
ΔΟΜΗ ΠΡΩΤΕΪΝΩΝ
Στη δομή των πρωτεϊνών, υπάρχουν τέσσερα επίπεδα μοριακής οργάνωσης: πρωτογενείς, δευτεροταγείς, τριτοταγείς και τεταρτοταγείς δομές. Τα δύο πρώτα επίπεδα είναι χαρακτηριστικά όλων των πρωτεϊνών. Τριτοταγείς και τεταρτοταγείς δομές εμφανίζονται μόνο σε σφαιρικές πρωτεΐνες.
Πρωτογενής δομή πρωτεϊνών
|
Σχηματισμός πεπτιδικού δεσμού |
Η κύρια δομή των πρωτεϊνών είναι η αλληλουχία των υπολειμμάτων αμινοξέων στην πολυπεπτιδική αλυσίδα. Η σειρά των αμινοξέων σε μια πρωτεΐνη καθορίζεται γενετικά από την αλληλουχία των νουκλεοτιδίων στο DNA. Ένα πολυπεπτίδιο σχηματίζεται από την αλληλεπίδραση της καρβοξυλικής ομάδας ενός αμινοξέος με την αμινομάδα ενός άλλου αμινοξέος - ενός πεπτιδικού δεσμού.
Η κεφαλή (NH2-) ενός αμινοξέος συνδέεται με την ουρά (-COOH) ενός άλλου αμινοξέος. Ένας πεπτιδικός δεσμός (-CO-NH-) σχηματίζεται μεταξύ αμινοξέων, ο οποίος είναι ο μόνος τύπος δεσμού στην πρωτογενή δομή μιας πρωτεΐνης. Όπως φαίνεται από το παραπάνω διάγραμμα, όταν σχηματίζεται ένας πεπτιδικός δεσμός, απελευθερώνεται νερό. Η διάσπαση του πεπτιδικού δεσμού κατά την υδρόλυση συνοδεύεται από την προσθήκη νερού στη θέση του διασπασμένου δεσμού. Το τελικό προϊόν της υδρόλυσης πρωτεϊνών και πολυπεπτιδίων είναι τα ελεύθερα αμινοξέα.
Ο πεπτιδικός δεσμός είναι ισχυρότερος μεμονωμένα ομόλογαμεταξύ άνθρακα και αζώτου, αφού ο ταυτομερισμός που προκύπτει είναι διπλάσιος κατά 40%. Για τον ίδιο λόγο, σε μια πολυπεπτιδική αλυσίδα, η περιστροφή είναι δυνατή μόνο γύρω από τα άτομα άνθρακα που σχετίζονται με τη ρίζα
Ο σκελετός όλων των πολυπεπτιδίων είναι ο ίδιος. Οι πολυπεπτιδικές αλυσίδες διαφέρουν ως προς τη φύση και την αλληλουχία των ριζών. Ένα πολυπεπτίδιο ονομάζεται από τον αριθμό των υπολειμμάτων αμινοξέων που το αποτελούν: διπεπτίδιο, τριπεπτίδιο κ.λπ.
Οι πρωτεΐνες είναι πολυπεπτίδια που περιέχουν περισσότερα από 50 υπολείμματα αμινοξέων. Η απλούστερη πρωτεΐνη είναι η ινσουλίνη. Περιέχει μόνο 51 υπολείμματα αμινοξέων. Η ριβονουκλεάση περιλαμβάνει 124 υπολείμματα, αιμοσφαιρίνη 574.
Στις πρωτεΐνες, η αλληλουχία των αμινοξέων, δηλαδή η πρωτογενής δομή, είναι αυστηρά καθορισμένη. Η αντικατάσταση ενός υπολείμματος αμινοξέος με ένα άλλο παράγει μια νέα πρωτεΐνη. Έτσι, στη βόεια ινσουλίνη υπάρχει ένα υπόλειμμα σερίνης στην ένατη θέση και στην ινσουλίνη προβάτου υπάρχει ένα υπόλειμμα γλυκίνης. Στην ινσουλίνη ανθρώπου και αλόγου, οι διαφορές αφορούν τρία υπολείμματα αμινοξέων - το όγδοο, το ένατο και το δέκατο. Όλες οι αναφερόμενες ινσουλίνες έχουν διαφορετικές πρωτογενείς δομές. σκίουροι διαφορετικούς οργανισμούςμε την ίδια συνάρτηση λέγονται ομόλογα.
Δευτερογενής δομή πρωτεϊνών
Υπάρχουν δύο κύριοι τύποι δευτερογενούς δομής πρωτεϊνών: σπειροειδές και διπλωμένο στρώμα.
Σπείρες . Λόγω της ελεύθερης περιστροφής των δεσμών γύρω από το άτομο α-άνθρακα στην πολυπεπτιδική αλυσίδα, η γραμμικότητα της πολυπεπτιδικής αλυσίδας διαταράσσεται. Αυτό οδηγεί στο σχηματισμό σπειρών. Υπάρχουν 3 είδη σπιράλ.
1. Χαρακτηριστικό της κερατίνης α-σ πειρατικό. Η πολυπεπτιδική αλυσίδα της κερατίνης είναι, σαν να λέγαμε, τυλιγμένη γύρω από έναν φανταστικό κύλινδρο. Τα πηνία είναι το ένα δίπλα στο άλλο συγκρατούνται με δεσμούς υδρογόνουμεταξύ του οξυγόνου ενός πεπτιδικού δεσμού και του υδρογόνου ενός άλλου πεπτιδικού δεσμού. Οι δεσμοί υδρογόνου είναι 20 φορές πιο αδύναμοι από τους ομοιοπολικούς δεσμούς μεταξύ οξυγόνου και υδρογόνου, αλλά λόγω της αφθονίας τους συγκρατούν την έλικα αρκετά σταθερά.
2. β -σπειροειδήςπου βρίσκεται σε βακτηριακή πρωτεΐνη. Μια στροφή της έλικας p αποτελείται από 22 υπολείμματα αμινοξέων, η β-έλικα είναι ένας κοίλος σωλήνας και η α-έλικα είναι ένας γεμάτος κύλινδρος.
3. Σπασμένη σπείραχαρακτηριστικό του κολλαγόνου. Αυτός ο τύπος έλικας είναι συνέπεια της υψηλής περιεκτικότητας σε γλυκίνη και προλίνη με υδροξυπρολίνη σε κολλαγόνο - αμινοξέα που διαταράσσουν την «ορθότητα» της έλικας.
ΜΕ στρώση τοιχοποιίας χαρακτηριστικό της πρωτεΐνης μεταξιού - ινώδες. Η κατεύθυνση των γειτονικών αλυσίδων στο διπλωμένο στρώμα είναι αντίθετη (αντιπαράλληλη) Οι αλυσίδες είναι η μία δίπλα στην άλλη συγκρατείται από δεσμούς υδρογόνου.
Οι έλικες και τα διπλωμένα στρώματα σε ινιδιακές πρωτεΐνες συχνά δημιουργούν υπερ-δευτερεύουσες δομές ή υπερπηνία. Έτσι, 7 α-έλικες κερατίνης δίνουν μια υπερέλικα. Με τη σειρά τους, 11 υπερσπείρες κερατίνης σχηματίζουν ένα μικροϊνίδιο μαλλιών.
Η δευτερογενής δομή των σφαιρικών πρωτεϊνών δεν είναι τόσο ομοιόμορφη όσο αυτή των ινιδιακών πρωτεϊνών. Έτσι, στο μόριο της μυοσφαιρίνης, το 77% της πολυπεπτιδικής αλυσίδας είναι ελικοειδές και το 23% δεν είναι ελικοειδές. Ο βαθμός περιέλιξης της ινσουλίνης είναι 60%, η αλβουμίνη αυγού είναι 40%, η πεψίνη είναι 28%. Η πολυπεπτιδική αλυσίδα της χυμοθρυψίνης δεν περιέχει σχεδόν καθόλου ελικοειδείς περιοχές, αλλά υπάρχουν πτυχές, στρώματα, βρόχοι, κάμψεις κ.λπ.
Στη δομή των σφαιρικών πρωτεϊνών με μοριακό βάρος πάνω από 20 χιλ. Ναι, η έννοια διακρίνεται τομέα - μικρές περιοχές 100-150 υπολειμμάτων αμινοξέων με χαρακτηριστική δομή. Ονομάζονται δομικοί τομείς.
Μεταξύ τομέων και μεμονωμένων δομικών στοιχείων εντός ενός τομέα υπάρχουν τα λεγόμενα τμήματα μεντεσέδων . Συχνά, πολλές παρόμοιες περιοχές του ίδιου τύπου βρίσκονται σε μία πρωτεΐνη.
Υπάρχει επίσης μια έννοια λειτουργικό τομέα . Στην τελευταία περίπτωση, ένας ή περισσότεροι δομικοί τομείς μαζί σχηματίζουν μια λειτουργικά απομονωμένη περιοχή στο μόριο πρωτεΐνης: μια θέση υποστρώματος, το περιβάλλον του ενεργού κέντρου ενός ενζύμου ή αναστολέα, ένα κανάλι ιόντων στη μεμβράνη κ.λπ.
Τριτογενής δομή- θέση της πολυπεπτιδικής αλυσίδας (σπειροειδής, ελαφρώς ελικοειδής ή μη ελικοειδής) σε τρισδιάστατο χώρο.
Παρά τη φαινομενική διαταραχή της σφαιρικής μπάλας, η δομή της είναι αυστηρά καθορισμένη και έχει κάποιες κανονικότητες.
1. Οι πολυπεπτιδικές αλυσίδες στο σφαιρίδιο είναι συσκευασμένες πολύ σφιχτά.
2. Συνήθως οι πολικές ομάδες της πρωτεΐνης βρίσκονται στην επιφάνεια του σφαιριδίου, και οι υδρόφοβες ρίζες κρύβονται μέσα σε αυτό.
Ακετόνη" href="/text/category/atceton/" rel="bookmark">ακετόνη, η πρωτεΐνη κατακρημνίζεται. Αυτή η κατακρήμνιση ονομάζεται αλάτισμα έξω. Ο μηχανισμός εξάλειψης είναι ότι τα ιόντα αλατιού και τα μόρια αλκοόλης και ακετόνης, που έχουν ένα δικό τους ισχυρό κέλυφος ενυδάτωσης, αφαιρούν νερό από το μόριο της πρωτεΐνης. Διαφορετικές πρωτεΐνες αλατίζονται σε διαφορετικές συγκεντρώσεις αλατιού. Οι σφαιρίνες αλατίζονται σε ένα ημι-κορεσμένο διάλυμα θειικού αμμωνίου και οι λευκωματίνες μόνο σε ένα κορεσμένο διάλυμα αυτού του άλατος. Το κλασματικό αλάτισμα χρησιμοποιείται για τον διαχωρισμό και τον καθαρισμό πρωτεϊνών.
Ορισμένες πρωτεΐνες καθιζάνουν σε pH που αντιστοιχεί σε ισοηλεκτρικό σημείο. Έτσι, η καζεΐνη καθιζάνει σε pH 4,7, αφού σε αυτή την τιμή pH τα μόρια δεν έχουν φορτίο και γρήγορα συσσωματώνονται σε μεγάλα σωματίδια που είναι ασταθή στο διάλυμα. Άλλες πρωτεΐνες είναι πιο σταθερές και η καθίζηση τους απαιτεί δράση και στους δύο παράγοντες σταθερότητας της πρωτεΐνης.
Αιμοκάθαρση πρωτεϊνών
Χάρη σε μεγάλα μεγέθη, τα μόρια πρωτεΐνης δεν διεισδύουν μέσω ορισμένων μεμβρανών. σελοφάν, κύστη ψαριού κ.λπ. Αυτή η ιδιότητα χρησιμοποιείται για τον καθαρισμό πρωτεϊνών από ακαθαρσίες χαμηλού μοριακού βάρους, δηλαδή για αιμοκάθαρση.
Ένα διάλυμα πρωτεΐνης αναμεμειγμένο με άλατα χύνεται σε μια πλαστική σακούλα, η σακούλα τοποθετείται σε ένα δοχείο μέσω του οποίου ρέει απεσταγμένο νερό. Μικρά ιόντα αλατιού και άλλες ουσίες διεισδύουν μέσω του σελοφάν στο νερό και απομακρύνονται, ενώ το διάλυμα πρωτεΐνης παραμένει στη σακούλα.
Φόρτωση πρωτεΐνης
Στην πρωτεΐνη, κατά κανόνα, το άθροισμα των όξινων, αρνητικά φορτισμένων αμινοξέων (γλουταμινικό, ασπαρτικό) δεν είναι ίσο με το άθροισμα των βασικών, θετικά φορτισμένων αμινοξέων (λυσίνη, αργινίνη, ιστιδίνη). Εξαιτίας αυτού, οι πρωτεΐνες στο νερό έχουν είτε θετικό είτε αρνητικό φορτίο. Όταν ένα διάλυμα μιας τέτοιας πρωτεΐνης οξινίζεται (σε περίσσεια Η+), ο ιονισμός των καρβοξυλομάδων θα καταστέλλεται και έρχεται μια στιγμή που το άθροισμα των θετικά φορτισμένων ομάδων θα είναι ίσο με το άθροισμα των αρνητικά φορτισμένων ομάδων. Σε αυτή την περίπτωση, το μόριο της πρωτεΐνης ως σύνολο δεν έχει φορτίο. Αυτή η κατάσταση της πρωτεΐνης ονομάζεται ισο εηλεκτρικό, και το pH στο οποίο εμφανίζεται η ισοηλεκτρική κατάσταση ονομάζεται ισοηλεκτρικό σημείο (IEP). Το IET είναι ένα από τα πιο σημαντικά χαρακτηριστικά της πρωτεΐνης.
Με περαιτέρω οξίνιση του διαλύματος, η πρωτεΐνη φορτίζεται θετικά. Τα μόρια πρωτεΐνης επαναφορτίζονται. Εάν πάρετε μια θετικά φορτισμένη πρωτεΐνη, τότε όταν αλκαλοποιηθεί, αποκτά πρώτα μια ισοηλεκτρική κατάσταση και στη συνέχεια φορτίζεται αρνητικά.
Ο γενικός κανόνας είναι ο εξής: σε pH κάτω από το IET, μια πρωτεΐνη είναι θετικά φορτισμένη και είναι κατιόν, και σε pH πάνω από το IET, είναι αρνητικά φορτισμένη και είναι ανιόν.
Η διαφορά στο φορτίο των πρωτεϊνών επιτρέπει τον διαχωρισμό τους σε ένα σταθερό ηλεκτρικό πεδίο. Αυτή η μέθοδος διαχωρισμού ονομάζεται ηλεκτροφόρηση.
Η ιοντοανταλλακτική χρωματογραφία βασίζεται επίσης στη διαφορά στο φορτίο των διαχωρισμένων ουσιών του μείγματος.
Μετουσίωσης πρωτεϊνών
Η μετουσίωση είναι κάθε μη υδρολυτική αλλαγή στη δομή των πρωτεϊνών, που συνοδεύεται από αλλαγή στη βιολογική τους δραστηριότητα και λειτουργία. Η μετουσίωση μπορεί να προκληθεί από πολλούς παράγοντες: βρασμό, υψηλή θερμοκρασία, υπεριώδη και ιοντίζουσα ακτινοβολία, υπερβολική πίεση, άλατα βαρέων μετάλλων, ακραίες τιμές pH (ισχυρά οξέα και αλκάλια), ορισμένες οργανικές ενώσεις.
Η θέρμανση και τα διάφορα είδη ακτινοβολίας καταστρέφουν το υδρογόνο και ιοντικούς δεσμούς. Ισχυρά οξέα, αλκάλια και πυκνά διαλύματα αλάτων διασπούν τους ιοντικούς δεσμούς. Βαριά μέταλλασχηματίζουν ισχυρούς δεσμούς με τα καρβοξυανιόντα και διασπούν τους ιοντικούς δεσμούς. Οι οργανικοί διαλύτες και τα απορρυπαντικά διαταράσσουν τις υδρόφοβες αλληλεπιδράσεις και σπάνε τους δεσμούς υδρογόνου στις πρωτεΐνες.
Κατά τη διάρκεια της μετουσίωσης, όλοι οι ασθενείς δεσμοί σε μια πρωτεΐνη αλλάζουν ή καταστρέφονται: υδρογόνο, ηλεκτροστατικοί, υδρόφοβοι κ.λπ., αλλά οι πεπτιδικοί δεσμοί παραμένουν άθικτοι.
Τα σημάδια της μετουσίωσης είναι:
1) αλλαγή στη διαλυτότητα. Η διαλυμένη στο νερό πρωτεΐνη κατακρημνίζεται ή, αντίθετα, η αδιάλυτη πρωτεΐνη μεταφέρεται σε διάλυμα.
2) αλλαγή στην οπτική δραστηριότητα, για παράδειγμα, η γωνία περιστροφής του επιπέδου της πολωμένης δέσμης.
3) η εμφάνιση νέων αντιδραστικών ομάδων κρυμμένων πριν από τη μετουσίωση μέσα στο πρωτεϊνικό σφαιρίδιο.
4) το κύριο και πρώτο σημάδι της μετουσίωσης είναι η απώλεια λειτουργίας. Η δομική πρωτεΐνη χαλαρώνει, τα ένζυμα χάνουν την καταλυτική τους δράση κ.λπ.
Αφού απελευθερωθεί από τον μετουσιωτικό παράγοντα, η πρωτεΐνη ανακτά σταδιακά τις αρχικές της ιδιότητες. Αυτή η διαδικασία ονομάζεται αναγέννηση.
Οπτικές ιδιότητες πρωτεϊνών
Με εξαίρεση τις χρωμοπρωτεΐνες, οι πρωτεΐνες δεν έχουν χρώμα. Οι πρωτεΐνες απορροφούν το υπεριώδες φως με μέγιστο στα λ=280 nm λόγω των αρωματικών αμινοξέων. Το δεύτερο μέγιστο απορρόφησης στα λ=216 nm ανήκει στον πεπτιδικό δεσμό.
Τα διαλύματα πρωτεΐνης είναι διαφανή, αλλά έχουν ωχρότητα - η θολότητα είναι ορατή όταν φωτίζεται από το πλάι. Οι ιδιότητες που αναφέρονται χρησιμοποιούνται για τον ποσοτικό προσδιορισμό της πρωτεΐνης.
ΜΟΝΟΝΟΥΚΛΕΟΤΙΔΙΑ


Πουρίνη Γουανίνη Αδενίνη
Πυριμιδίνη Κυτοσίνη Θυμίνη Ουρακίλη
Εκτός από τις αναφερόμενες βάσεις, υπάρχουν μεθυλιωμένα, θείο και άλλα παράγωγα αζωτούχων βάσεων. Καλούνται δευτερεύουσες βάσεις. Για παράδειγμα, στα προκαρυωτικά υπάρχουν: ριβοθυμιδίνη, ινοσίνη, ξανθίνη, υποξανθίνη κλπ. Συνολικά είναι γνωστές περίπου 60 αζωτούχες βάσεις.
Οι βάσεις αζώτου και οι ενώσεις που κατασκευάζονται από αυτές απορροφούν εντατικά το φως στην υπεριώδη περιοχή (260-280 nm). Αυτή η ιδιότητα χρησιμοποιείται για τον ποσοτικό προσδιορισμό ουσιών που περιέχουν αζωτούχες βάσεις.
https://pandia.ru/text/78/240/images/image009_58.jpg" alt="http://*****/biohimija_severina/img/B5873p267-a1.jpg" align="left" width="289" height="203 src=">Важным производным нуклеозидов является !} κατασκήνωση. Σχηματίζεται από ΑΤΡ με τη συμμετοχή του ενζύμου αδενυλική κυκλάση. Το cAMP εμπλέκεται στη ρύθμιση των μεταβολικών διεργασιών στο κύτταρο. Συγκεκριμένα, δρα ως δεύτερος αγγελιοφόρος στη δράση ορισμένων ορμονών στο κύτταρο. .
Οι ενώσεις που κατασκευάζονται όπως τα νουκλεοτίδια αποτελούν μέρος ορισμένων πολύπλοκων ενζύμων, που εκτελούν το ρόλο συνένζυμα. Συχνά, η σύνθεση τέτοιων συνενζύμων περιέχει αζωτούχες ουσίες που διαφέρουν στη δομή από τις βάσεις πουρίνης και πυριμιδίνης. Δεν συντίθενται στο σώμα των ζώων, αλλά προέρχονται από τις τροφές (βιταμίνες).
μονονουκλεοτίδιο φλαβίνης ( FMN) - φωσφορυλιωμένο ριβοφλαβίνη(βιταμίνη Β2).
δινουκλεοτίδιο αδενίνης φλαβίνης ( ΦΑΝΤΑΣΙΟΠΛΗΞΙΑ) αποτελείται από δύο νουκλεοτίδια AMP και FMN.

Το συνένζυμο Α ενεργοποιεί και μεταφέρει ακυλικές ρίζες, οι οποίες προσκολλώνται στην ομάδα SH χρησιμοποιώντας έναν δεσμό θειοαιθέρα.
Ανάλογα με το οξύ που μεταφέρουν, οι ενώσεις ονομάζονται: ακετυλο-συνένζυμο Α, μηλονυλο-συνένζυμο Α, ηλεκτρυλο-συνένζυμο Α.
ΝΟΥΚΛΕΪΚΑ ΟΞΕΑ
Νουκλεϊκά οξέα- Το δεοξυριβονουκλεϊκό οξύ (DNA) και το ριβονουκλεϊκό οξύ (RNA) είναι πολυμερή δεοξυριβονουκλεοτιδίων και ριβονουκλεοτιδίων, αντίστοιχα. Τα μονονουκλεοτίδια στα νουκλεϊκά οξέα συνδέονται μέσω ενός υπολείμματος φωσφορικό οξύμεταξύ ο πέμπτος άνθρακας της ριβόζηςΚαι τρίτος άνθρακας της ριβόζηςγειτονικό νουκλεοτίδιο.
Δομή του DNA
Το 1950, ο Chargaff ανακάλυψε μια σειρά από μοτίβα στη νουκλεοτιδική σύνθεση του DNA, τα οποία αργότερα ονομάστηκαν κανόνες Chargaff. Αυτοί είναι οι κανόνες: 1) Pur=Pir, 2) A=T, 3) G=C, 4) A+C=G+T. Οι κανόνες του Chargaff βοήθησαν στη διαμόρφωση του συμπληρωματικού μοντέλου της δομής του DNA.
Πρωτογενής δομή του DNAαντιπροσωπεύεται από πολυνουκλεοτιδικές αλυσίδες.
|
 Δομή του DNA
Δομή του DNAΥπάρχουν διάφορα μοτίβα στις αλυσίδες DNA:
1) Σε ιούς και προκαρυώτες, σχεδόν ολόκληρη η αλληλουχία DNA είναι μοναδική· στους ευκαρυώτες, το 30-40% του DNA αποτελείται από επαναλαμβανόμενες αλληλουχίες, ειδικά πολλές επαναλαμβανόμενες τομές DNA στην περιοχή του κεντρομερούς.
2) Οι αλυσίδες DNA δεν έχουν κλάδους.
3) Υπάρχουν πολλές (χιλιάδες) αντίστροφες ακολουθίες τρεξίματος στο DNA - παλίνδρομα, «αντιστροφές». Παραδείγματα μετατόπισης στα ρωσικά: «ο κάπρος πίεσε τη μελιτζάνα». Τα παλίνδρομα σχηματίζουν δομές σε σχήμα σταυρού - φουρκέτες, οι οποίες παίζουν σημαντικό ρόλο στη ρύθμιση της γονιδιακής έκφρασης (εργασία).
Δευτερογενής δομή του DNA
Το 1953, οι J. Watson και F. Crick διαπίστωσαν ότι το DNA είναι μια διπλή έλικα αντιπαράλληλοςπολυνουκλεοτιδικές αλυσίδες. Οι αλυσίδες συγκρατούνται η μία κοντά στην άλλη με δεσμούς υδρογόνου που σχηματίζονται μεταξύ αζωτούχων βάσεων, με διπλό δεσμό μεταξύ αδενίνης και θυμίνης και τριπλό δεσμό μεταξύ κυτοσίνης και γουανίνης. Έξω από τη διπλή έλικα του DNA υπάρχει μια ραχοκοκαλιά από φωσφορικό σάκχαρο.
Οι συμπληρωματικές συνδεδεμένες αζωτούχες βάσεις είναι στραμμένες προς τα μέσα. Σε μια στοίβα, οι αζωτούχες βάσεις μετατοπίζονται μεταξύ τους. Υπάρχουν διάφοροι τύποι έλικας DNA:
1) έλικα τύπου Β, που βρέθηκε κατά την αντιγραφή του DNA.
2) έλικα τύπου Α, που παρατηρείται κατά τη μεταγραφή.
3) μια σπείρα τύπου Ζ, στριμμένη προς τα αριστερά, και όχι προς τα δεξιά όπως μια σπείρα Α ή Β, εμφανίζεται κατά τη διάρκεια της διέλευσης.
4) Έχουν επίσης περιγραφεί έλικες τύπου C και SBS. Το τελευταίο δεν είναι στριμμένο.
Οι ιοί μπορεί να περιέχουν μονόκλωνο DNA.
Τριτογενής δομή του DNA
Στα προκαρυωτικά, τα μόρια του DNA είναι κυκλικά. Στους ευκαρυώτες, τα άκρα του DNA είναι ελεύθερα - αυτό είναι γραμμική μορφή DNA. Οι ιοί έχουν γραμμικό και κυκλικό DNA.
Οι προκαρυώτες δεν έχουν πυρήνα. Σε αυτά, το DNA μαζί με τις πρωτεΐνες συνδέεται με την κυτταροπλασματική μεμβράνη, σχηματίζοντας ένα νουκλεοειδές.
Στους ευκαρυώτες, το DNA διαχωρίζεται από το υπόλοιπο κύτταρο με την πυρηνική μεμβράνη. Κατά τη διάρκεια της ενδιάμεσης φάσης, το ευκαρυωτικό DNA συγκεντρώνεται νήματα χρωματίνης. Η χρωματίνη περιέχει πρωτεΐνες εκτός από το DNA. 50% πρωτεΐνη χρωματίνης - ιστόνες. Οι ιστόνες περιέχουν μεγάλος αριθμόςυπολείμματα διαμινοκαρβοξυλικού οξέος: αργινίνη και λυσίνη. Αυτές είναι πολύ συντηρητικές σφαιρικές πρωτεΐνες που είναι σχεδόν ίδιες σε όλους τους ευκαρυώτες. Το δεύτερο μισό των πρωτεϊνών χρωματίνης είναι πρωτεΐνες μη ιστόνης, που χαρακτηρίζονται από μεγάλη ποικιλομορφία.
 Η χρωματίνη έχει πολλά επίπεδα οργάνωσης:
Η χρωματίνη έχει πολλά επίπεδα οργάνωσης:
1) Νουκλεοσώματα. Σχεδόν δύο στροφές DNA τυλίγονται σε έναν πυρήνα τεσσάρων ζευγών μορίων ιστόνης. Αυτό - πυρήνας. Βρίσκεται ανάμεσα στις κρούστες συνδέτης- 40 ζεύγη νουκλεοτιδίων, μερικώς καλυμμένα από πρωτεΐνες ιστόνης και (ή) μη ιστονικές πρωτεΐνες ή δεν καλύπτονται καθόλου από πρωτεΐνες. Οι ιστόνες εμπλέκονται στη γονιδιακή ενεργοποίηση και καταστολή σε μεταγραφικό επίπεδο.
2) Σωληνοειδή: 6-10 νουκλεοσώματα δίνουν μια στροφή του σωληνοειδούς.
3) Βρόχοι. Στη ραχοκοκαλιά των πρωτεϊνών μη ιστόνης, βρίσκονται βρόχοι 30-90 χιλιάδων ζευγών βάσεων, στους οποίους η αρχή και το τέλος βρίσκονται κοντά.
4) Υψηλότερο επίπεδοΗ οργάνωση του DNA στους ευκαρυώτες είναι το χρωμόσωμα. Η βάση ενός χρωμοσώματος είναι μια πρωτεϊνική μήτρα στην οποία συνδέεται το DNA. Στα άκρα του χρωμοσώματος υπάρχουν τμήματα DNA που ονομάζονται τελομερή. Η αναπαραγωγή μπορεί να ξεκινήσει από τα τελομερή. Τα τελομερή προστατεύουν τα άκρα των χρωμοσωμάτων από την αποικοδόμηση.
