- управляемые. По механизму управления: электро-, хемо- и механоуправляемые;
- неуправляемые. Не имеют воротного механизма и всегда открыты, ионы идут постоянно, но медленно.
Потенциал покоя — это разность электрических потенциалов между наружной и внутренней средой клетки.
Механизм формирования потенциалов покоя. Непосредственная причина потенциала покоя — это неодинаковая концентрация анионов и катионов внутри и вне клетки. Во-первых, такое расположение ионов обосновано разницей проницаемости. Во-вторых, ионов калия выходит из клетки значительно больше, чем натрия.
Потенциал действия — это возбуждение клетки, быстрое колебание мембранного потенциала вследствие диффузии ионов в клетку и из клетки.
При действии раздражителя на клетки возбудимой ткани сначала очень быстро активируются и инактивируются натриевые каналы, затем с некоторым опозданием активируются и инактивируются калиевые каналы.
Вследствие этого ионы быстро диффундируют в клетку или из нее согласно электрохимическому градиенту. Это и есть возбуждение. По изменению величин и знака заряда клетки выделяют три фазы:
- 1-я фаза — деполяризация. Уменьшение заряда клетки до нуля. Натрий движется к клетке согласно концентрационному и электрическому градиенту. Условие движения: открыты ворота натриевого канала;
- 2-я фаза — инверсия. Изменение знака заряда на противоположный. Инверсия предполагает две части: восходящую и нисходящую.
Восходящая часть. Натрий продолжает двигаться в клетку согласно концентрационному градиенту, но вопреки электрическому градиенту (он препятствует).
Нисходящая часть. Калий начинает выходить из клетки согласно концентрационному и электрическому градиенту. Открыты ворота калиевого канала;
- 3-я фаза — реполяризация. Калий продолжает выходить из клетки согласно концентрационному, но вопреки электрическому градиенту.
Критерии возбудимости
При развитии потенциала действия происходит изменение возбудимости ткани. Это изменение протекает по фазам. Состояние исходной поляризации мембраны характерно отражает мембранный потенциал покоя, которому соответствует исходное состояние возбудимости а, следовательно, исходное состояние возбудимой клетки. Это нормальный уровень возбудимости. Период предспайка — период самого начала потенциала действия. Возбудимость ткани слегка повышена. Эта фаза возбудимости — первичная экзальтация (первичная супернормальная возбудимость). Во время развития предспайка мембранный потенциал приближается к критическому уровню деполяризации и для достижения этого уровня сила раздражителя может быть меньше пороговой.
В период развития спайка (пикового потенциала) идет лавинообразное поступление ионов натрия внутрь клетки, в результате чего происходит перезарядка мембраны, и она утрачивает способность отвечать возбуждением на раздражители сверхпороговой силы. Эта фаза возбудимости получила название абсолютной рефрактерности, т.е. абсолютной невозбудимости, которая длится до конца перезарядки мембраны. Абсолютная рефрактерность мембраны возникает в связи с тем, что натриевые каналы полностью открываются, а затем инактивируются.
После окончания фазы перезарядки возбудимость ее постепенно восстанавливается до исходного уровня — это фаза относительной рефрактерности, т.е. относительной невозбудимости. Она продолжается до восстановления заряда мембраны до величины, соответствующей критическому уровню деполяризации. Поскольку в этот период мембранный потенциал покоя еще не восстановлен, то возбудимость ткани понижена, и новое возбуждение может возникнуть только при действии сверхпорогового раздражителя. Снижение возбудимости в фазу относительной рефрактерности связано с частичной инактивацией натриевых каналов и активацией калиевых каналов.
Следующему периоду соответствует повышенный уровень возбудимости: фаза вторичной экзальтации или вторичной супернормальной возбудимости. Так как мембранный потенциал в эту фазу ближе к критическому уровню деполяризации, по сравнению с состоянием покоя исходной поляризации, то порог раздражения снижен, т.е. возбудимость клетки повышена. В эту фазу новое возбуждение может возникнуть при действии раздражителей подпороговой силы. Натриевые каналы в эту фазу инактивированы не полностью. Мембранный потенциал увеличивается — возникает состояние гиперполяризации мембраны. Удаляясь от критического уровня деполяризации, порог раздражения слегка повышается, и новое возбуждение может возникнуть только при действии раздражителей сверхпороговой величины.
Механизм возникновения мембранного потенциала покоя
Каждая клетка в состоянии покоя характеризуется наличием трансмембранной разности потенциалов (потенциала покоя). Обычно разность зарядов между внутренней и внешней поверхностями мембран составляет от -80 до -100 мВ и может быть измерена с помощью наружного и внутриклеточного микроэлектродов (рис. 1).
Разность потенциалов между наружной и внутренней сторонами мембраны клетки в состоянии ее покоя называют мембранным потенциалом (потенциалом покоя).
Создание потенциала покоя обеспечивается двумя основными процессами — неравномерным распределением неорганических ионов между внутри- и внеклеточным пространством и неодинаковой проницаемостью для них клеточной мембраны. Анализ химического состав вне- и внутриклеточной жидкости свидетельствует о крайне неравномерном распределении ионов (табл. 1).
В состоянии покоя внутри клетки много анионов органических кислот и ионов К+, концентрация которых в 30 раз больше, чем снаружи; ионов Na+, наоборот, снаружи клетки в 10 раз больше, чем внутри; СI- также больше снаружи.
В покое мембрана нервных клеток наиболее проницаема для К+, менее — для СI- и очень мало проницаема для Na+/ Проницаемость мембраны нервного волокна для Na+ B покое в 100 раз меньше, чем для K+. Для многих анионов органических кислот мембрана в покое совсем непроницаема.
Рис. 1. Измерение потенциала покоя мышечного волокна (А) с помощью внутриклеточного микроэлектрода: М — микрозлектрод; И — индифферентный электрод. Луч на экране осциллографа (В) показывает, что до прокола мембраны микроэлектродом разность потенциалов между М и И была равна нулю. В момент прокола (показан стрелкой) обнаружена разность потенциалов, указывающая, что внутренняя сторона мембраны заряжена отрицательно по отношению к ее наружной поверхности (по Б.И. Ходорову)
Таблица. Внутри- и внеклеточные концентрации ионов мышечной клетки теплокровного животного, ммоль/л (по Дж. Дудел)
|
Внутриклеточная концентрация |
Внеклеточная концентрация |
|
|
А- (анионы органических соединений) |
В силу градиента концентраций К+ выходит на наружную поверхность клетки, вынося свой положительный заряд. Высокомолекулярные анионы не могут следовать за К+ из-за непроницаемости для них мембраны. Ион Na+ также не может возместить ушедшие ионы калия, ибо проницаемость мембраны для него значительно меньше. СI- по градиенту концентраций может перемешаться только внутрь клетки, увеличивая тем самым отрицательный заряд внутренней поверхности мембраны. Вследствие такого перемещения ионов возникает поляризация мембраны, когда наружная ее поверхность заряжается положительно, а внутренняя — отрицательно.
Электрическое поле, которое создастся на мембране, активно вмешивается в распределение ионов между внутренним и наружным содержимым клетки. По мере возрастания положительного заряда на наружной поверхности клетки иону К+ как положительно заряженному становится все труднее перемещаться изнутри наружу. Он движется как бы в гору. Чем больше величина положительного заряда на наружной поверхности, тем меньшее количество ионов К+ может выходить на поверхность клетки. При определенной величине потенциала на мембране количество ионов К+, пересекающих мембрану в том и другом направлении, оказывается равным, т.е. концентрационный градиент калия уравновешивается имеющимся на мембране потенциалом. Потенциал, при котором диффузионный поток ионов становится равным потоку одноименных ионов, идущих в обратном направлении, называют потенциалом равновесия для данного иона. Для ионов К+ потенциал равновесия равен -90 мВ. В миелинизированных нервных волокнах величина потенциала равновесия для ионов СI- близка к значению мембранного потенциала покоя (-70 мВ). Поэтому, несмотря на то что концентрация ионов СI- снаружи волокна больше, чем внутри его, не отмечается их одностороннего тока в соответствии с градиентом концентраций. В этом случае разность концентраций сбалансирована потенциалом, имеющимся на мембране.
