.
Ένα σημαντικό επιχείρημα υπέρ της ορθότητας αυτής της παρουσίασης ήταν το γεγονός ότι η πιθανή διαφορά μεταξύ της εξωτερικής και της εσωτερικής πλευράς της μεμβράνης μυϊκής ίνας (περίπου 90 mV) ήταν κοντά στο μετρούμενο σε πειράματα χρησιμοποιώντας ένα ενδοκυτταρικό μικροηλεκτρόδρομο.
Θεωρήθηκε επίσης ότι η αύξηση της συγκέντρωσης ιόντων σε ֹ στο εξωτερικό κύτταρο του κυττάρου και κατά συνέπεια, η συνέπεια της συγκέντρωσης αυτών των ιόντων και στις δύο πλευρές της μεμβράνης οδηγεί σε πτώση του δυναμικού ανάπαυσης και σε ένα Ορισμένο εύρος συγκέντρωσης, αυτές οι βάρδιες συμπίπτουν ποσοτικά με τις δυνατότητες που υπολογίζονται από τον τύπο Nernst.
Ωστόσο, η πιο σημαντική, άμεση, απόδειξη της ορθότητας αυτών των ιδεών ελήφθησαν από τον Α. Hodgkin με τους υπαλλήλους (1962) σε πειράματα με την αντικατάσταση του πρωτοπλάσματος σε γιγαντιαίες νευρικές ίνες του μαλακίου καλαμάρια με λύσεις αλατιού. Από τις ίνες που έχει διάμετρο περίπου 1 ml, προσεκτικά συμπιέζεται το πρωτόπλασμα και το κέλυφος πτέρυγας γεμίστηκε με τεχνητή άλμη.
Στην περίπτωση που η συγκέντρωση ιόντων καλίου σε αυτό το διάλυμα ήταν κοντά στην ενδοκυτταρική, η διαφορά πιθανής πιθανότητας προσδιορίστηκε μεταξύ της εσωτερικής και της εξωτερικής πλευράς της μεμβράνης, περίπου ίσο με το δυναμικό της ανθοφορίας της κανονικής ίνας (50-80 mV). Η μείωση της συγκέντρωσης ιόντων σε ֹ στην εσωτερική διάλυμα οδήγησε σε φυσική μείωση ή ακόμη και διαστρέβλωση του δυναμικού ανάπαυσης.
Τέτοια πειράματα έδειξαν ότι η κλίση συγκέντρωσης των ιόντων σε ֹ είναι πράγματι ο κύριος παράγοντας που προσδιορίζει το μέγεθος του δυναμικού της ανάπαυσης της νευρικής ίνας.
Μαζί με τα ιόντα σε ֹ στην εμφάνιση του δυναμικού ανάπαυσης, τα ιόντα Na ֹ συμμετέχουν, διαχέουν στο πρωτόπλασμα από εξωκυτταρικό υγρό, όπου η συγκέντρωσή τους είναι μεγάλη. Διάχυση Αυτό είναι πολύ δύσκολο να μειώσει τη διαπερατότητα του νατρίου μόνο της μεμβράνης. Ωστόσο, η διάχυση μέσω της μεμβράνης μέσα στο πρωτόπλασμα, τα ιόντα Na ֺ ανέχονται τα θετικά τους φορτία εδώ, τα οποία ελαφρώς μειώνει το μέγεθος του δυνατού του δυναμικού ανάπαυσης που δημιουργείται από τη διάχυση από τα κύτταρα ιόντων σε ֺ. Αυτό εξηγεί το γεγονός ότι το δυναμικό της ειρήνης των περισσότερων νευρικών κυττάρων και ινών έχει τιμή όχι 90 mV, όπως θα πρέπει να αναμένεται εάν το δυναμικό ECHTOT δημιουργήθηκε μόνο από ιόντα σε ֺ αλλά 60-70 MV.
Έτσι, η ποσότητα του δυναμικού ανάπαυσης των νευρικών ινών και των κυττάρων προσδιορίζεται από την αναλογία του αριθμού των θετικών φορτισμένων ιόντων σε ֺ διάχυση ανά μονάδα χρόνου από το εξωτερικό των κυττάρων και τα θετικά φορτισμένα ιόντα Na διαχέονται μέσω της μεμβράνης προς την αντίθετη κατεύθυνση. Ποια είναι η αναλογία παραπάνω, τόσο μεγαλύτερη είναι το μέγεθος του δυναμικού ανάπαυσης και αντίστροφα.
text_fields.
text_fields.
arrow_upward
Το δυναμικό της μεμβράνης ανάπαυσης (MPP) ή Πιθανή ανάπαυση (PP) Καλέστε τη διαφορά στις πιθανότητες ενός κυττάρου ανάπαυσης μεταξύ των εσωτερικών και των εξωτερικών πλευρών της μεμβράνης. Η εσωτερική πλευρά της κυτταρικής μεμβράνης φορτίζεται αρνητικά προς την εξωτερική. Λαμβάνοντας το δυναμικό της εξωτερικής λύσης για μηδέν, το MPP καταγράφεται με το σημάδι "μείον". αξία MppΕξαρτάται από τον τύπο του υφάσματος και ποικίλλει από -9 έως -100 mV. Συνεπώς, στη μεμβράνη κυττάρων ανάπαυσης Πολωμένη.Μειώνοντας το ποσό του MPP που ονομάζεται αποπόλωσηαυξάνουν - υπερπολύση,Επαναφορά της τιμής προέλευσης Mpp- σχετικά μεπόλωσηΜεμβράνες.
Οι κύριες διατάξεις της θεωρίας της μεμβράνης προέλευσης MppΠηγαίνετε κάτω στο επόμενο. Σε κατάσταση ανάπαυσης, η κυτταρική μεμβράνη είναι καλά διαπερατή για ιόντα Κ + (σε αριθμό κυττάρων και για SG), λιγότερο διαπερατό για Na + και σχεδόν αδιαπέραστο για ενδοκυτταρικές πρωτεΐνες και άλλα οργανικά ιόντα. Τα ιόντα Κ + διάχυτα από ένα κύτταρο κατά μήκος μιας κλίσης συγκέντρωσης και τα ατομικά ανιόντα παραμένουν στο κυτταρόπλασμα, εξασφαλίζοντας την εμφάνιση της πιθανής διαφοράς μέσω της μεμβράνης.