Με κάθε αντιγραφή, τα τελομερή μικραίνουν. Έχοντας φτάσει σε ένα ορισμένο κρίσιμα μικρό μέγεθος τελομερών, το κύτταρο πεθαίνει. Η τελομεράση είναι ένα ένζυμο που αποκαθιστά το μήκος των τελομερών, καθιστώντας το κύτταρο αθάνατο. Η τελομεράση υπάρχει στα αναπαραγωγικά όργανα, στέλεχος Καικαρκινικά κύτταρα, αλλά όχι σε άλλα κύτταρα. Στο κέντρο του χρωμοσώματος βρίσκεται κεντρομερίδιο- επίσης μη κωδικοποιητικό DNA, το οποίο διασφαλίζει τον σωστό διαχωρισμό των χρωμοσωμάτων κατά την κυτταρική διαίρεση.
Το μεγαλύτερο μέρος του DNA είναι σε βρόχους. Εδώ βρίσκονται τα γονίδια. Κάθε βρόχος περιέχει ένα ή περισσότερα γονίδια. Οι βρόχοι αλληλεπιδρούν με τη χρωμοσωμική μήτρα μέσω μη κωδικοποιητικών περιοχών DNA.
Φυσικοχημικές ιδιότητες του DNA
Ένα χρωμόσωμα είναι ένα μόριο DNA. Οι προκαρυώτες έχουν μόνο ένα χρωμόσωμα. Τα μεγέθη του DNA ποικίλλουν από 5000 νουκλεοτίδια στους ιούς έως 5 δισεκατομμύρια (το μήκος του είναι 8 cm) στον άνθρωπο. Το μήκος του DNA όλων των χρωμοσωμάτων ενός ανθρώπινου κυττάρου είναι περίπου 2 m.
Το DNA είναι μια λευκή ινώδης μάζα. Τα διαλύματα είναι πολύ παχύρρευστα. Το ιξώδες αυξάνεται με την αύξηση του μοριακού βάρους. Τα διαλύματα DNA απορροφούν το υπεριώδες φως με μέγιστο στα 260 nm. Το DNA στο νερό είναι αρνητικά φορτισμένο.
Σε όξινα, αλκαλικά περιβάλλοντα σε θερμοκρασία ° C παρουσία φορμαμίδης, ουρίας και ορισμένων άλλων παραγόντων, εμφανίζεται απόκλιση των πολυνουκλεοτιδικών αλυσίδων DNA - μετουσίωση. Η μετουσίωση διασπά τους δεσμούς υδρογόνου - DNA" λιώνει"Η θερμοκρασία τήξης θεωρείται αυτή στην οποία το DNA μετουσιώνεται κατά το ήμισυ (καταστρέφονται οι μισοί δεσμοί υδρογόνου). Κατά την τήξη παρατηρείται αύξηση της οπτικής πυκνότητας των διαλυμάτων στα 260 nm - υπερχρωμικό αποτέλεσμα.
Όσο περισσότερα ζεύγη G-C στο DNA, τόσο υψηλότερο είναι το σημείο τήξης, γιατί Ζεύγη G-Cισχυρότερο από το Α-Τ επειδή συγκρατούνται μεταξύ τους με τρεις δεσμούς υδρογόνου.
Μετά από μείωση της θερμοκρασίας, το DNA που μετουσιώνεται από τη θερμότητα αποκαθιστά τη δευτερογενή δομή του, λαμβάνει χώρα επαναδιάταξη ή ανόπτησηοξέα.
Αν το DNA διαφορετικές πηγέςυποβάλλονται σε μετουσίωση και ανόπτηση στο μείγμα, τότε θα συμβεί υβριδισμός ξένων αλυσίδων DNA σύμφωνα με τους νόμους της συμπληρωματικότητας. Ο υβριδισμός των αλυσίδων DNA και RNA είναι δυνατός. Σε αυτή την περίπτωση, σχηματίζεται ένα υβρίδιο νουκλεϊκό οξύ, στην οποία η μία αλυσίδα είναι RNA, η άλλη είναι DNA.
Τα αμινοξέα είναι αμφοτερικές ενώσεις που συνδέονται μεταξύ τους σε ένα μόριο πρωτεΐνης χρησιμοποιώντας πεπτιδικούς δεσμούς.
Τα α-αμινοξέα μπορούν να συνδεθούν ομοιοπολικά μεταξύ τους χρησιμοποιώντας πεπτιδικούς δεσμούς
Σε σχέση μεταξύ τους, οι πεπτιδικές ομάδες βρίσκονται υπό γωνία.
Η γραμμική αλληλουχία των υπολειμμάτων αμινοξέων σε μια πολυπεπτιδική αλυσίδα ονομάζεται «πρωτογενής δομή μιας πρωτεΐνης».
Ειδικότητα ειδών πρωτεϊνών
Η ατομικότητα των μορίων πρωτεΐνης καθορίζεται από τη σειρά εναλλαγής των αμινοξέων στην πρωτεΐνη. Ωστόσο, πολλές πρωτεΐνες, ενώ εκτελούν την ίδια λειτουργία, διαφέρουν ελαφρώς στη δομή μεταξύ διαφορετικών εκπροσώπων του ίδιου είδους. Ένα παράδειγμα είναι οι πρωτεΐνες της ομάδας αίματος στον άνθρωπο. Αυτή η ποικιλομορφία πρωτεϊνών καθορίζει την ατομική ειδικότητα των οργανισμών.
Κληρονομικές αλλαγές στην πρωτογενή δομή. Πολυμορφισμός πρωτεΐνης
ΠΗ πρωτογενής δομή των πρωτεϊνών προγραμματίζεται από την αλληλουχία των νουκλεοτιδίων στο DNA. Η διαγραφή, η εισαγωγή ή η αντικατάσταση ενός νουκλεοτιδίου στο DNA οδηγεί σε αλλαγή στη δομή της συντιθέμενης πρωτεΐνης. Εάν η αλλαγή στην αλληλουχία aA δεν είναι θανατηφόρα, αλλά προσαρμοστική ή τουλάχιστον ουδέτερη, τότε η νέα πρωτεΐνη μπορεί να κληρονομηθεί και να παραμείνει στον πληθυσμό. Ως αποτέλεσμα, προκύπτουν νέες πρωτεΐνες με παρόμοιες λειτουργίες. Αυτό πολυμορφισμός πρωτεΐνης. Παραδείγματα πολυμορφισμού: ανθρώπινη αιμοσφαιρίνη (εμβρυϊκή, εμβρυϊκή και ενήλικη αιμοσφαιρίνη Α
Κληρονομικές πρωτεϊνοπάθειες: δρεπανοκυτταρική αναιμία, άλλα παραδείγματα.
Οι κληρονομικές πρωτεϊνοπάθειες αναπτύσσονται ως αποτέλεσμα βλάβης του γενετικού μηχανισμού, και επομένως των πρωτεϊνών
αναιμία με την ανακάλυψη ερυθρών αιμοσφαιρίων σε σχήμα ημισελήνου στο αίμα του. Η ασθένεια ονομάζεται δρεπανοκυτταρική αναιμία και προκαλείται από μια αλλαγή στην πρωτογενή δομή του HLA.
Στο μόριο της αιμοσφαιρίνης S, 2 β-αλυσίδες αποδείχθηκαν μεταλλαγμένες, στις οποίες το γλουταμικό στη θέση 6 αντικαταστάθηκε από βαλίνη που περιείχε μια υδρόφοβη ρίζα. Τα «δρεπανοειδή» ερυθρά αιμοσφαίρια δεν περνούν καλά από τα τριχοειδή αγγεία του ιστού· συχνά φράζουν τα αιμοφόρα αγγεία και έτσι δημιουργούν τοπική υποξία.
Οικογενής υπερχοληστερολαιμίαείναι μια γενετική ασθένεια που χαρακτηρίζεται από υψηλά επίπεδα χοληστερόλης στο αίμα και LDL, καθώς και από την πρώιμη εμφάνιση καρδιαγγειακών παθήσεων. Πολλοί ασθενείς έχουν μεταλλάξεις στο γονίδιο του υποδοχέα LDL, το οποίο κωδικοποιεί την αντίστοιχη πρωτεΐνη υποδοχέα LDL ή την απολιποπρωτεΐνη Β (apo-B), η οποία είναι το μέρος της LDL που συνδέεται με τον υποδοχέα
2. Διαμόρφωση πρωτεϊνικών μορίων (δευτερογενείς και τριτοταγείς δομές)
Δευτερεύουσα δομήΜια πρωτεΐνη είναι μια μέθοδος τοποθέτησης μιας πολυπεπτιδικής αλυσίδας στην οποία λαμβάνει χώρα η αλληλεπίδραση των πεπτιδικών ομάδων με το σχηματισμό δεσμών υδρογόνου μεταξύ τους.Στις σφαιρικές πρωτεΐνες κυριαρχεί η α-έλικα, στις ινιδώδεις πρωτεΐνες κυριαρχεί η β-δομή. 
α-Έλικας είναι μια δεξιόστροφη έλικα, που σχηματίζεται από δεσμούς υδρογόνου μεταξύ των πεπτιδικών ομάδων 1 και 4, 4 και 7, 7 και 10 λόγω του σχηματισμού δεσμών υδρογόνου μεταξύ των ατόμων οξυγόνου των ομάδων καρβονυλίου και των ατόμων αζώτου των αμινομάδων.
β-Δομή σχηματίζεται λόγω του σχηματισμού πολλών δεσμών υδρογόνου μεταξύ ατόμων πεπτιδικών ομάδων γραμμικών περιοχών μιας πολυπεπτιδικής αλυσίδας που δημιουργεί κάμψεις, ή μεταξύ διαφορετικών πολυπεπτιδικών αλυσίδων.
Εάν οι συνδεδεμένες πολυπεπτιδικές αλυσίδες κατευθύνονται προς αντίθετες κατευθύνσεις, δημιουργείται μια αντιπαράλληλη β-δομή, αλλά εάν τα Ν- και C-άκρα των πολυπεπτιδικών αλυσίδων συμπίπτουν, σχηματίζεται η δομή ενός παράλληλου β-πτυχωμένου στρώματος ( 
Τριτογενής δομή πρωτεϊνών
Η τριτογενής δομή των πρωτεϊνών είναι μια τρισδιάστατη χωρική δομή που σχηματίζεται λόγω αλληλεπιδράσεων μεταξύ των ριζών αμινοξέων
Δεσμοί που εμπλέκονται στο σχηματισμό της τριτογενούς δομής των πρωτεϊνών
Υδροφοβικές αλληλεπιδράσεις
Οι υδρόφοβες ρίζες αμινοξέων τείνουν να συνδυάζονται εντός της σφαιρικής δομής
Ιοντικοί δεσμοί και δεσμοί υδρογόνουΚαι Ομοιοπολικούς δεσμούς
δισουλφιδικούς δεσμούς, που σχηματίζεται λόγω της αλληλεπίδρασης ομάδων SH δύο υπολειμμάτων κυστεΐνης.
Ενεργή θέση πρωτεϊνών- ένα συγκεκριμένο τμήμα του μορίου πρωτεΐνης, που βρίσκεται στην εσοχή του, που σχηματίζεται από ρίζες που συλλέγονται κατά τον σχηματισμό της τριτογενούς δομής, αλληλεπιδρά με το μόριο του υποστρώματος
διαμορφωτική αστάθεια- μια τάση για αλλαγές στη διαμόρφωση λόγω της θραύσης ορισμένων και του σχηματισμού άλλων αδύναμων δεσμών. Η διαμόρφωση της πρωτεΐνης αλλάζει όταν αλλάζουν οι χημικές και φυσικές ιδιότητες του περιβάλλοντος, όταν η πρωτεΐνη αλληλεπιδρά με άλλα μόρια. Σε αυτή την περίπτωση, συμβαίνει μια αλλαγή στη χωρική δομή και τη διαμόρφωση της πρωτεΐνης στο σύνολό της.
3 . Τεταρτοταγής δομή πρωτεϊνών. Συνεργατικές αλλαγές στη διαμόρφωση του πρωτομερούς. Παραδείγματα δομής και λειτουργίας ολιγομερών πρωτεϊνών: αιμοσφαιρίνη (σε σύγκριση με μυοσφαιρίνη, αλλοστερικά ένζυμα).
Ο αριθμός και η σχετική θέση των πολυπεπτιδικών αλυσίδων στο διάστημα ονομάζεται "τεταρτοταγής δομή πρωτεϊνών". Οι μεμονωμένες πολυπεπτιδικές αλυσίδες σε μια τέτοια πρωτεΐνη είναι πρωτομερή ή υπομονάδες. Συνδυάζουν υδρόφοβα, ιοντικά, υδρογόνο.
Κάθε πρωτομερές αλληλεπιδρά με το άλλο σε πολλά σημεία. Οι υπομονάδες στα ολιγομερή αλληλεπιδρούν πολύ στενά μεταξύ τους, οπότε οποιαδήποτε αλλαγή στη διαμόρφωση οποιασδήποτε υπομονάδας συνεπάγεται απαραίτητα μια αλλαγή σε άλλες υπομονάδες. Αυτό το αποτέλεσμα ονομάζεται συνεργατική αλληλεπίδραση.
Για παράδειγμα, στην αιμοσφαιρίνη, μια τέτοια αλληλεπίδραση υπομονάδων στους πνεύμονες επιταχύνει την προσθήκη O 2 στην αιμοσφαιρίνη κατά 300 φορές.
Μυοσφαιρίνηανήκουν στην κατηγορία των πρωτεϊνών που περιέχουν αίμη, δηλ. περιέχει μια προσθετική ομάδα - αίμη, που σχετίζεται με το πρωτεϊνικό μέρος της απομυοσφαιρίνης. Η μυοσφαιρίνη ταξινομείται ως σφαιρική πρωτεΐνη. έχει μόνο μία πολυπεπτιδική αλυσίδα. Η μυοσφαιρίνη εμπλέκεται στην αποθήκευση οξυγόνου.
Η τριτογενής δομή μοιάζει με ένα συμπαγές σφαιρίδιο 
ΑιμοσφαιρίνεςΌπως και η μυοσφαιρίνη, ταξινομούνται ως αιμοπρωτεΐνες, αλλά έχουν τεταρτοταγή δομή (αποτελούνται από 4 πολυπεπτιδικές αλυσίδες), η οποία καθιστά δυνατή τη ρύθμιση των λειτουργιών τους.
Η αιμοσφαιρίνη Α είναι η κύρια αιμοσφαιρίνη του σώματος των ενηλίκων, αποτελεί περίπου το 98% της συνολικής ποσότητας αιμοσφαιρίνης, τετραμερές, αποτελείται από 2 πολυπεπτιδικές αλυσίδες α και 2 β (2α2β).
Αιμοσφαιρίνη Α 2 2%. Αποτελείται από 2 α- και 2 δ-αλυσίδες.
Τα πρωτομερή αιμοσφαιρίνης, όπως η απομυοσφαιρίνη, αποτελούνται από 8 έλικες κουλουριασμένες σε μια πυκνή σφαιρική δομή που περιέχει έναν εσωτερικό υδρόφοβο πυρήνα και μια «τσέπη» για τη δέσμευση της αίμης. αλλά η τετραμερής δομή της αιμοσφαιρίνης είναι πιο περίπλοκη από τη μυοσφαιρίνη.
Αλλοστερικά ένζυμαείναι ένζυμα των οποίων η δραστηριότητα ρυθμίζεται όχι μόνο από τον αριθμό των μορίων του υποστρώματος, αλλά και από άλλες ουσίες που ονομάζονται τελεστές. έχουν αλλοστερικό κέντρο μακριά από το καταλυτικό ενεργό κέντρο; Οι τελεστές προσκολλώνται στο ένζυμο μη ομοιοπολικά σε αλλοστερικά (ρυθμιστικά) κέντρα.
4 . Έννοια των ενζύμων. Ειδικότητα ενζυμικής δράσης. Ενζυμικοί συμπαράγοντες. Εξάρτηση του ρυθμού των ενζυματικών αντιδράσεων από τη συγκέντρωση του υποστρώματος, το ένζυμο, τη θερμοκρασία και το pH. Αρχές ποσοτικοποίησης ενζύμων. Μονάδες δραστηριότητας.
Τα ένζυμα είναι πρωτεΐνες. αυξάνουν τον ρυθμό μιας χημικής αντίδρασης, αλλά δεν καταναλώνονται.
Τα απλά ένζυμα αποτελούνται μόνο από ΑΑ και τα πολύπλοκα ένζυμα αποτελούνται από 2 μέρη: πρωτεΐνη (αποένζυμο) και μη πρωτεΐνη (συμπαράγοντας). Εάν ένας συμπαράγοντας είναι στενά συνδεδεμένος με το αποένζυμο, ονομάζεται προσθετική ομάδα, εάν είναι χαλαρά συνδεδεμένος, ονομάζεται συνένζυμο.
Σε πολυπεπτιδική αλυσίδα. Σημαντικά χαρακτηριστικά της πρωτογενούς δομής είναι τα διατηρημένα μοτίβα - συνδυασμοί αμινοξέων που παίζουν βασικό ρόλο στις πρωτεϊνικές λειτουργίες. Τα διατηρημένα μοτίβα διατηρούνται σε όλη την εξέλιξη των ειδών και μπορούν συχνά να χρησιμοποιηθούν για την πρόβλεψη της λειτουργίας μιας άγνωστης πρωτεΐνης. Συντηρητικά κίνητρα- σύντομες αλληλουχίες νουκλεοτιδίων στο DNA ή αμινοξέων σε μια πρωτεΐνη που διατηρούνται κατά τη διαδικασία της εξέλιξης, καθώς αυτά τα νουκλεοτίδια ή τα αμινοξέα είναι απαραίτητα για την εκτέλεση οποιωνδήποτε διεργασιών στο κύτταρο.
Δευτερεύουσα δομή- τοπική διάταξη ενός θραύσματος πολυπεπτιδικής αλυσίδας, σταθεροποιημένο με δεσμούς υδρογόνου. α-έλικες- πυκνές στροφές γύρω από τον μακρύ άξονα του μορίου, μια στροφή είναι 3,6 υπολείμματα αμινοξέων και το βήμα της έλικας είναι 0,54 nm, η έλικα σταθεροποιείται με δεσμούς υδρογόνου μεταξύ των πεπτιδικών ομάδων Η και Ο, σε απόσταση 4 μονάδων χώρια. β- φύλλα(διπλωμένα στρώματα) - αρκετές ζιγκ-ζαγκ πολυπεπτιδικές αλυσίδες στις οποίες σχηματίζονται δεσμοί υδρογόνου μεταξύ αμινοξέων ή διαφορετικών πρωτεϊνικών αλυσίδων που είναι σχετικά απομακρυσμένες μεταξύ τους (0,347 nm ανά υπόλειμμα αμινοξέος) στην πρωτοταγή δομή.
Τριτογενής δομή— χωρική δομή της πολυπεπτιδικής αλυσίδας (ένα σύνολο χωρικών συντεταγμένων των ατόμων που αποτελούν την πρωτεΐνη). Δομικά αποτελείται από δευτερεύοντα στοιχεία δομής σταθεροποιημένα διάφοροι τύποιαλληλεπιδράσεις στις οποίες οι υδρόφοβες αλληλεπιδράσεις παίζουν κρίσιμο ρόλο. Στη σταθεροποίηση της τριτογενούς δομής συμμετέχουν τα ακόλουθα: ομοιοπολικούς δεσμούς(μεταξύ δύο υπολειμμάτων κυστεΐνης υπάρχουν δισουλφιδικές γέφυρες). ιοντικούς δεσμούςμεταξύ αντίθετα φορτισμένων πλευρικών ομάδων υπολειμμάτων αμινοξέων. δεσμούς υδρογόνου; υδρόφιλες-υδρόφοβες αλληλεπιδράσειςΌταν αλληλεπιδρά με τα περιβάλλοντα μόρια του νερού, το μόριο της πρωτεΐνης «τείνει» να διπλώνεται έτσι ώστε οι μη πολικές πλευρικές ομάδες αμινοξέων να απομονώνονται από το υδατικό διάλυμα. πολικές υδρόφιλες πλευρικές ομάδες εμφανίζονται στην επιφάνεια του μορίου.
Τεταρτογενής δομή- (υπομονάδα, τομέας) — αμοιβαία διευθέτησηαρκετές πολυπεπτιδικές αλυσίδες ως μέρος ενός συμπλόκου πρωτεϊνών. Μόρια πρωτεΐνης, που αποτελούν μέρος μιας πρωτεΐνης με τεταρτοταγή δομή, σχηματίζονται ξεχωριστά στα ριβοσώματα και μόνο μετά την ολοκλήρωση της σύνθεσης σχηματίζουν μια κοινή υπερμοριακή δομή.
Σύνθεση ΑΚ πρωτεϊνών, πεπτιδικός δεσμός και φυσικοχημικός του. χαρακτήρας
Κύρια μέρη και δομικά στοιχείαΤα συστατικά ενός μορίου πρωτεΐνης είναι αμινοξέα. Τα προϊόντα διατροφής περιέχουν 20 αμινοξέα, 8 από τα οποία δεν συντίθενται στον ανθρώπινο οργανισμό και είναι απαραίτητοι διατροφικοί παράγοντες (βαλίνη, λευκίνη, ισολευκίνη, θρεονίνη, φαινυλαλανίνη, τρυπτοφάνη, μεθειονίνη, λυσίνη). Για παιδιά απαραίτητο αμινοξύείναι η ιστιδίνη. Τα υπόλοιπα αμινοξέα μπορούν να αντικατασταθούν, δηλαδή μπορούν να συντεθούν στο σώμα (αλανίνη, ασπαρτικό οξύ, γλυκόλη, γλυκίνη, γλουταμικό οξύ, προλίνη, σερίνη, τυροσίνη, κυστίνη, κυστεΐνη)
Η ανάγκη για μη απαραίτητα αμινοξέα ικανοποιείται κυρίως μέσω της σύνθεσης στον οργανισμό και εν μέρει μέσω της πρόσληψής τους στην τροφή. Για τη μελέτη της σύστασης αμινοξέων των πρωτεϊνών, χρησιμοποιείται ένας συνδυασμός όξινης (HC1), αλκαλικής [Ba(OH)2] και, λιγότερο συχνά, ενζυματικής υδρόλυσης. Υδροφόβος(μη πολική, αφόρτιστη): γλυκίνη, αλανίνη, βαλίνη, λευκίνη, ισολευκίνη, μεθειονίνη, προλίνη, φαινυλαλανίνη, τρυπτοφάνη. Υδρόφιλο(πολικό, αφόρτιστο): ασπαραγίνη, γλουταμίνη, σερίνη, θρεονίνη, τυροσίνη, κυστεΐνη. Υδρόφιλο (αρνητικό φορτίο): ασπαρτικό, γλουταμικό οξύ. Υδρόφιλο(πολ. φορτίο): λυσίνη, αργινίνη, ιστιδίνη.
Πεπτιδικός δεσμός- ένας τύπος δεσμού αμιδίου που εμφανίζεται κατά τον σχηματισμό πρωτεϊνών και πεπτιδίων ως αποτέλεσμα της αλληλεπίδρασης της α-αμινο ομάδας (-NH 2) ενός αμινοξέος με την α-καρβοξυλική ομάδα (-COOH) ενός άλλου αμινοξέος . Ο πεπτιδικός δεσμός έχει το χαρακτηριστικό ενός μερικού διπλού δεσμού, επομένως είναι βραχύτερος από τους άλλους δεσμούς του πεπτιδικού σκελετού και, ως αποτέλεσμα, έχει μικρή κινητικότητα. Ηλεκτρονική δομήΟ πεπτιδικός δεσμός καθορίζει την επίπεδη, άκαμπτη δομή της πεπτιδικής ομάδας. Τα επίπεδα των πεπτιδικών ομάδων βρίσκονται σε γωνία μεταξύ τους. Οι πεπτιδικοί δεσμοί είναι πολύ ισχυροί και δεν σπάνε αυθόρμητα όταν φυσιολογικές συνθήκεςπου υπάρχουν στα κύτταρα (ουδέτερο περιβάλλον, θερμοκρασία σώματος). Σε εργαστηριακές συνθήκες, η υδρόλυση των πρωτεϊνικών πεπτιδικών δεσμών πραγματοποιείται σε σφραγισμένη αμπούλα με συμπυκνωμένο (6 mol/l) υδροχλωρικό οξύ, σε θερμοκρασία μεγαλύτερη από 105 ° C, και η πλήρης υδρόλυση της πρωτεΐνης σε ελεύθερα αμινοξέα λαμβάνει χώρα σε περίπου μία ημέρα. Στους ζωντανούς οργανισμούς, οι πεπτιδικοί δεσμοί στις πρωτεΐνες σπάνε με τη βοήθεια ειδικών πρωτεολυτικών ενζύμων (από τα αγγλικά, πρωτεΐνη-πρωτεΐνη, λύση-καταστροφή), που ονομάζονται επίσης πρωτεάσες ή πεπτιδικές υδρολάσες.
Αντιδράσεις χρώματος σε αμινοξέα.
Αντίδραση νινυδρίνης: έγχρωμο διάλυμα για α-αμινοξέα όταν θερμαίνεται (μπλε χρώμα).
Αντίδραση ξανθοπρωτεΐνης , κίτρινο χρώμα όταν βράζεται με συμπυκνωμένο νιτρικό οξύ. Μετά την προσθήκη συμπυκνωμένου αλκαλίου, το κίτρινο χρώμα γίνεται πορτοκαλί. Αρωματικά ΑΑ (φαινυλαλανίνη, τυροσίνη και τρυπτοφάνη).

R-tionΤο Adamkiewicz, τρυπτοφάνη σε όξινο περιβάλλον, αλληλεπιδρώντας με όξινες αλδεΰδες, σχηματίζει προϊόντα συμπύκνωσης κόκκινου-ιώδους.

R-tionΦόλια , Τα αμινοξέα που περιέχουν ομάδες σουλφυδρυλίου - SH, υφίστανται αλκαλική υδρόλυση για να σχηματίσουν θειούχο νάτριο Na2S. Το τελευταίο, αλληλεπιδρώντας με το plumbite νατρίου (που σχηματίζεται κατά την αντίδραση μεταξύ οξικού μολύβδου και NaOH), σχηματίζει ένα μαύρο ή καφέ ίζημα θειούχου μολύβδου PbS.

R-tion Millona, Αυτή είναι μια αντίδραση στο αμινοξύ τυροσίνη. Το αντιδραστήριο Millon (διάλυμα HgNO 3 και Hg(NO 2) 2 σε αραιό HNO 3 που περιέχει ένα μείγμα HNO 2) αντιδρά με τυροσίνη για να σχηματίσει το άλας υδραργύρου του νίτρο παραγώγου της τυροσίνης, χρωματισμένο ροζ-κόκκινο.

Εξάρτηση της πρωτεϊνικής διαμόρφωσης από την πρωτογενή δομή τους.
Κάθε πρωτεΐνη βασίζεται σεπολυπεπτιδική αλυσίδα. Δεν είναι απλώς επιμήκης στο χώρο, αλλά οργανώνεται σε μια τρισδιάστατη δομή. Χρησιμοποιώντας ανάλυση περίθλασης ακτίνων Χ, οι Pauling και Corey προσδιόρισαν τις γωνίες των πεπτιδικών δεσμών, αποδεικνύοντας την παρουσία μιας άκαμπτης, επίπεδης (επίπεδης) δομής της πολυπεπτιδικής αλυσίδας. Αν και η διαμορφωτική του κινητικότητα είναι περιορισμένη, η κινητικότητα γύρω από απλούς δεσμούς στο άτομο άνθρακα άλφα είναι δυνατή. Οι γωνίες περιστροφής των απλών δεσμών ονομάζονται στρέψη: η γωνία περιστροφής γύρω από τον δεσμό N-C συμβολίζεται με φ, η γωνία περιστροφής γύρω από Συνδέσεις S-S- ψ. Οι υποκαταστάτες σε σχέση με τον πεπτιδικό δεσμό μπορεί να είναι σε θέση cis ή trans, με τον trans πεπτιδικό δεσμό να είναι πιο σταθερός.
Διαμόρφωση- χωρική σχετική διάταξη τμημάτων του μορίου που στερεώνονται άκαμπτα με ομοιοπολικούς δεσμούς (για παράδειγμα: ανήκουν στη σειρά L στερεοϊσομερών ή στη σειρά D). Για τις πρωτεΐνες η έννοια χρησιμοποιείται επίσης Διαμόρφωσημόριο πρωτεΐνης - μια συγκεκριμένη, αλλά όχι παγωμένη, όχι αμετάβλητη σχετική διάταξη των μερών του μορίου. Δεδομένου ότι η διαμόρφωση ενός μορίου πρωτεΐνης σχηματίζεται με τη συμμετοχή αδύναμων τύπων δεσμών, είναι κινητό (ικανό να αλλάξει) και η πρωτεΐνη μπορεί να αλλάξει τη δομή της. Ανάλογα με τις περιβαλλοντικές συνθήκες, ένα μόριο μπορεί να υπάρχει σε διαφορετικές διαμορφωτικές καταστάσεις, οι οποίες μεταμορφώνονται εύκολα το ένα στο άλλο. Ενεργειακά ευνοϊκές για πραγματικές συνθήκες είναι μόνο μία ή περισσότερες διαμορφωτικές καταστάσεις μεταξύ των οποίων υπάρχει μια ισορροπία.
