Все водоросли хорошо различаются по набору фотосинтезирующих пигментов. Такие группы в систематике растений имеют статус отделов.
Основным пигментом всех водорослей является зеленый пигмент хлорофилл. Известно четыре типа хлорофилла, которые различаются по своей структуре: хлорофилл a – присутствует у всех водорослей и высших растений; хлорофилл b – встречается у зеленых, харовых, евгленовых водорослей и у высших растений: растения, содержащие этот хлорофилл, всегда имеют ярко-зеленую окраску; хлорофилл c – встречается у гетероконтных водорослей; хлорофилл d – редкая форма, встречается у красных и синезеленых водорослей. Большинство фотосинтезирующих растений содержат два разных хлорофилла, одним из которых всегда является хлорофилл a. В некоторых случаях вместо второго хлорофилла присутствуют билипротеины . У синезеленых и красных водорослей встречаются два вида билипротеинов: фикоцианин – синий пигмент, фикоэритрин – красный пигмент.
Обязательными пигментами, входящими в фотосинтетические мембраны, являются желтые пигменты – каротиноиды . Они отличаются от хлорофиллов спектром поглощаемого света и, как полагают, выполняют защитную функцию, предохраняя молекулы хлорофилла от разрушительного воздействия молекулярного кислорода.
Кроме перечисленных пигментов у водорослей встречаются: фукоксантин – золотистый пигмент; ксантофилл – бурый пигмент.
Конец работы -
Эта тема принадлежит разделу:
Водоросли
Рыбохозяйственный университет.. институт биологии моря им а в жирмунского дво ран.. л л арбузова..
Если Вам нужно дополнительный материал на эту тему, или Вы не нашли то, что искали, рекомендуем воспользоваться поиском по нашей базе работ:
Что будем делать с полученным материалом:
Если этот материал оказался полезным ля Вас, Вы можете сохранить его на свою страничку в социальных сетях:
| Твитнуть |
Все темы данного раздела:
Клеточные покровы
Клеточные покровы обеспечивают устойчивость внутреннего содержимого клеток к внешним воздействиям и придают клеткам определенную форму. Покровы проницаемы для воды и растворенных в ней низкомолекул
Жгутики
Монадные вегетативные клетки и монадные стадии в жизненном цикле (зооспоры и гаметы) водорослей снабжены жгутиками – длинными и довольно толстыми выростами клеток, снаружи покрытыми плазмалеммой. И
Митохондрии
Митохондрии встречаются в клетках эукариотных водорослей. Форма и строение митохондрий в клетках водорослей более разнообразны по сравнению с митохондриями высших растений. Они могут быть округлыми
Пластиды
Пигменты в клетках эукариотных водорослей находятся в пластидах, как и у всех растений. У водорослей встречается два типа пластид: окрашенные хлоропласты (хроматофоры) и бесцветные лейкопласты (ами
Ядро и митотический аппарат
Ядро водорослей имеет типичную для эукариотов структуру. Количество ядер в клетке может варьировать от одного до нескольких. Снаружи ядро покрыто оболочкой, состоящей из двух мембран, наружная мемб
Монадный (жгутиковый) тип структуры таллома
Наиболее характерным признаком, определяющим этот тип структуры, является наличие жгутиков, с помощью которых монадные организмы активно передвигаются в водной среде (рис. 9, А). Подвижные ж
Ризоподиальный (амебоидный) тип структуры
Наибоее существенные признаки амебоидого типа структуры – отсутствие прочных клеточных покровов и способность к амебоидному движению, с помощью временно образующихся на поверхности клетки ци
Пальмеллоидный (гемимонадный) тип структуры
Характерным для этого типа структуры является сочетание неподвижного растительного образа жизни с наличием клеточных органелл, свойственных монадным организмам: сократительные вакуоли, стигма, жгут
Коккоидный тип структуры
Этот тип объединяет одноклеточные и колониальные водоросли, неподвижные в вегетативном состоянии. Клетки коккоидного типа одеты оболочкой и имеют протопласт растительного типа (тонопласт без сократ
Трихальный (нитчатый) тип структуры
Характерной особенностью нитчатого типа структуры является нитевидное расположение неподвижных клеток, которые образуются вегетативным путем в результате клеточного деления, происходящего преимущес
Гетеротрихальный (разнонитчатый) тип структуры
Разнонитчатый тип возник на базе нитчатого типа. Разнонитчатое слоевище состоит большей частью из горизонтальных, стелящихся по субстрату нитей, выполняющих функцию прикрепления, и вертикальных, по
Паренхиматозный (тканевой) тип структуры
Одно из направлений эволюции разнонитчатого слоевища было связано с возникновением паренхиматозных слоевищ. Способность к неограниченному росту и делению клеток в разных направлениях привела к обра
Сифональный тип структуры
Сифональный (неклеточный) тип структуры характеризуется отсутствием внутри слоевища, достигающего сравнительно крупных, обычно макроскопических размеров и определенной степени дифференцировки, клет
Сифонокладальный тип структуры
Основным признаком сифонокладального типа структуры является способность к образованию из первичного неклеточного слоевища сложно устроенных слоевищ, состоящих из первично многоядерных сегментов. В
Бесполое размножение
Бесполое размножение водорослей осуществляется с помощью специализированных клеток – спор. Спорообразование обычно сопровождается делением протопласта на части и выходом продуктов деления из
Простое деление
Этот способ размножения встречается только у одноклеточных форм водорослей. Наиболее просто происходит деление у клеток, имеющих амебоидный тип строения тела.