Η προκύπτουσα δυνητική διαφορά εμποδίζει την έξοδο να + από το κύτταρο και σε ορισμένα από τα σημασία της είναι η ισορροπία μεταξύ της απόδοσης του Κ + κατά μήκος της κλίσης συγκέντρωσης και της εισόδου αυτών των κατιόντων σύμφωνα με την ηλεκτρική κλίση. Το δυναμικό της μεμβράνης στην οποία επιτυγχάνεται αυτή η ισορροπία δυναμικό ισορροπίαςalom.Η αξία του μπορεί να υπολογιστεί από την εξίσωση Nernst:
Οπου ΕΔ.- δυναμικό ισορροπίας για ΠΡΟΣ ΤΗΝ + ; R. - σταθερά αερίου · Τ.- απόλυτη θερμοκρασία. ΦΑ. - Αριθμός Faraday. Π- Valence K + (+1), [Kn +] - [k + vn] -Εξωτερικές και εσωτερικές συγκεντρώσεις σε + -
Εάν πάτε από τους φυσικούς λογάριθμους έως δεκαδικό και υποκαταστήστε τις αριθμητικές τιμές των σταθερών, η εξίσωση θα λάβει τη φόρμα: 
Στους νωτιαί νευρώνες (Πίνακας 1.1) E K \u003d -90 MV. Το μέγεθος του MPP, μετρούμενο από μικροηλεκτρικούς, αισθητά κάτω - 70 mV.
Πίνακας 1.1.. Συγκέντρωση ορισμένων ιόντων μέσα και έξω από τις μυϊκές μορφές των θηλαστικών
| Και αυτος |
Συγκέντρωση |
(mmol / ln 2 o) |
Διορθωτικό δυναμικό (MV) |
|
μέσα στο κελί |
Εξωτερικά κελιά |
||
| Na +. | 15,0 | 150,0 | |
| K +. | 150,0 | 5,5 | |
| Sl - | 125,0 | ||
|
Το δυναμικό της μεμβράνης ανάπαυσης \u003d -70 mV |
|||
Εάν το δυναμικό μεμβράνης του κυττάρου έχει τη φύση του καλίου, στη συνέχεια, σύμφωνα με την εξίσωση Nernst, η τιμή του θα πρέπει να μειώνεται γραμμικά με μείωση της βαθμίδας συγκέντρωσης αυτών των ιόντων, για παράδειγμα, με αύξηση της συγκέντρωσης του k + σε εξωκυτταρικό υγρό . Ωστόσο, η γραμμική εξάρτηση του μεγέθους του MPP (το δυναμικό της μεμβράνης ανάπαυσης) από την κλίση συγκέντρωσης σε + υπάρχει μόνο στη συγκέντρωση του Κ + στο εξωκυτταρικό υγρό πάνω από 20 mm. Σε χαμηλότερες συγκεντρώσεις Κ + έξω από το κύτταρο, η καμπύλη εξάρτησης του ΕΙ από τον λογάριθμο της αναλογίας συγκέντρωσης καλίου από το εξωτερικό όσο και στο εσωτερικό του κυττάρου διαφέρει από το θεωρητικό. Εξηγήστε τις καθιερωμένες αποκλίσεις της πειραματικής εξάρτησης της τιμής MPP και της κλίσης συγκέντρωσης Κ + θεωρητικά υπολογιζόμενη σύμφωνα με την εξίσωση Nernst μπορεί να επιτρέπεται να επιτρέπεται η MPP των διεισδυτικών κυττάρων όχι όχι μόνο με κάλιο, αλλά και δυναμικό ισορροπίας χλωρίου. Διόρθωση ομοίως με το προηγούμενο, μπορείτε να γράψετε:
Τα μεγέθη των δυνατοτήτων ισορροπίας νατρίου και χλωρίου για τους νωτιαί νευρώνες (Πίνακας 1.1) είναι ίσοι, αντίστοιχα, +60 και -70 mV. Η τιμή του ΕΙ είναι ίση με το μέγεθος του MPP. Αυτό υποδεικνύει μια παθητική κατανομή ιόντων χλωρίου μέσω μιας μεμβράνης σύμφωνα με τις χημικές και ηλεκτρικές κλίσεις. Για ιόντα νατρίου, οι χημικές και ηλεκτρικές κλίσεις κατευθύνονται μέσα στο κύτταρο.
Η συμβολή καθενός από τις δυνατότητες ισορροπίας στην τιμή MPP προσδιορίζεται από τον λόγο μεταξύ της διαπερατότητας της κυτταρικής μεμβράνης για κάθε ένα από αυτά τα ιόντα. Ο υπολογισμός του μεγέθους του δυναμικού της μεμβράνης εκτελείται χρησιμοποιώντας την εξίσωση Goldman:

E m.- δυναμικό μεμβράνης · R.- σταθερά αερίου · Τ.- απόλυτη θερμοκρασία. ΦΑ.- τον αριθμό του Faraday. R K, p naκαι R Cl -σταθερές της διαπερατότητας της μεμβράνης για το Κ + Na + και Cl, αντίστοιχα. [ΠΡΟΣ ΤΗΝ + Ν. ], [ Κ. + Vn, [ Na. + Ν. [ Na. + Vn], [CL - H] και [CL - V] - συγκεντρώσεις K +, Na + και Cl Outside (H) και μέσα (HL) κύτταρα.
Αντικατάσταση αυτής της εξίσωσης που λαμβάνονται σε πειραματικές μελέτες των συγκεντρώσεων ιόντων και η ποσότητα του MPP, μπορεί να αποδειχθεί ότι για έναν γιγαντιαίο άξονα, το καλαμάρι πρέπει να είναι η ακόλουθη σχέση των σταθερών διαπερατότητας του p έως p na: p c1 \u003d Ι: 0,04: 0,45. Προφανώς, καθώς η μεμβράνη είναι διαπερατή για ιόντα νατρίου (p n α =/ 0) Και το δυναμικό ισορροπίας για αυτά τα ιόντα έχει ένα σύμβολο συν, η εισαγωγή του τελευταίου μέσα στα κύτταρα σε χημικές και ηλεκτρικές κλίσεις θα μειώσει την ηλεκτροενεργοποίηση του κυτταρόπλασμα, δηλ. Αυξήστε το MPP (το δυναμικό ανάπαυσης της μεμβράνης).
Με την αύξηση της συγκέντρωσης ιόντων καλίου στο εξωτερικό διάλυμα πάνω από 15 mM MPP, η αναλογία των σταθερών διαπερατότητας αλλάζει στην κατεύθυνση ενός σημαντικότερου υπέρβασης του "P έως το P Na και P C1. P έως: P NA: P C1 \u003d 1: 0.025: 0.4. Σε τέτοιες συνθήκες, η MPP προσδιορίζεται σχεδόν αποκλειστικά από την κλίση των ιόντων καλίου, επομένως η πειραματική και θεωρητική εξάρτηση της τιμής του MPP από τον λογάριθμο της σχέσης των συγκεντρώσεων καλίου έξω και μέσα στα κύτταρα αρχίζουν να συμπίπτουν.