Οι μεταβάσεις από τη μια διαμορφωτική κατάσταση στην άλλη διασφαλίζουν τη λειτουργία του μορίου της πρωτεΐνης. Πρόκειται για αναστρέψιμες διαμορφωτικές αλλαγές (που βρίσκονται στο σώμα, για παράδειγμα, κατά τη διάρκεια της αγωγής μιας νευρικής ώθησης, κατά τη μεταφορά οξυγόνου από την αιμοσφαιρίνη). Όταν η διαμόρφωση αλλάζει, μερικοί από τους ασθενείς δεσμούς καταστρέφονται και σχηματίζονται νέοι δεσμοί αδύναμου τύπου. Οι ινώδεις πρωτεΐνες είναι σταθερές, αδιάλυτες ουσίες στο νερό και αραιά αλατούχα διαλύματα. Διατεταγμένες παράλληλα μεταξύ τους κατά μήκος ενός άξονα, οι πολυπεπτιδικές αλυσίδες σχηματίζουν μακριές ίνες (ινίδια) ή στρώματα με διαμόρφωση β-δομής. Παραδείγματα ινιδιακών πρωτεϊνών: κολλαγόνο τενόντων και οστικού ιστού, κερατίνη μαλλιών, κερατώδεις σχηματισμοί, δέρμα, νύχια και φτερά, ελαστίνη ελαστικού συνδετικού ιστού.
Οι σφαιρικές πρωτεΐνες είναι ενώσεις των οποίων οι πολυπεπτιδικές αλυσίδες περιελίσσονται σφιχτά σε συμπαγείς σφαιρικές ή σφαιρικές δομές με α-έλικα. Οι περισσότερες σφαιρικές πρωτεΐνες είναι διαλυτές σε υδατικά διαλύματακαι διαχέεται εύκολα. Αυτά περιλαμβάνουν σχεδόν όλα τα επί του παρόντος γνωστά ένζυμα, καθώς και αντισώματα, ορισμένες ορμόνες και πολλές πρωτεΐνες που εκτελούν μια λειτουργία μεταφοράς, για παράδειγμα, αλβουμίνη ορού και αιμοσφαιρίνη. Ορισμένες πρωτεΐνες ανήκουν στον ενδιάμεσο τύπο. Όπως οι ινιδώδεις πρωτεΐνες, αποτελούνται από μακριές δομές σε σχήμα ράβδου και ταυτόχρονα, όπως οι σφαιρικές πρωτεΐνες, είναι διαλυτές σε υδατικά αλατούχα διαλύματα. Αυτές οι πρωτεΐνες περιλαμβάνουν: μυοσίνη - δομικό στοιχείομυς, το ινωδογόνο είναι πρόδρομος του ινώδους που εμπλέκεται στην πήξη του αίματος.
Ο συνδυασμός αμινοξέων μέσω πεπτιδικών δεσμών δημιουργεί μια γραμμική πολυπεπτιδική αλυσίδα που ονομάζεται πρωτογενής πρωτεϊνική δομή
Λαμβάνοντας υπόψη ότι 20 αμινοξέα εμπλέκονται στη σύνθεση πρωτεϊνών και η μέση πρωτεΐνη περιέχει 500 υπολείμματα αμινοξέων, μπορούμε να μιλήσουμε για έναν ασύλληπτο αριθμό πιθανών πρωτεϊνών. Περίπου 100 χιλιάδες διαφορετικές πρωτεΐνες έχουν βρεθεί στο ανθρώπινο σώμα.
Για παράδειγμα, 2 αμινοξέα (αλανίνη και σερίνη) σχηματίζουν 2 πεπτίδια Ala-Ser και Ser-Ala. 3 αμινοξέα θα δώσουν ήδη 6 παραλλαγές του τριπεπτιδίου. 20 αμινοξέα – 1018 διαφορετικά πεπτίδια σε μήκος μόλις 20 αμινοξέων (υποθέτοντας ότι κάθε αμινοξύ χρησιμοποιείται μόνο μία φορά).
Η μεγαλύτερη πρωτεΐνη που είναι σήμερα γνωστή είναι τιτίνης- είναι συστατικό των σαρκομερών μυοκυττάρων, το μοριακό βάρος των διαφόρων ισομορφών του κυμαίνεται από 3000 έως 3700 kDa. Η τιτίνη του ανθρώπινου πέλματος αποτελείται από 38.138 αμινοξέα.
Η πρωτογενής δομή των πρωτεϊνών, δηλ. η αλληλουχία των αμινοξέων σε αυτό προγραμματίζεται από την αλληλουχία των νουκλεοτιδίων στο DNA. Η απώλεια, η εισαγωγή ή η αντικατάσταση ενός νουκλεοτιδίου στο DNA οδηγεί σε αλλαγή στη σύνθεση αμινοξέων και, κατά συνέπεια, στη δομή της συντιθέμενης πρωτεΐνης.
Ένα τμήμα μιας πρωτεϊνικής αλυσίδας μήκους 6 αμινοξέων (Ser-Cys-Tyr-Lei-Glu-Ala)
(οι πεπτιδικοί δεσμοί επισημαίνονται με κίτρινο φόντο, τα αμινοξέα επισημαίνονται με πλαίσιο)
Εάν η αλλαγή στην αλληλουχία αμινοξέων δεν είναι θανατηφόρα, αλλά προσαρμοστική ή τουλάχιστον ουδέτερη, τότε η νέα πρωτεΐνη μπορεί να κληρονομηθεί και να παραμείνει στον πληθυσμό. Ως αποτέλεσμα, προκύπτουν νέες πρωτεΐνες με παρόμοιες λειτουργίες. Αυτό το φαινόμενο ονομάζεται πολυμορφισμόςπρωτεΐνες.
Για πολλές πρωτεΐνες, ανιχνεύεται έντονος δομικός συντηρητισμός. Για παράδειγμα, η ορμόνη ινσουλίνη πρόσωποδιαφέρει από αισιόδοξοςμόνο τρία αμινοξέα, από χοιρινό– ανά ένα αμινοξύ (αλανίνη αντί για θρεονίνη).
Η αλληλουχία και η αναλογία των αμινοξέων στην πρωτογενή δομή καθορίζει τον σχηματισμό δευτερεύων, τριτογενήςΚαι τετραδικόςδομές.
Γονοτυπική ετερογένεια
Ως αποτέλεσμα του γεγονότος ότι κάθε γονίδιο στον άνθρωπο υπάρχει σε δύο αντίγραφα (αλληλόμορφα) και μπορεί να υπόκειται σε μεταλλάξεις (αντικατάσταση, διαγραφή, εισαγωγή) και ανασυνδυασμούς που δεν επηρεάζουν σοβαρά τη λειτουργία της κωδικοποιημένης πρωτεΐνης, εμφανίζεται γονιδιακός πολυμορφισμόςκαι, κατά συνέπεια, πολυμορφισμός πρωτεΐνης. Αναδύονται ολόκληρες οικογένειες σχετικών πρωτεϊνών που έχουν παρόμοιες αλλά διαφορετικές ιδιότητες και λειτουργίες.
Για παράδειγμα, υπάρχει περίπου 300 ΔΙΑΦΟΡΕΤΙΚΟΙ ΤΥΠΟΙαιμοσφαιρίνη, μερικά από αυτά είναι απαραίτητα για διαφορετικά στάδιαοντογένεση: για παράδειγμα, η HbP είναι εμβρυϊκή, σχηματίζεται τον πρώτο μήνα ανάπτυξης, η HbF είναι εμβρυϊκή, απαραίτητη σε μεταγενέστερα στάδια της εμβρυϊκής ανάπτυξης, η HbA και η HbA2 είναι η αιμοσφαιρίνη ενηλίκων. Η ποικιλομορφία εξασφαλίζεται από τον πολυμορφισμό των αλυσίδων σφαιρίνης: η αιμοσφαιρίνη P περιέχει αλυσίδες 2ξ και 2ε, η HbF περιέχει αλυσίδες 2α και 2γ, η HbA περιέχει αλυσίδες 2α και 2β και η HbA2 περιέχει 2α και 2δ αλυσίδες.
Στο δρεπανοκυτταρική αναιμίαστην έκτη θέση της β-αλυσίδας της αιμοσφαιρίνης, το γλουταμινικό οξύ αντικαθίσταται από τη βαλίνη. Αυτό οδηγεί σε σύνθεση αιμοσφαιρίνη S (HbS)- μια αιμοσφαιρίνη που πολυμερίζεται σε μορφή δεοξυ και σχηματίζει κλώνους. Ως αποτέλεσμα, τα ερυθρά αιμοσφαίρια παραμορφώνονται, παίρνουν σχήμα δρεπανιού (μπανάνα), χάνουν την ελαστικότητα και καταστρέφονται όταν περνούν από τριχοειδή αγγεία. Αυτό τελικά οδηγεί σε μειωμένη οξυγόνωση των ιστών και νέκρωση.
Οι ομάδες αίματος AB0 εξαρτώνται από τη δομή ενός ειδικού υδατάνθρακα στη μεμβράνη των ερυθρών αιμοσφαιρίων. Οι διαφορές στη δομή των υδατανθράκων οφείλονται σε διαφορετική ειδικότητα και δραστηριότητα ένζυμο γλυκοσυλ τρανσφεράση, ικανό να τροποποιήσει τον αρχικό ολιγοσακχαρίτη. Το ένζυμο έχει τρεις παραλλαγές και συνδέει είτε Ν-ακετυλογαλακτόζη είτε γαλακτόζη στον ολιγοσακχαρίτη των μεμβρανών των ερυθροκυττάρων ή το ένζυμο δεν συνδέει πρόσθετες ομάδες σακχαρίτη (ομάδα 0).
Ως αποτέλεσμα, τα άτομα με ομάδα αίματος Α0 έχουν έναν ολιγοσακχαρίτη με Ν-ακετυλογαλακτοζαμίνη συνδεδεμένη σε αυτό στα ερυθρά αιμοσφαίρια τους, αυτά με ομάδα αίματος Β0 έχουν ολιγοσακχαρίτη με γαλακτόζη, 00 έχουν μόνο «καθαρό» ολιγοσακχαρίτη, όσοι έχουν ομάδα αίματος ΑΒ έχουν ένας ολιγοσακχαρίτης και Ν-ακετυλογαλακτοζαμίνη, και με γαλακτόζη.
ΕΝΟΤΗΤΑ 1 ΔΟΜΗ, ΙΔΙΟΤΗΤΕΣ ΚΑΙ ΛΕΙΤΟΥΡΓΙΕΣ ΤΩΝ ΠΡΩΤΕΪΝΩΝ
ΕΝΟΤΗΤΑ 1 ΔΟΜΗ, ΙΔΙΟΤΗΤΕΣ ΚΑΙ ΛΕΙΤΟΥΡΓΙΕΣ ΤΩΝ ΠΡΩΤΕΪΝΩΝ
Δομή ενότητας | Θέματα |
Αρθρωτή μονάδα 1 | 1.1. Δομική οργάνωση πρωτεϊνών. Στάδια σχηματισμού φυσικής πρωτεϊνικής διαμόρφωσης 1.2. Βασικά στοιχεία της πρωτεϊνικής λειτουργίας. Φάρμακα ως συνδέτες που επηρεάζουν τη λειτουργία των πρωτεϊνών 1.3. Μετουσίωσης πρωτεϊνών και δυνατότητα αυθόρμητης ανανέωσής τους |
Αρθρωτή μονάδα 2 | 1.4. Χαρακτηριστικά της δομής και της λειτουργίας των ολιγομερών πρωτεϊνών χρησιμοποιώντας το παράδειγμα της αιμοσφαιρίνης 1.5. Διατήρηση της φυσικής πρωτεϊνικής διαμόρφωσης υπό κυτταρικές συνθήκες 1.6. Ποικιλία πρωτεϊνών. Οικογένειες πρωτεϊνών χρησιμοποιώντας το παράδειγμα των ανοσοσφαιρινών 1.7. Φυσικοχημικές ιδιότητες πρωτεϊνών και μέθοδοι διαχωρισμού τους |
Αρθρωτή ενότητα 1 ΔΟΜΙΚΗ ΟΡΓΑΝΩΣΗ ΤΩΝ ΜΟΝΟΜΕΡΩΝ ΠΡΩΤΕΪΝΩΝ ΚΑΙ ΟΙ ΒΑΣΙΚΕΣ ΤΗΣ ΛΕΙΤΟΥΡΓΙΑΣ ΤΟΥΣ
Μαθησιακοί στόχοι Να είναι σε θέση:
1. Χρησιμοποιήστε τη γνώση σχετικά με τα δομικά χαρακτηριστικά των πρωτεϊνών και την εξάρτηση των πρωτεϊνικών λειτουργιών από τη δομή τους για να κατανοήσετε τους μηχανισμούς ανάπτυξης κληρονομικών και επίκτητων πρωτεϊνοπαθειών.
2. Εξηγήστε τους μηχανισμούς της θεραπευτικής δράσης ορισμένων φαρμάκων ως συνδετών που αλληλεπιδρούν με τις πρωτεΐνες και αλλάζουν τη δραστηριότητά τους.
3. Χρησιμοποιήστε τη γνώση σχετικά με τη δομή και τη διαμορφωτική αστάθεια των πρωτεϊνών για να κατανοήσετε τη δομική και λειτουργική αστάθεια και την τάση τους για μετουσίωση υπό μεταβαλλόμενες συνθήκες.
4. Εξηγήστε τη χρήση μετουσιωτικών παραγόντων ως μέσων για την αποστείρωση ιατρικών υλικών και εργαλείων, καθώς και ως αντισηπτικών.
Ξέρω:
1. Επίπεδα δομικής οργάνωσης πρωτεϊνών.
2. Η σημασία της πρωτογενούς δομής των πρωτεϊνών, η οποία καθορίζει τη δομική και λειτουργική ποικιλομορφία τους.
3. Ο μηχανισμός σχηματισμού του ενεργού κέντρου στις πρωτεΐνες και η ειδική αλληλεπίδρασή του με τον συνδέτη, που αποτελεί τη βάση της λειτουργίας των πρωτεϊνών.
4. Παραδείγματα της επίδρασης εξωγενών προσδεμάτων (φάρμακα, τοξίνες, δηλητήρια) στη διαμόρφωση και λειτουργική δραστηριότητα των πρωτεϊνών.
5. Αιτίες και συνέπειες μετουσίωσης πρωτεΐνης, παράγοντες που προκαλούν μετουσίωση.
6. Παραδείγματα χρήσης μετουσιωτικών παραγόντων στην ιατρική ως αντισηπτικά και μέσα για την αποστείρωση ιατρικών εργαλείων.
ΘΕΜΑ 1.1. ΔΟΜΙΚΗ ΟΡΓΑΝΩΣΗ ΠΡΩΤΕΪΝΩΝ. ΣΤΑΔΙΑ ΔΙΑΜΟΡΦΩΣΗΣ ΜΗΤΘΕΝΤΩΝ
ΠΡΩΤΕΪΝΕΣ ΔΙΑΜΟΡΦΩΣΕΙΣ
Οι πρωτεΐνες είναι πολυμερή μόρια των οποίων τα μονομερή είναι μόνο 20 α-αμινοξέα. Το σύνολο και η σειρά συνδυασμού των αμινοξέων σε μια πρωτεΐνη καθορίζεται από τη δομή των γονιδίων στο DNA των ατόμων. Κάθε πρωτεΐνη, σύμφωνα με τη συγκεκριμένη δομή της, επιτελεί τη δική της λειτουργία. Το σύνολο των πρωτεϊνών ενός δεδομένου οργανισμού καθορίζει τα φαινοτυπικά χαρακτηριστικά του, καθώς και την παρουσία κληρονομικών ασθενειών ή μια προδιάθεση για την ανάπτυξή τους.
1. Αμινοξέα που συνθέτουν τις πρωτεΐνες. Πεπτιδικός δεσμός.Οι πρωτεΐνες είναι πολυμερή κατασκευασμένα από μονομερή - 20 α-αμινοξέα, ο γενικός τύπος των οποίων είναι
Τα αμινοξέα διαφέρουν ως προς τη δομή, το μέγεθος και τις φυσικοχημικές ιδιότητες των ριζών που συνδέονται με το άτομο α-άνθρακα. Οι λειτουργικές ομάδες αμινοξέων καθορίζουν τα χαρακτηριστικά των ιδιοτήτων των διαφορετικών α-αμινοξέων. Οι ρίζες που βρίσκονται στα α-αμινοξέα μπορούν να χωριστούν σε διάφορες ομάδες:
Προλίνη,Σε αντίθεση με τα άλλα 19 μονομερή πρωτεΐνης, δεν είναι αμινοξύ, αλλά ιμινοξύ· η ρίζα στην προλίνη σχετίζεται τόσο με το άτομο α-άνθρακα όσο και με την ιμινο ομάδα
 Τα αμινοξέα ποικίλλουν ως προς τη διαλυτότητα στο νερό.Αυτό οφείλεται στην ικανότητα των ριζών να αλληλεπιδρούν με το νερό (ένυδρο).
Τα αμινοξέα ποικίλλουν ως προς τη διαλυτότητα στο νερό.Αυτό οφείλεται στην ικανότητα των ριζών να αλληλεπιδρούν με το νερό (ένυδρο).
ΠΡΟΣ ΤΗΝ υδρόφιλοςπεριλαμβάνουν ρίζες που περιέχουν ανιονικές, κατιονικές και πολικές αφόρτιστες λειτουργικές ομάδες.
ΠΡΟΣ ΤΗΝ υδροφόβοςπεριλαμβάνουν ρίζες που περιέχουν μεθυλομάδες, αλειφατικές αλυσίδες ή δακτυλίους.
2. Οι πεπτιδικοί δεσμοί συνδέουν αμινοξέα για να σχηματίσουν πεπτίδια.Κατά τη διάρκεια της σύνθεσης πεπτιδίων, η α-καρβοξυλική ομάδα ενός αμινοξέος αλληλεπιδρά με την α-αμινο ομάδα ενός άλλου αμινοξέος για να σχηματιστεί πεπτιδικός δεσμός:
Οι πρωτεΐνες είναι πολυπεπτίδια, δηλ. γραμμικά πολυμερή α-αμινοξέων που συνδέονται με πεπτιδικό δεσμό (Εικ. 1.1.)
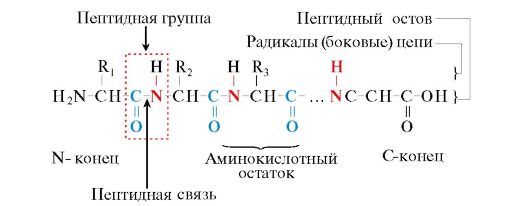 Ρύζι. 1.1. Όροι που χρησιμοποιούνται για την περιγραφή της δομής των πεπτιδίων
Ρύζι. 1.1. Όροι που χρησιμοποιούνται για την περιγραφή της δομής των πεπτιδίων
Τα μονομερή των αμινοξέων που αποτελούν τα πολυπεπτίδια ονομάζονται υπολείμματα αμινοξέων.Μια αλυσίδα επαναλαμβανόμενων ομάδων - NH-CH-CO- έντυπα πεπτιδική ραχοκοκαλιά.Ένα υπόλειμμα αμινοξέος που έχει μια ελεύθερη α-αμινο ομάδα ονομάζεται Ν-τελική και ένα που έχει μια ελεύθερη α-καρβοξυλική ομάδα ονομάζεται C-τελική. Τα πεπτίδια γράφονται και διαβάζονται από το Ν-άκρο στο C-άκρο.
Ο πεπτιδικός δεσμός που σχηματίζεται από την ιμινο ομάδα της προλίνης διαφέρει από άλλους πεπτιδικούς δεσμούς: το άτομο αζώτου της πεπτιδικής ομάδας στερείται υδρογόνου,
Αντίθετα, υπάρχει ένας δεσμός με μια ρίζα, ως αποτέλεσμα του οποίου η μία πλευρά του δακτυλίου περιλαμβάνεται στη ραχοκοκαλιά του πεπτιδίου:
 Τα πεπτίδια διαφέρουν ως προς τη σύνθεση αμινοξέων, τον αριθμό των αμινοξέων και τη σειρά σύνδεσης αμινοξέων, για παράδειγμα, τα Ser-Ala-Glu-Gis και His-Glu-Ala-Ser είναι δύο διαφορετικά πεπτίδια.
Τα πεπτίδια διαφέρουν ως προς τη σύνθεση αμινοξέων, τον αριθμό των αμινοξέων και τη σειρά σύνδεσης αμινοξέων, για παράδειγμα, τα Ser-Ala-Glu-Gis και His-Glu-Ala-Ser είναι δύο διαφορετικά πεπτίδια.
Οι πεπτιδικοί δεσμοί είναι πολύ ισχυροί και η χημική τους μη ενζυματική υδρόλυση απαιτεί σκληρές συνθήκες: η πρωτεΐνη που αναλύεται υδρολύεται σε πυκνό υδροχλωρικό οξύ σε θερμοκρασία περίπου 110° για 24 ώρες. Σε ένα ζωντανό κύτταρο, οι πεπτιδικοί δεσμοί μπορούν να σπάσουν πρωτεολυτικά ένζυμα,που ονομάζεται πρωτεάσεςή πεπτιδικές υδρολάσες.
3. Πρωτογενής δομή πρωτεϊνών.Τα υπολείμματα αμινοξέων στις πεπτιδικές αλυσίδες διαφορετικών πρωτεϊνών δεν εναλλάσσονται τυχαία, αλλά είναι διατεταγμένα με μια ορισμένη σειρά. Η γραμμική αλληλουχία ή σειρά εναλλαγής των υπολειμμάτων αμινοξέων σε μια πολυπεπτιδική αλυσίδα ονομάζεται πρωτογενής δομή της πρωτεΐνης.
Η πρωτογενής δομή κάθε μεμονωμένης πρωτεΐνης κωδικοποιείται στο μόριο DNA (σε μια περιοχή που ονομάζεται γονίδιο) και πραγματοποιείται κατά τη μεταγραφή (αντιγραφή πληροφοριών σε mRNA) και τη μετάφραση (σύνθεση της πρωτογενούς δομής της πρωτεΐνης). Κατά συνέπεια, η πρωταρχική δομή των πρωτεϊνών ενός μεμονωμένου ατόμου είναι πληροφορίες που μεταδίδονται κληρονομικά από τους γονείς στα παιδιά, οι οποίες καθορίζουν τα δομικά χαρακτηριστικά των πρωτεϊνών ενός δεδομένου οργανισμού, από τα οποία εξαρτάται η λειτουργία των υπαρχουσών πρωτεϊνών (Εικ. 1.2.).
 Ρύζι. 1.2. Η σχέση μεταξύ του γονότυπου και της διαμόρφωσης των πρωτεϊνών που συντίθενται στο σώμα του ατόμου
Ρύζι. 1.2. Η σχέση μεταξύ του γονότυπου και της διαμόρφωσης των πρωτεϊνών που συντίθενται στο σώμα του ατόμου
Κάθε μία από τις περίπου 100.000 μεμονωμένες πρωτεΐνες στο ανθρώπινο σώμα έχει μοναδικόςπρωτογενής δομή. Τα μόρια του ίδιου τύπου πρωτεΐνης (για παράδειγμα, λευκωματίνη) έχουν την ίδια εναλλαγή υπολειμμάτων αμινοξέων, γεγονός που διακρίνει τη λευκωματίνη από οποιαδήποτε άλλη μεμονωμένη πρωτεΐνη.
Η αλληλουχία των υπολειμμάτων αμινοξέων σε μια πεπτιδική αλυσίδα μπορεί να θεωρηθεί μια μορφή καταγραφής πληροφοριών. Αυτές οι πληροφορίες καθορίζουν τη χωρική διάταξη της γραμμικής πεπτιδικής αλυσίδας σε μια πιο συμπαγή τρισδιάστατη δομή που ονομάζεται διαμόρφωσησκίουρος. Η διαδικασία σχηματισμού μιας λειτουργικά ενεργής πρωτεϊνικής διαμόρφωσης ονομάζεται πτυσσόμενος
4. Διαμόρφωση πρωτεΐνης.Η ελεύθερη περιστροφή στη ραχοκοκαλιά του πεπτιδίου είναι δυνατή μεταξύ του ατόμου αζώτου της πεπτιδικής ομάδας και του γειτονικού ατόμου α-άνθρακα, καθώς και μεταξύ του α-ατόμου άνθρακα και του άνθρακα της ομάδας καρβονυλίου. Λόγω της αλληλεπίδρασης λειτουργικών ομάδων υπολειμμάτων αμινοξέων, η πρωτογενής δομή των πρωτεϊνών μπορεί να αποκτήσει πιο πολύπλοκες χωρικές δομές. Στις σφαιρικές πρωτεΐνες, υπάρχουν δύο κύρια επίπεδα αναδίπλωσης της διαμόρφωσης των πεπτιδικών αλυσίδων: δευτερεύωνΚαι τριτογενής δομή.
Δευτερογενής δομή πρωτεϊνώνείναι μια χωρική δομή που σχηματίζεται ως αποτέλεσμα του σχηματισμού δεσμών υδρογόνου μεταξύ των λειτουργικών ομάδων -C=O και -NH- της πεπτιδικής ραχοκοκαλιάς. Σε αυτή την περίπτωση, η πεπτιδική αλυσίδα μπορεί να αποκτήσει κανονικές δομές δύο τύπων: α-έλικεςΚαι β-δομές.
ΣΕ α-έλικεςΣχηματίζονται δεσμοί υδρογόνου μεταξύ του ατόμου οξυγόνου της καρβονυλικής ομάδας και του υδρογόνου του αμιδικού αζώτου του 4ου αμινοξέος από αυτό. πλευρικές αλυσίδες υπολειμμάτων αμινοξέων
βρίσκονται κατά μήκος της περιφέρειας της σπείρας, χωρίς να συμμετέχουν στο σχηματισμό της δευτερεύουσας δομής (Εικ. 1.3.).
Οι μαζικές ρίζες, ή ρίζες που φέρουν ίσα φορτία, εμποδίζουν το σχηματισμό μιας α-έλικας. Το υπόλειμμα προλίνης, που έχει δομή δακτυλίου, διακόπτει την α-έλικα, αφού λόγω της έλλειψης υδρογόνου στο άτομο αζώτου στην πεπτιδική αλυσίδα είναι αδύνατο να σχηματιστεί δεσμός υδρογόνου. Ο δεσμός μεταξύ αζώτου και ατόμου άνθρακα είναι μέρος του δακτυλίου προλίνης, έτσι η ραχοκοκαλιά του πεπτιδίου κάμπτεται σε αυτό το σημείο.
β-Δομήσχηματίζεται μεταξύ των γραμμικών περιοχών της πεπτιδικής ραχοκοκαλιάς μιας πολυπεπτιδικής αλυσίδας, σχηματίζοντας έτσι διπλωμένες δομές. Μπορούν να σχηματιστούν πολυπεπτιδικές αλυσίδες ή μέρη τους παράλληλοή αντιπαράλληλες β-δομές.Στην πρώτη περίπτωση, τα Ν- και C-άκρα των αλληλεπιδρώντων πεπτιδικών αλυσίδων συμπίπτουν και στη δεύτερη έχουν την αντίθετη κατεύθυνση (Εικ. 1.4).
Ρύζι. 1.3. Δευτερεύουσα δομή πρωτεΐνης - α-έλικα
 Ρύζι. 1.4. Παράλληλες και αντιπαράλληλες δομές β-φύλλων
Ρύζι. 1.4. Παράλληλες και αντιπαράλληλες δομές β-φύλλων
Οι β-δομές υποδεικνύονται με φαρδιά βέλη: A - Αντιπαράλληλη β-δομή. Β - Παράλληλες δομές β-φύλλων
Σε ορισμένες πρωτεΐνες, οι β-δομές μπορούν να σχηματιστούν λόγω του σχηματισμού δεσμών υδρογόνου μεταξύ ατόμων της πεπτιδικής ραχοκοκαλιάς διαφορετικών πολυπεπτιδικών αλυσίδων.
Βρίσκεται επίσης σε πρωτεΐνες περιοχές με ακανόνιστες δευτερεύουσεςδομή, η οποία περιλαμβάνει κάμψεις, βρόχους και στροφές της ραχοκοκαλιάς του πολυπεπτιδίου. Συχνά βρίσκονται σε μέρη όπου αλλάζει η κατεύθυνση της πεπτιδικής αλυσίδας, για παράδειγμα, όταν σχηματίζεται μια παράλληλη δομή β-φύλλου.
Με βάση την παρουσία α-έλικων και β-δομών, οι σφαιρικές πρωτεΐνες μπορούν να χωριστούν σε τέσσερις κατηγορίες.