Деление амебоидных форм
Фрагментация
Фрагментация присуща всем группам многоклеточных водорослей и проявляется в разных формах: образование гормогониев, регенерация оторвавшихся частей слоевища, спонтанное отпадание ветвей, отрастание
Размножение побегами, столонами, выводковыми почками, клубеньками, акинетами
У тканевых форм зеленых, бурых и красных водорослей вегетативное размножение приобретает свою законченную форму, которая мало отличается от вегетативного размножения высших растений. Сохраняя спосо
Половое размножение
Половое размножение у водорослей связано с половым процессом, который заключается в слиянии двух клеток, в результате чего образуется зигота, вырастающая в новую особь или дающая зооспоры.
Смена ядерных фаз
При половом процессе в результате слияния гамет и их ядер происходит удвоение числа хромосом в ядре. На определенном этапе цикла развития, при мейозе, происходит редукция числа хромосом, в результа
Эндофиты/ эндозоиты, или эндосимбионты
Эндосимбионты, или внутриклеточные симбионты – водоросли, живущие в тканях или клетках других организмов (беспозвоночных животных или водорослей). Они образуют своеобразную экологическую гру
Отдел синезеленые водоросли (цианобактерии) – cyanophyta
В названии отдела (от греч. cyanos – синий) отражена характерная особенность этих водорослей – окраска таллома, связанная с относительно высоким содержанием синего пигмента фикоцианина. Циан
Порядок– Chroococcales
Встречаются в виде одноклеточных "простых" индивидов или чаще образуют слизистые колонии. При делении клеток в двух плоскостях возникают однослойные пластинчатые колонии. Деление в трех п
Отдел красные водоросли– rhodophyta
Название отдела происходит от греческого слова rhodon ("родон") –розовый. Окраска красных водорослей обусловлена различным сочетанием пигментов. Она бывает от серого и фиолетового
Порядок Бангиевые–Bangiales
Род Порфира имеет слоевище в виде тонкой блестящей пластины с ровными или складчатыми краями, состоящей из одного-двух слоев плотно соединенных клеток. Основание пластины обычно переходит в
Порядок Родимениевые – Rhodymeniales
Род Спарлингия (Родимения) – плоские пластины до 45 см высоты, листовидной и клиновидной формы, расширенные и пальчато рассеченные вверху, от светло-розового или светло-оранжевого до
Порядок Кораллиновые – Corallinales
Род Кораллина – членистые вееровидно разветвленные кустики до 10 см высоты, разветвленные, известковые, от розово-сиреневого до почти белого цвета. Размножается бесполым и половым путем. Спо
Порядок Гигартиновые – Gigartinales
Род Хондрус – плотные кожистые хрящеватые кустики до 20 см высоты, 3–4-кратно разветвленные, светло-желтого, светло-розового, пурпурно-темно-красного цвета. Растет в нижней части литорали и
Порядок Церамиевые – Ceramiales
Род Церамиум – нежные пушистые членистые кустики до 10 см высоты, дихотомически или поочередно разветвленные, темно-желтого с розоватым оттенком цвета. Ветвление двух-четырех порядков, конеч
Отдел диатомовые водоросли – bacillariophyta
Отдел называется Диатомовые водоросли (от греч. di - два, tome - разрез, рассечение), или Бациллярии (bacillum – палочка). Включает одноклеточные одиночные или колониальные орг
Отдел гетероконтные (разножгутиковые) водоросли – heterokontophyta
У всех гетероконтов похоже устроен жгутиковый аппарат. Имеется 2 жгутика, причем на одном из них есть очень характерные трубчатые трехчленные перистые выросты, или волоски – мастигонемы. Именно нал
Систематика