Έτσι, η παρουσία σταθερής διαφοράς στις δυνατότητες μεταξύ κυτταρόπλασμα και του εξωτερικού μέσου σε ένα κύτταρο ανάπαυσης οφείλεται στις υπάρχουσες κλίσεις συγκέντρωσης για Κ +, Na + και Cl και η διαφορετική διαπερατότητα της μεμβράνης για αυτά τα ιόντα. Ο κύριος ρόλος στη δημιουργία του MPP παίζει τη διάχυση ιόντων καλίου από το κελί στην εξωτερική βαθμολογία. Μαζί με αυτό, ο MPP προσδιορίζεται επίσης με δυναμικά ισορροπίας νάτριο και χλώριο και η συμβολή καθενός από αυτά καθορίζεται από τις σχέσεις μεταξύ της διαπερατότητας της μεμβράνης πλάσματος των στοιχείων κυττάρων για τα ιόντα.
Όλοι οι παράγοντες που αναφέρονται παραπάνω είναι οι λεγόμενοι ιωνικό στοιχείοMPP (το δυναμικό της μεμβράνης ανάπαυσης). Δεδομένου ότι ούτε οι δυνατότητες ούτε οι δυνατότητες ισορροπίας του νατρίου είναι ίσες με το MPP. Το κύτταρο πρέπει να απορροφά το Na + και να χάσει το +. Η σταθερότητα των συγκεντρώσεων αυτών των ιόντων στο κύτταρο διατηρείται λόγω της λειτουργίας της Na + K + -Atphase.
Ωστόσο, ο ρόλος αυτής της ιονικής αντλίας δεν περιορίζεται στη διατήρηση βαθμίδων του νατρίου και του καλίου. Είναι γνωστό ότι η αντλία ισχύος νατρίου είναι ηλεκτρική και στη λειτουργία του υπάρχει ένα καθαρό ρεύμα θετικών επιβαρύνσεων από ένα κύτταρο σε εξωκυτταρικό υγρό, το οποίο προκαλεί αύξηση της ηλεκτρονεθαίρισης του κυτταρόπλασμα σε σχέση με το μέσο. Η ηλεκτρική ενέργεια της αντλίας νατρίου ανιχνεύθηκε σε πειράματα σε γιγαντιαίους νευρώνες μαλακίων. Η έγχυση ηλεκτροφόρου των ιόντων Na + στο σώμα ενός ενιαίου νευρώνα προκάλεσε την υπερδιαγιοποίηση της μεμβράνης κατά τη διάρκεια της οποίας το MPP ήταν σημαντικά χαμηλότερο από την τιμή του δυναμικού ισορροπίας καλίου. Αυτή η υπερδιβυλίωση εξασθενίστηκε με τη μείωση της θερμοκρασίας του διαλύματος, στην οποία εντοπίστηκε το κύτταρο και καταστέλθηκε με έναν ειδικό αναστολέα Na +, K + -Atphase -Atashase.
Από τα παραπάνω, ακολουθεί ότι το MPP μπορεί να χωριστεί σε δύο συστατικά - "Ιόν"και "Μεταβολικός".Το πρώτο συστατικό εξαρτάται από τις κλίσεις συγκέντρωσης των ιόντων και των διαπερατών μεμβράνης γι 'αυτούς. Το δεύτερο, το "μεταβολικό" οφείλεται στην ενεργό μεταφορά του νατρίου και του καλίου και έχει διπλό αντίκτυπο Mpp.Από τη μία πλευρά, η αντλία νατρίου διατηρεί κλίσεις συγκέντρωσης μεταξύ του κυτταρόπλασμα και του εξωτερικού περιβάλλοντος. Από την άλλη πλευρά, η ηλεκτρική αντλία νατρίου έχει άμεση επίδραση στο MPP. Η συμβολή της στην ποσότητα MPP εξαρτάται από την πυκνότητα του ρεύματος "άντλησης" (ρεύμα ανά μονάδα ανύψωσης της επιφάνειας της κυτταρικής μεμβράνης) και της αντίστασης μεμβράνης.
Δυναμικό δράσης μεμβράνης
text_fields.
text_fields.
arrow_upward
Εάν το νευρικό ή μυς είναι ενοχλητικό πάνω από το όριο διέγερσης, τότε η PPP του νεύρου ή των μυών θα μειωθεί γρήγορα και για σύντομο χρονικό διάστημα (Millisecond) θα εμφανιστεί μεμβράνη επαναφόρτισης: η εσωτερική του πλευρά θα φορτιστεί θετικά σε σχέση με τον εξωτερικό. το Μια σύντομη αλλαγή στο MPP, η οποία συμβαίνει όταν το κύτταρο είναι ενθουσιασμένο, το οποίο το σχήμα μιας μόνο κορυφής στην οθόνη του παλμογράφου καλείται Δυνατότητα δράσης μεμβράνης (MPD).
Το MPD σε νευρικούς και μυϊκούς ιστούς παρουσιάζεται όταν μια μείωση στην απόλυτη τιμή του MPP (αποπόλωση της μεμβράνης) σε μια ορισμένη κρίσιμη τιμή που ονομάζεται Όριο παραγωγήςMpd. Στις γιγαντιαίες νευρικές ίνες, το Kalmar MPD είναι ίσο με 60 mV. Όταν εμφανιστεί η αποπόλωση της μεμβράνης σε -45 mV (κατώφλι παραγωγής MPD) (Εικ. 1.15).
Σύκο. 1.15 Το δυναμικό της νευρικής ίνας (α) και η μεταβολή της αγωγιμότητας της μεμβράνης για ιόντα νατρίου και καλίου (Β).Κατά τη διάρκεια της εμφάνισης MPD στο Squid Acone, η αντίσταση μεμβράνης μειώνει 25 φορές, από 1000 έως 40 ohm. Cm2, ενώ το ηλεκτρικό δοχείο δεν αλλάζει. Η υποδεικνυόμενη μείωση της αντοχής της μεμβράνης οφείλεται σε αύξηση της διαπερατότητας ιόντων της μεμβράνης κατά τη διέγερση.
Σύμφωνα με το πλάτος του (100-120 mV) του MPD (το δυναμικό της μεμβράνης δράσης) κατά 20-50 MV υπερβαίνει την τιμή του MPP (το δυναμικό της μεμβράνης της ειρήνης). Με άλλα λόγια, η εσωτερική πλευρά της μεμβράνης για μικρό χρονικό διάστημα καθίσταται θετικά σε σχέση με το εξωτερικό, "υπέρβαση" ή Χρέωση αναστροφής.
Από την εξίσωση Goldman προκύπτει ότι μόνο η αύξηση της διαπερατότητας της μεμβράνης για ιόντα νατρίου μπορεί να οδηγήσει σε τέτοιες αλλαγές στο δυναμικό του κλιβάνου. Η τιμή Ε είναι πάντα μικρότερη από την τιμή του MPP, επομένως η αύξηση της διαπερατότητας της μεμβράνης για το Κ + θα αυξήσει την απόλυτη τιμή του MPP. Το δυναμικό ισορροπίας του νατρίου έχει ένα σύμβολο συν, επομένως μια απότομη αύξηση της διαπερατότητας της μεμβράνης για αυτά τα κατιόντα οδηγεί σε μια επαναφόρτιση της μεμβράνης.