Ρύζι. 1.5. Δευτερεύουσα δομή της μυοσφαιρίνης (Α) και της β-αλυσίδας της αιμοσφαιρίνης (Β), που περιέχει οκτώ α-έλικες

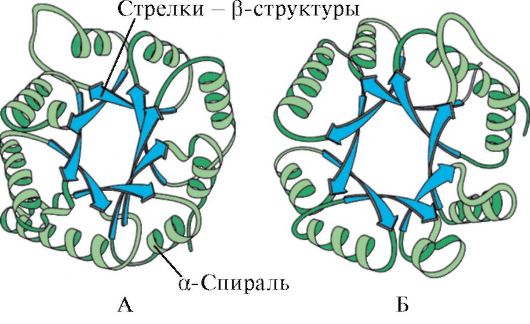 Ρύζι. 1.6. Δευτερογενής δομή της ισομεράσης της τριοσοφωσφορικής και της περιοχής πυροσταφυλικής κινάσης
Ρύζι. 1.6. Δευτερογενής δομή της ισομεράσης της τριοσοφωσφορικής και της περιοχής πυροσταφυλικής κινάσης
 Ρύζι. 1.7. Δευτερογενής δομή της σταθερής περιοχής της ανοσοσφαιρίνης (Α) και του ενζύμου υπεροξειδίου δισμουτάση (Β)
Ρύζι. 1.7. Δευτερογενής δομή της σταθερής περιοχής της ανοσοσφαιρίνης (Α) και του ενζύμου υπεροξειδίου δισμουτάση (Β)
ΣΕ τέταρτη κατηγορίαπεριελάμβανε πρωτεΐνες που περιέχουν μια μικρή ποσότητα κανονικών δευτερογενών δομών. Αυτές οι πρωτεΐνες περιλαμβάνουν μικρές πρωτεΐνες πλούσιες σε κυστεΐνη ή μεταλλοπρωτεΐνες.
Τριτοταγής δομή πρωτεΐνης- ένας τύπος διαμόρφωσης που σχηματίζεται λόγω αλληλεπιδράσεων μεταξύ ριζών αμινοξέων, οι οποίες μπορούν να βρίσκονται σε σημαντική απόσταση μεταξύ τους στην πεπτιδική αλυσίδα. Οι περισσότερες πρωτεΐνες σχηματίζουν μια χωρική δομή που μοιάζει με σφαιρίδιο (σφαιρικές πρωτεΐνες).
Δεδομένου ότι οι υδρόφοβες ρίζες αμινοξέων τείνουν να συνδυάζονται μέσω των λεγόμενων υδρόφοβες αλληλεπιδράσειςκαι διαμοριακές δυνάμεις van der Waals, σχηματίζεται ένας πυκνός υδρόφοβος πυρήνας μέσα στο πρωτεϊνικό σφαιρίδιο. Οι υδρόφιλες ιονισμένες και μη ιονισμένες ρίζες βρίσκονται κυρίως στην επιφάνεια της πρωτεΐνης και καθορίζουν τη διαλυτότητά της στο νερό.
 Ρύζι. 1.8. Τύποι δεσμών που προκύπτουν μεταξύ των ριζών αμινοξέων κατά τον σχηματισμό της τριτοταγούς δομής μιας πρωτεΐνης
Ρύζι. 1.8. Τύποι δεσμών που προκύπτουν μεταξύ των ριζών αμινοξέων κατά τον σχηματισμό της τριτοταγούς δομής μιας πρωτεΐνης
1 - ιοντικός δεσμός- εμφανίζεται μεταξύ θετικά και αρνητικά φορτισμένες λειτουργικές ομάδες.
2 - δεσμός υδρογόνου- εμφανίζεται μεταξύ μιας υδρόφιλης μη φορτισμένης ομάδας και οποιασδήποτε άλλης υδρόφιλης ομάδας.
3 - υδρόφοβες αλληλεπιδράσεις- προκύπτουν μεταξύ υδρόφοβων ριζών.
4 - δισουλφιδικός δεσμός- σχηματίζεται λόγω της οξείδωσης των ομάδων SH υπολειμμάτων κυστεΐνης και της αλληλεπίδρασής τους μεταξύ τους
Τα υδρόφιλα υπολείμματα αμινοξέων που βρίσκονται μέσα στον υδρόφοβο πυρήνα μπορούν να αλληλεπιδράσουν μεταξύ τους χρησιμοποιώντας ιωνικόςΚαι δεσμούς υδρογόνου(Εικ. 1.8).
Οι ιοντικοί δεσμοί και οι δεσμοί υδρογόνου, καθώς και οι υδρόφοβες αλληλεπιδράσεις, είναι ασθενείς: η ενέργειά τους δεν είναι πολύ μεγαλύτερη από την ενέργεια της θερμικής κίνησης των μορίων σε θερμοκρασία δωματίου. Η διαμόρφωση της πρωτεΐνης διατηρείται με το σχηματισμό πολλών τέτοιων αδύναμων δεσμών. Δεδομένου ότι τα άτομα που συνθέτουν μια πρωτεΐνη βρίσκονται σε συνεχή κίνηση, είναι δυνατό να σπάσουν ορισμένοι ασθενείς δεσμοί και να σχηματιστούν άλλοι, γεγονός που οδηγεί σε ελαφρές μετακινήσεις μεμονωμένων τμημάτων της πολυπεπτιδικής αλυσίδας. Αυτή η ιδιότητα των πρωτεϊνών να αλλάζουν τη διαμόρφωση ως αποτέλεσμα της διάσπασης ορισμένων και του σχηματισμού άλλων αδύναμων δεσμών ονομάζεται διαμορφωτική αστάθεια.
Το ανθρώπινο σώμα έχει συστήματα που υποστηρίζουν ομοιοσταση- συνέπεια εσωτερικό περιβάλλονεντός ορισμένων αποδεκτών ορίων για ένα υγιές σώμα. Υπό συνθήκες ομοιόστασης, μικρές αλλαγές στη διαμόρφωση δεν διαταράσσουν τη συνολική δομή και λειτουργία των πρωτεϊνών. Η λειτουργικά ενεργή διαμόρφωση μιας πρωτεΐνης ονομάζεται εγγενής διαμόρφωση.Αλλαγές στο εσωτερικό περιβάλλον (για παράδειγμα, η συγκέντρωση γλυκόζης, ιόντων Ca, πρωτονίων κ.λπ.) οδηγούν σε αλλαγές στη διαμόρφωση και διαταραχή των πρωτεϊνικών λειτουργιών.
Η τριτοταγής δομή ορισμένων πρωτεϊνών σταθεροποιείται δισουλφιδικούς δεσμούς,που σχηματίζεται λόγω της αλληλεπίδρασης των ομάδων -SH δύο υπολειμμάτων
 Ρύζι. 1.9. Σχηματισμός δισουλφιδικού δεσμού σε μόριο πρωτεΐνης
Ρύζι. 1.9. Σχηματισμός δισουλφιδικού δεσμού σε μόριο πρωτεΐνης
κυστεΐνη (Εικ. 1.9). Οι περισσότερες ενδοκυτταρικές πρωτεΐνες δεν έχουν ομοιοπολικούς δισουλφιδικούς δεσμούς στην τριτοταγή δομή τους. Η παρουσία τους είναι χαρακτηριστική των πρωτεϊνών που εκκρίνονται από το κύτταρο, γεγονός που εξασφαλίζει τη μεγαλύτερη σταθερότητά τους σε εξωκυτταρικές συνθήκες. Έτσι, υπάρχουν δισουλφιδικοί δεσμοί στα μόρια της ινσουλίνης και των ανοσοσφαιρινών.
Ινσουλίνη- μια πρωτεϊνική ορμόνη που συντίθεται στα β-κύτταρα του παγκρέατος και εκκρίνεται στο αίμα ως απόκριση σε αύξηση της συγκέντρωσης γλυκόζης στο αίμα. Στη δομή της ινσουλίνης, υπάρχουν δύο δισουλφιδικοί δεσμοί που συνδέουν τις πολυπεπτιδικές αλυσίδες Α και Β και ένας δισουλφιδικός δεσμός εντός της αλυσίδας Α (Εικ. 1.10).
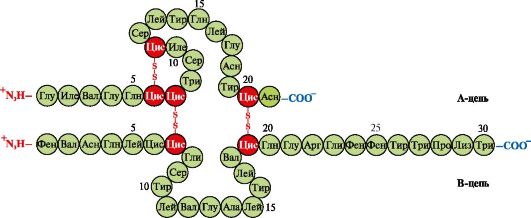 Ρύζι. 1.10. Δισουλφιδικοί δεσμοί στη δομή της ινσουλίνης
Ρύζι. 1.10. Δισουλφιδικοί δεσμοί στη δομή της ινσουλίνης
5. Υπερδευτερογενής δομή πρωτεϊνών.Σε πρωτεΐνες με διαφορετική πρωτογενή δομή και λειτουργίες, μερικές φορές ανιχνεύονται παρόμοιοι συνδυασμοί και σχετικές θέσεις δευτερευουσών δομών,που ονομάζονται υπερδευτερεύουσα δομή. Καταλαμβάνει μια ενδιάμεση θέση μεταξύ των δευτερογενών και τριτογενών δομών, αφού είναι ένας συγκεκριμένος συνδυασμός στοιχείων της δευτερογενούς δομής στο σχηματισμό της τριτοταγούς δομής της πρωτεΐνης. Οι υπερδευτερεύουσες δομές έχουν συγκεκριμένα ονόματα, όπως «α-έλικα-turn-a-helix», «φερμουάρ λευκίνης», «δάκτυλα ψευδαργύρου» κ.λπ. Τέτοιες υπερδευτερεύουσες δομές είναι χαρακτηριστικές των πρωτεϊνών που δεσμεύουν το DNA.
"Φερμουάρ λευκίνης."Αυτός ο τύπος υπερδευτερογενούς δομής χρησιμοποιείται για την ένωση δύο πρωτεϊνών μεταξύ τους. Στην επιφάνεια των αλληλεπιδρώντων πρωτεϊνών υπάρχουν α-έλικες περιοχές που περιέχουν τουλάχιστον τέσσερα υπολείμματα λευκίνης. Τα υπολείμματα λευκίνης στην α-έλικα βρίσκονται σε απόσταση έξι αμινοξέων μεταξύ τους. Δεδομένου ότι κάθε στροφή της α-έλικας περιέχει 3,6 υπολείμματα αμινοξέων, οι ρίζες λευκίνης βρίσκονται στην επιφάνεια κάθε δεύτερης στροφής. Τα υπολείμματα λευκίνης της α-έλικας μιας πρωτεΐνης μπορούν να αλληλεπιδράσουν με υπολείμματα λευκίνης μιας άλλης πρωτεΐνης (υδρόφοβες αλληλεπιδράσεις), συνδέοντάς τα μεταξύ τους (Εικ. 1.11.). Πολλές πρωτεΐνες που δεσμεύουν το DNA λειτουργούν σε ολιγομερή σύμπλοκα όπου οι μεμονωμένες υπομονάδες συνδέονται μεταξύ τους με «φερμουάρ λευκίνης».
 Ρύζι. 1.11. "Φερμουάρ λευκίνης" μεταξύ α-ελικοειδών περιοχών δύο πρωτεϊνών
Ρύζι. 1.11. "Φερμουάρ λευκίνης" μεταξύ α-ελικοειδών περιοχών δύο πρωτεϊνών
Ένα παράδειγμα τέτοιων πρωτεϊνών είναι οι ιστόνες. Ιστόνες- πυρηνικές πρωτεΐνες, οι οποίες περιλαμβάνουν ένας μεγάλος αριθμός απόθετικά φορτισμένα αμινοξέα - αργινίνη και λυσίνη (έως 80%). Τα μόρια ιστόνης συνδυάζονται σε ολιγομερή σύμπλοκα που περιέχουν οκτώ μονομερή χρησιμοποιώντας «φερμουάρ λευκίνης», παρά το σημαντικό ομώνυμο φορτίο αυτών των μορίων.
"Δάχτυλο ψευδάργυρου"- μια παραλλαγή υπερδευτερογενούς δομής, χαρακτηριστική των πρωτεϊνών που δεσμεύουν το DNA, έχει τη μορφή επιμήκους θραύσματος στην επιφάνεια της πρωτεΐνης και περιέχει περίπου 20 υπολείμματα αμινοξέων (Εικ. 1.12). Το σχήμα του «εκτεταμένου δακτύλου» υποστηρίζεται από ένα άτομο ψευδαργύρου συνδεδεμένο με τέσσερις ρίζες αμινοξέων - δύο υπολείμματα κυστεΐνης και δύο υπολείμματα ιστιδίνης. Σε ορισμένες περιπτώσεις, αντί για υπολείμματα ιστιδίνης, υπάρχουν υπολείμματα κυστεΐνης. Δύο υπολείμματα κυστεΐνης που βρίσκονται κοντά διαχωρίζονται από τα άλλα δύο υπολείμματα Gisili με μια αλληλουχία Cys που αποτελείται από περίπου 12 υπολείμματα αμινοξέων. Αυτή η περιοχή της πρωτεΐνης σχηματίζει μια α-έλικα, οι ρίζες της οποίας μπορούν να συνδεθούν ειδικά με τις ρυθμιστικές περιοχές της κύριας αύλακας του DNA. Ατομική εξειδίκευση δέσμευσης
 Ρύζι. 1.12. Η πρωταρχική δομή της περιοχής των πρωτεϊνών που δεσμεύουν το DNA που σχηματίζουν τη δομή του «δαχτύλου ψευδαργύρου» (τα γράμματα υποδεικνύουν τα αμινοξέα που συνθέτουν αυτή τη δομή)
Ρύζι. 1.12. Η πρωταρχική δομή της περιοχής των πρωτεϊνών που δεσμεύουν το DNA που σχηματίζουν τη δομή του «δαχτύλου ψευδαργύρου» (τα γράμματα υποδεικνύουν τα αμινοξέα που συνθέτουν αυτή τη δομή)
Η ρυθμιστική πρωτεΐνη που δεσμεύει το DNA εξαρτάται από την αλληλουχία των υπολειμμάτων αμινοξέων που βρίσκονται στην περιοχή του δακτύλου ψευδαργύρου. Τέτοιες δομές περιέχουν, συγκεκριμένα, υποδοχείς για στεροειδείς ορμόνες που εμπλέκονται στη ρύθμιση της μεταγραφής (ανάγνωση πληροφοριών από DNA σε RNA).
ΘΕΜΑ 1.2. ΒΑΣΕΙΣ ΤΗΣ ΛΕΙΤΟΥΡΓΙΑΣ ΠΡΩΤΕΪΝΩΝ. ΤΑ ΦΑΡΜΑΚΑ ΩΣ ΠΡΟΣΔΕΣΤΕΣ ΠΟΥ ΕΠΗΡΕΑΖΟΥΝ ΤΗ ΛΕΙΤΟΥΡΓΙΑ ΤΩΝ ΠΡΩΤΕΪΝΩΝ
1. Το ενεργό κέντρο της πρωτεΐνης και η αλληλεπίδρασή της με τον συνδέτη.Κατά τον σχηματισμό της τριτοταγούς δομής, σχηματίζεται μια περιοχή στην επιφάνεια μιας λειτουργικά ενεργής πρωτεΐνης, συνήθως σε μια εσοχή, που σχηματίζεται από ρίζες αμινοξέων που βρίσκονται πολύ μακριά η μία από την άλλη στην πρωτογενή δομή. Αυτή η περιοχή, η οποία έχει μια μοναδική δομή για μια δεδομένη πρωτεΐνη και είναι ικανή να αλληλεπιδρά ειδικά με ένα συγκεκριμένο μόριο ή ομάδα παρόμοιων μορίων, ονομάζεται θέση σύνδεσης πρωτεΐνης-συνδέτη ή ενεργή θέση. Οι συνδέτες είναι μόρια που αλληλεπιδρούν με πρωτεΐνες.
Υψηλή ειδικότηταΗ αλληλεπίδραση της πρωτεΐνης με τον συνδέτη εξασφαλίζεται από τη συμπληρωματικότητα της δομής του ενεργού κέντρου προς τη δομή του συνδέτη.
Συμπληρωματικότητα- αυτή είναι η χωρική και χημική αντιστοιχία των επιφανειών που αλληλεπιδρούν. Το ενεργό κέντρο δεν πρέπει μόνο χωρικά να αντιστοιχεί στον συνδέτη που περιλαμβάνεται σε αυτό, αλλά και δεσμοί (ιοντικές, υδρογόνου και υδρόφοβες αλληλεπιδράσεις) πρέπει να σχηματίζονται μεταξύ των λειτουργικών ομάδων των ριζών που περιλαμβάνονται στο ενεργό κέντρο και του συνδέτη που συγκρατούν τον συνδέτη. στο ενεργό κέντρο (Εικ. 1.13 ).
 Ρύζι. 1.13. Συμπληρωματική αλληλεπίδραση πρωτεΐνης με συνδέτη
Ρύζι. 1.13. Συμπληρωματική αλληλεπίδραση πρωτεΐνης με συνδέτη
Μερικοί συνδέτες, όταν συνδέονται με το ενεργό κέντρο μιας πρωτεΐνης, παίζουν βοηθητικό ρόλο στη λειτουργία των πρωτεϊνών. Τέτοιοι συνδετήρες ονομάζονται συμπαράγοντες και οι πρωτεΐνες που περιέχουν ένα μη πρωτεϊνικό μέρος ονομάζονται σύνθετες πρωτεΐνες(σε αντίθεση με τις απλές πρωτεΐνες, που αποτελούνται μόνο από το πρωτεϊνικό μέρος). Το μη πρωτεϊνικό μέρος, σταθερά συνδεδεμένο με την πρωτεΐνη, ονομάζεται ομάδα προσθετικών.Για παράδειγμα, η μυοσφαιρίνη, η αιμοσφαιρίνη και τα κυτοχρώματα περιέχουν μια προσθετική ομάδα, την αίμη, που περιέχει ένα ιόν σιδήρου, σταθερά συνδεδεμένη με το ενεργό κέντρο. Οι σύνθετες πρωτεΐνες που περιέχουν αίμη ονομάζονται αιμοπρωτεΐνες.
Όταν ειδικοί συνδέτες συνδέονται με πρωτεΐνες, η λειτουργία αυτών των πρωτεϊνών εκδηλώνεται. Έτσι, η λευκωματίνη, η πιο σημαντική πρωτεΐνη στο πλάσμα του αίματος, επιδεικνύει τη λειτουργία μεταφοράς της συνδέοντας υδρόφοβους συνδέτες, όπως λιπαρά οξέα, χολερυθρίνη, ορισμένα φάρμακα κ.λπ. στο ενεργό κέντρο (Εικ. 1.14).
Οι συνδέτες που αλληλεπιδρούν με την τρισδιάστατη δομή της πεπτιδικής αλυσίδας μπορεί να είναι όχι μόνο οργανικά και ανόργανα μόρια χαμηλού μοριακού βάρους, αλλά και μακρομόρια:
DNA (παραδείγματα με πρωτεΐνες δέσμευσης DNA που συζητήθηκαν παραπάνω).
Πολυσακχαρίτες;
 Ρύζι. 1.14. Σχέση μεταξύ γονότυπου και φαινοτύπου
Ρύζι. 1.14. Σχέση μεταξύ γονότυπου και φαινοτύπου
Η μοναδική πρωτογενής δομή των ανθρώπινων πρωτεϊνών, που κωδικοποιείται στο μόριο DNA, πραγματοποιείται στα κύτταρα με τη μορφή μοναδικής διαμόρφωσης, δομής ενεργού κέντρου και πρωτεϊνικών λειτουργιών
Σε αυτές τις περιπτώσεις, η πρωτεΐνη αναγνωρίζει μια συγκεκριμένη περιοχή του συνδέτη που είναι ανάλογη και συμπληρωματική προς τη θέση δέσμευσης. Έτσι, στην επιφάνεια των ηπατοκυττάρων υπάρχουν πρωτεΐνες υποδοχείς για την ορμόνη ινσουλίνη, η οποία έχει επίσης δομή πρωτεΐνης. Η αλληλεπίδραση της ινσουλίνης με τον υποδοχέα προκαλεί αλλαγή στη διαμόρφωση της και ενεργοποίηση των συστημάτων σηματοδότησης, οδηγώντας στην αποθήκευση θρεπτικών ουσιών στα ηπατοκύτταρα μετά τα γεύματα.
Ετσι, Η λειτουργία των πρωτεϊνών βασίζεται στην ειδική αλληλεπίδραση του ενεργού κέντρου της πρωτεΐνης με τον συνδέτη.
2. Δομή τομέα και ο ρόλος του στη λειτουργία των πρωτεϊνών.Οι μακριές πολυπεπτιδικές αλυσίδες σφαιρικών πρωτεϊνών συχνά αναδιπλώνονται σε αρκετές συμπαγείς, σχετικά ανεξάρτητες περιοχές. Έχουν ανεξάρτητη τριτογενή δομή, που θυμίζει αυτή των σφαιρικών πρωτεϊνών και ονομάζονται τομείς.Χάρη στη δομή του τομέα των πρωτεϊνών, η τριτοταγής δομή τους είναι πιο εύκολο να σχηματιστεί.
Στις πρωτεΐνες της περιοχής, οι θέσεις δέσμευσης συνδέτη βρίσκονται συχνά μεταξύ των περιοχών. Έτσι, η θρυψίνη είναι ένα πρωτεολυτικό ένζυμο που παράγεται από το εξωκρινές τμήμα του παγκρέατος και είναι απαραίτητο για την πέψη των πρωτεϊνών των τροφίμων. Έχει δομή δύο τομέων και το κέντρο δέσμευσης της θρυψίνης με τον συνδέτη της - την πρωτεΐνη τροφής - βρίσκεται στο αυλάκι μεταξύ των δύο περιοχών. Στο ενεργό κέντρο δημιουργούνται οι απαραίτητες συνθήκες για την αποτελεσματική σύνδεση μιας συγκεκριμένης θέσης πρωτεΐνης τροφής και την υδρόλυση των πεπτιδικών της δεσμών.
Διαφορετικές περιοχές σε μια πρωτεΐνη μπορούν να μετακινηθούν μεταξύ τους όταν το ενεργό κέντρο αλληλεπιδρά με τον συνδέτη (Εικ. 1.15).
Εξοκινάση- ένα ένζυμο που καταλύει τη φωσφορυλίωση της γλυκόζης χρησιμοποιώντας ATP. Η ενεργή θέση του ενζύμου βρίσκεται στη σχισμή μεταξύ των δύο περιοχών. Όταν η εξοκινάση συνδέεται με τη γλυκόζη, οι περιοχές που την περιβάλλουν κλείνουν και το υπόστρωμα παγιδεύεται, όπου συμβαίνει φωσφορυλίωση (βλ. Εικ. 1.15).
 Ρύζι. 1.15. Σύνδεση περιοχών εξοκινάσης με γλυκόζη
Ρύζι. 1.15. Σύνδεση περιοχών εξοκινάσης με γλυκόζη
Σε ορισμένες πρωτεΐνες, οι περιοχές εκτελούν ανεξάρτητες λειτουργίες με σύνδεση σε διάφορους συνδέτες. Τέτοιες πρωτεΐνες ονομάζονται πολυλειτουργικές.
3. Τα φάρμακα είναι συνδέτες που επηρεάζουν τη λειτουργία των πρωτεϊνών.Η αλληλεπίδραση των πρωτεϊνών με τους συνδέτες είναι ειδική. Ωστόσο, λόγω της διαμορφωτικής αστάθειας της πρωτεΐνης και του ενεργού της κέντρου, είναι δυνατό να επιλεγεί μια άλλη ουσία που θα μπορούσε επίσης να αλληλεπιδράσει με την πρωτεΐνη στο ενεργό κέντρο ή σε άλλο μέρος του μορίου.
Μια ουσία παρόμοια στη δομή με έναν φυσικό συνδέτη ονομάζεται δομικό ανάλογο του συνδέτηή ένα μη φυσικό πρόσδεμα. Αλληλεπιδρά επίσης με την πρωτεΐνη στο ενεργό κέντρο. Ένα δομικό ανάλογο ενός συνδέτη μπορεί και τα δύο να ενισχύσει τη λειτουργία της πρωτεΐνης (αγωνιστής),και να το μειώσει (ανταγωνιστής).Ο συνδέτης και τα δομικά του ανάλογα ανταγωνίζονται μεταξύ τους για σύνδεση με την πρωτεΐνη στην ίδια θέση. Τέτοιες ουσίες ονομάζονται ανταγωνιστικοί διαμορφωτές(ρυθμιστές) των πρωτεϊνικών λειτουργιών. Πολλά φάρμακα δρουν ως αναστολείς πρωτεΐνης. Μερικά από αυτά λαμβάνονται με χημική τροποποίηση φυσικών προσδεμάτων. Αναστολείς των πρωτεϊνικών λειτουργιών μπορεί να είναι φάρμακα και δηλητήρια.
Η ατροπίνη είναι ένας ανταγωνιστικός αναστολέας των Μ-χολινεργικών υποδοχέων.Η ακετυλοχολίνη είναι ένας νευροδιαβιβαστής για τη μετάδοση των νευρικών ερεθισμάτων μέσω των χολινεργικών συνάψεων. Για να πραγματοποιηθεί η διέγερση, η ακετυλοχολίνη που απελευθερώνεται στη συναπτική σχισμή πρέπει να αλληλεπιδράσει με την πρωτεΐνη υποδοχέα της μετασυναπτικής μεμβράνης. Βρέθηκαν δύο τύποι χολινεργικοί υποδοχείς:
Μ υποδοχέαςεκτός από την ακετυλοχολίνη, αλληλεπιδρά επιλεκτικά με τη μουσκαρίνη (τοξίνη μύγας). Μ - χολινεργικοί υποδοχείς υπάρχουν στους λείους μύες και, όταν αλληλεπιδρούν με την ακετυλοχολίνη, προκαλούν τη συστολή τους.
Η υποδοχέαςδεσμεύεται ειδικά με τη νικοτίνη. Οι Ν-χολινεργικοί υποδοχείς βρίσκονται στις συνάψεις των γραμμωτών σκελετικών μυών.
Ειδικός αναστολέας Μ-χολινεργικοί υποδοχείςείναι ατροπίνη. Βρίσκεται στα φυτά της μπελαντόνας και της χένμπας.
 Η ατροπίνη έχει λειτουργικές ομάδες παρόμοιες στη δομή με την ακετυλοχολίνη και τη χωρική τους διάταξη, επομένως είναι ένας ανταγωνιστικός αναστολέας των Μ-χολινεργικών υποδοχέων. Λαμβάνοντας υπόψη ότι η σύνδεση της ακετυλοχολίνης με τους Μ-χολινεργικούς υποδοχείς προκαλεί συστολή των λείων μυών, η ατροπίνη χρησιμοποιείται ως φάρμακο που ανακουφίζει από τον σπασμό τους. (αντισπασμωδικό).Έτσι, είναι γνωστή η χρήση της ατροπίνης για τη χαλάρωση των μυών των ματιών κατά την εξέταση του βυθού, καθώς και για την ανακούφιση από σπασμούς κατά τη διάρκεια του γαστρεντερικού κολικού. Μ-χολινεργικοί υποδοχείς υπάρχουν επίσης στο κεντρικό νευρικό σύστημα(ΚΝΣ), επομένως μεγάλες δόσεις ατροπίνης μπορεί να προκαλέσουν ανεπιθύμητη αντίδραση από το κεντρικό νευρικό σύστημα: κινητική και νοητική διέγερση, παραισθήσεις, σπασμούς.
Η ατροπίνη έχει λειτουργικές ομάδες παρόμοιες στη δομή με την ακετυλοχολίνη και τη χωρική τους διάταξη, επομένως είναι ένας ανταγωνιστικός αναστολέας των Μ-χολινεργικών υποδοχέων. Λαμβάνοντας υπόψη ότι η σύνδεση της ακετυλοχολίνης με τους Μ-χολινεργικούς υποδοχείς προκαλεί συστολή των λείων μυών, η ατροπίνη χρησιμοποιείται ως φάρμακο που ανακουφίζει από τον σπασμό τους. (αντισπασμωδικό).Έτσι, είναι γνωστή η χρήση της ατροπίνης για τη χαλάρωση των μυών των ματιών κατά την εξέταση του βυθού, καθώς και για την ανακούφιση από σπασμούς κατά τη διάρκεια του γαστρεντερικού κολικού. Μ-χολινεργικοί υποδοχείς υπάρχουν επίσης στο κεντρικό νευρικό σύστημα(ΚΝΣ), επομένως μεγάλες δόσεις ατροπίνης μπορεί να προκαλέσουν ανεπιθύμητη αντίδραση από το κεντρικό νευρικό σύστημα: κινητική και νοητική διέγερση, παραισθήσεις, σπασμούς.
Η διτιλίνη είναι ένας ανταγωνιστικός αγωνιστής των Η-χολινεργικών υποδοχέων, αναστέλλοντας τη λειτουργία των νευρομυϊκών συνάψεων.