Ион Na+ по градиенту концентраций должен был бы входить внутрь клетки (его потенциал равновесия составляет +60 мВ), и наличие отрицательного заряда внутри клетки не должно было бы препятствовать этому потоку. В этом случае входящий Na+ нейтрализовал бы отрицательные заряды внутри клетки. Однако этого в действительности не происходит, так как мембрана в покое малопроницаема для Na+.
Важнейшим механизмом, поддерживающим низкую внутриклеточную концентрацию ионов Na+ и высокую концентрацию ионов К+, является натрий-калиевый насос (активный транспорт). Известно, что в клеточной мембране имеется система переносчиков, каждый из которых связывается стремя находящимися внутри клетки ионами Na+ и выводит их наружу. С наружной стороны переносчик связывается с двумя находящимися вне клетки ионами К+ которые переносятся в цитоплазму. Энергообеспечение работы систем переносчиков обеспечивается АТФ. Функционирование насоса по такой системе приводит к следующим результатам:
- поддерживается высокая концентрация ионов К+ внутри клетки, что обеспечивает постоянство величины потенциала покоя. Вследствие того что за один цикл обмена ионов из клетки выводится на один положительный ион больше, чем вводится, активный транспорт играет роль в создании потенциала покоя. В этом случае говорят об электрогенном насосе, поскольку он сам создает небольшой, но постоянный ток положительных зарядов из клетки, а потому вносит прямой вклад в формирование отрицательного потенциала внутри нее. Однако величина вклада электрогенного насоса в общее значение потенциала покоя обычно невелика и составляет несколько милливольт;
- поддерживается низкая концентрация ионов Na + внутри клетки, что, с одной стороны, обеспечивает работу механизма генерации потенциала действия, с другой — обеспечивает сохранение нормальных осмолярности и объема клетки;
- поддерживая стабильный концентрационный градиент Na + , натрий-калиевый насос способствует сопряженному К+, Na+ -транспорту аминокислот и Сахаров через клеточную мембрану.
Таким образом, возникновение трансмембранной разности потенциалов (потенциала покоя) обусловлено высокой проводимостью клеточной мембраны в состоянии покоя для ионов К + , СI-, ионной асимметрией концентраций ионов К + и ионов СI-, работой систем активного транспорта (Na+/K+ -АТФаза), которые создают и поддерживают ионную асимметрию.
Потенциал действия нервного волокна, нервный импульс
Потенциал действия - это кратковременное колебание разности потенциалов мембраны возбудимой клетки, сопровождающееся изменением ее знака заряда.
Потенциал действия является основным специфическим признаком возбуждения. Его регистрация свидетельствует о том, что клетка или ее структуры ответили на воздействие возбуждением. Однако, как уже отмечалось, ПД в некоторых клетках может возникать спонтанно (самопроизвольно). Такие клетки содержатся в водителях ритма сердца, стенках сосудов, нервной системе. ПД используется как носитель информации, передающий ее в виде электрических сигналов (электрическая сигнализации) по афферентным и эфферентным нервным волокнам, проводящей системе сердца, а также для инициирования сокращения мышечных клеток.
Рассмотрим причины и механизм генерации ПД в афферентных нервных волокнах, образующих первично воспринимающие сенсорные рецепторы. Непосредственной причиной возникновения (генерации) ПД в них является рецепторный потенциал.
Если измерять разность потенциалов на мембране ближайшего к нервному окончанию перехвата Ранвье, то в промежутках между воздействиями на капсулу тельца Пачини она остается неизменной (70 мВ), а во время воздействия деполяризуется почти одновременно с деполяризацией рецепторной мембраны нервного окончания.
При увеличении силы давления на тельце Пачини, вызывающей возрастание рецепторного потенциала до 10 мВ, в ближайшем перехвате Ранвье обычно регистрируется быстрое колебание мембранного потенциала, сопровождающееся перезарядкой мембраны — потенциал действия (ПД), или нервный импульс (рис. 2). Если сила давления на тельце возрастет еще больше, амплитуда рецепторного потенциала увеличивается и в нервном окончании генерируется уже ряд потенциалов действия с определенной частотой.

Рис. 2. Схематическое представление механизма преобразования рецепторного потенциала в потенциал действия (нервный импульс) и распространения импульса по нервному волокну
Суть механизма генерации ПД состоит в том, что рецепторный потенциал вызывает возникновение локальных круговых токов между деполяризованной рецепторной мембраной немиелинизированной части нервного окончания и мембраной первого перехвата Ранвье. Эти токи, носителями которых являются ионы Na+, К+, СI- и другие минеральные ионы, «протекают» не только вдоль, но и поперек мембраны нервного волокна в области перехвата Ранвье. В мембране перехватов Ранвье в отличие от рецепторной мембраны самого нервного окончания имеется большая плотность ионных потенциалзависимых натриевых и калиевых каналов.
При достижении на мембране перехвата Ранвье величины деполяризации около 10 мВ происходит открытие быстрых потенциалзависимых натриевых каналов и через них в аксоплазму по электрохимическому градиенту устремляется поток ионов Na+. Он обусловливает быструю деполяризацию и перезарядку мембраны перехвата Ранвье. Однако одновременно с открытием быстрых потенциалзависимых натриевых каналов в мембране перехвата Ранвье открываются медленные потенциалзависимые калиевые каналы и из аксоилазмы начинают выходить ионы К+ Их выход запаздывает по отношению ко входу ионов Na+. Таким образом, входящие с большой скоростью в аксоплазму ионы Na+ быстро деполяризуют и перезаряжают на короткое время (0,3-0,5 мс) мембрану, а выходящие ионы К+ восстанавливают исходное распределение зарядов на мембране (реполяризуют мембрану). В результате во время механического воздействия на тельце Пачини силой, равной или превышающей пороговую, на мембране ближайшего перехвата Ранвье наблюдается кратковременное колебание потенциала в виде быстрой деполяризации и реполяризации мембраны, т.е. генерируется ПД (нервный импульс).
Поскольку непосредственной причиной генерации ПД является рецепторный потенциал, то его в этом случае еще называют генераторным потенциалом. Число генерируемых в единицу времени одинаковых по амплитуде и длительности нервных импульсов пропорционально амплитуде рецепторного потенциала, а следовательно, силе давления на рецептор. Процесс преобразования информации о силе воздействия, заложенной в амплитуде рецепторного потенциала, в число дискретных нервных импульсов получил название дискретного кодирования информации.
Более подробно ионные механизмы и временная динамика процессов генерации ПД изучены в экспериментальных условиях при искусственном воздействии на нервное волокно электрическим током различной силы и длительности.
Природа потенциала действия нервного волокна (нервного импульса)
Мембрана нервного волокна в точке локализации раздражающего электрода отвечает на воздействие очень слабого тока, еще не достигшего порогового значения. Этот ответ получил название локального, а колебание разности потенциалов на мембране — локального потенциала.
Локальный ответ на мембране возбудимой клетки может предшествовать возникновению потенциала действия или возникать как самостоятельный процесс. Он представляет собой кратковременное колебание (деполяризация и реполяризация) потенциала покоя, не сопровождающееся перезарядкой мембраны. Деполяризация мембраны при развитиии локального потенциала обусловлена опережающим входом в аксоплазму ионов Na+, а реполяризация — запаздывающим выходом из аксоплазмы ионов К+.