Κατά τη διάρκεια της MPD, η διαπερατότητα της μεμβράνης για ιόντα νατρίου αυξάνεται. Οι υπολογισμοί έδειξαν ότι εάν η αναλογία των σταθερών σταθερών της διαπερατότητας της μεμβράνης για Κ +, Na + και Sg είναι 1: 0,04: 0,45, στη συνέχεια στο MPD - P έως P Na: p \u003d 1: 20: 0,45. Επομένως, σε κατάσταση διέγερσης της μεμβράνης ινών της Nevice, δεν χάνει απλώς τη διαπερατότητα των εκλογικών ιόντων, αλλά, αντίθετα, γίνεται επιλεκτικά διαπερατό για ιόντα νατρίου. Η αύξηση της διαπερατότητας του νατρίου της μεμβράνης σχετίζεται με το άνοιγμα των πιθανών εξαρτώμενων καναλιών νατρίου.
Ο μηχανισμός που παρέχει άνοιγμα και κλείσιμο των διαύλων ιόντων κλήθηκε Πύλη καναλιών.Είναι αποδεκτό Δραστηριοποίηση(m) και Απενεργοποιητικόςη) πύλη. Το κανάλι ιόντων μπορεί να είναι σε τρεις βασικές καταστάσεις: κλειστή (η πύλη M κλειστό, H-Open), ανοιχτά (m- και h-gates είναι ανοιχτά) και απενεργοποιημένα (m-gates είναι ανοιχτά, η πύλη H είναι κλειστή) ( Εικόνα 1.16).
Σύκο. 1.16 Το σχήμα της ενεργοποίησης (Μ) και της αδρανοποίησης (Η) της πύλης του καναλιού νατρίου που αντιστοιχεί στην κλειστή (Α), ανοικτή (ενεργοποίηση, β) και αδρανοποιημένη (C).
Η αποπόλωση της μεμβράνης που προκαλείται από ένα ενοχλητικό ερέθισμα, όπως ένα ηλεκτρικό ρεύμα, ανοίγει την πύλη M-πύλη των καναλιών νατρίου (μετάβαση από την κατάσταση Α Β) και παρέχει την εμφάνιση θετικών χρεώσεων στη ροή θετικών χρεώσεων - ιόντα νατρίου. Αυτό οδηγεί σε περαιτέρω αποπόλωση της μεμβράνης, η οποία, με τη σειρά του, αυξάνει τον αριθμό των ανοικτών καναλιών νατρίου και επομένως αυξάνει την διαπερατότητα του νατρίου της μεμβράνης. Η "αναγεννητική" αποπόλωση της μεμβράνης προκύπτει, ως αποτέλεσμα της οποίας το δυναμικό του εσωτερικού της μεμβράνης επιδιώκει να επιτύχει ένα δυναμικό ισορροπίας νατρίου.
Ο λόγος για την παύση της ανάπτυξης του MPD (το δυναμικό της μεμβράνης της δράσης) και η επαναπόλωση της μεμβράνης του κελιού είναι:
αλλά) Αυξήστε την αποπόλωση της μεμβράνης, δηλ. Όταν το ΕΙ - "Ε ΝΑ, ως αποτέλεσμα της οποίας μειώνεται μια ηλεκτροχημική κλίση για ιόντα νατρίου, ίσο με το Ε Μ -\u003e Ε. Με άλλα λόγια, η δύναμη μειώνεται, το "σπρώχνει" νάτριο μέσα στο κελί.
σι) Η αποπόλωση της μεμβράνης παράγει τη διαδικασία απενεργοποίησης των καναλιών νατρίου (κλείνοντας την πύλη H, κατάσταση στο κανάλι), το οποίο αναστέλλει την ανάπτυξη της διαπερατότητας του νατρίου της μεμβράνης και οδηγεί στη μείωση του.
σε) Η αποπόλωση μεμβράνης αυξάνει τη διαπερατότητα του για ιόντα καλίου. Το ρεύμα του καλίου εξόδου επιδιώκει να μετατοπίσει το δυναμικό της μεμβράνης προς το δυναμικό ισορροπίας καλίου.
Η μείωση του ηλεκτροχημικού δυναμικού για ιόντα νατρίου και η απενεργοποίηση των καναλιών νατρίου μειώνει το μέγεθος του εισερχόμενου ρεύματος νατρίου. Σε ένα συγκεκριμένο χρονικό σημείο, το μέγεθος του εισερχόμενου ρεύματος νατρίου συγκρίνεται με το αυξημένο εξερχόμενο ρεύμα - η ανάπτυξη IPD τερματίζεται. Όταν το συνολικό εξερχόμενο ρεύμα υπερβαίνει την εισερχόμενη, αρχίζει η επαναπόλωση της μεμβράνης, η οποία είναι επίσης αναγεννητική. Η εκκίνηση επαναποσάτισης οδηγεί στο κλείσιμο των πύλων ενεργοποίησης (m), το οποίο μειώνει την διαπερατότητα του νατρίου της μεμβράνης, επιταχύνει την επαναπόλωση και ο τελευταίος αυξάνει τον αριθμό των κλειστών καναλιών κλπ.
Η φάση επαναπολλατισμού PDA σε μερικά κύτταρα (για παράδειγμα, σε καρδιομυοκύτταρα και αριθμό κυττάρων λείου μυός) μπορεί να επιβραδυνθεί, σχηματίζοντας οροπέδιοPD, λόγω σύνθετων αλλαγών κατά τη στιγμή των εισερχόμενων και αποχωρητών ρευμάτων μέσω της μεμβράνης. Το απόγευμα, η υπερδιολωμάτωση ή / και η αποπόλωση της μεμβράνης μπορεί να εμφανιστούν. Αυτά είναι τα λεγόμενα Δυναμικά.Η διαδρομή Hyperpolarization έχει διπλή φύση: ιωνικόςκαι μεταβολισμόςkui.Το πρώτο οφείλεται στο γεγονός ότι η διαπερατότητα του καλίου στη μεμβράνη νευρικού ινών παραμένει για κάποιο χρονικό διάστημα (δεκάδες και ακόμη και εκατοντάδες χιλιοστόνηση) ανυψώνονται μετά τη δημιουργία MPD και μετατοπίζεται το δυναμικό της μεμβράνης προς το δυναμικό ισορροπίας καλίου. Παρακολουθήστε την υπερδιαγίτιση μετά την διέγερση της ρυθμικής κυττάρου οφείλεται κυρίως στην ενεργοποίηση της ηλεκτρικής αντλίας νατρίου, λόγω της συσσώρευσης ιόντων νατρίου στο κύτταρο.