Ископаемые кокколиты известны из мезозойских отложений и были обильны на протяжении больней части юрского и в меловом периодах. Максимального разнообразия примнезиофитовые достигли в позднем мелу,
Отдел криптофитовые водоросли (криптомонады) – cryptophyta
Отдел назван по типовому роду Криптомонас (от греч. kryptos – скрытый, monas - особь). Включает одноклеточные, подвижные, монадные организмы. Клетки криптофитов
А б в г д
Рис. 53. Внешний вид криптофитовых водорослей (по: Г.А. Белякова и др., 2006): А – Родомонас, Б – Хроомонас, В – Криптомонас, Г – Хиломонас, Д – Гониомонас
может фор
Отдел зеленые водоросли – chlorophyta
Зеленые водоросли - самый обширный из всех отделов водорослей, насчитывающий по разным оценкам от 4 до 13 - 20 тысяч видов. Все они имеют зеленый цвет слоевищ, что обусловлено преобладанием в хлоро
Порядок Улотриксовые – Ulotrichales
Род Улотрикс (рис. 54). Виды Улотрикса обитают чаще в пресных, реже в морских, солоноватых водоемах и в почве. Они прикрепляются к подводным предметам, формируя ярко-зеленые кустики р
Порядок Бриопсидовые– Bryopsidales
Большинство видов встречается в пресных и солоноватых водоемах. Некоторые из них растут на почве, на камнях, песке и иногда на солончаках.
Род Бриопсис – нитевидные кустики до 6-8 с
Порядок Вольвоксовые - Volvocales
Род Хламидомонада (рис. 57)включает свыше 500 видов одноклеточных водорослей, которые обитают в пресных, мелких, хорошо прогреваемых и загрязненных водоемах: прудах, лужах, канавах и т.п. Пр
Отдел Харофитовые (Харовые) – Charophyta
Харофитовые – линия пресноводных зеленых водорослей, приведшая к высшим растениям. Это формы преимущественно с нитчатым талломом. Нередко таллом вертикальный, расчлененный и несет о
Отдел динофитовые (динофлагеллаты) – dinophyta
1. Название отдела происходит от греч. dineo - вращаться. Объединяет преимущественно одноклеточные монадные, реже коккоидные, амебоидные или пальмеллоидные, иногда колониальн
Отдел euglenozoa – евгленовые
Отдел назва по типовому роду - Euglena (от греч. еu – хорошо развитый, glene – зрачок, глаз). Объединяет одиночных монадных или амебоидных представителей. Изредка встречаются к
Словарь терминов
Автогамия – половое размножение, при котором сливаются два сестринских гаплоидных ядра в общей цитоплазме.
Автоспора – структура бесполого размножения, представляющая собой
Дальневосточный государственный технический
рыбохозяйственный университет
Институт биологии моря им. А.В. Жирмунского ДВО РАН
Л.Л. Арбузова
И.Р. Левенец
Водоросли

Рецензенты:
– В.Г.Чавтур, д.б.н., профессор кафедры Морской биологии и аквакультуры Дальневосточного государственного университета
– С.В.Нестерова, к.б.н., старший научный сотрудник лаборатории флоры Дальнего Востока Ботанического сада-института ДВО РАН
Арбузова Л.Л., Левенец И.Р. Водоросли: Уч. пос. Владивосток: Дальрыбвтуз, ИБМ ДВО РАН, 2010. 177с.
В пособии приведены современные сведения об анатомии, морфологии, систематике, образе жизни и практическом значении водорослей.
Учебное пособие предназначено для студентов-бакалавров по направлениям «Водные биоресурсы и аквакультура» и «Экология и природопользование» дневной и заочной формы обучения, магистров экологии, биологии, ихтиологии и рыбоводства.
©Дальневосточный государственный
технический рыбохозяйственный
университет, 2010
©Институт биологии моря им. А.В. Жирмунского ДВО РАН, 2010
ISBN ……………………..
Введение ………………………………………………………………
1. Строение клеток водорослей ……………………………………
2. Общая характеристика водорослей ……………………………
2.1. Типы питания ……………………………………………………
2.2. Типы талломов …………………………………………………..