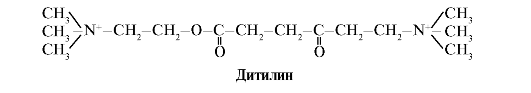 Οι νευρομυϊκές συνάψεις των σκελετικών μυών περιέχουν Η-χολινεργικούς υποδοχείς. Η αλληλεπίδρασή τους με την ακετυλοχολίνη οδηγεί σε μυϊκές συσπάσεις. Κατά τη διάρκεια ορισμένων χειρουργικών επεμβάσεων, καθώς και σε ενδοσκοπικές μελέτες, χρησιμοποιούνται φάρμακα που προκαλούν χαλάρωση των σκελετικών μυών (μυοχαλαρωτικά).Αυτές περιλαμβάνουν τη διθιλίνη, η οποία είναι ένα δομικό ανάλογο της ακετυλοχολίνης. Προσκολλάται στους Η-χολινεργικούς υποδοχείς, αλλά σε αντίθεση με την ακετυλοχολίνη, καταστρέφεται πολύ αργά από το ένζυμο ακετυλοχολινεστεράση. Ως αποτέλεσμα του παρατεταμένου ανοίγματος των διαύλων ιόντων και της επίμονης εκπόλωσης της μεμβράνης, διαταράσσεται η αγωγή των νευρικών ερεθισμάτων και επέρχεται μυϊκή χαλάρωση. Αρχικά, αυτές οι ιδιότητες ανακαλύφθηκαν στο δηλητήριο curare, γι' αυτό και ονομάζονται τέτοια φάρμακα σαν curare.
Οι νευρομυϊκές συνάψεις των σκελετικών μυών περιέχουν Η-χολινεργικούς υποδοχείς. Η αλληλεπίδρασή τους με την ακετυλοχολίνη οδηγεί σε μυϊκές συσπάσεις. Κατά τη διάρκεια ορισμένων χειρουργικών επεμβάσεων, καθώς και σε ενδοσκοπικές μελέτες, χρησιμοποιούνται φάρμακα που προκαλούν χαλάρωση των σκελετικών μυών (μυοχαλαρωτικά).Αυτές περιλαμβάνουν τη διθιλίνη, η οποία είναι ένα δομικό ανάλογο της ακετυλοχολίνης. Προσκολλάται στους Η-χολινεργικούς υποδοχείς, αλλά σε αντίθεση με την ακετυλοχολίνη, καταστρέφεται πολύ αργά από το ένζυμο ακετυλοχολινεστεράση. Ως αποτέλεσμα του παρατεταμένου ανοίγματος των διαύλων ιόντων και της επίμονης εκπόλωσης της μεμβράνης, διαταράσσεται η αγωγή των νευρικών ερεθισμάτων και επέρχεται μυϊκή χαλάρωση. Αρχικά, αυτές οι ιδιότητες ανακαλύφθηκαν στο δηλητήριο curare, γι' αυτό και ονομάζονται τέτοια φάρμακα σαν curare.
ΘΕΜΑ 1.3. ΜΕΤΟΥΣΩΣΗ ΠΡΩΤΕΪΝΩΝ ΚΑΙ ΔΥΝΑΤΟΤΗΤΑ ΤΗΣ ΑΥΘΟΡΜΗΤΑΣ ΑΝΑΚΑΤΑΣΚΕΥΗΣ ΤΟΥΣ
1. Δεδομένου ότι η φυσική διαμόρφωση των πρωτεϊνών διατηρείται λόγω αδύναμων αλληλεπιδράσεων, αλλαγών στη σύνθεση και τις ιδιότητες του περιβάλλοντος που περιβάλλει την πρωτεΐνη, η έκθεση σε χημικά αντιδραστήρια και φυσικούς παράγοντες προκαλούν αλλαγή στη διάταξή τους (ιδιότητα διαμορφωτικής αστάθειας). Το σπάσιμο ενός μεγάλου αριθμού δεσμών οδηγεί στην καταστροφή της φυσικής διαμόρφωσης και στη μετουσίωση των πρωτεϊνών.
Μετουσίωσης πρωτεϊνών- αυτή είναι η καταστροφή της φυσικής τους διαμόρφωσης υπό την επίδραση μετουσιωτικών παραγόντων, που προκαλείται από τη ρήξη αδύναμων δεσμών που σταθεροποιούν τη χωρική δομή της πρωτεΐνης. Η μετουσίωση συνοδεύεται από την καταστροφή της μοναδικής τρισδιάστατης δομής και του ενεργού κέντρου της πρωτεΐνης και την απώλεια της βιολογικής της δραστηριότητας (Εικ. 1.16).
Όλα τα μετουσιωμένα μόρια μιας πρωτεΐνης αποκτούν μια τυχαία διαμόρφωση που διαφέρει από άλλα μόρια της ίδιας πρωτεΐνης. Οι ρίζες αμινοξέων που σχηματίζουν το ενεργό κέντρο αποδεικνύονται ότι είναι χωρικά απομακρυσμένες η μία από την άλλη, δηλ. η ειδική θέση δέσμευσης της πρωτεΐνης με τον συνδέτη καταστρέφεται. Κατά τη διάρκεια της μετουσίωσης, η πρωτογενής δομή των πρωτεϊνών παραμένει αμετάβλητη.
Εφαρμογή μετουσιωτικών παραγόντων στη βιολογική έρευνα και ιατρική.Σε βιοχημικές μελέτες, πριν από τον προσδιορισμό των ενώσεων χαμηλού μοριακού βάρους σε βιολογικό υλικό, οι πρωτεΐνες συνήθως αφαιρούνται πρώτα από το διάλυμα. Το τριχλωροξικό οξύ (TCA) χρησιμοποιείται συχνότερα για το σκοπό αυτό. Μετά την προσθήκη TCA στο διάλυμα, οι μετουσιωμένες πρωτεΐνες καθιζάνουν και απομακρύνονται εύκολα με διήθηση (Πίνακας 1.1.)
Στην ιατρική, οι μετουσιωτικοί παράγοντες χρησιμοποιούνται συχνά για την αποστείρωση ιατρικών εργαλείων και υλικών σε αυτόκλειστα (ο παράγοντας μετουσίωσης είναι υψηλή θερμοκρασία) και ως αντισηπτικά (οινόπνευμα, φαινόλη, χλωραμίνη) για τη θεραπεία μολυσμένων επιφανειών που περιέχουν παθογόνο μικροχλωρίδα.
2. Αυθόρμητη επανενεργοποίηση πρωτεΐνης- απόδειξη του ντετερμινισμού της πρωτογενούς δομής, διαμόρφωσης και λειτουργίας των πρωτεϊνών. Οι μεμονωμένες πρωτεΐνες είναι προϊόντα ενός γονιδίου που έχουν την ίδια αλληλουχία αμινοξέων και αποκτούν την ίδια διαμόρφωση στο κύτταρο. Το θεμελιώδες συμπέρασμα ότι η πρωτογενής δομή μιας πρωτεΐνης περιέχει ήδη πληροφορίες για τη διαμόρφωση και τη λειτουργία της έγινε με βάση την ικανότητα ορισμένων πρωτεϊνών (ιδιαίτερα, της ριβονουκλεάσης και της μυοσφαιρίνης) να επαναφέρουν αυθόρμητα - αποκαθιστούν τη φυσική τους διαμόρφωση μετά τη μετουσίωση.
Ο σχηματισμός χωρικών πρωτεϊνικών δομών πραγματοποιείται με τη μέθοδο της αυτοσυναρμολόγησης - μια αυθόρμητη διαδικασία στην οποία μια πολυπεπτιδική αλυσίδα, η οποία έχει μια μοναδική πρωτογενή δομή, τείνει να υιοθετήσει μια διαμόρφωση με τη χαμηλότερη ελεύθερη ενέργεια σε διάλυμα. Η ικανότητα αναγέννησης πρωτεϊνών που διατηρούν την πρωταρχική τους δομή μετά τη μετουσίωση περιγράφηκε σε ένα πείραμα με το ένζυμο ριβονουκλεάση.
Η ριβονουκλεάση είναι ένα ένζυμο που διασπά τους δεσμούς μεταξύ μεμονωμένων νουκλεοτιδίων σε ένα μόριο RNA. Αυτή η σφαιρική πρωτεΐνη έχει μία πολυπεπτιδική αλυσίδα, η τριτοταγής δομή της οποίας σταθεροποιείται από πολλούς ασθενείς και τέσσερις δισουλφιδικούς δεσμούς.
Η επεξεργασία της ριβονουκλεάσης με ουρία, η οποία διασπά τους δεσμούς υδρογόνου στο μόριο, και έναν αναγωγικό παράγοντα, που σπάει τους δισουλφιδικούς δεσμούς, οδηγεί σε μετουσίωση του ενζύμου και απώλεια της δραστηριότητάς του.
Η απομάκρυνση των μετουσιωτικών παραγόντων με αιμοκάθαρση οδηγεί σε αποκατάσταση της πρωτεϊνικής διαμόρφωσης και λειτουργίας, δηλ. στην αναγέννηση. (Εικ. 1.17).
 Ρύζι. 1.17. Μετουσίωση και ανανέωση της ριβονουκλεάσης
Ρύζι. 1.17. Μετουσίωση και ανανέωση της ριβονουκλεάσης
Α - φυσική διαμόρφωση της ριβονουκλεάσης, στην τριτοταγή δομή της οποίας υπάρχουν τέσσερις δισουλφιδικοί δεσμοί. Β - μετουσιωμένο μόριο ριβονουκλεάσης.
Β - επανενεργοποιημένο μόριο ριβονουκλεάσης με αποκατασταθείσα δομή και λειτουργία
1. Συμπληρώστε τον πίνακα 1.2.
Πίνακας 1.2. Ταξινόμηση αμινοξέων σύμφωνα με την πολικότητα των ριζών
2. Γράψτε τον τύπο του τετραπεπτιδίου:
Asp - Pro - Fen - Liz
α) επισημάνετε τις επαναλαμβανόμενες ομάδες στο πεπτίδιο που σχηματίζουν τη ραχοκοκαλιά του πεπτιδίου και τις μεταβλητές ομάδες που αντιπροσωπεύονται από ρίζες αμινοξέων.
β) επισημάνετε τα Ν- και C-άκρα.
γ) επισημάνετε τους πεπτιδικούς δεσμούς.
δ) γράψτε ένα άλλο πεπτίδιο που αποτελείται από τα ίδια αμινοξέα.
ε) μετρήστε τον αριθμό των πιθανών παραλλαγών ενός τετραπεπτιδίου με παρόμοια σύνθεση αμινοξέων.
3. Εξηγήστε το ρόλο της πρωτογενούς δομής των πρωτεϊνών χρησιμοποιώντας το παράδειγμα μιας συγκριτικής ανάλυσης δύο δομικά όμοιων και εξελικτικά κοντινών πεπτιδικών ορμονών της νευροϋπόφυσης των θηλαστικών - ωκυτοκίνη και βαζοπρεσίνη (Πίνακας 1.3).
Πίνακας 1.3. Δομή και λειτουργίες της ωκυτοκίνης και της βαζοπρεσίνης
 Για αυτό:
Για αυτό:
α) Συγκρίνετε τη σύνθεση και την αλληλουχία αμινοξέων δύο πεπτιδίων.
β) βρείτε την ομοιότητα της πρωτογενούς δομής των δύο πεπτιδίων και την ομοιότητα της βιολογικής τους δράσης.
γ) βρείτε διαφορές στη δομή δύο πεπτιδίων και διαφορές στις λειτουργίες τους.
δ) εξάγετε ένα συμπέρασμα σχετικά με την επίδραση της πρωτογενούς δομής των πεπτιδίων στις λειτουργίες τους.
4. Περιγράψτε τα κύρια στάδια σχηματισμού της διαμόρφωσης των σφαιρικών πρωτεϊνών (δευτερογενείς, τριτογενείς δομές, η έννοια της υπερδευτερογενούς δομής). Υποδείξτε τους τύπους δεσμών που εμπλέκονται στο σχηματισμό πρωτεϊνικών δομών. Ποιες ρίζες αμινοξέων μπορούν να συμμετέχουν στο σχηματισμό υδρόφοβων αλληλεπιδράσεων, ιοντικών, δεσμών υδρογόνου.
Δώσε παραδείγματα.
5. Ορίστε την έννοια της «διαμόρφωσης αστάθειας των πρωτεϊνών», αναφέρετε τους λόγους ύπαρξης και σημασίας της.
6. Αναπτύξτε το νόημα της ακόλουθης φράσης: «Η λειτουργία των πρωτεϊνών βασίζεται στην ειδική αλληλεπίδρασή τους με τον συνδέτη», χρησιμοποιώντας τους όρους και εξηγώντας τη σημασία τους: διαμόρφωση πρωτεΐνης, ενεργό κέντρο, συνδετήρας, συμπληρωματικότητα, πρωτεϊνική λειτουργία.
7. Χρησιμοποιώντας ένα παράδειγμα, εξηγήστε τι είναι οι τομείς και ποιος είναι ο ρόλος τους στη λειτουργία των πρωτεϊνών.
ΚΑΘΗΚΟΝΤΑ ΑΥΤΟΕΛΕΓΧΟΥ
1. Αγώνας.
Λειτουργική ομάδα σε ρίζα αμινοξέων:
Α. Καρβοξυλική ομάδα Β. Υδροξυλομάδα Γ Ομάδα γουανιδίνης Δ. Ομάδα θειόλης Ε. Αμινο ομάδα
2. Επιλέξτε τις σωστές απαντήσεις.
Τα αμινοξέα με πολικές αφόρτιστες ρίζες είναι:
Α. Cis B. Asn
Β. Γλου Γ. Τρεις
3. Επιλέξτε τις σωστές απαντήσεις.
Ρίζες αμινοξέων:
Α. Παρέχετε ειδικότητα της πρωτογενούς δομής Β. Συμμετέχετε στο σχηματισμό της τριτογενούς δομής
Β. Βρίσκονται στην επιφάνεια της πρωτεΐνης, επηρεάζουν τη διαλυτότητά της Δ. Σχηματίζουν το ενεργό κέντρο
Δ. Συμμετέχουν στο σχηματισμό πεπτιδικών δεσμών
4. Επιλέξτε τις σωστές απαντήσεις.
Υδροφοβικές αλληλεπιδράσεις μπορεί να δημιουργηθούν μεταξύ των ριζών αμινοξέων:
A. Tre Lay B. Pro Three
B. Met Ile G. Tir Ala D. Val Fen
5. Επιλέξτε τις σωστές απαντήσεις.
Μεταξύ των ριζών αμινοξέων μπορούν να σχηματιστούν ιονικοί δεσμοί:
Α. Γλν Ασπ Β. Απρ Λιζ
B. Liz Glu G. Gis Asp D. Asn Apr
6. Επιλέξτε τις σωστές απαντήσεις.
Οι δεσμοί υδρογόνου μπορούν να σχηματιστούν μεταξύ των ριζών αμινοξέων:
Α. Ser Gln Β. Cis Tre
Β. Asp Liz G. Glu Asp D. Asn Tre
7. Αγώνας.
Τύπος δεσμού που εμπλέκεται στο σχηματισμό της δομής της πρωτεΐνης:
Α. Πρωτογενής δομή Β. Δευτερεύουσα δομή
Β. Τριτογενής δομή
Δ. Υπερδευτερογενής δομή Ε. Διαμόρφωση.
1. Δεσμοί υδρογόνου μεταξύ ατόμων της ραχοκοκαλιάς του πεπτιδίου
2. Αδύναμοι δεσμοί μεταξύ λειτουργικών ομάδων ριζών αμινοξέων
3. Δεσμοί μεταξύ α-αμινο και α-καρβοξυλικών ομάδων αμινοξέων
8. Επιλέξτε τις σωστές απαντήσεις. Τρυψίνη:
Α. Πρωτεολυτικό ένζυμο Β. Περιέχει δύο τομείς
Β. Υδρολύει το άμυλο
Δ. Ο ενεργός ιστότοπος βρίσκεται μεταξύ των τομέων. Δ. Αποτελείται από δύο πολυπεπτιδικές αλυσίδες.
9. Επιλέξτε τις σωστές απαντήσεις. Ατροπίνη:
Α. Νευροδιαβιβαστής
Β. Δομικό ανάλογο της ακετυλοχολίνης
Β. Αλληλεπιδρά με Η-χολινεργικούς υποδοχείς
Δ. Ενισχύει την αγωγή των νευρικών ερεθισμάτων μέσω των χολινεργικών συνάψεων
Δ. Ανταγωνιστικός αναστολέας Μ-χολινεργικών υποδοχέων
10. Επιλέξτε τις σωστές δηλώσεις. Σε πρωτεΐνες:
Α. Η κύρια δομή περιέχει πληροφορίες σχετικά με τη δομή της ενεργού τοποθεσίας της
Β. Το ενεργό κέντρο διαμορφώνεται στο επίπεδο της πρωτογενούς δομής
Β. Η διαμόρφωση στερεώνεται άκαμπτα με ομοιοπολικούς δεσμούς
Δ. Η ενεργή θέση μπορεί να αλληλεπιδράσει με μια ομάδα παρόμοιων προσδεμάτων
λόγω της διαμορφωτικής αστάθειας των πρωτεϊνών Δ. Αλλαγή περιβάλλον, μπορεί να επηρεάσει τη συγγένεια του ενεργού
κέντρο σε συνδέτη
1. 1-B, 2-G, 3-B.
3. Α, Β, Γ, Δ.
7. 1-B, 2-D, 3-A.
8. Α, Β, Γ, Δ.
ΒΑΣΙΚΟΙ ΟΡΟΙ ΚΑΙ ΕΝΝΟΙΕΣ
1. Πρωτεΐνες, πολυπεπτίδια, αμινοξέα
2. Πρωτογενείς, δευτερογενείς, τριτοταγείς πρωτεϊνικές δομές
3. Διαμόρφωση, διαμόρφωση φυσικής πρωτεΐνης
4. Ομοιοπολικοί και αδύναμοι δεσμοί στην πρωτεΐνη
5. Διαμορφωτική αστάθεια
6. Πρωτεϊνική ενεργή θέση
7. Συνδέματα
8. Αναδίπλωση πρωτεΐνης
9. Δομικά ανάλογα προσδεμάτων
10. Πρωτεΐνες τομέα
11. Απλές και σύνθετες πρωτεΐνες
12. Μετουσίωσης πρωτεϊνών, μετουσιωτικοί παράγοντες
13. Επανενεργοποίηση πρωτεϊνών
Λύνω προβλήματα
«Δομική οργάνωση των πρωτεϊνών και η βάση της λειτουργίας τους»
1. Η κύρια λειτουργία της πρωτεΐνης - αιμοσφαιρίνης Α (HbA) είναι η μεταφορά οξυγόνου στους ιστούς. Στον ανθρώπινο πληθυσμό, είναι γνωστές πολλαπλές μορφές αυτής της πρωτεΐνης με αλλοιωμένες ιδιότητες και λειτουργία - οι λεγόμενες μη φυσιολογικές αιμοσφαιρίνες. Για παράδειγμα, η αιμοσφαιρίνη S, που βρίσκεται στα ερυθρά αιμοσφαίρια ασθενών με δρεπανοκυτταρική αναιμία (HbS), έχει βρεθεί ότι έχει χαμηλή διαλυτότητα υπό συνθήκες χαμηλής μερικής πίεσης οξυγόνου (όπως συμβαίνει στο φλεβικό αίμα). Αυτό οδηγεί στο σχηματισμό συσσωματωμάτων αυτής της πρωτεΐνης. Η πρωτεΐνη χάνει τη λειτουργία της, κατακρημνίζεται και αποκτούν τα ερυθρά αιμοσφαίρια ακανόνιστο σχήμα(μερικά από αυτά σχηματίζουν δρεπανοειδές σχήμα) και καταστρέφονται πιο γρήγορα από το συνηθισμένο στη σπλήνα. Ως αποτέλεσμα, αναπτύσσεται δρεπανοκυτταρική αναιμία.
Η μόνη διαφορά στην πρωτογενή δομή της HbA βρέθηκε στη Ν-τελική περιοχή της β-αλυσίδας της αιμοσφαιρίνης. Συγκρίνετε τις Ν-τελικές περιοχές του β-κλώνου και δείξτε πώς οι αλλαγές στην πρωτογενή δομή της πρωτεΐνης επηρεάζουν τις ιδιότητες και τις λειτουργίες της.
 Για αυτό:
Για αυτό:
α) Να γράψετε τους τύπους των αμινοξέων με τους οποίους διαφέρει η HbA και να συγκρίνετε τις ιδιότητες αυτών των αμινοξέων (πολικότητα, φορτίο).
β) εξάγετε συμπέρασμα σχετικά με τον λόγο της μείωσης της διαλυτότητας και της διακοπής της μεταφοράς οξυγόνου στους ιστούς.
2. Το σχήμα δείχνει ένα διάγραμμα της δομής μιας πρωτεΐνης που έχει κέντρο δέσμευσης με έναν συνδέτη (ενεργό κέντρο). Εξηγήστε γιατί η πρωτεΐνη είναι επιλεκτική στην επιλογή του συνδέτη. Για αυτό:
α) θυμηθείτε ποιο είναι το ενεργό κέντρο μιας πρωτεΐνης και σκεφτείτε τη δομή του ενεργού κέντρου της πρωτεΐνης που φαίνεται στο σχήμα.
β) γράψτε τους τύπους των ριζών αμινοξέων που αποτελούν το ενεργό κέντρο.
γ) σχεδιάστε έναν συνδέτη που θα μπορούσε να αλληλεπιδράσει ειδικά με την ενεργό θέση της πρωτεΐνης. Αναφέρετε σε αυτό τις λειτουργικές ομάδες που μπορούν να σχηματίσουν δεσμούς με τις ρίζες αμινοξέων που αποτελούν το ενεργό κέντρο.
δ) υποδεικνύουν τους τύπους δεσμών που προκύπτουν μεταξύ του συνδετήρα και των ριζών αμινοξέων του ενεργού κέντρου.
ε) εξηγήστε σε τι βασίζεται η ειδικότητα της αλληλεπίδρασης πρωτεΐνης-προσδέματος.
 3.
Το σχήμα δείχνει την ενεργό θέση της πρωτεΐνης και αρκετών προσδεμάτων.
3.
Το σχήμα δείχνει την ενεργό θέση της πρωτεΐνης και αρκετών προσδεμάτων.
Προσδιορίστε ποιος συνδέτης είναι πιο πιθανό να αλληλεπιδράσει με την ενεργό θέση της πρωτεΐνης και γιατί.
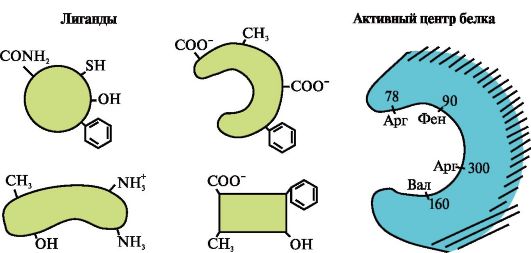 Ποιοι τύποι δεσμών προκύπτουν κατά τον σχηματισμό ενός συμπλόκου πρωτεΐνης-συνδέτη;
Ποιοι τύποι δεσμών προκύπτουν κατά τον σχηματισμό ενός συμπλόκου πρωτεΐνης-συνδέτη;
4. Δομικά ανάλογα φυσικών πρωτεϊνικών προσδεμάτων μπορούν να χρησιμοποιηθούν ως φάρμακα για την τροποποίηση της δραστηριότητας των πρωτεϊνών.
Η ακετυλοχολίνη είναι ένας μεσολαβητής μετάδοσης διέγερσης στις νευρομυϊκές συνάψεις. Όταν η ακετυλοχολίνη αλληλεπιδρά με πρωτεΐνες - υποδοχείς της μετασυναπτικής μεμβράνης των σκελετικών μυών, ανοίγουν οι δίαυλοι ιόντων και εμφανίζεται μυϊκή σύσπαση. Το Ditilin είναι ένα φάρμακο που χρησιμοποιείται σε ορισμένες επεμβάσεις για τη χαλάρωση των μυών, καθώς διαταράσσει τη μετάδοση των νευρικών ερεθισμάτων μέσω των νευρομυϊκών συνάψεων. Εξηγήστε τον μηχανισμό δράσης της διτιλίνης ως μυοχαλαρωτικού. Για αυτό:
α) Να γράψετε τους τύπους της ακετυλοχολίνης και της διθιλίνης και να συγκρίνετε τις δομές τους.
β) να περιγράψετε τον μηχανισμό της χαλαρωτικής δράσης της διτιλίνης.
5. Σε ορισμένες ασθένειες, η θερμοκρασία του σώματος του ασθενούς αυξάνεται, κάτι που θεωρείται ως προστατευτική αντίδραση του οργανισμού. Ωστόσο, οι υψηλές θερμοκρασίες είναι επιζήμιες για τις πρωτεΐνες του σώματος. Εξηγήστε γιατί σε θερμοκρασίες άνω των 40 °C η λειτουργία των πρωτεϊνών διαταράσσεται και δημιουργείται κίνδυνος για την ανθρώπινη ζωή. Για να το κάνετε αυτό, θυμηθείτε:
1) Η δομή των πρωτεϊνών και οι δεσμοί που συγκρατούν τη δομή της στη φυσική διαμόρφωση.
2) Πώς αλλάζει η δομή και η λειτουργία των πρωτεϊνών με την αύξηση της θερμοκρασίας;
3) Τι είναι η ομοιόσταση και γιατί είναι σημαντική για τη διατήρηση της ανθρώπινης υγείας.
Modular unit 2 OLIGOMERICK PROTEINS ΩΣ ΣΤΟΧΟΙ ΡΥΘΜΙΣΤΙΚΩΝ ΕΠΙΡΡΟΕΩΝ. ΔΟΜΗ ΚΑΙ ΛΕΙΤΟΥΡΓΙΚΗ ΠΟΙΚΙΛΟΛΟΓΙΑ ΠΡΩΤΕΪΝΩΝ. ΜΕΘΟΔΟΙ ΔΙΑΧΩΡΙΣΜΟΥ ΚΑΙ ΚΑΘΑΡΙΣΜΟΥ ΠΡΩΤΕΪΝΩΝ
Μαθησιακοί στόχοι Να είναι σε θέση:
1. Χρησιμοποιήστε γνώσεις σχετικά με τα χαρακτηριστικά της δομής και τις λειτουργίες των ολιγομερών πρωτεϊνών για να κατανοήσετε τους προσαρμοστικούς μηχανισμούς ρύθμισης των λειτουργιών τους.
2. Εξηγήστε το ρόλο των συνοδών στη σύνθεση και τη διατήρηση της πρωτεϊνικής διαμόρφωσης υπό κυτταρικές συνθήκες.
3. Εξηγήστε την ποικιλία των εκδηλώσεων της ζωής από την ποικιλία των δομών και των λειτουργιών των πρωτεϊνών που συντίθενται στο σώμα.
4. Αναλύστε τη σχέση μεταξύ της δομής των πρωτεϊνών και της λειτουργίας τους χρησιμοποιώντας παραδείγματα σύγκρισης σχετικών αιμοπρωτεϊνών - μυοσφαιρίνης και αιμοσφαιρίνης, καθώς και εκπροσώπων των πέντε τάξεων πρωτεϊνών της οικογένειας ανοσοσφαιρινών.
5. Εφαρμόστε γνώσεις σχετικά με τις ιδιαιτερότητες των φυσικών και χημικών ιδιοτήτων των πρωτεϊνών για να επιλέξετε μεθόδους καθαρισμού τους από άλλες πρωτεΐνες και ακαθαρσίες.
6. Ερμηνεύστε τα αποτελέσματα των ποσοτικών και ποιοτική σύνθεσηπρωτεΐνες πλάσματος αίματος για επιβεβαίωση ή διευκρίνιση της κλινικής διάγνωσης.
Ξέρω:
1. Χαρακτηριστικά της δομής των ολιγομερών πρωτεϊνών και προσαρμοστικοί μηχανισμοί για τη ρύθμιση των λειτουργιών τους χρησιμοποιώντας το παράδειγμα της αιμοσφαιρίνης.
2. Η δομή και οι λειτουργίες των συνοδών και η σημασία τους για τη διατήρηση της φυσικής διαμόρφωσης των πρωτεϊνών υπό κυτταρικές συνθήκες.
3. Αρχές συνδυασμού πρωτεϊνών σε οικογένειες με βάση την ομοιότητα της διαμόρφωσης και των λειτουργιών τους χρησιμοποιώντας το παράδειγμα των ανοσοσφαιρινών.
4. Μέθοδοι διαχωρισμού πρωτεϊνών με βάση τα χαρακτηριστικά των φυσικοχημικών ιδιοτήτων τους.
5. Ηλεκτροφόρηση πλάσματος αίματος ως μέθοδος αξιολόγησης της ποιοτικής και ποσοτικής σύνθεσης των πρωτεϊνών.