Η αιτία της αποπόλωσης, η ανάπτυξη μετά τη δημιουργία MPD (δυναμικό μεμβράνης δράσης), είναι η συσσώρευση ιόντων καλίου στην εξωτερική επιφάνεια της μεμβράνης. Το τελευταίο, ως εξής από την εξίσωση Goldman, οδηγεί σε αύξηση του MPP (το δυναμικό της μεμβράνης της ειρήνης).
Με την απενεργοποίηση των καναλιών νατρίου, μια σημαντική ιδιότητα των νευρικών ινών είναι συνδεδεμένη, που ονομάζεταιπυρίμαχος .
Στη διάρκεια ΣούπαςλαούτοΑνθεκτικό περίοδος Η νευρική ίνα χάνει εντελώς την ικανότητα να είναι ενθουσιασμένη υπό τη δράση ενός ερεθισμού οποιασδήποτε εξουσίας.
ΣυγγενήςΔιαθλιμότηταΜετά το απόλυτο, που χαρακτηρίζεται από ένα υψηλότερο όριο για την εμφάνιση του MPD (δυναμικό δράσης μεμβράνης).
Η ιδέα των διαδικασιών μεμβράνης που συμβαίνουν κατά τη διάρκεια της διέγερσης της νευρικής ίνας χρησιμεύει ως βάση για την κατανόηση και το φαινόμενο Κατάλυμα.Στην καρδιά της στέγασης του ιστού σε μικρή αύξηση της αύξησης του ερεθιστικού ρεύματος είναι μια αύξηση του κατωφλίου διέγερσης, μπροστά από την αργή αποπόλωση της μεμβράνης. Η αύξηση του κατωφλίου διέγερσης καθορίζεται σχεδόν εξ ολοκλήρου από την αδρανοποίηση των καναλιών νατρίου. Ο ρόλος της αύξησης της διαπερατότητας του καλίου της μεμβράνης στην ανάπτυξη καταλυμάτων είναι ότι οδηγεί σε πτώση της αντίστασης μεμβράνης. Λόγω της αντίστασης στην αντίσταση, ο ρυθμός αποπόλωσης της μεμβράνης γίνεται ακόμη πιο αργός. Η ταχύτητα διαμονής είναι υψηλότερη από όσο μεγαλύτερη είναι ο αριθμός των διαύλων νατρίου με το δυναμικό ανάπαυσης είναι σε μια αδρανοποιημένη κατάσταση, τόσο μεγαλύτερη είναι η ταχύτητα ανάπτυξης της απενεργοποίησης και όσο υψηλότερη είναι η διαπερατότητα του καλίου της μεμβράνης.
Διέγερση
text_fields.
text_fields.
arrow_upward
Η διέγερση της νευρικής ίνας διεξάγεται εις βάρος των τοπικών ρευμάτων μεταξύ μεμβράνης ενθουσιασμού και ανάπαυσης. Η ακολουθία συμβάντων σε αυτή την περίπτωση εμφανίζεται στην παρακάτω φόρμα.
Όταν εφαρμόζεται ερεθισμός σημείων επί του νευρικού ίνες στο αντίστοιχο τμήμα της μεμβράνης, συμβαίνει το δυναμικό δράσης. Η εσωτερική πλευρά της μεμβράνης σε αυτό το σημείο αποδεικνύεται ότι φορτίζεται θετικά προς τη γειτονική, ανάπαυση. Μεταξύ των σημείων ινών που έχουν διαφορετικό δυναμικό, συμβαίνει ρεύμα (τοπικό ρεύμα),Κατευθύνεται από το ενθουσιασμένο (σημάδι (+) στο εσωτερικό της μεμβράνης) σε ένα μηχάνημα (σημάδι (-) στο εσωτερικό της μεμβράνης) στο τμήμα ινών. Αυτό το ρεύμα έχει αποπολωτική επίδραση στην μεμβράνη ινών σε περιοχή ανάπαυσης και όταν επιτευχθεί το κρίσιμο επίπεδο αποπόλωσης της μεμβράνης σε αυτή την ενότητα, η MPD προκύπτει (δυναμικό δράσης μεμβράνης). Αυτή η διαδικασία κατανέμεται σταθερά σε όλα τα τμήματα της νευρικής ίνας.
Σε μερικά κύτταρα (νευρώνες, λείοι μύες), το MPD δεν έχει φύση νατρίου, αλλά οφείλεται στην είσοδο των ιόντων Ca2 + με δυνητικά εξαρτώμενα κανάλια ασβεστίου. Στα καρδιομυοκύτταρα, η παραγωγή MPD συσχετίζεται με εισερχόμενα ρεύματα ασβεστίου νατρίου και νατρίου.
Σε αυτό το θέμα, θα ληφθούν υπόψη δύο κατιόντα - νάτριο (Na) και κάλιο (Κ). Μιλώντας για ανιόντα, λαμβάνουμε υπόψη ότι μια ορισμένη ποσότητα ανιόντων είναι στο εξωτερικό και στο εσωτερικό της κυτταρικής μεμβράνης.
Η μορφή κυττάρων εξαρτάται από τον οποίο ανήκει ο ιστός. Σύμφωνα με μορφή Τα κύτταρα μπορούν να είναι:
· Κυλινδρικά και κυβικά (κύτταρα του δέρματος).
· Δυοειδή (ερυθρά αιμοσφαίρια).
· Χαρακτήρας (αυγό);
· Πρώτα (λείο μυς).
· Αστέρι και πυραμιδικό (νευρικά κύτταρα).
· Μην έχετε σταθερή μορφή - Amoeboboid (λευκοκύτταρα).
Το κύτταρο έχει κοντά Ιδιότητες: Τροφοδοτεί, αναπτύσσεται, πολλαπλά, αποκατασταθεί, προσαρμοστεί στο περιβάλλον μέσο, \u200b\u200bανταλλαγές με ενέργεια και ουσίες με Περιβαλλοντικός, εκτελεί λειτουργίες που είναι εγγενείς σε αυτό (ανάλογα με τον οποίο ο ιστός έχει ένα δεδομένο κελί). Επιπλέον, το κελί έχει διεγερτικότητα.
Διεγερτικότητα – Αυτή η κυτταρική ικανότητα ως απάντηση στον ερεθισμό για να μετακινηθεί από την κατάσταση της ανάπαυσης στην κατάσταση της δραστηριότητας.
Ο ερεθισμός μπορεί να προέρχεται από ένα εξωτερικό περιβάλλον ή περιστασιακά μέσα στο κελί. Οι ερεισίτες προκαλώντας τον ενθουσιασμό μπορεί να είναι: ηλεκτρική, χημική, μηχανική, θερμοκρασία και άλλα κίνητρα.