Если воздействовать на мембрану электрическим током возрастающей силы, то при се величине, называемой пороговой, деполяризация мембраны может достигнуть критического уровня — Е к, при котором происходит открытие быстрых потенциалзависимых натриевых каналов. В результате через них происходит лавинообразно нарастающее поступление в клетку ионов Na+. Вызываемый процесс деполяризации приобретает самоускоряющийся характер, и локальный потенциал перерастает в потенциал действия.
Уже упоминалось, что характерным признаком ПД является кратковременная инверсия (перемена) знака заряда на мембране. Снаружи она на короткое время (0,3-2 мс) становится заряженной отрицательно, а внутри — положительно. Величина инверсии может составлять до 30 мВ, а величина всего потенциала действия — 60-130 мВ (рис. 3).
Таблица. Сравнительная характеристика локального потенциала и потенциала действия
|
Характеристика |
Локальный потенциал |
Потенциал действия |
|
Проводимость |
Распространяется местно, на 1-2 мм с затуханием (декрементом) |
Распространяется без затухания на большие расстояния по всей длине нервного волокна |
|
Закон «силы» |
Подчиняется |
Не подчиняется |
|
Закон «все или ничего» |
Не подчиняется |
Подчиняется |
|
Явление суммации |
Суммируется, возрастает при повторных частых подпороговых раздражениях |
Не суммируется |
|
Величина амплитуды |
||
|
Способность к возбудимости |
Увеличивается |
Уменьшается вплоть до полной невозбудимости (рефрактерность) |
|
Величина раздражителя |
Подпороговая |
Пороговая и сверхпороговая |
Потенциал действия в зависимости от характера изменения зарядов на внутренней поверхности мембраны подразделяют на фазы деполяризации, реполяризации и гиперполяризации мембраны. Деполяризацией называют всю восходящую часть ПД, на которой выделяют участки, соответствующие локальному потенциалу (от уровня Е 0 до Е к ), быстрой деполяризации (от уровня Е к до уровня 0 мВ), инверсии знака заряда (от 0 мВ до пикового значения или начала реполяризации). Реполяризацией называют нисходящую часть ПД, которая отражает процесс восстановления исходной поляризации мембраны. Вначале реполяризация осуществляется быстро, но, приближаясь к уровню Е 0 , скорость се может замедляться и этот участок называют следовой отрицательностью (или следовым отрицательным потенциалом). У некоторых клеток вслед за реполяризацией развивается гиперполяризация (возрастание поляризации мембраны). Ее называют следовым положительным потенциалом.
Начальную высокоамплитудную быстропротекающую часть ПД называют также пик, или спайк. Он включает фазы деполяризации и быстрой реполяризации.
В механизме развития ПД важнейшая роль принадлежит потенциалзависимым ионным каналам и неодновременному увеличению проницаемости клеточной мембраны для ионов Na+ и К+. Так, при действии на клетку электрического тока он вызывает деполяризацию мембраны и, когда заряд мембраны уменьшается до критического уровня (Е к), открываются потенциалзависимые натриевые каналы. Как уже упоминалось,эти каналы образованы встроенными в мембрану белковыми молекулами, внутри которых имеются пора и два воротных механизма. Один из воротных механизмов — активационный обеспечивает (при участии сегмента 4) открытие (активацию) канала при деполяризации мембраны, а второй (при участии внутриклеточной петли между 3-м и 4-м доменами) — его инактивацию, развивающуюся при перезарядке мембраны (рис. 4). Поскольку оба этих механизма быстро изменяют положение ворот канала, то потенциалзависимые натриевые каналы являются быстрыми ионными каналами. Это обстоятельство имеет определяющее значение для генерации ПД в возбудимых тканях и для его проведения по мембранам нервных и мышечных волокон.

Рис. 3. Потенциал действия, его фазы и ионные токи (а, о). Описание в тексте

Рис. 4. Положение ворот и состояние активности потенциалзависимых натриевого и калиевого каналов при различных уровнях поляризации мембраны
Чтобы потенциалзависимый натриевый канал мог пропускать внутрь клетки ионы Na+, необходимо открыть лишь активационные ворота, поскольку инактивационные в условиях покоя открыты. Это и происходит, когда деполяризация мембраны достигает уровня Е к (рис. 3, 4).
Открытие активационных ворот натриевых каналов приводит к лавинообразному вхождению натрия внутрь клетки, движимому действием сил его электрохимического градиента. Поскольку ионы Na+ несут положительный заряд, то они нейтрализуют избыток отрицательных зарядов на внутренней поверхности мембраны, снижают разность потенциалов на мембране и деполяризуют ее. Вскоре ионы Na+ придают внутренней поверхности мембраны избыток положительных зарядов, что сопровождается инверсией (сменой) знака заряда с отрицательного на положительный.
Однако натриевые каналы остаются открытыми лишь около 0,5 мс и через этот промежуток времени от момента начала
ПД закрываются инактивационные ворота, натриевые каналы становятся инактивированными и непроницаемыми для ионов Na+, поступление которых внутрь клетки резко ограничивается.
С момента деполяризации мембраны до уровня Е к наблюдаются также активация калиевых каналов и открытие их ворот для ионов К+. Ионы К+ под действием сил концентрационного градиента выходят из клетки, вынося из нее положительные заряды. Однако воротный механизм калиевых каналов является медленно функционирующим и скорость выхода положительных зарядов с ионами К+ из клетки наружу запаздывает по отношению ко входу ионов Na+. Поток ионов К+, удаляя из клетки избыток положительных зарядов, обусловливает восстановление на мембране исходного распределения зарядов или ее реполяризацию, и на се внутренней стороне через мгновение от момента перезарядки восстанавливается отрицательный заряд.
Возникновение ПД на возбудимых мембранах и последующее восстановление исходного потенциала покоя на мембране оказываются возможными потому, что динамика входа в клетку и выхода из клетки положительных зарядов ионов Na+ и К+ различна. Вход иона Na+ по времени опережает выход иона К+. Если бы эти процессы были равновесными, то разность потенциалов на мембране не изменялась бы. Развитие способности к возбуждению и генерации ПД возбудимыми мышечными и нервными клетками было обусловлено формированием в их мембране двух типов разноскоростных ионных каналов — быстрых натриевых и медленных калиевых.
Для генерации одиночного ПД требуется поступление в клетку относительно небольшого числа ионов Na+, которое не нарушает его распределения вне и внутри клетки. При генерации большого числа ПД распределение ионов по обе стороны мембраны клетки могло бы нарушиться. Однако в нормальных условиях это предотвращается работой Na+, К+ -насоса.
В естественных условиях в нейронах ЦНС потенциал действия первично возникает в области аксонного холмика, в афферентных нейронах — в ближайшем к сенсорному рецептору перехвате Ранвье нервного окончания, т.е. в тех участках мембраны, где имеются быстрые селективные потенциалзависимые натриевые каналы и медленные калиевые каналы. В других типах клеток (например, пейсмекерных, гладких миоцитах) в возникновении ПД играют роль не только натриевые и калиевые, но и кальциевые каналы.
Механизмы восприятия и преобразования в ПД сигналов во вторично чувствующих сенсорных рецепторах отличаются от механизмов, разобранных для первично чувствствующих рецепторов. В этих рецепторах восприятие сигналов осуществляется специализированными нейросенсорными (фоторецепторные, обонятельные) или сенсоэпителиальными (вкусовые, слуховые, вестибулярные) клетками. В каждой из этих чувствительных клеток имеется свой, особый механизм восприятия сигналов. Однако во всех клетках энергия воспринимаемого сигнала (раздражителя) преобразуется в колебание разности потенциалов плазматической мембраны, т.е. в рецепторный потенциал.