2.3. Размножение водорослей ……………………………………..
2.4. Жизненные циклы водорослей ……………………………….
3. Экологические группы водорослей …………………………….
3.1. Водоросли водных местообитаний …………………………..
3.1.1. Фитопланктон ………………………………………………….
3.1.2. Фитобентос ……………………………………………………..
3.1.3. Водоросли экстремальных водных экосистем ……………
3.2. Водоросли вневодных местообитаний ………………………
3.2.1. Аэрофильные водоросли …………………………………….
3.2.2. Эдафофильные водоросли ………………………………….
3.2.3. Литофильные водоросли ……………………………………..
4. Роль водорослей в природе и практическое значение ………
5. Современная систематика водорослей ………………………..
5.1. Прокариотические водоросли ………………………………..
5.1.1. Отдел Синезеленые водоросли ………………………
5.2. Эукариотические водоросли ………………………………….
5.2.1. Отдел Красные водоросли …………………………….
5.2.2. Отдел Диатомовые водоросли ………………………..
5.2.3. Отдел Гетероконтные водоросли …………………….
Класс Бурые водоросли …………………………………
Класс Золотистые водоросли …………………............
Класс Синуровые водоросли …………………………..
Класс Феотамниевые водоросли ………………………
Класс Рафидовые водоросли ………………………….
Класс Евстигмовые водоросли …………………………
Класс Желтозеленые водоросли ………………………
5.2.4. Отдел Примнезиофитовые водоросли ……………….
5.2.5. Отдел Криптофитовые водоросли ……………………
5.2.6. Отдел Зеленые водоросли …………………….………
5.2.7. Отдел Харовые водоросли …………………………….
5.2.8. Отдел Динофитовые водоросли ………………………
5.2.9. Отдел Евгленовые водоросли …………………………
Литература ……………………………………………………………
Словарь терминов …………………………………………………….
Приложение …………………………………………………………….
ВВЕДЕНИЕ
Водоросли традиционно объединяют разнородную группу талломных, фотосинтезирующих, споровых бессосудистых организмов. Как у всех низших растений, органы размножения водорослей лишены покровов, тело не разделено на органы, отсутствуют ткани. Среди водорослей встречаются как эукариотические, так и прокариотические формы. Последние, в отличие от хлоробактерий, в процессе фотосинтеза выделяют в среду свободный кислород.
Водоросли занимают господствующее положение как в пресных, так и в морских водоемах. Являясь основными продуцентами, они в значительной мере определяют рыбопродуктивность водных экосистем. Благодаря фотосинтезирующей деятельности водоросли обогащают воду кислородом и уменьшают количество углекислоты. Они обладают уникальной способностью аккумулировать из окружающей водной среды различные вредные вещества, а также выделять в среду метаболиты, которые подавляют рост патогенных микроорганизмов. Водоросли, изменяя химический состав воды, нередко способствуют ее очищению. Качественный и количественный состав групп водорослей является важным показателем экологического состояния водоемов. Ряд видов используют в качестве индикаторов загрязнения водной среды.
Изучение водорослей является важным этапом при подготовке специалистов в области марикультуры, рыбоводства и морской экологии. Знания по строению, экологии и систематике водорослей являются базовыми для изучения гибробиологии, ихтиологии, экологии, ихтиотоксикологии; они также необходимы для оценки сырьевой базы водоемов и составления промысловых прогнозов.
В последнее время благодаря современным методическим приемам получены новые сведения о тонком строении, физиологии и биохимии водорослей, что вызвало пересмотр традиционных представлений. Наибольшие изменения претерпела систематика низших растений, к которым относятся водоросли. В то же время в учебной литературе по ботанике современные сведения о систематике и строении водорослей не отражаются, а специальная литература по фикологии не доступна для широкой студенческой аудитории.
В данном учебном пособии приведены новейшие сведения о строении, морфологии, таксономии, экологии и практическом значении водорослей. Дано описание наиболее значимых таксонов водорослей.
Учебное пособие предназначено для студентов-бакалавров по направлениям «Водные биоресурсы и аквакультура» и «Экология и природопользование» дневной и заочной форм обучения, магистров в области экологии, ихтиологии, рыбоводства и аквакультуры.
В подготовке материалов для данного пособия принимали участие преподаватели Дальрыбвтуза и специалисты в области гидробиологии и фикологии ДВО РАН.