Το κελί μπορεί να βρίσκεται σε δύο βασικές καταστάσεις - μόνο και σε διέγερση. Την ειρήνη και τον ενθουσιασμό του κυττάρου αλλιώς κλήση - Μεμβράνη ανάπαυσης και μεμβράνης.
Όταν το κελί δεν αντιμετωπίζει ερεθισμό, είναι σε ηρεμία. Το κλουβί είναι διαφορετικά Δυνατότητα ανάπαυσης μεμβράνης (MPP).
Σε κατάσταση ανάπαυσης, η εσωτερική επιφάνεια της μεμβράνης του φορτίζεται αρνητικά και το εξωτερικό είναι θετικό. Αυτό εξηγείται από το γεγονός ότι μέσα στο κελί υπάρχουν πολλά ανιόντα και λίγα κατιόντα, και η κατιόν του κατιόντος κυριαρχείται από το κύτταρο.
Δεδομένου ότι το κελί είναι παρόν Ηλεκτρικά τέληΗ ηλεκτρική ενέργεια που δημιουργήθηκε από αυτά μπορεί να μετρηθεί. Το μέγεθος του δυναμικού μεμβράνης ανάπαυσης είναι: - 70 mV, (μείον 70, επειδή μέσα στο αρνητικό φορτίο κυττάρων). Αυτή η τιμή είναι υπό όρους, καθώς κάθε κελί μπορεί να έχει δικό του δυναμικό.
Σε μια κατάσταση των πόρων ανάπαυσης, οι μεμβράνες είναι ανοικτές σε ιόντα καλίου και κλειστά στα ιόντα νατρίου. Αυτό σημαίνει ότι τα ιόντα καλίου μπορούν εύκολα να διεισδύσουν στο κλουβί και να το αφήσουν. Τα ιόντα νατρίου δεν μπορούν να εισέλθουν στο κλουβί, καθώς οι πόροι της μεμβράνης είναι κλειστές. Αλλά ένας μικρός αριθμός ιόντων νατρίου διεισδύει σε ένα κύτταρο, επειδή προσελκύονται από ένα μεγάλο αριθμό ανιόντων που βρίσκονται στην εσωτερική επιφάνεια της μεμβράνης (πολυδιάστατες χρεώσεις προσελκύονται). Αυτή η κίνηση των ιόντων είναι Παθητικός , Δεδομένου ότι δεν απαιτεί ενεργειακά έξοδα.
Για την κανονική ζωή, η κυτταρική τιμή του MPP του πρέπει να παραμείνει σε σταθερό επίπεδο. Ωστόσο, η κίνηση των ιόντων του νατρίου και του καλίου μέσω της μεμβράνης προκαλεί δονήσεις αυτής της τιμής, η οποία μπορεί να οδηγήσει σε μείωση ή αύξηση της αξίας: - 70 mV.
Προκειμένου η τιμή MPP να παραμείνει σχετικά σταθερή αρχίζει να ενεργεί το λεγόμενο Αντλία νατρίου - καλύμματος . Η λειτουργία του έγκειται στο γεγονός ότι καθαρίζεται με ιόντα νατρίου από τα κύτταρα και τα ιόντα καλίου που εγχέονται στο κλουβί. Είναι μια ορισμένη αναλογία ιόντων νατρίου και καλίου σε ένα κύτταρο και το κύτταρο δημιουργεί την επιθυμητή τιμή MPP. Το έργο της αντλίας είναι ενεργό μηχανισμό , Επειδή απαιτεί ενέργεια.
Η πηγή ενέργειας στο κελί είναι ΑΤΡ. Το ATP δίνει ενέργεια μόνο κατά τη διάσπαση σε απλούστερο οξύ - ADP, με δεσμευτική συμμετοχή στην αντίδραση ενζύμου ΑΤΡ:
ATP + ATF-AZA ADF + Ενέργεια
Όλα τα ζωντανά κύτταρα έχουν την ικανότητα να επηρεάζονται από ερεθιστικά να μετακινηθούν από την κατάσταση της φυσιολογικής ανάπαυσης στην κατάσταση της δραστηριότητας ή τον ενθουσιασμό.
Διέγερση - Πρόκειται για ένα συγκρότημα ενεργών ηλεκτρικών, χημικών και λειτουργικών αλλαγών σε ειδικούς ιστούς (νευρικό, μυϊκό ή σίδηρο), το οποίο το ύφασμα ανταποκρίνεται στην εξωτερική επιρροή. Ένας σημαντικός ρόλος στη διέγερση αναπαράγεται από ηλεκτρικές διαδικασίες που εξασφαλίζουν τη διέγερση των νευρικών ινών και των κορυφαίων ιστών σε ένα ενεργό (λειτουργικό) κατάσταση.
Δυναμικό μεμβράνης
Τα ζωντανά κύτταρα έχουν σημαντική ιδιότητα: η εσωτερική επιφάνεια του κελιού φορτίζεται πάντα αρνητικά σε σχέση με την εξωτερική του πλευρά. Υπάρχει μια διαφορά στις δυνατότητες μεταξύ της εξωτερικής επιφάνειας του κυττάρου, τα κύτταρα που φορτώθηκαν στην ηλεκτροπροστασία σε σχέση με το πρωτόπλασμα και την εσωτερική πλευρά της κυτταρικής μεμβράνης υπάρχει μια πιθανή διαφορά που κυμαίνεται στην περιοχή των 60-70 mV. Σύμφωνα με τον P. G. Kozheuk (2001), στο νευρικό κύτταρο, αυτή η διαφορά κυμαίνεται από 30 έως 70 mV. Η διαφορά των δυνατοτήτων μεταξύ των εξωτερικών και των εσωτερικών πλευρών της κυτταρικής μεμβράνης ονομάζεται δυναμικό μεμβράνηςή Πιθανή ανάπαυση (Εικ. 2.1).
Το δυναμικό ανάπαυσης της μεμβράνης υπάρχει στη μεμβράνη μέχρις ότου το κύτταρο είναι ζωντανό και εξαφανίζεται με το θάνατο του κυττάρου. L. Γαλβανία το 1794 έδειξε ότι αν βλάψετε το νεύρο ή το μυ, κάνοντας διατομή Και βάζοντας στο κατεστραμμένο τμήμα και στον τόπο βλάβης στα ηλεκτρόδια που συνδέονται με το γαλβανόμετρο, το γαλβανόμετρο θα δείξει το ρεύμα που ρέει πάντοτε από το άθικτο τμήμα του υφάσματος στον τόπο της κοπής. Αυτή η ροή κάλεσε ανάπαυση. Με τη φυσιολογική ουσία του, το ρεύμα ανάπαυσης και το δυναμικό της μεμβράνης της ανάπαυσης είναι το ίδιο. Η πιθανή διαφορά που μετράται σε αυτή την εμπειρία είναι 30-50 mV, αφού όταν ο ιστός έχει υποστεί βλάβη, μέρος του ρεύματος είναι χαλασμένο στον ενδοκυτταρικό χώρο και τη γύρω δομή του υγρού. Η πιθανή διαφορά μπορεί να υπολογιστεί χρησιμοποιώντας το Nernst Formula:
Όπου r είναι σταθερά αερίου, T - απόλυτη θερμοκρασία, F είναι ο αριθμός του Faraday, [K] VN. και [k] nar. - Συγκέντρωση του καλίου μέσα και έξω από το κύτταρο.