Таким образом, ключевым моментом в механизмах преобразования сенсорными клетками воспринимаемых сигналов в рецепторный потенциал является изменение проницаемости ионных каналов в ответ на воздействие. Открытие Na+, Са 2+ , К+ -ионных каналов при восприятии и преобразовании сигнала достигается в этих клетках при участии G-белков, вторых внутриклеточных посредников, связывании с лигандами, фосфорилировании ионных каналов. Как правило, возникший в сенсорных клетках рецепторный потенциал вызывает высвобождение из них в синаптическую щель нейромедиатора, который обеспечивает передачу сигнала на постсинаптическую мембрану афферентного нервного окончания и генерацию на его мембране нервного импульса. Эти процессы подробно описаны в главе, посвященной сенсорным системам.
Потенциал действия может быть охарактеризован амплитудой и продолжительностью, которые для одного и того же нервного волокна остаются одинаковыми при распространении ПД по волокну. Поэтому потенциал действия называют дискретным потенциалом.
Между характером воздействия на сенсорные рецепторы и числом ПД, возникших в афферентном нервном волокне в ответ на воздействие, имеется определенная связь. Она заключается в том, что на большие но силе или продолжительности воздействия в нервном волокне формируется большее число нервных импульсов, т.е. при усилении воздействия в нервную систему будут посылаться от рецептора импульсы большей частоты. Процессы преобразования информации о характере воздействия в частоту и другие параметры нервных импульсов, передаваемых в ЦНС, получили название дискретного кодирования информации.
Для объяснения происхождения потенциала покоя были предложены различные теории. У истоков современного понимания этой проблемы стоит работа В. Ю. Чаговца, который в 1896 г., будучи студентом-медиком, высказал мысль об ионной природе биоэлектрических процессов и сделал попытку применить теорию электролитической диссоциации Аррениуса для объяснения происхождения этих потенциалов. В дальнейшем в 1902 г. Ю. Бернштейном была развита мембранно-ионная теория, которая модифицирована и экспериментально обоснована А. Ходжкином и А. Хаксли (1952) и в настоящее время пользуется широким признанием. Согласно этой теории, биоэлектрические потенциалы обусловлены неодинаковой концентрацией ионов Кֹ, Naֹ, Сl" внутри и вне клетки и различной проницаемостью для них поверхностной мембраны.
Протоплазма нервных и мышечных клеток содержит в 30-50 раз больше ионов калия, в 8-10 раз меньше ионов натрия и в 50 раз меньше ионов хлора, чём внеклеточная жидкость.
Препятствием для быстрого выравнивания этой разности концентраций является тончайшая (около 100 Å) плазматическая мембрана, покрывающая живые клетки.
|
Представления о структуре этой мембраны строятся на основании данных, полученных методами электронной микроскопии, оптической микроскопии, диффракции рентгеновых лучей и химического анализа. Предполагают, что мембрана состоит из двойного слоя молекул фосфолипидов, покрытого изнутри слоем белковых молекул, а снаружи слоем молекул сложных углеводов - мукополисахаридов. Трехслойная структура мембраны схематически изображена на рис. 116 . Рис. 116. Схема молекулярной структуры мембраны. Показан бимолекулярный липидныи слой Z (кружками обозначены полярные группы фосфолипидов) и два нелипидных монослоя: наружный - мукополисахаридный - Х, внутренний - белковый - Y (по Робертсону). |
В клеточной мембране имеются тончайшие канальцы - «поры» диаметром в несколько ангстрем. Через эти канальцы молекулы воды и других веществ, а такжо ионы, имеющие соответствующий размеру пор диаметр, входят в клетку выходят из нее.
На структурных элементах мембраны фиксируются личные ионы, что придает стенкам ее пор тот или иной заряд и тем самым затрудняет или облегчает прохождение через них ионов. Так, предполагается, что наличие в мембране диссоциированных фосфатных и карбоксильных групп является причиной того, что мембрана нервных волокон значительно менее проницаема для анионов, чем для катионов.
Проницаемость мембраны для различных катионов также неодинакова, и она закономерно изменяется при разных функциональных состояниях ткани. В покое мембрана нервных волокон примерно в 20-100 раз более проницаема для ионов Кֹ, чем для ионов Naֹ, а при возбуждении натриевая проницаемость начинает значительно превышать калиевую проницаемость мембраны.
Для того чтобы понять механизм возникновения мембранного потенциала покоя с точки зрения теории Бернштейна - Ходжкина, рассмотрим модельный опыт. Первую половину сосуда (рис. 117 ), разделенного искусственной полупроницаемой мембраной, поры которой свободно пропускают положительно заряженные ионы Кֹ и не пропускают отрицательно заряженных ионов SO"4, наполняют концентрированным раствором K2SО4, а левую половину наполняют также раствором K2SО4, но меньшей концентрации.
 |
Вследствие существования концентрационного градиента ионы Кֹ начнут диффундировать через мембрану преимущественно из правой половины сосуда (где их концентрация равна С1) в левую (с концентрацией С2). Соответственно отрицательно заряженные анионы SO"4, для которых мембрана непроницаема, будут концентрироваться в правой половине сосуда у поверхности мембраны. Своим отрицательным зарядом они электростатически будут удерживать ионы Кֹ на поверхности мембраны слева. В результате мембрана поляризуется: между двумя ее поверхностями возникает разность потенциалов. Рис. 117. Возникновение разности потенциалов на искусственной мембране, разделяющей растворы K2SО4 разной концентрации (С1 и С2). Мембрана избирательно проницаема для катионов Кֹ (маленькие кружки) и не пропускает анионы SО"4 (большие кружки). 1 и 2 - электроды, опущенные в раствор; 3 - электроизмерительный прибор. |
Если теперь в правую и левую половины сосуда опустить электроды, то электроизмерительный прибор обнаружит наличие разности потенциалов, при этом раствор с меньшей концентрацией ионов K2SО4, в который происходит преимущественно диффузия положительно заряженных ионов Кֹ, приобретает положительный заряд по отношению к раствору с большей концентрацией K2SО4.
Разность потенциалов (Е) в рассмотренном случае может быть подсчитана по формуле Нернста:

Есть много оснований считать, что сходные отношения имеют место и в живом нервном волокне, поскольку концентрация ионов Кֹ в протоплазме более, чем в 30 роз превышает концентрацию этих ионов в наружном растворе, а органические (белковые и др.) анионы протоплазмы через мембрану практически не проникают.
В состоянии физиологического покоя диффузия положительно заряженных ионов Кֹ из протоплазмы во внешнюю жидкость придает наружной поверхности мембраны положительный заряд, а внутренней - отрицательный.
Важным доводом в пользу правильности этого представления послужил тот факт, что рассчитанная по формуле Нернста разность потенциалов между наружной и внутренней сторона мембраны мышечного волокна (около 90 мв) оказалась близкой к измеренной в опытах с помощью внутриклеточного микроэлектрода.
Было установлено также, что повышение концентрации ионов Кֹ во внешней среде клетки, а следовательно, cнижение разности концентрации этих ионов по обе стороны мембраны приводят к падению потенциала покоя, причем в определенном диапазоне концентрацией эти сдвиги количественно хорошо совпадают с расчитанными по формуле Нернста.
Однако наиболее важные, прямые, доказательства правильности этих представлений были получены А. Ходжкиным с сотрудниками (1962) в опытах с заменой протоплазмы в гигантских нервных волокнах моллюска кальмара солевыми растворами. Из волокна, имеющего диаметр около 1 мл, осторожно выдавливали протоплазму и спавшуюся оболочку заполняли искусственным солевым раствором.