ΘΕΜΑ 1.4. ΧΑΡΑΚΤΗΡΙΣΤΙΚΑ ΤΗΣ ΔΟΜΗΣ ΚΑΙ ΤΗΣ ΛΕΙΤΟΥΡΓΙΑΣ ΤΩΝ ΟΛΙΓΟΜΕΡΩΝ ΠΡΩΤΕΪΝΩΝ ΜΕ ΤΟ ΠΑΡΑΔΕΙΓΜΑ ΤΗΣ ΑΙΜΟΓΛΟΒΙΝΗΣ
1. Πολλές πρωτεΐνες περιέχουν πολλές πολυπεπτιδικές αλυσίδες. Τέτοιες πρωτεΐνες ονομάζονται ολιγομερές,και μεμονωμένες αλυσίδες - πρωτομερή.Τα πρωτομερή των ολιγομερών πρωτεϊνών συνδέονται με πολλούς ασθενείς μη ομοιοπολικούς δεσμούς (υδρόφοβο, ιοντικό, υδρογόνο). ΑΛΛΗΛΕΠΙΔΡΑΣΗ
τα πρωτομερή πραγματοποιούνται χάρη σε συμπληρωματικότητατις επιφάνειες επαφής τους.
Ο αριθμός των πρωτομερών στις ολιγομερείς πρωτεΐνες μπορεί να ποικίλλει πολύ: η αιμοσφαιρίνη περιέχει 4 πρωτομερή, το ένζυμο ασπαρτική αμινοτρανσφεράση έχει 12 πρωτομερή και η πρωτεΐνη του ιού του μωσαϊκού του καπνού περιέχει 2120 πρωτομερή συνδεδεμένα με μη ομοιοπολικούς δεσμούς. Κατά συνέπεια, οι ολιγομερείς πρωτεΐνες μπορεί να έχουν πολύ υψηλά μοριακά βάρη.
Η αλληλεπίδραση ενός πρωτομερούς με άλλα μπορεί να θεωρηθεί μια ειδική περίπτωση αλληλεπίδρασης πρωτεΐνης-προσδέματος, αφού κάθε πρωτομερές χρησιμεύει ως συνδετήρας για άλλα πρωτομερή. Ο αριθμός και η μέθοδος σύνδεσης των πρωτομερών σε μια πρωτεΐνη ονομάζεται δομή τεταρτοταγούς πρωτεΐνης.
Οι πρωτεΐνες μπορεί να περιέχουν πρωτομερή της ίδιας ή διαφορετικής δομής, για παράδειγμα, τα ομοδιμερή είναι πρωτεΐνες που περιέχουν δύο πανομοιότυπα πρωτομερή και τα ετεροδιμερή είναι πρωτεΐνες που περιέχουν δύο διαφορετικά πρωτομερή.
Εάν οι πρωτεΐνες περιέχουν διαφορετικά πρωτομερή, τότε μπορούν να σχηματιστούν πάνω τους κέντρα δέσμευσης με διαφορετικούς συνδέτες που διαφέρουν στη δομή. Όταν ένας συνδέτης δεσμεύεται στην ενεργό θέση, η λειτουργία αυτής της πρωτεΐνης εκδηλώνεται. Ένα κέντρο που βρίσκεται σε διαφορετικό πρωτομερές ονομάζεται αλλοστερικό (διαφορετικό από το ενεργό). Επικοινωνώντας αλλοστερικός συνδέτης ή τελεστής,εκτελεί μια ρυθμιστική λειτουργία (Εικ. 1.18). Η αλληλεπίδραση του αλλοστερικού κέντρου με τον τελεστή προκαλεί διαμορφωτικές αλλαγές στη δομή ολόκληρης της ολιγομερικής πρωτεΐνης λόγω της διαμορφωτικής της αστάθειας. Αυτό επηρεάζει τη συγγένεια της ενεργού θέσης για ένα συγκεκριμένο συνδετήρα και ρυθμίζει τη λειτουργία αυτής της πρωτεΐνης. Μια αλλαγή στη διαμόρφωση και τη λειτουργία όλων των πρωτομερών κατά τη διάρκεια της αλληλεπίδρασης μιας ολιγομερούς πρωτεΐνης με τουλάχιστον έναν συνδέτη ονομάζεται συνεργατικές διαμορφωτικές αλλαγές. Οι δράσεις που ενισχύουν τη λειτουργία των πρωτεϊνών ονομάζονται ενεργοποιητές,και τελεστές που αναστέλλουν τη λειτουργία του - αναστολείς.
Έτσι, οι ολιγομερείς πρωτεΐνες, καθώς και οι πρωτεΐνες με δομή τομέα, έχουν μια νέα ιδιότητα σε σύγκριση με τις μονομερείς πρωτεΐνες - την ικανότητα αλλοστερικής ρύθμισης των λειτουργιών (ρύθμιση με σύνδεση διαφορετικών προσδεμάτων στην πρωτεΐνη). Αυτό μπορεί να φανεί συγκρίνοντας τις δομές και τις λειτουργίες δύο στενά συνδεδεμένων σύνθετων πρωτεϊνών, της μυοσφαιρίνης και της αιμοσφαιρίνης.
 Ρύζι. 1.18. Σχήμα δομής διμερούς πρωτεΐνης
Ρύζι. 1.18. Σχήμα δομής διμερούς πρωτεΐνης
2. Σχηματισμός χωρικών δομών και λειτουργία μυοσφαιρίνης.
Η μυοσφαιρίνη (Mb) είναι μια πρωτεΐνη που βρίσκεται στους κόκκινους μύες, η κύρια λειτουργία της οποίας είναι να δημιουργεί αποθέματα Ο 2 απαραίτητα για έντονη μυϊκή εργασία. Το Mb είναι μια σύνθετη πρωτεΐνη που περιέχει ένα πρωτεϊνικό μέρος - apoMb και ένα μη πρωτεϊνικό μέρος - την αίμη. Η πρωταρχική δομή του apoMB καθορίζει τη συμπαγή σφαιρική του διαμόρφωση και τη δομή του ενεργού κέντρου, στο οποίο συνδέεται το μη πρωτεϊνικό τμήμα της μυοσφαιρίνης, η αίμη. Το οξυγόνο που προέρχεται από το αίμα στους μύες συνδέεται με τις αίμες Fe+2 στη μυοσφαιρίνη. Το Mb είναι μια μονομερής πρωτεΐνη που έχει πολύ υψηλή συγγένεια για το O 2, επομένως η απελευθέρωση οξυγόνου από τη μυοσφαιρίνη συμβαίνει μόνο κατά τη διάρκεια έντονης μυϊκής εργασίας, όταν η μερική πίεση του O 2 μειώνεται απότομα.
Σχηματισμός της διαμόρφωσης Mv.Στους κόκκινους μύες, στα ριβοσώματα κατά τη μετάφραση, συντίθεται η πρωταρχική δομή του ΜΒ, που αντιπροσωπεύεται από μια συγκεκριμένη αλληλουχία 153 υπολειμμάτων αμινοξέων. Η δευτερεύουσα δομή του Mb περιέχει οκτώ α-έλικες, που ονομάζονται με λατινικά γράμματα από το Α έως το Η, μεταξύ των οποίων υπάρχουν μη ελικοειδείς περιοχές. Η τριτογενής δομή του Mb έχει τη μορφή συμπαγούς σφαιριδίου, στην εσοχή του οποίου βρίσκεται το ενεργό κέντρο μεταξύ των α-έλικων F και E (Εικ. 1.19).
 Ρύζι. 1.19. Δομή μυοσφαιρίνης
Ρύζι. 1.19. Δομή μυοσφαιρίνης
3. Χαρακτηριστικά της δομής και της λειτουργίας του ενεργού κέντρου του MV.Το ενεργό κέντρο του Mb σχηματίζεται κυρίως από υδρόφοβες ρίζες αμινοξέων, σε μεγάλη απόσταση μεταξύ τους στην πρωτογενή δομή (για παράδειγμα, Tri 3 9 και Fen 138) Ελάχιστα διαλυτά στο νερό συνδέτες - αίμη και O 2 - προσκολλώνται στο ενεργό κέντρο. Η αίμη είναι ένας ειδικός συνδέτης του apoMB (Εικ. 1.20), η βάση του οποίου αποτελείται από τέσσερις δακτυλίους πυρρολίου που συνδέονται με γέφυρες μεθενυλίου. στο κέντρο υπάρχει ένα άτομο Fe+ 2 συνδεδεμένο με τα άτομα αζώτου των δακτυλίων πυρρολίου με τέσσερις δεσμούς συντονισμού. Στο ενεργό κέντρο του Mb, εκτός από τις υδρόφοβες ρίζες αμινοξέων, υπάρχουν επίσης υπολείμματα δύο αμινοξέων με υδρόφιλες ρίζες - Gis E 7(Gis 64) και GIS F 8(Το 93 του) (Εικ. 1.21).
 Ρύζι. 1.20. Η δομή της αίμης - το μη πρωτεϊνικό μέρος της μυοσφαιρίνης και της αιμοσφαιρίνης
Ρύζι. 1.20. Η δομή της αίμης - το μη πρωτεϊνικό μέρος της μυοσφαιρίνης και της αιμοσφαιρίνης
 Ρύζι. 1.21. Εντόπιση της αίμης και του Ο2 στην ενεργό θέση της απομυοσφαιρίνης και των πρωτομερών της αιμοσφαιρίνης
Ρύζι. 1.21. Εντόπιση της αίμης και του Ο2 στην ενεργό θέση της απομυοσφαιρίνης και των πρωτομερών της αιμοσφαιρίνης
Η αίμη συνδέεται ομοιοπολικά με το His F8 μέσω ενός ατόμου σιδήρου. Το O 2 προσκολλάται στο σίδερο στην άλλη πλευρά του επιπέδου αίμης. Το E 7 του είναι απαραίτητο για τον σωστό προσανατολισμό του O 2 και διευκολύνει την προσθήκη οξυγόνου στην αίμη Fe + 2
GIS F 8σχηματίζει δεσμό συντονισμού με το Fe+2 και σταθεροποιεί την αίμη στο ενεργό κέντρο. Gis E 7απαραίτητο για τον σωστό προσανατολισμό στο ενεργό κέντρο ενός άλλου συνδέτη - O 2 κατά την αλληλεπίδρασή του με την αίμη Fe + 2. Το μικροπεριβάλλον της αίμης δημιουργεί συνθήκες για ισχυρή αλλά αναστρέψιμη δέσμευση του O 2 με το Fe + 2 και εμποδίζει το νερό να εισέλθει στην υδρόφοβη ενεργή θέση, γεγονός που μπορεί να οδηγήσει στην οξείδωσή του σε Fe + 3.
Η μονομερής δομή του Mb και το ενεργό κέντρο του καθορίζει την υψηλή συγγένεια της πρωτεΐνης για το O 2.
4. Ολιγομερής δομή της Hb και ρύθμιση της συγγένειας της Hb προς τους συνδέτες O 2. Ανθρώπινες αιμοσφαιρίνες- μια οικογένεια πρωτεϊνών, όπως η μυοσφαιρίνη, που σχετίζεται με σύνθετες πρωτεΐνες (αιμοπρωτεΐνες). Έχουν τετραμερή δομή και περιέχουν δύο α-αλυσίδες, αλλά διαφέρουν ως προς τη δομή των άλλων δύο πολυπεπτιδικών αλυσίδων (2α-, 2χ-αλυσίδες). Η δομή της δεύτερης πολυπεπτιδικής αλυσίδας καθορίζει τα χαρακτηριστικά της λειτουργίας αυτών των μορφών Hb. Περίπου το 98% της αιμοσφαιρίνης στα ερυθρά αιμοσφαίρια ενός ενήλικα είναι αιμοσφαιρίνη Α(2α-, 2ρ-αλυσίδες).
Κατά την ανάπτυξη του εμβρύου λειτουργούν δύο κύριοι τύποι αιμοσφαιρινών: εμβρυϊκή Hb(2α, 2ε), το οποίο βρίσκεται στο πρώιμα στάδιαανάπτυξη του εμβρύου και αιμοσφαιρίνη F (εμβρυϊκή)- (2α, 2γ), που αντικαθιστά την πρώιμη εμβρυϊκή αιμοσφαιρίνη στον έκτο μήνα της ενδομήτριας ανάπτυξης και μόνο μετά τη γέννηση αντικαθίσταται από Hb A.
Το HB A είναι μια πρωτεΐνη που σχετίζεται με τη μυοσφαιρίνη (MB) που βρίσκεται στα ερυθρά αιμοσφαίρια του ενήλικου ανθρώπου. Η δομή των μεμονωμένων πρωτομερών του είναι παρόμοια με αυτή της μυοσφαιρίνης. Οι δευτερογενείς και τριτοταγείς δομές της μυοσφαιρίνης και των πρωτομερών αιμοσφαιρίνης είναι πολύ παρόμοιες, παρά το γεγονός ότι στην πρωτογενή δομή των πολυπεπτιδικών τους αλυσίδων μόνο 24 υπολείμματα αμινοξέων είναι πανομοιότυπα (η δευτερογενής δομή των πρωτομερών αιμοσφαιρίνης, όπως η μυοσφαιρίνη, περιέχει οκτώ α-έλικες, χαρακτηρίζεται με λατινικά γράμματα από το A έως το H, και η τριτογενής δομή έχει τη μορφή συμπαγούς σφαιριδίου). Αλλά σε αντίθεση με τη μυοσφαιρίνη, η αιμοσφαιρίνη έχει μια ολιγομερή δομή, που αποτελείται από τέσσερις πολυπεπτιδικές αλυσίδες που συνδέονται με μη ομοιοπολικούς δεσμούς (Εικόνα 1.22).
Κάθε πρωτομερές Hb σχετίζεται με ένα μη πρωτεϊνικό μέρος - την αίμη και τα γειτονικά πρωτομερή. Η σύνδεση του πρωτεϊνικού τμήματος της Hb με την αίμη είναι παρόμοια με εκείνη της μυοσφαιρίνης: στο ενεργό κέντρο της πρωτεΐνης, τα υδρόφοβα μέρη της αίμης περιβάλλονται από υδρόφοβες ρίζες αμινοξέων με εξαίρεση τα His F 8 και His E 7, που βρίσκονται και στις δύο πλευρές του επιπέδου της αίμης και παίζουν παρόμοιο ρόλο στη λειτουργία της πρωτεΐνης και στη σύνδεσή της με το οξυγόνο (βλ. δομή της μυοσφαιρίνης).
 Ρύζι. 1.22. Ολιγομερής δομή της αιμοσφαιρίνης
Ρύζι. 1.22. Ολιγομερής δομή της αιμοσφαιρίνης
Εκτός, Gis E 7εκτελεί ένα σημαντικό πρόσθετο ρόλοστη λειτουργία του Nv. Η ελεύθερη αίμη έχει 25.000 φορές μεγαλύτερη συγγένεια για το CO από ό,τι για το Ο2. Το CO σχηματίζεται σε μικρές ποσότητες στο σώμα και, δεδομένης της υψηλής συγγένειάς του για την αίμη, θα μπορούσε να διαταράξει τη μεταφορά του O 2 που είναι απαραίτητο για τη ζωή των κυττάρων. Ωστόσο, στη σύνθεση της αιμοσφαιρίνης, η συγγένεια της αίμης για το μονοξείδιο του άνθρακα υπερβαίνει τη συγγένεια για το O 2 μόνο κατά 200 φορές λόγω της παρουσίας του His E 7 στο ενεργό κέντρο. Το υπόλοιπο αυτού του αμινοξέος δημιουργεί τις βέλτιστες συνθήκες για τη δέσμευση της αίμης στο O 2 και εξασθενεί την αλληλεπίδραση της αίμης με το CO.
5. Η κύρια λειτουργία του HB είναι η μεταφορά του O2 από τους πνεύμονες στους ιστούς.Σε αντίθεση με τη μονομερή μυοσφαιρίνη, η οποία έχει πολύ υψηλή συγγένεια για το O2 και εκτελεί τη λειτουργία της αποθήκευσης οξυγόνου στους κόκκινους μύες, η ολιγομερής δομή της αιμοσφαιρίνης παρέχει:
1) γρήγορος κορεσμός της HB με οξυγόνο στους πνεύμονες.
2) την ικανότητα του HB να απελευθερώνει οξυγόνο στους ιστούς σε σχετικά υψηλή μερική πίεση O 2 (20-40 mm Hg).
3) η δυνατότητα ρύθμισης της συγγένειας της Hb προς το O 2.
6. Συνεργατικές αλλαγές στη διαμόρφωση των πρωτομερών αιμοσφαιρίνης επιταχύνουν τη δέσμευση του O 2 στους πνεύμονες και την απελευθέρωσή του στους ιστούς. Στους πνεύμονες, η υψηλή μερική πίεση του O 2 προάγει τη δέσμευσή του με Hb στην ενεργό θέση τεσσάρων πρωτομερών (2α και 2β). Το ενεργό κέντρο κάθε πρωτομερούς, όπως και στη μυοσφαιρίνη, βρίσκεται ανάμεσα σε δύο α-έλικες (F και E) σε έναν υδρόφοβο θύλακα. Περιέχει ένα μη πρωτεϊνικό μέρος - αίμη, συνδεδεμένο με το πρωτεϊνικό τμήμα με πολλές ασθενείς υδρόφοβες αλληλεπιδράσεις και έναν ισχυρό δεσμό μεταξύ Fe 2 + αίμη και His F 8 (βλ. Εικ. 1.21).
Στην δεοξυαιμοσφαιρίνη, λόγω αυτού του δεσμού με το His F 8, το άτομο Fe 2 + προεξέχει από το επίπεδο της αίμης προς την ιστιδίνη. Η δέσμευση του O 2 με το Fe 2 + συμβαίνει στην άλλη πλευρά της αίμης στην περιοχή His E 7 χρησιμοποιώντας έναν απλό ελεύθερο δεσμό συντονισμού. Το Ε 7 του παρέχει βέλτιστες συνθήκες για τη δέσμευση του Ο 2 με τον αιμικό σίδηρο.
Η προσθήκη O 2 στο άτομο Fe + 2 ενός πρωτομερούς προκαλεί την κίνησή του στο επίπεδο της αίμης, ακολουθούμενη από το υπόλειμμα ιστιδίνης που σχετίζεται με αυτό
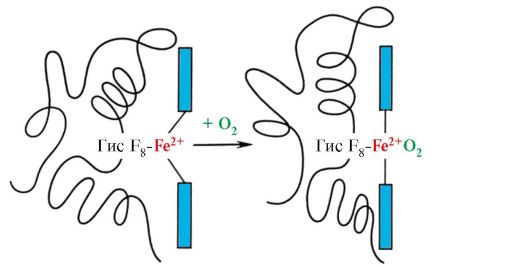 Ρύζι. 1.23. Αλλαγή στη διαμόρφωση του πρωτομερούς αιμοσφαιρίνης όταν συνδυάζεται με O 2
Ρύζι. 1.23. Αλλαγή στη διαμόρφωση του πρωτομερούς αιμοσφαιρίνης όταν συνδυάζεται με O 2
Αυτό οδηγεί σε μια αλλαγή στη διαμόρφωση όλων των πολυπεπτιδικών αλυσίδων λόγω της διαμορφωτικής τους αστάθειας. Η αλλαγή της διαμόρφωσης άλλων αλυσίδων διευκολύνει την αλληλεπίδρασή τους με τα επόμενα μόρια O 2.
Το τέταρτο μόριο O 2 προσκολλάται στην αιμοσφαιρίνη 300 φορές ευκολότερα από το πρώτο (Εικ. 1.24).
 Ρύζι. 1.24. Συνεργατικές αλλαγές στη διαμόρφωση των πρωτομερών αιμοσφαιρίνης κατά την αλληλεπίδρασή της με το Ο2
Ρύζι. 1.24. Συνεργατικές αλλαγές στη διαμόρφωση των πρωτομερών αιμοσφαιρίνης κατά την αλληλεπίδρασή της με το Ο2
Στους ιστούς, κάθε επόμενο μόριο O 2 αποκόπτεται πιο εύκολα από το προηγούμενο, επίσης λόγω συνεργατικών αλλαγών στη διαμόρφωση των πρωτομερών.
7. CO 2 και H+ που σχηματίζονται κατά τον καταβολισμό οργανική ύλη, μειώνουν τη συγγένεια της αιμοσφαιρίνης για το O 2 ανάλογα με τη συγκέντρωσή τους. Η ενέργεια που απαιτείται για τη λειτουργία του κυττάρου παράγεται κυρίως στα μιτοχόνδρια κατά την οξείδωση οργανικών ουσιών με χρήση O 2 που απελευθερώνεται από τους πνεύμονες από την αιμοσφαιρίνη. Ως αποτέλεσμα της οξείδωσης των οργανικών ουσιών, σχηματίζονται τα τελικά προϊόντα της αποσύνθεσής τους: CO 2 και K 2 O, η ποσότητα των οποίων είναι ανάλογη με την ένταση των συνεχιζόμενων διεργασιών οξείδωσης.
Το CO 2 διαχέεται από τα κύτταρα στο αίμα και διεισδύει στα ερυθρά αιμοσφαίρια, όπου, υπό τη δράση του ενζύμου carbanhydrase, μετατρέπεται σε ανθρακικό οξύ. Αυτό το ασθενές οξύ διασπάται σε ένα πρωτόνιο και ένα διττανθρακικό ιόν.
Τα H+ είναι ικανά να ενωθούν με τις ρίζες Του 14 6 στις α- και β-αλυσίδες της αιμοσφαιρίνης, δηλ. σε περιοχές απομακρυσμένες από την αίμη. Η πρωτονίωση της αιμοσφαιρίνης μειώνει τη συγγένειά της για το O 2, προάγει την απομάκρυνση του O 2 από το oxyHb, το σχηματισμό deoxyHb και αυξάνει την παροχή οξυγόνου στους ιστούς αναλογικά με τον αριθμό των πρωτονίων που σχηματίζονται (Εικ. 1.25).
Μια αύξηση στην ποσότητα του απελευθερωμένου οξυγόνου ανάλογα με την αύξηση της συγκέντρωσης Η+ στα ερυθρά αιμοσφαίρια ονομάζεται φαινόμενο Bohr (που πήρε το όνομά του από τον Δανό φυσιολόγο Christian Bohr, ο οποίος ανακάλυψε πρώτος αυτό το φαινόμενο).
Στους πνεύμονες, μια υψηλή μερική πίεση οξυγόνου προάγει τη σύνδεσή του με το deoxyHb, το οποίο μειώνει τη συγγένεια της πρωτεΐνης για το H +. Τα απελευθερωμένα πρωτόνια υπό τη δράση του ανθρακικού οξέος αντιδρούν με διττανθρακικά για να σχηματίσουν CO 2 και H 2 O

 Ρύζι. 1,25. Εξάρτηση της συγγένειας της Hb για το O 2 από τη συγκέντρωση του CO 2 και των πρωτονίων (φαινόμενο Bohr):
Ρύζι. 1,25. Εξάρτηση της συγγένειας της Hb για το O 2 από τη συγκέντρωση του CO 2 και των πρωτονίων (φαινόμενο Bohr):
ΕΝΑ- επίδραση της συγκέντρωσης CO 2 και H+ στην απελευθέρωση O 2 από το σύμπλοκο με HB (φαινόμενο Bohr). σι- οξυγόνωση δεοξυαιμοσφαιρίνης στους πνεύμονες, σχηματισμός και απελευθέρωση CO 2.
Το CO 2 που προκύπτει εισέρχεται στον κυψελιδικό χώρο και απομακρύνεται με εκπνεόμενο αέρα. Έτσι, η ποσότητα οξυγόνου που απελευθερώνεται από την αιμοσφαιρίνη στους ιστούς ρυθμίζεται από τα προϊόντα του καταβολισμού των οργανικών ουσιών: όσο πιο έντονη είναι η διάσπαση των ουσιών, για παράδειγμα κατά τη διάρκεια σωματικής άσκησης, τόσο μεγαλύτερη είναι η συγκέντρωση CO 2 και H + και τόσο περισσότερο οξυγόνο οι ιστοί λαμβάνουν ως αποτέλεσμα της μείωσης της συγγένειας της Hb για το O 2.
8. Αλλοστερική ρύθμιση της συγγένειας της Hb για το O2 από τον συνδέτη - 2,3-διφωσφογλυκερικό.Στα ερυθροκύτταρα, ο αλλοστερικός συνδέτης της αιμοσφαιρίνης, το 2,3-διφωσφογλυκερικό (2,3-BPG), συντίθεται από το προϊόν της οξείδωσης της γλυκόζης - 1,3-διφωσφογλυκερικό. Υπό κανονικές συνθήκες, η συγκέντρωση της 2,3-BPG είναι υψηλή και συγκρίσιμη με τη συγκέντρωση της Hb. Το 2,3-BPG έχει ισχυρό αρνητικό φορτίο -5.
 Το διφωσφογλυκερικό στα τριχοειδή ιστού, που συνδέεται με την δεοξυαιμοσφαιρίνη, αυξάνει την απελευθέρωση οξυγόνου στους ιστούς, μειώνοντας τη συγγένεια της Hb για το O 2.
Το διφωσφογλυκερικό στα τριχοειδή ιστού, που συνδέεται με την δεοξυαιμοσφαιρίνη, αυξάνει την απελευθέρωση οξυγόνου στους ιστούς, μειώνοντας τη συγγένεια της Hb για το O 2.
Στο κέντρο του τετραμερούς μορίου της αιμοσφαιρίνης βρίσκεται μια κοιλότητα. Σχηματίζεται από υπολείμματα αμινοξέων και των τεσσάρων πρωτομερών (βλ. Εικ. 1.22). Στα τριχοειδή των ιστών, η πρωτονίωση της Hb (φαινόμενο Bohr) οδηγεί στη ρήξη του δεσμού μεταξύ του σιδήρου αίμης και του Ο2. Σε ένα μόριο
η δεοξυαιμοσφαιρίνη, σε σύγκριση με την οξυαιμοσφαιρίνη, εμφανίζονται επιπλέον ιοντικοί δεσμοί που συνδέουν τα πρωτομερή, με αποτέλεσμα να αυξάνονται οι διαστάσεις της κεντρικής κοιλότητας σε σχέση με την οξυαιμοσφαιρίνη. Η κεντρική κοιλότητα είναι η θέση σύνδεσης του 2,3-BPG στην αιμοσφαιρίνη. Λόγω της διαφοράς στο μέγεθος της κεντρικής κοιλότητας, το 2,3-BPG μπορεί να προσκολληθεί μόνο στην δεοξυαιμοσφαιρίνη.
Το 2,3-BPG αλληλεπιδρά με την αιμοσφαιρίνη σε μια θέση μακριά από τα ενεργά κέντρα της πρωτεΐνης και ανήκει αλλοστερική(ρυθμιστικοί) συνδέτες, και η κεντρική κοιλότητα της Hb είναι αλλοστερικό κέντρο.Το 2,3-BPG έχει ισχυρό αρνητικό φορτίο και αλληλεπιδρά με πέντε θετικά φορτισμένες ομάδες των δύο β-αλυσίδων της Hb: τη Ν-τελική α-αμινο ομάδα του Val και τις ρίζες Lys 82 His 143 (Εικ. 1.26).
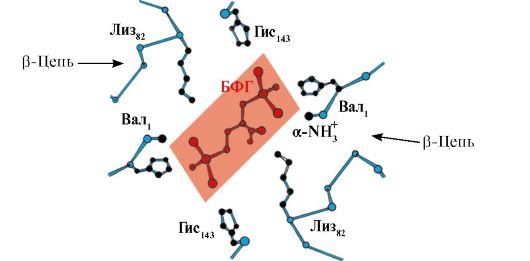 Ρύζι. 1.26. BPG στην κεντρική κοιλότητα της δεοξυαιμοσφαιρίνης
Ρύζι. 1.26. BPG στην κεντρική κοιλότητα της δεοξυαιμοσφαιρίνης
Το BPG συνδέεται με τρεις θετικά φορτισμένες ομάδες σε κάθε β-κλώνο.
Στα τριχοειδή ιστού, η προκύπτουσα δεοξυαιμοσφαιρίνη αλληλεπιδρά με 2,3-BPG και σχηματίζονται ιοντικοί δεσμοί μεταξύ των θετικά φορτισμένων ριζών των β-αλυσίδων και του αρνητικά φορτισμένου συνδέτη, οι οποίοι αλλάζουν τη διαμόρφωση της πρωτεΐνης και μειώνουν τη συγγένεια της Hb για το O2. . Η μείωση της συγγένειας της Hb για το O 2 συμβάλλει στην αποτελεσματικότερη απελευθέρωση του O 2 στον ιστό.