1. СТРОЕНИЕ КЛЕТОК ВОДОРОСЛЕЙ
Прокариотические водоросли по строению клетки сходны с бактериями: у них отсутствуют мембранные органеллы, такие, как ядро, хлоропласты, митохондрии, эндоплазматическая сеть, аппарат Гольджи.
Эукариотические водоросли содержат структурные элементы, характерные для клеток высших растений (рис. 1).

Рис. 1. Взрослая растительная клетка без вторичного утолщения стенки (схематизировано) при максимальном увеличении светового микроскопа (по:): 1 – стенка клетки, 2 – срединная пластинка, 3 – межклетник, 4 – плазмодесмы, 5 – плазмалемма, 6 – тонопласт, 7 – центральная вакуоль, 9 – ядро, 10 – ядерная оболочка, 11 – пора в ядерной оболочке, 12 – ядрышко, 13 – хроматин, 14 – хлоропласт, 15 – грана в хлоропласте, 16 – крахмальное зерно в хлоропласте, 17 – митохондрия, 18 – диктиосома, 19 – гранулярный эндоплазматический ретикулум, 20 – капелька запасного жира (липида) в цитоплазме, 21 – микротело, 22 – цитоплазма (гиалоплазма)
Приведенная схема растительной клетки в целом отражает строение клеток водорослей, однако многие водоросли, наряду с типичными растительными органеллами (пластиды, вакуоль с клеточным соком), содержат структуры, характерные для животных клеток (жгутики, стигму, нетипичные для растительных клеток оболочки).
Клеточные покровы
Клеточные покровы обеспечивают устойчивость внутреннего содержимого клеток к внешним воздействиям и придают клеткам определенную форму. Покровы проницаемы для воды и растворенных в ней низкомолекулярных веществ и легк пропускают солнечный свет. Клеточные покровы водорослей отличаются большим морфологическим и химическим разнообразием. В их состав входят полисахариды, белки, гликопротеиды, минеральные соли, пигменты, липиды, вода. В отличие от высших растений, в оболочках водорослей отсутствует лигнин.
В основе строения клеточных покровов лежит плазмалемма, или цитоплазматическая мембрана. У многих жгутиковых и амебоидных представителей клетки снаружи покрыты лишь плазмалеммой, которая не способна обеспечить постоянную форму тела. Такие клетки могут образовывать псевдоподии. По морфологии выделяют несколько типов псевдоподий. Чаще всего у водорослей встречаются ризоподии , представляющие собой нитевидные длинные, тонкие, разветвленные, иногда анастомозирующие цитоплазматические выросты. Внутри ризоподий находятся микрофиламенты. Лобоподии – широкие закругленные выпячивания цитоплазмы. Они встречаются у водорослей с амебоидным и с монадным типами дифференциации таллома. Реже у водорослей встречаются филоподии – тонкие подвижные образования, напоминающие щупальца, которые способны втягиваться в клетку.
У многих динофлагеллат тело покрыто чешуйками, расположенными на поверхности клетки. Чешуйки могут быть одиночными или смыкаться в сплошной покров – теку . Они могут быть органическими или неорганическими. Органические чешуйки встречаются на поверхности зеленых, золотистых, криптофитовых водорослей. В состав неорганических чешуек может входить или карбонат кальция, или кремнезем. Чешуйки из карбоната кальция – кокколиты – встречаются преимущественно у морских примнезиофитовых водорослей.
Нередко клетки жгутиковых и амебоидных водорослей располагаются в домиках, которые имеют в основном органическое происхождение. Их стенки могут быть тонкими и прозрачными (род Динобрион ) или более прочными и окрашенными за счет отложения в них солей железа и марганца (род Трахеломонас ). В домиках имеется обычно одно отверстие для выхода жгутика, иногда может быть несколько отверстий. При размножении водорослей домик не разрушается, чаще всего одна из образовавшихся клеток покидает его и строит новый домик.
Клеточный покров евгленовых водорослей называется пелликулой. Пелликула являет собой совокупность цитоплазматической мембраны и расположенных под ней белковых полос, микротрубочек и цистерн эндоплазматической сети.
У динофитовых водорослей клеточные покровы представлены амфиесмой. Амфиесма состоит из плазмалеммы и расположенной под ней совокупности уплощенных везикул, под которыми лежит слой микротрубочек. В везикулах ряда динофитовых могут располагаться целлюлозные пластинки; такая амфиесма называется текой , или панцирем (роды Цератиум, Перидиниум ).