В том случае, когда концентрация ионов калия в этом растворе была близка к внутриклеточной, между внутренней и наружной стороной мембраны устанавливалась разность потенциалов, примерно равная потенциалу покоя нормального волокна (50-80 мв). Уменьшение концентрации ионов Кֹ во внутреннем растворе приводило к закономерному снижению или даже извращению потенциала покоя.
Такие опыты показали, что концентрационный градиент ионов Кֹ действительно является основным фактором, определяющим величину потенциала покоя нервного волокна.
Наряду с ионами Кֹ в возникновении потенциала покоя принимают участие и ионы Naֹ, диффундирующие в протоплазму из внеклеточной жидкости, где их концентрация велика. Диффузия эта сильно затруднена низкой натриевой проницаемостью мембраны в покое. Тем не менее, диффундируя через мембрану внутрь протоплазмы, ионы Naֺ переносят сюда свои положительные заряды, что несколько уменьшает величину потенциала покоя, создаваемого диффузией из клетки ионов Кֺ. Этим объясняется тот факт, что потенциал покоя большинства нервных клеток и волокон имеет величину не 90 мв, как это следовало ожидать, если бы эхтот потенциал создавался только ионами Кֺ но 60-70 мв.
Таким образом, величина потенциала покоя нервных волокон и клеток определяется соотношением числа положительно заряженных ионов Кֺ, диффундирующих в единицу времени из клетки наружу, и положительно заряженных ионов Naֺ, диффундирующих через мембрану в противоположном направлении. Чем это соотношение выше, тем больше величина потенциала покоя, и наоборот.
text_fields
text_fields
arrow_upward
Мембранным потенциалом покоя (МПП) или потенциалом покоя (ПП) называют разность потенциалов покоящейся клетки между внутренней и наружной сторонами мембраны. Внутренняя сторона мембраны клетки заряжена отрицательно по отношению к наружной. Принимая потенциал наружного раствора за нуль, МПП записывают со знаком «минус». Величина МПП зависит от вида ткани и варьирует от -9 до -100 мв. Следовательно, в состоянии покоя клеточная мембрана поляризована. Уменьшение величины МПП называют деполяризацией, увеличение - гиперполяризацией, восстановление исходного значения МПП - ре поляризацией мембраны.
Основные положения мембранной теории происхождения МПП сводятся к следующему. В состоянии покоя клеточная мембрана хорошо проницаема для ионов К + (в ряде клеток и для СГ), менее проницаема для Na + и практически непроницаема для внутриклеточных белков и других органических ионов. Ионы К + диффундируют из клетки по концентрационному градиенту, а непроникающие анионы остаются в цитоплазме, обеспечивая появление разности потенциалов через мембрану.
Возникающая разность потенциалов препятствует выходу К + из клетки и при некотором ее значении наступает равновесие между выходом К + по концентрационному градиенту и входом этих катионов по возникшему электрическому градиенту. Мембранный потенциал, при котором достигается это равновесие, называется равновесным потенци алом. Его величина может быть рассчитана из уравнения Нернста:
где Е к - равновесный потенциал для К + ; R - газовая постоянная; Т - абсолютная температура; F - число Фарадея; п - валентность К + (+1), [К н + ] - [К + вн ] - наружная и внутренняя концентрации К + —
Если перейти от натуральных логарифмов к десятичным и подставить в уравнение числовые значения констант, то уравнение примет вид:
В спинальных нейронах (табл. 1.1) Е к = -90 мв. Величина МПП, измеренная с помощью микроэлектродов заметно ниже — 70 мв.
Таблица 1.1 . Концентрация некоторых ионов внутри и снаружи спинальных мотонейронов млекопитающих
| Ион |
Концентрация |
(ммоль/л Н 2 О) |
Разновесный потенциал (мв) |
|
внутри клетки |
снаружи клетки |
||
| Na + | 15,0 | 150,0 | |
| К + | 150,0 | 5,5 | |
| Сl — | 125,0 | ||
|
Мембранный потенциал покоя = -70 мв |
|||
Если мембранный потенциал клетки имеет калиевую природу, то, в соответствии с уравнением Нернста, его величина должна линейно снижаться с уменьшением концентрационного градиента этих ионов, например, при повышении концентрации К + во внеклеточной жидкости. Однако линейная зависимость величины МПП (Мембранный потенциал покоя) от градиента концентрации К + существует только при концентрации К + во внеклеточной жидкости выше 20 мМ. При меньших концентрациях К + снаружи клетки кривая зависимости Е м от логарифма отношения концентрации калия снаружи и внутри клетки отличается от теоретической. Объяснить установленные отклонения экспериментальной зависимости величины МПП и градиента концентрации К + теоретически рассчитанной по уравнению Нернста можно, допустив, что МПП возбудимых клеток определяется не только калиевым, но и натриевым, и хлорным равновесным потенциалами. Рассуждая аналогично с предыдущим, можно записать:
Величины натриевого и хлорного равновесных потенциалов для спинальных нейронов (табл. 1.1) равны соответственно +60 и -70 мв. Значение Е Cl равно величине МПП. Это свидетельствует о пассивном распределении ионов хлора через мембрану в соответстии с химическим и электрическим градиентами. Для ионов натрия химический и электрический градиенты направлены внутрь клетки.
Вклад каждого из равновесных потенциалов в величину МПП определяется соотношением между проницаемостью клеточной мембраны для каждого из этих ионов. Расчет величины мембранного потенциала производится с помощью уравнения Гольдмана:

Е m - мембранный потенциал; R - газовая постоянная; Т - абсолютная температура; F - число Фарадея; Р K , P Na и Р Cl - константы проницаемости мембраны для К + Na + и Сl, соответственно; [К + н ], [ K + вн , [ Na + н [ Na + вн ], [Сl — н ] и[Сl — вн ]- концентрации K + , Na + и Сl снаружи (н) и внутри (вн) клетки.
Подставляя в это уравнение полученные в экспериментальных исследованиях концентрации ионов и величину МПП, можно показать, что для гигантского аксона кальмара должно быть следующее соотношение констант проницаемости Р к: P Na: Р С1 = I: 0,04: 0,45. Очевидно, что, поскольку мембрана проницаема для ионов натрия (Р N a =/ 0) и равновесный потенциал для этих ионов имеет знак «плюс», то вход последних внутрь клетки по химическому и электрическому градиентам будет уменьшать электроотрицательность цитоплазмы, т.е. увеличивать МПП (Мембранный потенциал покоя).
При повышении концентрации ионов калия в наружном растворе выше 15 мМ МПП увеличивается и соотношение констант проницаемости меняется в сторону более значительного превышения» Р к над P Na и Р С1 . Р к: P Na: Р С1 = 1: 0.025: 0,4. В таких условиях МПП определяется почти исключительно градиентом ионов калия, поэтому экспериментальная и теоретическая зависимости величины МПП от логарифма отношения концентраций калия снаружи и внутри клетки начинают совпадать.
Таким образом, наличие стационарной разности потенциалов между цитоплазмой и наружной средой в покоящейся клетке обусловлено существующими концентрационными градиентами для К + , Na + и Сl и различной проницаемостью мембраны для этих ионов. Основную роль в генерации МПП играет диффузия ионов калия из клетки в наружный наствор. Наряду с этим, МПП определяется также натриевым и хлорным равновесными потенциалами и вклад каждого из них определяется отношениями между проницаемостями плазматической мембраны клетки для данных ионов.
Все факторы, перечисленные выше, составляют так называемую ионную компоненту МПП (Мембранный потенциал покоя). Поскольку, ни калиевый, ни натриевый равновесные потенциалы не равны МПП. клетка должна поглощать Na + и терять К + . Постоянство концентраций этих ионов в клетке поддерживается за счет работы Na + К + -АТФазы.