Σύκο. 2.1.
Ο λόγος για την εμφάνιση του δυναμικού ανάπαυσης για όλα τα κύτταρα. Μεταξύ του κυττάρου πρωτοπλασμάτων και του εξωκυτταρικού μέσου υπάρχει μια ανώμαλη κατανομή ιόντων (ασυμμετρία ιόντων). Η σύνθεση του αίματος ενός ανθρώπου σε ισορροπία άλατος μοιάζει με τη σύνθεση του ωκεανού νερού. Το εξωκυτταρικό μέσο στο κεντρικό νευρικό σύστημα περιέχει επίσης ένα πολύχρωμο νάτριο. Η σύνθεση ιόντων κυττάρων κυτταροπλάσματος φτωχών. Μέσα στα κύτταρα 8-10 φορές μικρότερο από τα ιόντα Na + και 50 φορές λιγότερο από τα ιόντα! ". Η κύρια κατιόν του κυτταροπλάσματος είναι Κ +. Η συγκέντρωσή του μέσα στο κύτταρο είναι 30 φορές υψηλότερη από ό, τι στο εξωκυτταρικό μέσο και περίπου ίσο με το Η εξωκυτταρική συγκέντρωση NA τα κύρια αντίθετα στοιχεία για το Κ + σε κυτταρόπλασμα είναι οργανικά ανιόντα, ειδικά ανιόντα ασπαραγινικά, ισταμίνη και άλλα αμινοξέα. Τέτοια ασυμμετρία είναι μια παραβίαση της θερμοδυναμικής ισορροπίας. Για να το αποκατασταθεί, τα ιόντα καλίου πρέπει να εγκαταλείψουν σταδιακά το κλουβί, και τα ιόντα νατρίου πρέπει να το αγωνίσουν. Ωστόσο, αυτό δεν συμβαίνει.
Το πρώτο εμπόδιο για την ευθυγράμμιση της διαφοράς στις συγκεντρώσεις ιόντων είναι η μεμβράνη πλάσματος του κελιού. Αποτελείται από ένα διπλό στρώμα μορίων φωσφολιπιδίων επικαλυμμένα από το εσωτερικό με ένα στρώμα πρωτεϊνικών μορίων και έξω από το στρώμα των υδατανθράκων (Mucopolisachariums). Ορισμένες από τις κυτταρικές πρωτεΐνες χτίζονται απευθείας σε ένα διπλό λιπιδικό στρώμα. Αυτές είναι εσωτερικές πρωτεΐνες.
Οι πρωτεΐνες μεμβράνης όλων των κυττάρων χωρίζονται σε πέντε κατηγορίες: Αντλίες, κανάλια, υποδοχείς, ένζυμα και Δομικές πρωτεΐνες. Γοβάκια Χρησιμεύουν για να μετακινήσετε ιόντα και μόρια έναντι βαθμίδων συγκέντρωσης χρησιμοποιώντας μεταβολική ενέργεια για αυτό. Πρωτεϊνικά κανάλια ή Πόροι Παρέχουν επιλεκτική διαπερατότητα (διάχυση) διαμέσου της μεμβράνης των αντίστοιχων ιόντων και μορίων. Πρωτεΐνες υποδοχέα, Διαθέτοντας υψηλή εξειδίκευση, αναγνωρίζουν και δεσμεύουν, συνδέοντας με τη μεμβράνη, πολλοί τύποι μορίων που απαιτούνται για τη ζωή των κυττάρων σε κάθε φορά. Ένζυμα Επιταχύνετε το ρεύμα Χημικές αντιδράσεις Στην επιφάνεια της μεμβράνης. Δομικές πρωτεΐνες Παρέχετε κυτταρική σύνδεση με όργανα και διατηρώντας μια υποκυτταρική δομή.
Όλες αυτές οι πρωτεΐνες είναι συγκεκριμένες, αλλά όχι αυστηρά. Σε ορισμένες συνθήκες, μία ή άλλη πρωτεΐνη μπορεί ταυτόχρονα την αντλία και το ένζυμο και τον υποδοχέα. Μέσα από τα κανάλια μεμβράνης των μορίων νερού, καθώς και τους αντίστοιχους πόρους των πόρων, τα ιόντα περιλαμβάνονται στο κελί και βγαίνουν από αυτό. Η διαπερατότητα της μεμβράνης για διάφορα κατιόντα δεν είναι η ίδια και ποικίλλει με διαφορετικές λειτουργικές καταστάσεις του ιστού. Στην υπόλοιπη μεμβράνη, 25 φορές πιο διαπερατή για ιόντα καλίου από ό, τι για ιόντα νατρίου, και όταν η διαπερατότητα του νατρίου είναι ενθουσιασμένη κατά περίπου 20 φορές το κάλιο. Σε κατάσταση ανάπαυσης, ίσες συγκεντρώσεις καλίου σε κυτταρόπλασμα και νάτριο στο εξωκυτταρικό μέσο θα πρέπει να εξασφαλίζουν ίσο αριθμό θετικών τελών και στις δύο πλευρές της μεμβράνης. Όσον αφορά τη διαπερατότητα των ιόντων καλίου άνω των 25 φορές, στη συνέχεια το κάλιο, αφήνοντας το κύτταρο, καθιστά την επιφάνεια του όλο και περισσότερο φορτισμένη σε σχέση με το εσωτερικό της μεμβράνης, κοντά στο οποίο τα αρνητικά φορτισμένα μόρια ασπαρμίας, ισταμίνη και άλλα συσσωρεύονται όλο και περισσότερο για τους πόρους της μεμβράνης Τα αμινοξέα, το "κυκλοφόρησε" κάλιο έξω από το κελί, αλλά "δεν τον δίνει" τον να πάει πολύ λόγω της αρνητικής τους επιβάρυνσης. Τα αρνητικά τέλη συσσωρεύονται από το εσωτερικό της μεμβράνης και με εξωτερική θετική. Υπάρχει μια διαφορά στις δυνατότητες. Το διάχυτο ρεύμα ιόντων νατρίου στο πρωτόπλασμα από εξωκυτταρικό υγρό διατηρεί αυτή τη διαφορά στο επίπεδο των 60-70 mV, χωρίς να το αφήνει να αυξηθεί. Το διάχυτο ρεύμα των ιόντων νατρίου μόνο είναι 25 φορές ασθενέστερη από το επόμενο ρεύμα των ιόντων καλίου. Τα ιόντα νατρίου, που διεισδύουν μέσα στο κύτταρο, μειώνουν το μέγεθος του δυναμικού ανάπαυσης, επιτρέποντάς του να διατηρηθεί σε ένα ορισμένο επίπεδο. Έτσι, η ποσότητα των μυών και των νευρικών δυνατοτήτων, καθώς και τις νευρικές ίνες, καθορίζεται από τον λόγο του αριθμού των θετικά φορτισμένων ιόντων καλίου που διαφέρει ανά μονάδα χρόνου από το εξωτερικό των κυττάρων και θετικά φορτισμένα ιόντα νατρίου που διαμορφώνεται μέσω της μεμβράνης στο αντίθετη κατεύθυνση. Ποια είναι η αναλογία παραπάνω, τόσο μεγαλύτερη είναι το μέγεθος του δυναμικού ανάπαυσης και αντίστροφα.