У диатомовых водорослей поверх плазмалеммы формируется особый клеточный покров – панцирь , состоящий главным образом из аморфного кремнезема. Помимо кремнезема в состав панциря входит примесь органических соединений и некоторых металлов (железо, алюминий, магний).
В клеточных стенках зеленых, желтозеленых, красных и бурых водорослей основным структурным компонентом служит целлюлоза, которая образует каркас (структурную основу), погруженный в матрикс (полужидкую среду), состоящий из пектина, гемицеллюлозы, альгиновой кислоты и других органических веществ.
Жгутики
Монадные вегетативные клетки и монадные стадии в жизненном цикле (зооспоры и гаметы) водорослей снабжены жгутиками – длинными и довольно толстыми выростами клеток, снаружи покрытыми плазмалеммой. Их количество, длина, морфология, место прикрепления, характер движения достаточно разнообразны у водорослей, но постоянны внутри родственных групп.
Жгутики могут прикрепляться на переднем конце клетки (апикально) или могут быть чуть сдвинуты вбок (субапикально); возможно их прикрепление сбоку клетки (латерально) и на брюшной стороне клетки (вентрально). Жгутики, одинаковые по морфологии, называют изоморфными , если они различаются – гетероморфными . Изоконтные – это жгутики одинаковой длины, гетероконтные – разной длины.
Жгутики имеют единый план строения. Можно выделить свободную часть (ундулиподию), переходную зону, базальное тело (кинетосому). Разные части жгутика отличаются по количеству и расположению микротрубочек, которые образуют скелет (рис. 2).

Рис. 2. Схема строения жгутиков водорослей (по: Л.Л.Великанов и др., 1981): 1 – продольный разрез жгутиков; 2, 3 – поперечный разрез через кончик жгутика; 4 – поперечный разрез через ундулиподию; 5 – переходная зона; 6 – поперечный разрез через основание жгутика – кинетосому
Ундулиподия (в переводе с латинского «волноножка») способна совершать ритмичные волнообразные движения. Ундулиподия представляет собой одетую мембраной аксонему. Аксонема состоит из девяти пар микротрубочек, расположенных по кругу, и пары микротрубочек в центре (рис. 2). Жгутики могут быть гладкими или покрыты чешуйками либо мастигонемами (волосками), а у динофитовых и криптофитовых они покрыты и чешуйками и волосками. Жгутики примнезиофитовых, криптофитовых и зеленых водорослей могут быть покрыты чешуйками разной формы и размеров.
Переходная зона . Функционально играет роль в укреплении жгутика в месте его выхода из клетки. У водорослей выделяют несколько типов структур переходной зоны: поперечная пластинка (динофитовые), звездчатая структура (зеленые), переходная спираль (гетероконтные), переходный цилиндр (примнезиофитовые и динофитовые).
Базальное тело или кинетосома . У этой части жгутика структура в виде полого цилиндра, стенка которого образована девятью триплетами микротрубочек. Функция кинетосомы – связь жгутика с плазмалеммой клетки. Базальные тела у ряда водорослей могут принимать участие в делении ядра и становиться центрами организации микротрубочек.
Митохондрии
Митохондрии встречаются в клетках эукариотных водорослей. Форма и строение митохондрий в клетках водорослей более разнообразны по сравнению с митохондриями высших растений. Они могут быть округлыми, нитевидными, в виде сети или неправильных очертаний. Их форма может варьировать в одной и той же клетке на разных стадиях жизненного цикла. Митохондрии покрыты оболочкой из двух мембран. Матрикс митоходрий содержит рибосомы и митохондриальную ДНК. Внутренняя мембрана образует складки – кристы (рис. 3).

Рис. 3. Строение митохондрий растений (по:): А–объемное изображение; Б–продольный срез; В – часть кристы с грибовидными выступами: 1– наружная мембрана, 2– внутренняя мембрана, 3–криста, 4–матрикс, 5–межмембранное пространство, 6–митохондриальные рибосомы, 7–гранула, 8– митохондриальная ДНК, 9–АТФ-сомы
Кристы водорослей бывают разнообразной формы: дисковидные (евгленовые водоросли), трубчатые (динофитовые водоросли), пластинчатые (зеленые, красные, криптомонадовые водоросли) (рис. 4).

Рис. 4. Различные типы крист митохондрий (по:): А – пластинчатые; Б – трубчатые; В – дисковидные; к - кристы
Самыми примитивными считаются дисковидные кристы.