Однако роль этого ионного насоса не ограничивается поддержанием градиентов натрия и калия. Известно, что натриевый насос электрогенен и при его функционировании возникает чистый поток положительных зарядов из клетки во внеклеточную жидкость, обуславливающий увеличение электроотрицательности цитоплазмы по отношению к среде. Электрогенность натриевого насоса была выявлена в опытах на гигантских нейронах моллюска. Электрофорети-ческая инъекция ионов Na + в тело одиночного нейрона вызывала гиперполяризацию мембраны, во время которой МПП был значительно ниже величины калиевого равновесного потенциала. Указанная гиперполяризация ослаблялась при снижении температур раствора, в котором находилась клетка, и подавлялась специфическим ингибитором Na + , К + -АТФазы уабаином.
Из сказанного следует, что МПП может быть разделен на две компоненты - «ионную» и «метаболическую». Первая компонента зависит от концентрационных градиентов ионов и мембранных проницаемостей для них. Вторая, «метаболическая», обусловлена активным транспортом натрия и калия и оказывает двоякое влияние на МПП. С одной стороны, натриевый насос поддерживает концентрационные градиенты между цитоплазмой и внешней средой. С другой, будучи электрогенным, натриевый насос оказывает прямое влияние на МПП. Вклад его в величину МПП зависит от плотности «насосного» тока (ток на единицу плошади поверхности мембраны клетки) и сопротивления мембраны.
Мембранный потенциал действия
text_fields
text_fields
arrow_upward
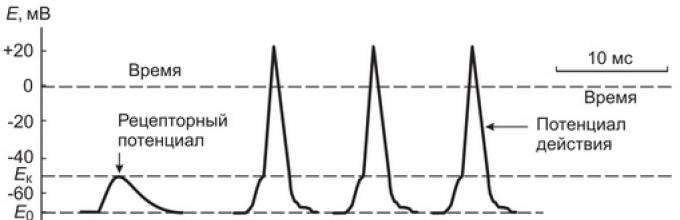
Если на нерв или мышцу нанести раздражение выше порога возбуждения, то МПП нерва или мышцы быстро уменьшится и на короткий промежуток времени (миллисекунда) произойдет перезарядка мембраны: ее внутренняя сторона станет заряженной положительно относительно наружной. Это кратковременное изменение МПП, происходящее при возбуждении клетки, которое на экране осциллографа имеет форму одиночного пика, называется мембранным потенциалом действия (МПД).
МПД в нервной и мышечной тканях возникает при снижении абсолютной величины МПП (деполяризации мембраны) до некоторого критического значения, называемого порогом генерации МПД. В гигантских нервных волокнах кальмара МПД равен — 60 мВ. При деполяризации мембраны до -45 мВ (порог генерации МПД) возникает МПД (рис. 1.15).
Рис. 1.15 Потенциал действия нервного волокна (А) и изменение проводимости мембраны для ионов натрия и калия (Б).Во время возникновения МПД в аксоне кальмара сопротивление мембраны уменьшается в 25 раз, с 1000 до 40 Ом.см 2 , тогда как электрическая емкость не изменяется. Указанное снижение сопротивления мембраны обусловлено увеличением ионной проницаемости мембраны при возбуждении.
По своей амплитуде (100-120 мВ) МПД (Мембранный потенциал действия) на 20-50 мВ превышает величину МПП (Мембранный потенциал покоя). Другими словами, внутренняя сторона мембраны на короткое время становится заряженной положительно по отношению к наружной, - «овершут» или реверсия заряда.
Из уравнения Гольдмана следует, что лишь увеличение проницаемости мембраны для ионов натрия может привести к таким изменениям мебранного потенциала. Значение Е к всегда меньше, чем величина МПП, поэтому повышение проницаемости мембраны для К + будет увеличивать абсолютное значение МПП. Натриевый равновесный потенциал имеет знак «плюс», поэтому резкое увеличение проницаемости мембраны для этих катионов приводит к перезарядке мембраны.
Во время МПД увеличивается проницаемость мембраны для ионов натрия. Расчеты показали, что если в состоянии покоя соотношение констант проницаемости мембраны для К + , Na + и СГ равно 1:0,04:0,45, то при МПД — Р к: P Na: Р = 1: 20: 0,45. Следовательно, в состоянии возбуждения мембрана нервного волокна не просто утрачивает свою избирательную ионную проницаемость, а, напротив, из избирательно проницаемой в покое для ионов калия она становится избирательно проницаемой для ионов натрия. Увеличение натриевой проницаемости мембраны связано с открыванием потенциал-зависимых натриевых каналов.
Механизм, который обеспечивает открывание и закрывание ионных каналов, получил название ворот канала. Принято различать активационные (m) и инактивационные (h) ворота. Ионный канал может находиться в трех основных состояниях: закрытом (m-ворота закрыты; h-открыты), открытом (m- и h-ворота открыты) и инактивированном (m-ворота открыты, h- ворота закрыты) (рис 1.16).
Рис. 1.16 Схема положения активационных (m) и инактивационных (h) ворот натриевых каналов, соответствующие закрытому (покой, А), открытому (активация, Б) и инактивированному (В) состояниям.
Деполяризация мембраны, вызываемая раздражающим стимулом, например, электрическим током, открывает m-ворота натриевых каналов (переход из состояния А в Б) и обеспечивает появление направленного внутрь потока положительных зарядов - ионов натрия. Это ведет к дальнейшей деполяризации мембраны, что, в свою очередь, увеличивает число открытых натриевых каналов и, следовательно, повышает натриевую проницаемость мембраны. Возникает «регенеративная» деполяризация мембраны, в результате которой потенциал внутренней стороны мембраны стремится достичь величины натриевого равновесного потенциала.
Причиной прекращения роста МПД (Мембранный потенциал действия) и реполяризации мембраны клетки является:
а) Увеличение деполяризации мембраны, т.е. когда Е м -» E Na , в результате чего снижается электрохимический градиент для ионов натрия, равный Е м -> E Na . Другими словами, уменьшается сила, «толкающая» натрий внутрь клетки;
б) Деполяризация мембраны порождает процесс инактивации натриевых каналов (закрывание h-ворот; состояние В канала), который тормозит рост натриевой проницаемости мембраны и ведет к ее снижению;
в) Деполяризация мембраны увеличивает ее проницаемость для ионов калия. Выходящий калиевый ток стремится сместить мембранный потенциал в сторону калиевого равновесного потенциала.
Снижение электрохимического потенциала для ионов натрия и инактивация натриевых каналов уменьшает величину входящего натриевого тока. В определенный момент времени величина входящего тока натрия сравнивается с возросшим выходящим током - рост МПД прекращается. Когда суммарный выходящий ток превышает входящий, начинается реполяризация мембраны, которая также имеет регенеративный характер. Начавшаяся реполяризация ведет к закрыванию активационных ворот (m), что уменьшает натриевую проницаемость мембраны, ускоряет реполяризацию, а последняя увеличивает число закрытых каналов и т.д.
Фаза реполяризации МПД в некоторых клетках (например, в кардиомиоцитах и ряде гладкомышечных клеток) может замедляться, формируя плато ПД, обусловленное сложными изменениями во времени входящих и выходящих токов через мембрану. В последействии МПД может возникнуть гиперполяризация или/и деполяризация мембраны. Это так называемые следовые потенциалы. Следовая гиперполяризация имеет двоякую природу: ионную и метаболичес кую. Первая связана с тем, что калиевая проницаемость в нервном волокне мембраны остается некоторое время (десятки и даже сотни миллисекунд) повышенной после генерации МПД и смещает мембранный потенциал в сторону калиевого равновесного потенциала. Следовая гиперполяризация после ритмической стимуляции клеток связана преимущественно с активацией электрогенного натриевого насоса, вследствие накопления ионов натрия в клетке.