Στους πνεύμονες, σε υψηλή μερική πίεση, το οξυγόνο αλληλεπιδρά με την Hb, ενώνοντας τον αιμικό σίδηρο. Σε αυτή την περίπτωση, η διαμόρφωση της πρωτεΐνης αλλάζει, η κεντρική κοιλότητα μειώνεται και 2,3-BPG μετατοπίζεται από το αλλοστερικό κέντρο
Έτσι, οι ολιγομερείς πρωτεΐνες έχουν νέες ιδιότητες σε σύγκριση με τις μονομερείς πρωτεΐνες. Προσκόλληση προσδεμάτων σε θέσεις
χωρικά απομακρυσμένα το ένα από το άλλο (αλλοστερικό), μπορεί να προκαλέσει αλλαγές διαμόρφωσης σε ολόκληρο το μόριο πρωτεΐνης. Λόγω της αλληλεπίδρασης με ρυθμιστικούς συνδέτες, εμφανίζεται μια αλλαγή στη διαμόρφωση και προσαρμογή της λειτουργίας του μορίου πρωτεΐνης στις περιβαλλοντικές αλλαγές.
ΘΕΜΑ 1.5. ΔΙΑΤΗΡΗΣΗ ΤΗΣ ΕΓΓΕΝΗΣ ΔΙΑΜΟΡΦΩΣΗΣ ΤΩΝ ΠΡΩΤΕΪΝΩΝ ΣΕ ΣΥΝΘΗΚΕΣ ΚΥΤΤΑΡΩΝ
Στα κύτταρα, κατά τη σύνθεση πολυπεπτιδικών αλυσίδων, τη μεταφορά τους μέσω μεμβρανών στα αντίστοιχα μέρη του κυττάρου, κατά τη διαδικασία αναδίπλωσης (σχηματισμός της φυσικής διαμόρφωσης) και κατά τη συναρμολόγηση ολιγομερών πρωτεϊνών, καθώς και κατά τη λειτουργία τους, ενδιάμεσο , επιρρεπείς σε συσσωμάτωση, ασταθείς διαμορφώσεις προκύπτουν στη δομή της πρωτεΐνης. Οι υδρόφοβες ρίζες, συνήθως κρυμμένες μέσα στο πρωτεϊνικό μόριο στην φυσική διαμόρφωση, εμφανίζονται στην επιφάνεια σε ασταθή διαμόρφωση και τείνουν να συνδυάζονται με ομάδες άλλων πρωτεϊνών που είναι ελάχιστα διαλυτές στο νερό. Στα κύτταρα όλων των γνωστών οργανισμών, έχουν βρεθεί ειδικές πρωτεΐνες που εξασφαλίζουν τη βέλτιστη αναδίπλωση των πρωτεϊνών των κυττάρων, σταθεροποιούν τη φυσική τους διαμόρφωση κατά τη λειτουργία και, κυρίως, διατηρούν τη δομή και τις λειτουργίες των ενδοκυτταρικών πρωτεϊνών όταν διαταράσσεται η ομοιόσταση. Αυτές οι πρωτεΐνες ονομάζονται "συνοδοί"που σημαίνει «νταντά» στα γαλλικά.
1. Μοριακές συνοδούς και ο ρόλος τους στην πρόληψη της μετουσίωσης των πρωτεϊνών.
Οι συνοδοί (CH) ταξινομούνται ανάλογα με τη μάζα των υπομονάδων τους. Οι συνοδοί υψηλού μοριακού βάρους έχουν μάζα από 60 έως 110 kDa. Μεταξύ αυτών, τρεις κατηγορίες έχουν μελετηθεί περισσότερο: Sh-60, Sh-70 και Sh-90. Κάθε κατηγορία περιλαμβάνει μια οικογένεια σχετικών πρωτεϊνών. Έτσι, το Sh-70 περιλαμβάνει πρωτεΐνες με μοριακό βάρος από 66 έως 78 kDa. Οι συνοδοί χαμηλού μοριακού βάρους έχουν μοριακό βάρος από 40 έως 15 kDa.
Ανάμεσα στους συνοδούς υπάρχουν συστατικόπρωτεΐνες, η υψηλή βασική σύνθεση των οποίων δεν εξαρτάται από τις επιδράσεις του στρες στα κύτταρα του σώματος και παρακίνητος,η σύνθεση των οποίων υπό κανονικές συνθήκες είναι αδύναμη, αλλά αυξάνεται απότομα υπό πίεση. Οι επαγώγιμες συνοδούς ονομάζονται επίσης «πρωτεΐνες θερμικού σοκ» επειδή ανακαλύφθηκαν για πρώτη φορά σε κύτταρα που εκτέθηκαν σε υψηλές θερμοκρασίες. Στα κύτταρα, λόγω της υψηλής συγκέντρωσης πρωτεϊνών, η αυθόρμητη επανενεργοποίηση των μερικώς μετουσιωμένων πρωτεϊνών είναι δύσκολη. Το Sh-70 μπορεί να αποτρέψει την έναρξη της μετουσίωσης και να βοηθήσει στην αποκατάσταση της φυσικής διαμόρφωσης των πρωτεϊνών. Molecular chaperones-70- μια εξαιρετικά διατηρημένη κατηγορία πρωτεϊνών που βρίσκεται σε όλα τα μέρη του κυττάρου: κυτταρόπλασμα, πυρήνας, ενδοπλασματικό δίκτυο, μιτοχόνδρια. Στο καρβοξυλικό άκρο της μονής πολυπεπτιδικής αλυσίδας Ш-70 υπάρχει μια περιοχή που είναι μια αυλάκωση ικανή να αλληλεπιδρά με πεπτίδια μήκους
από 7 έως 9 υπολείμματα αμινοξέων εμπλουτισμένα με υδρόφοβες ρίζες. Τέτοιες περιοχές στις σφαιρικές πρωτεΐνες εμφανίζονται περίπου κάθε 16 αμινοξέα. Το Sh-70 είναι ικανό να προστατεύει τις πρωτεΐνες από την απενεργοποίηση της θερμοκρασίας και να αποκαθιστά τη διαμόρφωση και τη δραστηριότητα των μερικώς μετουσιωμένων πρωτεϊνών.
2. Ο ρόλος των συνοδών στην αναδίπλωση πρωτεΐνης.Κατά τη διάρκεια της πρωτεϊνικής σύνθεσης στο ριβόσωμα, η Ν-τερματική περιοχή του πολυπεπτιδίου συντίθεται πριν από την C-τερματική. Για να σχηματιστεί η φυσική διαμόρφωση, απαιτείται η πλήρης αλληλουχία αμινοξέων της πρωτεΐνης. Στη διαδικασία της πρωτεϊνοσύνθεσης, οι chaperones-70, λόγω της δομής του ενεργού κέντρου τους, είναι σε θέση να κλείσουν περιοχές του πολυπεπτιδίου που είναι επιρρεπείς σε συσσωμάτωση, εμπλουτισμένες σε υδρόφοβες ρίζες αμινοξέων μέχρι να ολοκληρωθεί η σύνθεση (Εικόνα 1.27, Α. ).
 Ρύζι. 1.27. Συμμετοχή συνοδών στο δίπλωμα πρωτεΐνης
Ρύζι. 1.27. Συμμετοχή συνοδών στο δίπλωμα πρωτεΐνης
Α - συμμετοχή των chaperones-70 στην πρόληψη υδρόφοβων αλληλεπιδράσεων μεταξύ τμημάτων του συντιθέμενου πολυπεπτιδίου. Β - σχηματισμός της φυσικής διαμόρφωσης της πρωτεΐνης στο σύμπλεγμα συνοδών
Πολλές υψηλού μοριακές πρωτεΐνες που έχουν μια πολύπλοκη διαμόρφωση, όπως μια δομή τομέα, αναδιπλώνονται σε έναν ειδικό χώρο που σχηματίζεται από το Sh-60. Ш-60λειτουργούν ως ένα ολιγομερές σύμπλοκο που αποτελείται από 14 υπομονάδες. Σχηματίζουν δύο κοίλους δακτυλίους, καθένας από τους οποίους αποτελείται από επτά υπομονάδες, αυτοί οι δακτύλιοι συνδέονται μεταξύ τους. Κάθε υπομονάδα Sh-60 αποτελείται από τρεις περιοχές: κορυφαία (ακρορριζική), εμπλουτισμένη με υδρόφοβες ρίζες που αντιμετωπίζουν την κοιλότητα του δακτυλίου, ενδιάμεσο και ισημερινό (Εικ. 1.28).
 Ρύζι. 1.28. Δομή του συμπλέγματος chaperonin που αποτελείται από 14 Ш-60
Ρύζι. 1.28. Δομή του συμπλέγματος chaperonin που αποτελείται από 14 Ш-60
A - πλάγια όψη. Β - κάτοψη
Οι συντιθέμενες πρωτεΐνες, οι οποίες έχουν στοιχεία στην επιφάνεια χαρακτηριστικά των ξεδιπλωμένων μορίων, ιδιαίτερα υδρόφοβες ρίζες, εισέρχονται στην κοιλότητα των δακτυλίων συνοδού. Στο συγκεκριμένο περιβάλλον αυτών των κοιλοτήτων, αναζητούνται πιθανές διαμορφώσεις μέχρι να βρεθεί η μόνη που είναι ενεργειακά πιο ευνοϊκή (Εικ. 1.27, Β). Ο σχηματισμός διαμορφώσεων και η απελευθέρωση πρωτεΐνης συνοδεύεται από Υδρόλυση ATPστην περιοχή του Ισημερινού. Τυπικά, μια τέτοια αναδίπλωση που εξαρτάται από τον συνοδό απαιτεί σημαντική ποσότητα ενέργειας.
Εκτός από τη συμμετοχή στο σχηματισμό της τρισδιάστατης δομής των πρωτεϊνών και την αναγέννηση μερικώς μετουσιωμένων πρωτεϊνών, οι συνοδοί είναι επίσης απαραίτητοι για την εμφάνιση θεμελιωδών διεργασιών όπως η συναρμολόγηση ολιγομερών πρωτεϊνών, η αναγνώριση και η μεταφορά μετουσιωμένων πρωτεϊνών στα λυσοσώματα. μεταφορά πρωτεϊνών μέσω των μεμβρανών και συμμετοχή στη ρύθμιση της δραστηριότητας των πρωτεϊνικών συμπλεγμάτων.
ΘΕΜΑ 1.6. ΠΟΙΚΙΛΙΑ ΠΡΩΤΕΪΝΩΝ. ΟΙΚΟΓΕΝΕΙΕΣ ΠΡΩΤΕΪΝΩΝ: ΤΟ ΠΑΡΑΔΕΙΓΜΑ ΤΩΝ ΑΝΟΣΟσφαιρινών
1. Οι πρωτεΐνες παίζουν καθοριστικό ρόλο στη ζωή των μεμονωμένων κυττάρων και ολόκληρου του πολυκύτταρου οργανισμού και οι λειτουργίες τους είναι εκπληκτικά διαφορετικές. Αυτό καθορίζεται από τα χαρακτηριστικά της πρωτογενούς δομής και τις διαμορφώσεις των πρωτεϊνών, τη μοναδική δομή του ενεργού κέντρου και την ικανότητα δέσμευσης συγκεκριμένων προσδεμάτων.
Μόνο ένα πολύ μικρό κλάσμα όλων των πιθανών παραλλαγών των πεπτιδικών αλυσίδων μπορεί να υιοθετήσει μια σταθερή χωρική δομή. η πλειοψηφία
από αυτά μπορούν να λάβουν πολλές διαμορφώσεις με περίπου την ίδια ενέργεια Gibbs, αλλά με διαφορετικές ιδιότητες. Η πρωτογενής δομή των πιο γνωστών πρωτεϊνών που επιλέγονται από τη βιολογική εξέλιξη εξασφαλίζει εξαιρετική σταθερότητα μιας από τις διαμορφώσεις, η οποία καθορίζει τα χαρακτηριστικά της λειτουργίας αυτής της πρωτεΐνης.
2. Πρωτεϊνικές οικογένειες.Μέσα στο ίδιο βιολογικό είδος, οι υποκαταστάσεις υπολειμμάτων αμινοξέων μπορούν να οδηγήσουν στην εμφάνιση διαφορετικών πρωτεϊνών που εκτελούν σχετικές λειτουργίες και έχουν ομόλογες αλληλουχίεςαμινοξέα. Τέτοιες σχετικές πρωτεΐνες έχουν εντυπωσιακά παρόμοιες διαμορφώσεις: ο αριθμός και οι σχετικές θέσεις των α-έλικων και/ή των β-δομών και οι περισσότερες στροφές και στροφές των πολυπεπτιδικών αλυσίδων είναι παρόμοιες ή πανομοιότυπες. Πρωτεΐνες με ομόλογες περιοχές της πολυπεπτιδικής αλυσίδας, παρόμοια διαμόρφωση και σχετικές λειτουργίες ταξινομούνται σε οικογένειες πρωτεϊνών. Παραδείγματα οικογενειών πρωτεϊνών: πρωτεϊνάσες σερίνης, οικογένεια ανοσοσφαιρινών, οικογένεια μυοσφαιρίνης.
Πρωτεϊνάσες σερίνης- μια οικογένεια πρωτεϊνών που εκτελούν τη λειτουργία των πρωτεολυτικών ενζύμων. Αυτά περιλαμβάνουν πεπτικά ένζυμα - χυμοθρυψίνη, θρυψίνη, ελαστάση και πολλούς παράγοντες πήξης του αίματος. Αυτές οι πρωτεΐνες έχουν πανομοιότυπα αμινοξέα στο 40% των θέσεων τους και πολύ παρόμοια διαμόρφωση (Εικ. 1.29).

Ρύζι. 1.29. Χωρικές δομέςελαστάση (Α) και χυμοθρυψίνη (Β)
Ορισμένες υποκαταστάσεις αμινοξέων έχουν οδηγήσει σε αλλαγές στην εξειδίκευση του υποστρώματος αυτών των πρωτεϊνών και στην εμφάνιση λειτουργικής ποικιλομορφίας εντός της οικογένειας.
3. Οικογένεια ανοσοσφαιρινών.Σε εξέλιξη ανοσοποιητικό σύστημαΤεράστιο ρόλο παίζουν οι πρωτεΐνες της υπεροικογένειας των ανοσοσφαιρινών, η οποία περιλαμβάνει τρεις οικογένειες πρωτεϊνών:
Αντισώματα (ανοσοσφαιρίνες);
Υποδοχείς Τ-λεμφοκυττάρων;
Πρωτεΐνες του κύριου συμπλέγματος ιστοσυμβατότητας - MHC κλάσεις 1 και 2 (Major Histocompatibility Complex).
Όλες αυτές οι πρωτεΐνες έχουν δομή πεδίου, αποτελούνται από ομόλογες περιοχές που μοιάζουν με ανοσοποιητικό και εκτελούν παρόμοιες λειτουργίες: αλληλεπιδρούν με ξένες δομές, είτε διαλυμένες στο αίμα, λέμφο ή μεσοκυττάριο υγρό (αντισώματα), είτε βρίσκονται στην επιφάνεια των κυττάρων (δικά τους ή ξένο).
4. Αντισώματα- ειδικές πρωτεΐνες που παράγονται από τα Β λεμφοκύτταρα ως απόκριση στην είσοδο μιας ξένης δομής στο σώμα, που ονομάζονται αντιγόνο.
Χαρακτηριστικά της δομής των αντισωμάτων
Τα απλούστερα μόρια αντισωμάτων αποτελούνται από τέσσερις πολυπεπτιδικές αλυσίδες: δύο πανομοιότυπες ελαφριές - L, που περιέχουν περίπου 220 αμινοξέα, και δύο πανομοιότυπες βαριές - Η, που αποτελούνται από 440-700 αμινοξέα. Και οι τέσσερις αλυσίδες στο μόριο του αντισώματος συνδέονται με πολλούς μη ομοιοπολικούς δεσμούς και τέσσερις δισουλφιδικούς δεσμούς (Εικ. 1.30).
Οι ελαφριές αλυσίδες αντισωμάτων αποτελούνται από δύο τομείς: μια μεταβλητή περιοχή (VL), που βρίσκεται στη Ν-τελική περιοχή της πολυπεπτιδικής αλυσίδας και μια σταθερή περιοχή (CL), που βρίσκεται στο C-άκρο. Οι βαριές αλυσίδες έχουν συνήθως τέσσερις τομείς: μία μεταβλητή (VH), που βρίσκεται στο Ν-άκρο, και τρεις σταθερές περιοχές (CH1, CH2, CH3) (βλ. Εικ. 1.30). Κάθε τομέας ανοσοσφαιρίνης έχει μια υπερδομή β-φύλλου στην οποία δύο υπολείμματα κυστεΐνης συνδέονται με έναν δισουλφιδικό δεσμό.
Μεταξύ των δύο σταθερών περιοχών CH1 και CH2 υπάρχει μια περιοχή που περιέχει μεγάλο αριθμό υπολειμμάτων προλίνης, τα οποία εμποδίζουν το σχηματισμό μιας δευτερεύουσας δομής και την αλληλεπίδραση γειτονικών αλυσίδων Η σε αυτό το τμήμα. Αυτή η περιοχή άρθρωσης δίνει στο μόριο του αντισώματος ευελιξία. Μεταξύ των μεταβλητών περιοχών της βαριάς και της ελαφριάς αλυσίδας υπάρχουν δύο πανομοιότυπες θέσεις δέσμευσης αντιγόνου (ενεργές θέσεις για δέσμευση αντιγόνων), επομένως τέτοια αντισώματα ονομάζονται συχνά δισθενείς.Δεν εμπλέκεται ολόκληρη η αλληλουχία αμινοξέων των μεταβλητών περιοχών και των δύο αλυσίδων στη σύνδεση του αντιγόνου με το αντίσωμα, αλλά μόνο 20-30 αμινοξέα που βρίσκονται στις υπερμεταβλητές περιοχές κάθε αλυσίδας. Αυτές οι περιοχές είναι που καθορίζουν τη μοναδική ικανότητα κάθε τύπου αντισώματος να αλληλεπιδρά με το αντίστοιχο συμπληρωματικό αντιγόνο.
Τα αντισώματα είναι μια από τις γραμμές άμυνας του οργανισμού ενάντια σε εισβολείς ξένων οργανισμών. Η λειτουργία τους μπορεί να χωριστεί σε δύο στάδια: το πρώτο στάδιο είναι η αναγνώριση και η δέσμευση του αντιγόνου στην επιφάνεια ξένων οργανισμών, η οποία είναι δυνατή λόγω της παρουσίας θέσεων δέσμευσης αντιγόνου στη δομή του αντισώματος. το δεύτερο στάδιο είναι η έναρξη της διαδικασίας αδρανοποίησης και καταστροφής του αντιγόνου. Η ειδικότητα του δεύτερου σταδίου εξαρτάται από την κατηγορία των αντισωμάτων. Υπάρχουν πέντε κατηγορίες βαριών αλυσίδων, που διαφέρουν μεταξύ τους ως προς τη δομή των σταθερών περιοχών: α, δ, ε, γ και μ, σύμφωνα με τις οποίες διακρίνονται πέντε κατηγορίες ανοσοσφαιρινών: A, D, E, G και M.
Τα δομικά χαρακτηριστικά των βαριών αλυσίδων δίνουν στις περιοχές άρθρωσης και στις C-τερματικές περιοχές των βαριών αλυσίδων ένα χαρακτηριστικό διαμόρφωσης κάθε κατηγορίας. Μετά τη σύνδεση του αντιγόνου σε ένα αντίσωμα, οι διαμορφωτικές αλλαγές στις σταθερές περιοχές καθορίζουν την οδό για την απομάκρυνση του αντιγόνου.
 Ρύζι. 1. 30. Δομή τομέα IgG
Ρύζι. 1. 30. Δομή τομέα IgG
Ανοσοσφαιρίνες Μ
Οι ανοσοσφαιρίνες Μ έχουν δύο μορφές.
Μονομερής μορφή- 1η κατηγορία αντισωμάτων που παράγονται από την ανάπτυξη Β λεμφοκυττάρων. Στη συνέχεια, πολλά Β κύτταρα μεταπηδούν στην παραγωγή άλλων τάξεων αντισωμάτων, αλλά με την ίδια θέση δέσμευσης αντιγόνου. Το IgM είναι ενσωματωμένο στη μεμβράνη και δρα ως υποδοχέας αναγνώρισης αντιγόνου. Η ενσωμάτωση του IgM στην κυτταρική μεμβράνη είναι δυνατή λόγω της παρουσίας 25 υδρόφοβων υπολειμμάτων αμινοξέων στο τμήμα της ουράς της περιοχής.
Εκκριτική μορφή IgMπεριέχει πέντε μονομερείς υπομονάδες συνδεδεμένες μεταξύ τους με δισουλφιδικούς δεσμούς και μια επιπλέον αλυσίδα πολυπεπτιδίου J (Εικ. 1.31). Οι βαριές αλυσίδες μονομερών αυτής της μορφής δεν περιέχουν υδρόφοβη ουρά. Το πενταμερές έχει 10 θέσεις δέσμευσης αντιγόνου και επομένως είναι αποτελεσματικό στην αναγνώριση και απομάκρυνση του αντιγόνου που εισέρχεται πρώτο στο σώμα. Η εκκριτική μορφή του IgM είναι η κύρια κατηγορία αντισωμάτων που εκκρίνονται στο αίμα κατά την πρωτογενή ανοσοαπόκριση. Η σύνδεση του IgM σε ένα αντιγόνο αλλάζει τη διαμόρφωση του IgM και προκαλεί τη δέσμευσή του στο πρώτο συστατικό πρωτεΐνης του συστήματος συμπληρώματος (το σύστημα συμπληρώματος είναι ένα σύνολο πρωτεϊνών που εμπλέκονται στην καταστροφή του αντιγόνου) και την ενεργοποίηση αυτού του συστήματος. Εάν το αντιγόνο βρίσκεται στην επιφάνεια ενός μικροοργανισμού, το σύστημα συμπληρώματος προκαλεί παραβίαση της ακεραιότητας κυτταρική μεμβράνηκαι θάνατο του βακτηριακού κυττάρου.
Ανοσοσφαιρίνες G
Ποσοτικά, αυτή η κατηγορία ανοσοσφαιρινών κυριαρχεί στο αίμα (75% όλων των Igs). IgG - μονομερή, η κύρια κατηγορία αντισωμάτων που εκκρίνονται στο αίμα κατά τη διάρκεια μιας δευτερογενούς ανοσοαπόκρισης. Μετά την αλληλεπίδραση του IgG με τα επιφανειακά αντιγόνα των μικροοργανισμών, το σύμπλεγμα αντιγόνου-αντισώματος είναι σε θέση να δεσμεύει και να ενεργοποιεί πρωτεΐνες του συστήματος συμπληρώματος ή μπορεί να αλληλεπιδρά με συγκεκριμένους υποδοχείς μακροφάγων και ουδετερόφιλων. Η αλληλεπίδραση με τα φαγοκύτταρα οδηγεί
 Ρύζι. 1.31. Δομή της εκκριτικής μορφής του IgM
Ρύζι. 1.31. Δομή της εκκριτικής μορφής του IgM
στην απορρόφηση συμπλεγμάτων αντιγόνου-αντισώματος και στην καταστροφή τους στα κυτταρικά φαγοσώματα. Το IgG είναι η μόνη κατηγορία αντισωμάτων που είναι ικανά να διαπεράσουν τον φραγμό του πλακούντα και να παρέχουν ενδομήτρια προστασία του εμβρύου από λοιμώξεις.
Ανοσοσφαιρίνες Α
Η κύρια κατηγορία αντισωμάτων που υπάρχουν στις εκκρίσεις (γάλα, σάλιο, εκκρίσεις της αναπνευστικής οδού και του εντερικού σωλήνα). Το IgA εκκρίνεται κυρίως σε διμερή μορφή, όπου τα μονομερή συνδέονται μεταξύ τους μέσω μιας πρόσθετης αλυσίδας J (Εικ. 1.32).
Το IgA δεν αλληλεπιδρά με το σύστημα του συμπληρώματος και τα φαγοκυτταρικά κύτταρα, αλλά δεσμεύοντας σε μικροοργανισμούς, τα αντισώματα εμποδίζουν την προσκόλλησή τους στα επιθηλιακά κύτταρα και τη διείσδυσή τους στο σώμα.
Ανοσοσφαιρίνες Ε
Οι ανοσοσφαιρίνες Ε αντιπροσωπεύονται από μονομερή στα οποία οι βαριές ε-αλυσίδες περιέχουν, όπως και οι μ-αλυσίδες των ανοσοσφαιρινών Μ, μία μεταβλητή και τέσσερις σταθερές περιοχές. Μετά την έκκριση, η IgE συνδέεται με αυτήν
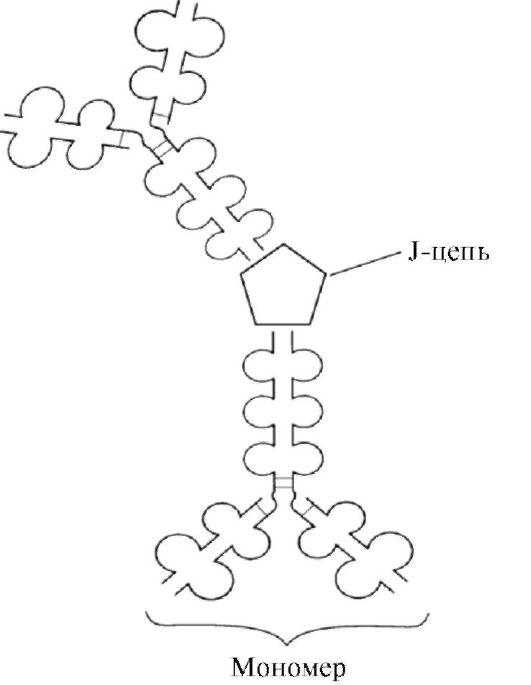 Ρύζι. 1.32. Δομή της IgA
Ρύζι. 1.32. Δομή της IgA
C-τελικές περιοχές με αντίστοιχους υποδοχείς στην επιφάνεια των ιστιοκυττάρων και των βασεόφιλων. Ως αποτέλεσμα, γίνονται υποδοχείς για αντιγόνα στην επιφάνεια αυτών των κυττάρων (Εικ. 1.33).
 Ρύζι. 1.33. Αλληλεπίδραση IgE με αντιγόνο στην επιφάνεια ενός ιστιοκυττάρου
Ρύζι. 1.33. Αλληλεπίδραση IgE με αντιγόνο στην επιφάνεια ενός ιστιοκυττάρου
Αφού το αντιγόνο συνδεθεί με τις αντίστοιχες θέσεις δέσμευσης αντιγόνου της IgE, τα κύτταρα λαμβάνουν ένα σήμα για να εκκρίνουν βιολογικά δραστικές ουσίες(ισταμίνη, σεροτονίνη), που ευθύνονται σε μεγάλο βαθμό για την ανάπτυξη της φλεγμονώδους αντίδρασης και για την εκδήλωση αλλεργικών αντιδράσεων όπως το άσθμα, η κνίδωση, ο αλλεργικός πυρετός.
Ανοσοσφαιρίνες Δ
Οι ανοσοσφαιρίνες D βρίσκονται σε πολύ μικρές ποσότητες στον ορό· είναι μονομερή. Οι βαριές δ αλυσίδες έχουν μία μεταβλητή και τρεις σταθερές περιοχές. Τα IgD δρουν ως υποδοχείς για τα Β λεμφοκύτταρα· άλλες λειτουργίες είναι ακόμη άγνωστες. Η αλληλεπίδραση ειδικών αντιγόνων με υποδοχείς στην επιφάνεια των Β-λεμφοκυττάρων (IgD) οδηγεί στη μετάδοση αυτών των σημάτων στο κύτταρο και στην ενεργοποίηση μηχανισμών που διασφαλίζουν τον πολλαπλασιασμό ενός δεδομένου κλώνου λεμφοκυττάρων.
ΘΕΜΑ 1.7. ΦΥΣΙΚΕΣ ΚΑΙ ΧΗΜΙΚΕΣ ΙΔΙΟΤΗΤΕΣ ΤΩΝ ΠΡΩΤΕΪΝΩΝ ΚΑΙ ΜΕΘΟΔΟΙ ΓΙΑ ΤΟ ΔΙΑΧΩΡΙΣΜΟ ΤΟΥΣ
1. Οι μεμονωμένες πρωτεΐνες διαφέρουν ως προς τις φυσικές και χημικές ιδιότητες:
Σχήμα μορίων;
Μοριακό βάρος;
Το συνολικό φορτίο, το μέγεθος του οποίου εξαρτάται από την αναλογία ανιονικών και κατιονικών ομάδων αμινοξέων.
Η αναλογία πολικών και μη πολικών ριζών αμινοξέων στην επιφάνεια των μορίων.
Βαθμοί αντοχής σε διάφορους μετουσιωτικούς παράγοντες.