Пигменты
Все водоросли хорошо различаются по набору фотосинтезирующих пигментов. Такие группы в систематике растений имеют статус отделов.
Основным пигментом всех водорослей является зеленый пигмент хлорофилл. Известно четыре типа хлорофилла, которые различаются по своей структуре: хлорофилл a – присутствует у всех водорослей и высших растений; хлорофилл b – встречается у зеленых, харовых, евгленовых водорослей и у высших растений: растения, содержащие этот хлорофилл, всегда имеют ярко-зеленую окраску; хлорофилл c – встречается у гетероконтных водорослей; хлорофилл d – редкая форма, встречается у красных и синезеленых водорослей. Большинство фотосинтезирующих растений содержат два разных хлорофилла, одним из которых всегда является хлорофилл a . В некоторых случаях вместо второго хлорофилла присутствуют билипротеины . У синезеленых и красных водорослей встречаются два вида билипротеинов: фикоцианин – синий пигмент, фикоэритрин – красный пигмент.
Обязательными пигментами, входящими в фотосинтетические мембраны, являются желтые пигменты – каротиноиды . Они отличаются от хлорофиллов спектром поглощаемого света и, как полагают, выполняют защитную функцию, предохраняя молекулы хлорофилла от разрушительного воздействия молекулярного кислорода.
Кроме перечисленных пигментов у водорослей встречаются: фукоксантин – золотистый пигмент; ксантофилл – бурый пигмент.
Пластиды
Пигменты в клетках эукариотных водорослей находятся в пластидах, как и у всех растений. У водорослей встречается два типа пластид: окрашенные хлоропласты (хроматофоры) и бесцветные лейкопласты (амилопласты). Хлоропласты водорослей в отличие от таковых высших растений значительно разнообразнее по форме и по строению (рис. 5).

Рис. 5. Схема строения хлоропластов у эукариотных водорослей (по:):1 – рибосомы; 2 – оболочка хлоропласта; 3 – опоясывающий тилакоид; 4 – ДНК; 5 – фикобилисомы; 6 – крахмал; 7 – две мембраны хлоропластной ЭПС; 8 – две мембраны оболочки хлоропласта; 9 – ламелла; 10 – запасной продукт; 11 – ядро; 12 – одна мембрана хлоропластной ЭПС; 13 – липид; 14 – грана; 15 – пиреноид. А – тилакоиды расположены по одному, отсутствует ХЭС - хлоропластная эндоплазматическая сеть (Rhodophyta); Б – ламеллы двухтилакоидные, две мембраны ХЭС (Cryptophyta); В – трехтилакоидные ламеллы, одна мембрана ХЭС (Dinophyta. Euglenophyta); Г – трехтилакоидные ламеллы, две мембраны ХЭС (Heterokontophyta, Prymnesiophyta); Д – двух-, шеститилакоидные ламеллы, отсутствует ХЭС (Chlorophyta)
Структурной фотосинтезирующей единицей эукариот и прокариот является тилакоид – плоский мембранный мешочек. В тилакоидные мембраны встроены пигментные системы и переносчики электронов. С тилакоидами связана световая фаза фотосинтеза. Темновая фаза фотосинтеза проходит в строме хлоропласта. Оболочка у зеленых и красных водорослей состоит из двух мембран. У других водорослей хлоропласт окружен дополнительными одной или двумя мембранами хлоропластной эндоплазматической сети (ХЭС ). У евгленовых и большинства динофитовых хлоропласт окружен тремя мембранами, а у гетероконтных и криптофитовых – четырьмя (рис. 5).
Ядро и митотический аппарат
Ядро водорослей имеет типичную для эукариотов структуру. Количество ядер в клетке может варьировать от одного до нескольких. Снаружи ядро покрыто оболочкой, состоящей из двух мембран, наружная мембрана покрыта рибосомами. Пространство между ядерными мембранами называется перинуклеарным . В нем могут находиться хлоропласты или лейкопласты, как у гетероконтных и криптофитовых. В матриксе ядра находится хроматин, представляющий ДНК в комплексе с основными белками – гистонами. Исключением являются динофитовые, у которых количество гистонов невелико и отсутствует нуклеосомная организация хроматина. Хроматиновые нити у этих водорослей уложены в виде восьмерок. В ядре бывает от одного до нескольких ядрышек, исчезающих или сохраняющихся во время митоза.