Το δεύτερο εμπόδιο στη δυνητική διαφορά σε ένα ορισμένο επίπεδο είναι η αντλία καλίου νατρίου (Εικ. 2.2). Ονομάστηκε κάλιο ή ιοντικό νατρίου, δεδομένου ότι πραγματοποιεί ενεργό ανασκαφή (άντληση) από το πρωτόπλασμα ιόντων νατρίου που διεισδύει σε αυτήν και την εισαγωγή ιόντων καλίου σε αυτό. Η πηγή ενέργειας για τη λειτουργία της αντλίας ιόντων είναι η διάσπαση του ΑΤΡ (τριφωσφορικού αδενοσίνης), η οποία συμβαίνει υπό την επίδραση του ενζύμου της αδενοσυνοδιρρυστάσης, που εντοπίζεται στην κυτταρική μεμβράνη και τα ίδια ιόντα που ενεργοποιούνται από τα ίδια ιόντα, δηλαδή, κάλιο και νατρίου (εξαρτώμενος από τον κάλιο ΑΤΡ-ΑΖΑ).

Σύκο. 2.2.
Αυτή είναι μια μεγάλη πρωτεΐνη που υπερβαίνει το πάχος της κυτταρικής μεμβράνης. Το μόριο αυτής της πρωτεΐνης, διείσδυση της μεμβράνης διαμέσου, δεσμεύεται από το εσωτερικό κυρίως νατρίου και ΑΤΡ, και με υπαίθριο κάλιο και διάφορους αναστολείς του τύπου των glyco-Zides. Αυτό συμβαίνει ένα ρεύμα μεμβράνης. Χάρη σε αυτό το ρεύμα, παρέχεται κατάλληλη κατεύθυνση της μεταφοράς ιόντων. Η μεταφορά ιόντων συμβαίνει σε τρία στάδια. Πρώτον, το ιόν συνδέεται με το μόριο φορέα, σχηματίζοντας ένα σύμπλοκο ιόντων-φορέα. Στη συνέχεια, αυτό το συγκρότημα περνά μέσα από τη μεμβράνη ή μεταφέρει τη φόρτιση μέσω αυτού. Στο τέλος - το ιόν απελευθερώνεται από τον φορέα στην αντίθετη πλευρά της μεμβράνης. Ταυτόχρονα, υπάρχει μια παρόμοια διαδικασία που φέρει ιόντα προς την αντίθετη κατεύθυνση. Εάν η αντλία μεταφέρει ένα ιόν νατρίου σε ένα ιόν καλίου, υποστηρίζει απλώς μια κλίση συγκέντρωσης και στις δύο πλευρές της μεμβράνης, αλλά δεν συμβάλλει στη δημιουργία δυναμικού μεμβράνης. Για να γίνει αυτή η συμβολή, η αντλία ιόντων πρέπει να μεταφέρει νατρίου και καλίου σε αναλογία 3: 2, δηλ. Σε 2 ιόντα καλίου που εισέρχονται στο κύτταρο, θα πρέπει να εξάγουμε 3 ιόντα νατρίου από το κύτταρο. Εργασία με το μέγιστο φορτίο, κάθε αντλία είναι ικανή να αντλείται μέσω της μεμβράνης περίπου 130 ιόντα καλίου και 200 \u200b\u200bιόντα νατρίου ανά δευτερόλεπτο. Αυτή είναι η οριακή ταχύτητα. Σε πραγματικές συνθήκες, το έργο κάθε αντλίας ρυθμίζεται σύμφωνα με τις ανάγκες του κελιού. Για τους περισσότερους νευρώνες, μια επιφάνεια μεμβράνης ενός τετραγωνικού μικρού μεγέθους αντιπροσώπευε 100 έως 200 αντλίες ιόντων. Κατά συνέπεια, η μεμβράνη οποιουδήποτε νευρικού κυττάρου περιέχει 1 εκατομμύριο ιόντα αντλίες ικανές να μετακινούνται μέχρι 200 \u200b\u200bεκατομμύρια ιόντα νατρίου ανά δευτερόλεπτο.
Έτσι, το δυναμικό της μεμβράνης (δυναμικό ανάπαυσης) δημιουργείται ως αποτέλεσμα τόσο των παθητικών όσο και των ενεργών μηχανισμών. Ο βαθμός συμμετοχής ορισμένων μηχανισμών σε διαφορετικά κύτταρα μη-Ειναλάκφ, από την οποία προκύπτει ότι το δυναμικό της μεμβράνης μπορεί να είναι άνισο σε διαφορετικές δομές. Η δραστικότητα των αντλιών μπορεί να εξαρτάται από τη διάμετρο των νευρικών ινών: ο λεπτότερος της ίνας, η αναλογία του μεγέθους της επιφάνειας στον όγκο του κυτταρόπλασμα παραπάνω, αντίστοιχα, η δραστηριότητα των αντλιών που απαιτούνται για τη διατήρηση της διαφοράς στο Οι συγκεντρώσεις ιόντων στην επιφάνεια και στο εσωτερικό της ίνας πρέπει να είναι μεγαλύτερες. Με άλλα λόγια, το δυναμικό της μεμβράνης μπορεί να εξαρτάται από τη δομή νευρικός ιστόςΈτσι, από το λειτουργικό του σκοπό. Η ηλεκτρική πόλωση της μεμβράνης είναι η κύρια προϋπόθεση που εξασφαλίζει τη διεγερσιμότητα του κελιού. Αυτή είναι η συνεχής ετοιμότητα για δράση. Αυτό το αποθεματικό δυναμική ενέργεια Κύτταρα που μπορούν να χρησιμοποιήσουν σε περίπτωση που το νευρικό σύστημα θα χρειαστεί την άμεση απάντησή του.