Причиной деполяризации, развивающейся после генерации МПД (Мембранный потенциал действия), является накопление ионов калия у наружной поверхности мембраны. Последнее, как это следует из уравнения Гольдмана, ведет к увеличению МПП (Мембранный потенциал покоя).
С инактивацией натриевых каналов связано важное свойство нервного волокна, называемое рефрактерностью .
Во время абсо лютного рефрактерного периода нервное волокно полностью утрачивает способность возбуждаться при действии раздражителя любой силы.
Относительная рефрактерность , следующая за абсолютной, характеризуется более высоким порогом возникновения МПД (Мембранный потенциал действия).
Представление о мембранных процессах, происходящих во время возбуждения нервного волокна, служит базой для понимания и явления аккомодации. В основе аккомодации ткани при малой крутизне нарастания раздражающего тока лежит повышение порога возбуждения, опережающее медленную деполяризацию мембраны. Повышение порога возбуждения почти целиком определяется инактивацией натриевых каналов. Роль повышения калиевой проницаемости мембраны в развитии аккомодации состоит в том, что оно приводит к падению сопротивления мембраны. Вследствие снижения сопротивления скорость деполяризации мембраны становится еще медленнее. Скорость аккомодации тем выше, чем большее число натриевых каналов при потенциале покоя находится в инактивированном состоянии, чем выше скорость развития инактивации и чем выше калиевая проницаемость мембраны.
Проведение возбуждения
text_fields
text_fields
arrow_upward
Проведение возбуждения по нервному волокну осуществляется за счет локальных токов между возбужденным и покоящимися участками мембраны. Последовательность событий в этом случае представляется в следующем виде.
При нанесении точечного раздражения на нервное волокно в соответствующем участке мембраны возникает потенциал действия. Внутренняя сторона мембраны в данной точке оказывается заряженной положительно по отношению к соседней, покоящейся. Между точками волокна, имеющими различный потенциал, возникает ток (локальный ток), направленный от возбужденного (знак (+) на внутренней стороне мембраны) к невозбужденному (знак (-) на внутренней стороне мембраны) к участку волокна. Этот ток оказывает деполяризующее влияние на мембрану волокна в покоящемся участке и при достижении критического уровня деполяризации мембраны в данном участке возникает МПД (Мембранный потенциал действия). Этот процесс последовательно распространяется по всем участкам нервного волокна.
В некоторых клетках (нейронах, гладких мышцах) МПД имеет не натриевую природу, а обусловлен входом ионов Ca 2+ по потенциал-зависимым кальциевым каналам. В кардиомиоцитах генерация МПД связана с входящими натриевым и натрий-кальциевым токами.
В данной теме будут рассмотрены два катиона – натрий (Na) и калий (К). Говоря об анионах, примем во внимание то, что определенное количество анионов находится у наружной и внутренней стороны мембраны клетки.
Форма клетки зависит от того, к какой ткани она принадлежит. По своей форме клетки могут быть:
· цилиндрические и кубические (клетки кожи);
· дисковидные (эритроциты);
· шаровидные (яйцеклетки);
· веретеновидные (гладкомышечные);
· звездчатые и пирамидные (нервные клетки);
· не имеющие постоянной формы – амебовидные (лейкоциты).
Клетка обладает рядом свойств: она питается, растет, размножается, восстанавливается, адаптируется к окружающей её среде, обменивается энергией и веществами с окружающей средой, выполняет присущие ей функции (в зависимости от того, к какой ткани принадлежит данная клетка). Кроме того, клетка обладает возбудимостью.
Возбудимость – это способность клетки в ответ на раздражения переходить из состояния покоя к состоянию активности.
Раздражения могут поступать из внешней среды или возникать внутри клетки. Раздражителями, вызывающими возбуждение, могут быть: электрические, химические, механические, температурные и иные стимулы.
Клетка может находиться в двух основных состояниях – в покое и в возбуждении. Покой и возбуждение клетки иначе называют – мембранный потенциал покоя и мембранный потенциал действия.
Когда клетка не испытывает ни каких раздражений, она находится в состоянии покоя. Покой клетки иначе называется мембранным потенциалом покоя (МПП).
В состоянии покоя внутренняя поверхность ее мембраны заряжена отрицательно, а наружная – положительно. Это объясняется тем, что внутри клетки находится много анионов и мало катионов, а за клеткой наоборот, преобладают катионы.
Поскольку в клетке присутствуют электрические заряды, то создаваемое ими электричество можно измерить. Величина мембранного потенциала покоя равна: - 70 мВ, (минус 70, поскольку внутри клетки отрицательный заряд). Данная величина условна, так как в каждой клетке может быть своя величина потенциала покоя.
В состоянии покоя поры мембраны открыты для ионов калия и закрыты для ионов натрия. Это означает, что ионы калия могут легко проникать в клетку и выходить из нее. Ионы натрия не могут поступать в клетку, поскольку для них закрыты поры мембраны. Но небольшое число ионов натрия проникает в клетку, потому что притягиваются большим количеством анионов, расположенных на внутренней поверхности мембраны (разноименные заряды притягиваются). Такое перемещение ионов является пассивным, поскольку не требует затрат энергии.
Для нормальной жизнедеятельности клетки величина ее МПП должна оставаться на постоянном уровне. Однако, перемещение ионов натрия и калия через мембрану вызывает колебания данной величины, что может привести к уменьшению или увеличению значения: - 70 мВ.
Для того, чтобы величина МПП оставалась относительно постоянной начинает действовать так называемый натрий – калиевый насос. Его функция заключается в том, что он ионы натрия убирает из клетки, а ионы калия нагнетает в клетку. Именно определенное соотношение ионов натрия и калия в клетке и за клеткой создает нужную величину МПП. Работа насоса является активным механизмом, поскольку для нее требуется энергия.
Источником энергии в клетке является АТФ. АТФ дает энергию только при расщеплении на более простую кислоту - АДФ, при обязательном участии в реакции фермента АТФ-азы:
АТФ + фермент АТФ-аза АДФ + энергия
Все живые клетки обладают способностью под влиянием раздражителей переходить из состояния физиологического покоя в состояние активности или возбуждения.
Возбуждение - это комплекс активных электрических, химических и функциональных изменений в возбудимых тканях (нервной, мышечной или железистой), которыми ткань отвечает на внешнее воздействие. Важную роль при возбуждении играют электрические процессы, обеспечивающие проведение возбуждения по нервным волокнам и приводящие ткани в активное (рабочее) состояние.
Мембранный потенциал
Живые клетки обладают важным свойством: внутренняя поверхность клетки всегда заряжена отрицательно по отношению к внешней ее стороне. Между внешней поверхностью клетки, заряженной электроположительно по отношению к протоплазме, и внутренней стороной клеточной мембраны существует разность потенциалов, которая колеблется в пределах 60-70 мВ. По данным П. Г. Ко- стюка (2001), у нервной клетки эта разность колеблется в пределах от 30 до 70 мВ. Разность потенциалов между внешней и внутренней сторонами мембраны клетки называют мембранным потенциалом, или потенциалом покоя (рис. 2.1).