2. Η διαλυτότητα της πρωτεΐνης εξαρτάταιστις ιδιότητες των πρωτεϊνών που αναφέρονται παραπάνω, καθώς και στη σύνθεση του μέσου στο οποίο διαλύεται η πρωτεΐνη (τιμές pH, σύνθεση άλατος, θερμοκρασία, παρουσία άλλων οργανικών ουσιών που μπορούν να αλληλεπιδράσουν με την πρωτεΐνη). Η ποσότητα φορτίου των μορίων πρωτεΐνης είναι ένας από τους παράγοντες που επηρεάζουν τη διαλυτότητά τους. Όταν το φορτίο στο ισοηλεκτρικό σημείο χαθεί, οι πρωτεΐνες συσσωματώνονται και καθιζάνουν ευκολότερα. Αυτό είναι ιδιαίτερα χαρακτηριστικό για μετουσιωμένες πρωτεΐνες, στις οποίες εμφανίζονται υδρόφοβες ρίζες αμινοξέων στην επιφάνεια.
Στην επιφάνεια ενός μορίου πρωτεΐνης υπάρχουν τόσο θετικά όσο και αρνητικά φορτισμένες ρίζες αμινοξέων. Ο αριθμός αυτών των ομάδων, και επομένως το συνολικό φορτίο των πρωτεϊνών, εξαρτάται από το pH του μέσου, δηλ. αναλογία συγκεντρώσεων ομάδων Η+ - και ΟΗ -. Σε όξινο περιβάλλονΗ αύξηση της συγκέντρωσης Η+ οδηγεί σε καταστολή της διάστασης των καρβοξυλομάδων -COO - + H+ > - COOH και σε μείωση του αρνητικού φορτίου των πρωτεϊνών. Σε αλκαλικό περιβάλλον, η δέσμευση περίσσειας ΟΗ - από πρωτόνια που σχηματίζονται κατά τη διάσταση των αμινομάδων -NH 3 + + OH - - NH 2 + H 2 O με το σχηματισμό νερού, οδηγεί σε μείωση του θετικού φορτίου των πρωτεϊνών . Η τιμή pH στην οποία μια πρωτεΐνη έχει καθαρό μηδενικό φορτίο ονομάζεται ισοηλεκτρικό σημείο (IEP).Στο IET, ο αριθμός των θετικά και αρνητικά φορτισμένων ομάδων είναι ο ίδιος, δηλ. η πρωτεΐνη βρίσκεται σε ισοηλεκτρική κατάσταση.
3. Διαχωρισμός μεμονωμένων πρωτεϊνών.Τα χαρακτηριστικά της δομής και της λειτουργίας του σώματος εξαρτώνται από το σύνολο των πρωτεϊνών που συντίθενται σε αυτό. Η μελέτη της δομής και των ιδιοτήτων των πρωτεϊνών είναι αδύνατη χωρίς την απομόνωση τους από το κύτταρο και τον καθαρισμό τους από άλλες πρωτεΐνες και οργανικά μόρια. Στάδια απομόνωσης και καθαρισμού μεμονωμένων πρωτεϊνών:
Καταστροφή κυττάρωνο ιστός που μελετάται και λαμβάνεται ένα ομογενοποιημένο προϊόν.
Διαχωρισμός του ομογενοποιημένου προϊόντος σε κλάσματαμε φυγοκέντρηση, λαμβάνοντας ένα πυρηνικό, μιτοχονδριακό, κυτοσολικό ή άλλο κλάσμα που περιέχει την επιθυμητή πρωτεΐνη.
Επιλεκτική θερμική μετουσίωση- βραχυπρόθεσμη θέρμανση ενός διαλύματος πρωτεΐνης, κατά την οποία ορισμένες από τις μετουσιωμένες πρωτεϊνικές ακαθαρσίες μπορούν να αφαιρεθούν (εάν η πρωτεΐνη είναι σχετικά σταθερή στη θερμότητα).
Αλάτισμα έξω.Διαφορετικές πρωτεΐνες καθιζάνουν σε διαφορετικές συγκεντρώσεις άλατος στο διάλυμα. Αυξάνοντας σταδιακά τη συγκέντρωση του άλατος, είναι δυνατό να ληφθεί ένας αριθμός ξεχωριστών κλασμάτων με κυρίαρχη περιεκτικότητα σε απομονωμένη πρωτεΐνη σε ένα από αυτά. Το θειικό αμμώνιο χρησιμοποιείται συχνότερα για κλασματοποίηση πρωτεϊνών. Οι πρωτεΐνες με τη μικρότερη διαλυτότητα καθιζάνουν σε χαμηλές συγκεντρώσεις αλάτων.
Διήθηση γέλης- μέθοδος κοσκινίσματος μορίων μέσω διογκωμένων κόκκων Sephadex (τρισδιάστατες πολυσακχαριτικές αλυσίδες δεξτράνης με πόρους). Η ταχύτητα με την οποία οι πρωτεΐνες περνούν μέσα από μια στήλη γεμάτη με Sephadex θα εξαρτηθεί από το μοριακό τους βάρος: όσο μικρότερη είναι η μάζα των μορίων πρωτεΐνης, τόσο πιο εύκολα διεισδύουν στους κόκκους και παραμένουν εκεί περισσότερο· όσο μεγαλύτερη είναι η μάζα, τόσο πιο γρήγορα εκλούονται από στήλη.
Υπερφυγοκέντρηση- μια μέθοδος που περιλαμβάνει την τοποθέτηση πρωτεϊνών σε ένα σωλήνα φυγοκέντρησης στον ρότορα μιας υπερφυγοκέντρου. Όταν ο ρότορας περιστρέφεται, ο ρυθμός καθίζησης των πρωτεϊνών είναι ανάλογος με το μοριακό τους βάρος: κλάσματα βαρύτερων πρωτεϊνών βρίσκονται πιο κοντά στον πυθμένα του δοκιμαστικού σωλήνα, ελαφρύτερα - πιο κοντά στην επιφάνεια.
Ηλεκτροφόρηση- μια μέθοδος που βασίζεται σε διαφορές στην ταχύτητα κίνησης των πρωτεϊνών σε ένα ηλεκτρικό πεδίο. Αυτή η τιμή είναι ανάλογη με το φορτίο των πρωτεϊνών. Η ηλεκτροφόρηση των πρωτεϊνών πραγματοποιείται σε χαρτί (σε αυτή την περίπτωση, η ταχύτητα της κίνησης των πρωτεϊνών είναι ανάλογη μόνο με το φορτίο τους) ή σε γέλη πολυακρυλαμιδίου με συγκεκριμένο μέγεθος πόρων (η ταχύτητα κίνησης της πρωτεΐνης είναι ανάλογη με το φορτίο και το μοριακό τους βάρος) .
Χρωματογραφία ανταλλαγής ιόντων- μέθοδος κλασματοποίησης που βασίζεται στη δέσμευση ιονισμένων ομάδων πρωτεϊνών με αντίθετα φορτισμένες ομάδες ρητινών ανταλλαγής ιόντων (αδιάλυτα πολυμερή υλικά). Η δύναμη της δέσμευσης της πρωτεΐνης με τη ρητίνη είναι ανάλογη με το φορτίο της πρωτεΐνης. Οι πρωτεΐνες που έχουν προσροφηθεί στο πολυμερές ανταλλαγής ιόντων μπορούν να ξεπλυθούν με αυξανόμενες συγκεντρώσεις διαλυμάτων NaCl. Όσο χαμηλότερο είναι το φορτίο πρωτεΐνης, τόσο χαμηλότερη είναι η συγκέντρωση του NaCl που απαιτείται για να ξεπλυθεί η πρωτεΐνη που είναι δεσμευμένη στις ιοντικές ομάδες της ρητίνης.
Χρωματογραφία συγγένειας- η πιο ειδική μέθοδος για την απομόνωση μεμονωμένων πρωτεϊνών Ένας συνδέτης μιας πρωτεΐνης συνδέεται ομοιοπολικά σε ένα αδρανές πολυμερές. Όταν ένα διάλυμα πρωτεΐνης διέρχεται μέσω μιας στήλης με ένα πολυμερές, μόνο η πρωτεΐνη που είναι ειδική για ένα δεδομένο πρόσδεμα προσροφάται στη στήλη λόγω της συμπληρωματικής δέσμευσης της πρωτεΐνης στον συνδέτη.
Διάλυση- μέθοδος που χρησιμοποιείται για την αφαίρεση ενώσεων χαμηλού μοριακού βάρους από διάλυμα απομονωμένης πρωτεΐνης. Η μέθοδος βασίζεται στην αδυναμία των πρωτεϊνών να περάσουν από μια ημιπερατή μεμβράνη, σε αντίθεση με τις ουσίες χαμηλού μοριακού βάρους. Χρησιμοποιείται για τον καθαρισμό πρωτεϊνών από ακαθαρσίες χαμηλού μοριακού βάρους, για παράδειγμα, άλατα μετά το αλάτισμα.
ΕΡΓΑΣΙΕΣ ΕΞΩΔΕΙΚΤΙΚΩΝ ΕΡΓΑΣΙΩΝ
1. Συμπληρώστε τον πίνακα. 1.4.
Πίνακας 1.4. Συγκριτική ανάλυσηδομή και λειτουργίες των σχετικών πρωτεϊνών - μυοσφαιρίνη και αιμοσφαιρίνη
α) θυμηθείτε τη δομή του ενεργού κέντρου των Mb και Hb. Τι ρόλο παίζουν οι υδρόφοβες ρίζες αμινοξέων στο σχηματισμό των ενεργών κέντρων αυτών των πρωτεϊνών; Περιγράψτε τη δομή του ενεργού κέντρου των Mb και Hb και τους μηχανισμούς προσκόλλησης των προσδεμάτων σε αυτό. Τι ρόλο παίζουν τα υπολείμματα His F 8 και His E 7 στη λειτουργία του ενεργού κέντρου του Mv iHv;
β) ποιες νέες ιδιότητες σε σύγκριση με τη μονομερή μυοσφαιρίνη έχει η στενά συγγενής ολιγομερής πρωτεΐνη, η αιμοσφαιρίνη; Εξηγήστε τον ρόλο των αλλαγών συνεργασίας στη διαμόρφωση των πρωτομερών στο μόριο της αιμοσφαιρίνης, την επίδραση των συγκεντρώσεων CO 2 και πρωτονίων στη συγγένεια της αιμοσφαιρίνης για το οξυγόνο, καθώς και τον ρόλο της 2,3-BPG στην αλλοστερική ρύθμιση της λειτουργίας της Hb .
2. Χαρακτηρίστε τους μοριακούς συνοδούς, δίνοντας προσοχή στη σχέση μεταξύ της δομής και της λειτουργίας τους.
3. Ποιες πρωτεΐνες ομαδοποιούνται σε οικογένειες; Χρησιμοποιώντας το παράδειγμα της οικογένειας των ανοσοσφαιρινών, αναγνωρίστε παρόμοια δομικά χαρακτηριστικά και σχετικές λειτουργίες των πρωτεϊνών αυτής της οικογένειας.
4. Συχνά απαιτούνται καθαρισμένες μεμονωμένες πρωτεΐνες για βιοχημικούς και ιατρικούς σκοπούς. Εξηγήστε ποια ΦΥΣΙΚΕΣ ΚΑΙ ΧΗΜΙΚΕΣ ΙΔΙΟΤΗΤΕΣΟι πρωτεΐνες βασίζονται στις μεθόδους που χρησιμοποιούνται για τον διαχωρισμό και τον καθαρισμό τους.
ΚΑΘΗΚΟΝΤΑ ΑΥΤΟΕΛΕΓΧΟΥ
1. Επιλέξτε τις σωστές απαντήσεις.
Λειτουργίες της αιμοσφαιρίνης:
Α. Μεταφορά Ο 2 από τους πνεύμονες στον ιστό Β. Μεταφορά Η + από τους ιστούς στους πνεύμονες
Β. Διατήρηση σταθερού pH αίματος Δ. Μεταφορά CO 2 από τους πνεύμονες στους ιστούς
Δ. Μεταφορά CO 2 από τους ιστούς στους πνεύμονες
2. Επιλέξτε τις σωστές απαντήσεις. Ligandα - Το πρωτομερές Hb είναι:Α. Heme
Β. Οξυγόνο
Β. CO G. 2,3-BPG
D. β-Πρωτομερές
3. Επιλέξτε τις σωστές απαντήσεις.
Αιμοσφαιρίνη σε αντίθεση με μυοσφαιρίνη:
Α. Έχει τεταρτογενή δομή
Β. Η δευτερεύουσα δομή αντιπροσωπεύεται μόνο από α-έλικες
Β. Ανήκει σε σύνθετες πρωτεΐνες
Δ. Αλληλεπιδρά με έναν αλλοστερικό συνδέτη Δ. Ομοιοπολικά δεσμευμένος στην αίμη
4. Επιλέξτε τις σωστές απαντήσεις.
Η συγγένεια της Hb για το O2 μειώνεται:
Α. Όταν προστίθεται ένα μόριο Ο 2 Β. Όταν αφαιρείται ένα μόριο Ο 2
Β. Κατά την αλληλεπίδραση με 2,3-BPG
Δ. Όταν προσκολλάται σε πρωτομερή H + D. Όταν μειώνεται η συγκέντρωση του 2,3-BPG
5. Αγώνας.
Οι τύποι HB χαρακτηρίζονται από:
Α. Σε δεοξυ μορφή σχηματίζει ινώδη συσσωματώματα Β. Περιέχει δύο α- και δύο δ-αλυσίδες
Β. Η κυρίαρχη μορφή Hb σε ερυθροκύτταρα ενηλίκων Δ. Περιέχει αίμη με Fe+ 3 στο ενεργό κέντρο
Δ. Περιέχει δύο α- και δύο γ-αλυσίδες 1. HbA 2.
6. Αγώνας.
Υποκαταστάτες Hb:
Α. Προσδένεται στην Hb στο αλλοστερικό κέντρο
Β. Έχει πολύ υψηλή συγγένεια με τη δραστική θέση της Hb
Β. Με την ένωση, αυξάνει τη συγγένεια της Hb για το O 2 G. Οξειδώνει το Fe+ 2 σε Fe+ 3
Δ. Έντυπα ομοιοπολικό δεσμόμε gisF8
7. Επιλέξτε τις σωστές απαντήσεις.
Συνοδοιπόροι:
Α. Πρωτεΐνες που υπάρχουν σε όλα τα μέρη του κυττάρου
Β. Η σύνθεση αυξάνεται υπό πίεση
Β. Συμμετέχουν στην υδρόλυση μετουσιωμένων πρωτεϊνών
Δ. Συμμετέχετε στη διατήρηση της φυσικής διαμόρφωσης των πρωτεϊνών
Δ. Δημιουργούν οργανίδια στα οποία σχηματίζεται η διαμόρφωση των πρωτεϊνών.
8. Ταίριασμα. Ανοσοσφαιρίνες:
Α. Η εκκριτική μορφή είναι πενταμερής.
Β. Κατηγορία Ig που διαπερνά τον φραγμό του πλακούντα
Β. Ig - υποδοχέας μαστοκυττάρων
Δ. Η κύρια κατηγορία Ig που υπάρχει στις εκκρίσεις των επιθηλιακών κυττάρων. Δ. Υποδοχέας Β-λεμφοκυττάρων, η ενεργοποίηση του οποίου διασφαλίζει τον πολλαπλασιασμό των κυττάρων
9. Επιλέξτε τις σωστές απαντήσεις.
Ανοσοσφαιρίνες Ε:
Α. Παράγεται από μακροφάγα Β. Έχουν βαριές ε αλυσίδες.
Β. Ενσωματωμένο στη μεμβράνη των Τ-λεμφοκυττάρων
Δ. Λειτουργούν ως υποδοχείς αντιγόνων μεμβράνης σε μαστοκύτταρα και βασεόφιλα
Δ. Υπεύθυνος για αλλεργικές αντιδράσεις
10. Επιλέξτε τις σωστές απαντήσεις.
Η μέθοδος για τον διαχωρισμό των πρωτεϊνών βασίζεται στις διαφορές στο μοριακό τους βάρος:
Α. Διήθηση γέλης
Β. Υπερφυγοκέντρηση
Β. Ηλεκτροφόρηση γέλης πολυακρυλαμιδίου Δ. Χρωματογραφία ανταλλαγής ιόντων
Δ. Χρωματογραφία συγγένειας
11. Διάλεξε την σωστή απάντηση.
Η μέθοδος διαχωρισμού πρωτεϊνών βασίζεται στις διαφορές στη διαλυτότητά τους στο νερό:
Α. Διήθηση γέλης Β. Αλάτισμα
Β. Χρωματογραφία ανταλλαγής ιόντων Δ. Χρωματογραφία συγγένειας
Δ. Ηλεκτροφόρηση σε γέλη πολυακρυλαμιδίου
ΠΡΟΤΥΠΑ ΑΠΑΝΤΗΣΕΩΝ ΣΤΙΣ ΕΡΓΑΣΙΕΣ ΑΥΤΟΕΛΕΓΧΟΥ
1. Α, Β, Γ, Δ
2. Α, Β, Γ, Δ
5. 1-B, 2-A, 3-G
6. 1-B, 2-B, 3-A
7. Α, Β, Δ, Δ
8. 1-G; 2-Β, 3-Β
ΒΑΣΙΚΟΙ ΟΡΟΙ ΚΑΙ ΕΝΝΟΙΕΣ
1. Ολιγομερείς πρωτεΐνες, πρωτομερές, τεταρτοταγής δομή πρωτεϊνών
2. Συνεργατικές αλλαγές στη διαμόρφωση του πρωτομερούς
3. Φαινόμενο Bohr
4. Αλλοστερική ρύθμιση πρωτεϊνικών λειτουργιών, αλλοστερικό κέντρο και αλλοστερικός τελεστής
5. Μοριακές συνοδούς, πρωτεΐνες θερμικού σοκ
6. Οικογένειες πρωτεϊνών (πρωτεάσες σερίνης, ανοσοσφαιρίνες)
7. Σχέση IgM-, G-, E-, A-δομής-συνάρτησης
8. Ολικό φορτίο πρωτεϊνών, ισοηλεκτρικό σημείο πρωτεϊνών
9. Ηλεκτροφόρηση
10. Αλάτισμα έξω
11. Διήθηση γέλης
12. Χρωματογραφία ανταλλαγής ιόντων
13. Υπερφυγοκέντρηση
14. Χρωματογραφία συγγένειας
15. Ηλεκτροφόρηση πρωτεϊνών πλάσματος αίματος
ΕΡΓΑΣΙΕΣ ΓΙΑ ΕΡΓΑΣΙΕΣ ΤΗΣ ΤΑΞΗΣ
1. Συγκρίνετε τις εξαρτήσεις των βαθμών κορεσμού της αιμοσφαιρίνης (Hb) και της μυοσφαιρίνης (Mb) με το οξυγόνο από τη μερική της πίεση στους ιστούς
 Ρύζι. 1.34. Εξάρτηση κορεσμού Mv καιNHοξυγόνο από τη μερική του πίεση
Ρύζι. 1.34. Εξάρτηση κορεσμού Mv καιNHοξυγόνο από τη μερική του πίεση
Σημειώστε ότι το σχήμα των καμπυλών κορεσμού πρωτεϊνών οξυγόνου είναι διαφορετικό: για τη μυοσφαιρίνη - μια υπερβολή, για την αιμοσφαιρίνη - ένα σιγμοειδές σχήμα.
1. συγκρίνετε τις τιμές της μερικής πίεσης του οξυγόνου στην οποία τα Mb και Hb είναι κορεσμένα με O 2 κατά 50%. Ποια από αυτές τις πρωτεΐνες έχει μεγαλύτερη συγγένεια για το O 2;
2. Ποια δομικά χαρακτηριστικά του Mb καθορίζουν την υψηλή του συγγένεια για το O 2;
3. Ποια δομικά χαρακτηριστικά του HB του επιτρέπουν να απελευθερώνει O2 στα τριχοειδή αγγεία των ιστών ηρεμίας (σε σχετικά υψηλή μερική πίεση O2) και να αυξάνει απότομα αυτή την απελευθέρωση στους εργαζόμενους μύες; Ποια ιδιότητα των ολιγομερών πρωτεϊνών παρέχει αυτό το αποτέλεσμα;
4. Υπολογίστε ποια ποσότητα O 2 (σε%) οξυγονωμένης αιμοσφαιρίνης δίνει στους μύες που ξεκουράζονται και λειτουργούν;
5. εξάγετε συμπεράσματα για τη σχέση της δομής μιας πρωτεΐνης και της λειτουργίας της.
2. Η ποσότητα οξυγόνου που απελευθερώνεται από την αιμοσφαιρίνη στα τριχοειδή αγγεία εξαρτάται από την ένταση των καταβολικών διεργασιών στους ιστούς (φαινόμενο Bohr). Πώς οι αλλαγές στον μεταβολισμό των ιστών ρυθμίζουν τη συγγένεια της Hb για το O2; Επίδραση του CO 2 και του H+ στη συγγένεια της Hb για το O 2
 1. περιγράψτε το φαινόμενο Bohr.
1. περιγράψτε το φαινόμενο Bohr.
2. προς ποια κατεύθυνση προχωρά η διαδικασία που φαίνεται στο διάγραμμα:
α) στα τριχοειδή αγγεία των πνευμόνων.
β) στα τριχοειδή των ιστών;
3. Ποια είναι η φυσιολογική σημασία του φαινομένου Bohr;
4. Γιατί η αλληλεπίδραση της Hb με το Η+ σε θέσεις απομακρυσμένες από την αίμη αλλάζει τη συγγένεια της πρωτεΐνης για το O 2;
3. Η συγγένεια της Hb για το O2 εξαρτάται από τη συγκέντρωση του συνδέτη της - 2,3-διφωσφογλυκερικό, που είναι ένας αλλοστερικός ρυθμιστής της συγγένειας της Hb για το O2. Γιατί η αλληλεπίδραση συνδέτη σε μια θέση μακριά από την ενεργό θέση επηρεάζει τη λειτουργία των πρωτεϊνών; Πώς το 2,3-BPG ρυθμίζει τη συγγένεια της Hb για το O2; Για να λύσετε το πρόβλημα, απαντήστε στις ακόλουθες ερωτήσεις:
1. πού και από τι συντίθεται το 2,3-διφωσφογλυκερικό (2,3-BPG); Γράψτε τον τύπο του, υποδείξτε το φορτίο αυτού του μορίου.
2. Με ποια μορφή αιμοσφαιρίνης (οξυ ή δεοξυ) αλληλεπιδρά η BPG και γιατί; Σε ποιο μέρος του μορίου Hb συμβαίνει η αλληλεπίδραση;
3. προς ποια κατεύθυνση συμβαίνει η διαδικασία που φαίνεται στο διάγραμμα;
α) στα τριχοειδή αγγεία των ιστών.
β) στα τριχοειδή αγγεία των πνευμόνων;
4. όπου η συγκέντρωση του συμπλέγματος θα πρέπει να είναι μεγαλύτερη
Nv-2,3-BFG:
α) στα τριχοειδή αγγεία των μυών σε ηρεμία,
β) στα τριχοειδή αγγεία των μυών που λειτουργούν (παρέχεται η ίδια συγκέντρωση BPG στα ερυθροκύτταρα);
5. Πώς θα αλλάξει η συγγένεια της HB για το οξυγόνο όταν ένα άτομο προσαρμοστεί σε συνθήκες μεγάλου υψομέτρου, εάν αυξηθεί η συγκέντρωση της BPG στα ερυθροκύτταρα; Ποια είναι η φυσιολογική σημασία αυτού του φαινομένου;
4. Η καταστροφή του 2,3-BPG κατά την αποθήκευση του συντηρημένου αίματος βλάπτει τις λειτουργίες του HB. Πώς θα αλλάξει η συγγένεια του HB για το O 2 στο διατηρημένο αίμα εάν η συγκέντρωση του 2,3-BPG στα ερυθροκύτταρα μπορεί να μειωθεί από 8 σε 0,5 mmol/l. Είναι δυνατή η μετάγγιση τέτοιου αίματος σε βαριά άρρωστους ασθενείς εάν η συγκέντρωση του 2,3-BPG αποκατασταθεί όχι νωρίτερα από τρεις ημέρες; Είναι δυνατόν να αποκατασταθούν οι λειτουργίες των ερυθρών αιμοσφαιρίων προσθέτοντας 2,3-BPG στο αίμα;
5. Θυμηθείτε τη δομή των απλούστερων μορίων ανοσοσφαιρίνης. Τι ρόλο παίζουν οι ανοσοσφαιρίνες στο ανοσοποιητικό σύστημα; Γιατί τα Igs ονομάζονται συχνά δισθενή; Πώς σχετίζεται η δομή των Igs με τη λειτουργία τους; (Περιγράψτε χρησιμοποιώντας ένα παράδειγμα μιας κατηγορίας ανοσοσφαιρινών.)
Φυσικοχημικές ιδιότητες πρωτεϊνών και μέθοδοι διαχωρισμού τους.
6. Πώς το καθαρό φορτίο μιας πρωτεΐνης επηρεάζει τη διαλυτότητά της;
α) προσδιορίστε το συνολικό φορτίο του πεπτιδίου σε pH 7
Ala-Glu-Tre-Pro-Asp-Liz-Cis
β) πώς θα αλλάξει το φορτίο αυτού του πεπτιδίου σε pH >7, pH<7, рН <<7?
γ) ποιο είναι το ισοηλεκτρικό σημείο μιας πρωτεΐνης (IEP) και σε ποιο περιβάλλον βρίσκεται;
IET αυτού του πεπτιδίου;
δ) σε ποια τιμή pH θα παρατηρηθεί η μικρότερη διαλυτότητα αυτού του πεπτιδίου.
7. Γιατί το ξινόγαλα, σε αντίθεση με το φρέσκο γάλα, «πήζει» όταν βράζεται (δηλαδή, η καζεΐνη πρωτεΐνης γάλακτος κατακρημνίζεται); Στο φρέσκο γάλα, τα μόρια της καζεΐνης έχουν αρνητικό φορτίο.
8. Η διήθηση γέλης χρησιμοποιείται για τον διαχωρισμό μεμονωμένων πρωτεϊνών. Ένα μείγμα που περιέχει πρωτεΐνες Α, Β, Γ με μοριακά βάρη ίσα με 160.000, 80.000 και 60.000, αντίστοιχα, αναλύθηκε με διήθηση γέλης (Εικ. 1.35). Οι διογκωμένοι κόκκοι γέλης είναι διαπερατοί σε πρωτεΐνες με μοριακό βάρος μικρότερο από 70.000. Ποια αρχή βασίζεται σε αυτή τη μέθοδο διαχωρισμού; Ποιο γράφημα αντικατοπτρίζει σωστά τα αποτελέσματα της κλασμάτωσης; Υποδείξτε τη σειρά με την οποία οι πρωτεΐνες Α, Β και Γ απελευθερώνονται από τη στήλη.
 Ρύζι. 1,35. Χρήση Διήθησης Γέλης για Διαχωρισμό Πρωτεϊνών
Ρύζι. 1,35. Χρήση Διήθησης Γέλης για Διαχωρισμό Πρωτεϊνών
9. Στο Σχ. 1.36, Α δείχνει ένα διάγραμμα ηλεκτροφόρησης σε χαρτί πρωτεϊνών ορού αίματος από ένα υγιές άτομο. Οι σχετικές ποσότητες πρωτεϊνικών κλασμάτων που λαμβάνονται χρησιμοποιώντας αυτή τη μέθοδο είναι: λευκωματίνες 54-58%, α1-σφαιρίνες 6-7%, α2-σφαιρίνες 8-9%, β-σφαιρίνες 13%, γ-σφαιρίνες 11-12%.
 Ρύζι. 1.36 Ηλεκτροφόρηση σε χαρτί πρωτεϊνών πλάσματος αίματος ενός υγιούς ατόμου (Α) και ενός ασθενούς (Β)
Ρύζι. 1.36 Ηλεκτροφόρηση σε χαρτί πρωτεϊνών πλάσματος αίματος ενός υγιούς ατόμου (Α) και ενός ασθενούς (Β)
Ι - γ-σφαιρίνες; II - β-σφαιρίνες; III -α 2 -σφαιρίνη; IV -α 2 -σφαιρίνη; V - λευκωματίνες
Πολλές ασθένειες συνοδεύονται από ποσοτικές αλλαγές στη σύνθεση των πρωτεϊνών του ορού (δυσπρωτεϊναιμία). Η φύση αυτών των αλλαγών λαμβάνεται υπόψη κατά τη διάγνωση και την αξιολόγηση της σοβαρότητας και του σταδίου της νόσου.
Χρησιμοποιώντας τα δεδομένα που δίνονται στον πίνακα. 1.5, κάντε μια εικασία για την ασθένεια, η οποία χαρακτηρίζεται από το ηλεκτροφορητικό προφίλ που παρουσιάζεται στο Σχ. 1.36.
Πίνακας 1.5. Αλλαγές στη συγκέντρωση πρωτεϊνών ορού στην παθολογία