Мембранный потенциал покоя присутствует на мембране до тех пор, пока клетка жива, и исчезает с гибелью клетки. Л. Гальвани еще в 1794 г. показал, что если повредить нерв или мышцу, сделав поперечное сечение и приложив к поврежденной части и к месту повреждения электроды, соединенные с гальванометром, то гальванометр покажет ток, который течет всегда от неповрежденной части ткани к месту разреза. Это течение он назвал током покоя. По своей физиологической сути ток покоя и мембранный потенциал покоя - одно и то же. Измеренная в данном опыте разность потенциалов составляет 30-50 мВ, поскольку при повреждении ткани часть тока шунтируется в межклеточном пространстве и окружающей исследуемую структуру жидкости. Разность потенциалов можно рассчитать по формуле Нернста:
где R - газовая постоянная, Т - абсолютная температура, F - число Фарадея, [К] вн. и [К] нар. - концентрация калия внутри и снаружи клетки.

Рис. 2.1.
Причина возникновения потенциала покоя общая для всех клеток. Между протоплазмой клетки и внеклеточной средой существует неравномерное распределение ионов (ионная асимметрия). Состав крови человека по солевому балансу напоминает состав океанской воды. Внеклеточная среда в центральной нервной системе также содержит много хлорида натрия. Ионный состав цитоплазмы клеток беднее. Внутри клеток в 8-10 раз меньше ионов Na + и в 50 раз меньше ионовС!". Основным катионом цитоплазмы является К + . Его концентрация внутри клетки в 30 раз выше, чем во внеклеточной среде, и приблизительно равняется внеклеточной концентрации Na Основными противоионами для К + в цитоплазме являются органические анионы, в частности анионы аспарагиновой, гистаминовой и других аминокислот. Такая асимметрия - это нарушение термодинамического равновесия. Для того чтобы восстановить его, ионы калия должны постепенно покидать клетку, а ионы натрия - стремиться в нее. Однако этого не происходит.
Первым препятствием для выравнивания разности концентраций ионов является плазматическая мембрана клетки. Она состоит из двойного слоя молекул фосфолипидов, покрытых изнутри слоем белковых молекул, а снаружи - слоем углеводов (мукополисахари- дов). Некоторая часть клеточных белков встроена непосредственно в двойной липидный слой. Это внутренние белки.
Мембранные белки всех клеток делят на пять классов: насосы, каналы, рецепторы, ферменты и структурные белки. Насосы служат для перемещения ионов и молекул против градиентов концентрации, используя для этого метаболическую энергию. Белковые каналы, или поры, обеспечивают избирательную проницаемость (диффузию) через мембрану соответствующих им по размеру ионов и молекул. Рецепторные белки, обладающие высокой специфичностью, распознают и связывают, прикрепляя к мембране, многие типы молекул, необходимых для жизнедеятельности клетки в каждый определенный момент времени. Ферменты ускоряют течение химических реакций у поверхности мембраны. Структурные белки обеспечивают соединение клеток в органы и поддержание субклеточной структуры.
Все эти белки специфичны, но не строго. В определенных условиях тот или иной белок может быть одновременно и насосом, и ферментом, и рецептором. Через каналы мембраны молекулы воды, а также соответствующие размерам пор ионы входят в клетку и выходят из нее. Проницаемость мембраны для различных катионов не одинакова и изменяется при разных функциональных состояниях ткани. В покое мембрана в 25 раз более проницаема для ионов калия, чем для ионов натрия, а при возбуждении натриевая проницаемость примерно в 20 раз превышает калиевую. В состоянии покоя равные концентрации калия в цитоплазме и натрия во внеклеточной среде должны обеспечить и равное количество положительных зарядов по обе стороны мембраны. Но поскольку проницаемость для ионов калия выше в 25 раз, то калий, выходя из клетки, делает ее поверхность все более положительно заряженной по отношению к внутренней стороне мембраны, около которой все более накапливаются слишком крупные для пор мембраны отрицательно заряженные молекулы аспарагиновой, гистаминовой и других аминокислот, «отпустивших» калий за пределы клетки, но «не дающих» ему уйти далеко благодаря своему отрицательному заряду. С внутренней стороны мембраны копятся отрицательные заряды, а с внешней - положительные. Возникает разность потенциалов. Диффузный ток ионов натрия в протоплазму из внеклеточной жидкости удерживает эту разность на уровне 60-70 мВ, не давая ей возрастать. Диффузный ток ионов натрия в покое в 25 раз слабее, чем встречный ток ионов калия. Ионы натрия, проникая внутрь клетки, снижают величину потенциала покоя, позволяя ей удерживаться на определенном уровне. Таким образом, величина потенциала покоя мышечных и нервных клеток, а также нервных волокон определяется соотношением числа положительно заряженных ионов калия, диффундирующих в единицу времени из клетки наружу, и положительно заряженных ионов натрия, диффундирующих через мембрану в противоположном направлении. Чем это соотношение выше, тем больше величина потенциала покоя, и наоборот.
Вторым препятствием, удерживающим разность потенциалов на определенном уровне, является натрий-калиевый насос (рис. 2.2). Он получил название натрий-калиевого или ионного, поскольку осуществляет активное выведение (выкачивание) из протоплазмы проникающих в нее ионов натрия и введение (нагнетание) в нее ионов калия. Источником энергии для работы ионного насоса является расщепление АТФ (аденозинтрифосфата), которое происходит под воздействием фермента аденозинтрифосфатазы, локализованного в мембране клетки и активируемого теми же ионами, т. е. калием и натрием (нагрий-калий-зависимая АТФ-аза).

Рис. 2.2.
Это крупный белок, превышающий по размеру толщину клеточной мембраны. Молекула этого белка, пронизывая мембрану насквозь, связывает с внутренней стороны преимущественно натрий и АТФ, а с наружной - калий и различные ингибиторы типа глико- зидов. При этом возникает мембранный ток. Благодаря этому току обеспечивается соответствующее направление переноса ионов. Перенос ионов происходит в три этапа. Сначала ион соединяется с молекулой переносчика, образуя комплекс ион-переносчик. Затем этот комплекс проходит через мембрану или переносит через нее заряд. В завершении - ион освобождается от переносчика на противоположной стороне мембраны. Одновременно происходит аналогичный процесс, переносящий ионы в противоположном направлении. Если насос осуществляет перенос одного иона натрия на один ион калия, то он просто поддерживает концентрационный градиент по обе стороны мембраны, но не вносит вклада в создание мембранного потенциала. Чтобы внести этот вклад, ионный насос должен переносить натрий и калий в соотношении 3:2, т. е. на 2 иона калия, поступающих в клетку, из клетки он должен выводить 3 иона натрия. Работая с максимальной нагрузкой, каждый насос способен перекачивать через мембрану около 130 ионов калия и 200 ионов натрия в секунду. Это предельная скорость. В реальных условиях работа каждого насоса регулируется в соответствии с потребностями клетки. У большинства нейронов на один квадратный микрон мембранной поверхности приходится от 100 до 200 ионных насосов. Следовательно, мембрана любой нервной клетки содержит 1 миллион ионных насосов, способных перемещать до 200 миллионов ионов натрия в секунду.
Таким образом, мембранный потенциал (потенциал покоя) создается в результате как пассивных, так и активных механизмов. Степень участия тех или иных механизмов в разных клетках неодинакова, из чего следует, что мембранный потенциал может быть неодинаковым в разных структурах. Активность насосов может зависеть от диаметра нервных волокон: чем тоньше волокно, тем отношение размера поверхности к объему цитоплазмы выше, соответственно, и активность насосов, необходимая для поддержания разницы концентраций ионов на поверхности и внутри волокна, должна быть больше. Другими словами, мембранный потенциал может зависеть от структуры нервной ткани, а значит, и от ее функционального назначения. Электрическая поляризация мембраны - главное условие, обеспечивающее возбудимость клетки. Это ее постоянная готовность к действию. Это запас потенциальной энергии клетки, который она может использовать в случае, если нервной системе понадобится ее немедленная реакция.