Митоз - непрямое деление водорослей, может протекать по-разному, но в целом схема этого процесса с 4 стадиями сохраняется (рис. 6).

Рис. 6. Последовательные фазы митоза: 1 – интерфаза; 2-4– профаза; 5– метафаза; 6– анафаза; 7-9–телофаза; 10– цитокинез
Профаза – самая длительная фаза митоза. В ней происходят важнейшие преобразования: ядро увеличивается в объеме, вместо слабозаметной хроматиновой сети в нем появляются хромосомы в виде тонких длинных изогнутых и слабо спирализированных нитей, образующих подобие клубка. С самого начала профазы видно, что хромосомы состоят из 2 нитей (результат их репликации в интерфазе). Половинки хромосом (хроматиды) располагаются параллельно друг другу. По мере развития профазы нити все более спирализуются, и образующиеся хромосомы все более укорачиваются и уплотняются.
В конце профазы выявляются индивидуальные морфологические признаки хромосом. Затем исчезают ядрышки, ядерная оболочка фрагментируется на отдельные короткие цистерны, неотличимые от элементов ЭПС, в результате чего нуклеоплазма смешивается с гиалоплазмой и образуется миксоплазма; из вещества ядра и цитоплазмы формируются ахроматиновые нити – веретено деления.
Веретено деления двухполюсное и состоит из пучков микротрубочек, тянущихся от одного полюса к другому. После разрушения ядерной оболочки каждая хромосома прикрепляется к нитям веретена деления при помощи своей центромеры. После прикрепления хромосом к веретену они выстраиваются в экваториальной плоскости клетки так, что все центромеры находятся на одинаковом расстоянии от его полюсов.
Метафаза . В этой фазе митоза хромосомы достигают максимального уплотнения и приобретают характерную форму, свойственную каждому виду растения. Обычно они бывают двуплечими, и в этих случаях в месте перегиба, именуемого центромерой, хромосомы соединяются с ахроматиновой нитью веретена. В метафазе отчетливо видно, что каждая хромосома состоит из двух дочерних хроматид. Расположены они более или менее параллельно в экваториальной плоскости клетки. К концу стадии каждая хромосома разделяется на две хроматиды, остающиеся соединенными только в центромерном участке. Позднее расщепляются и центромеры на две сестринские; при этом сестринские центромеры и хроматиды обращены к противоположным полюсам.
Анафаза . Самая короткая фаза митоза. Дочерние хромосомы – хроматиды – расходятся к противоположным полюсам клетки. Теперь свободные концы хроматид направлены к экватору, а кинетохоры – к полюсам. Полагают, что хроматиды расходятся в силу сокращения ахроматиновых нитей веретена, которые смыкаются с центромерой. Хромосомы вследствие раскручивания и удлинения становятся менее заметными. В центре клетки (по экватору) иногда уже на этой стадии появляются фрагменты клеточной стенки – фрагмопласт.
Телофаза . Продолжается процесс раскручивания - деспирализация и удлинение хромосом. Наконец, они теряются в поле зрения оптического микроскопа. Восстанавливаются оболочка ядра и ядрышко. Идет тот же процесс, что и в профазе, только в обратном порядке. Хромосомы теперь имеют по одной хроматиде. Структура интерфазного ядра восстанавливается, веретено изменяет бочковидную форму на конусовидную.
Так заканчивается кариотомия – деление ядра, затем наступает плазмотомия. Между дочерними клетками распределяются органеллы цитоплазмы, причем некоторые из них (диктиосомы, митохондрии и пластиды) претерпевают значительные видоизменения. Наконец, происходит цитокинез – образование клеточной стенки между дочерними ядрами. Из прежней клетки образовались две новые; в каждой из них имеется ядро, содержащее диплоидное число хромосом.
В зависимости от поведения ядерной оболочки у водорослей различают закрытый , полузакрытый и открытый митозы. При закрытом митозе расхождение хромосом происходит без нарушения ядерной оболочки. При полузакрытом митозе ядерная оболочка сохраняется в течение всего митоза, за исключением полярных зон. При открытом митозе ядерная оболочка исчезает в профазе. В зависимости от формы веретена деления различают плевромитоз и ортомитоз.
При плевромитозе в метафазе не образуется метафазная пластинка и веретено представлено двумя полуверетенами, расположенными под углом друг к другу вне или внутри ядра. При ортомитозе в метафазе хромосомы становятся по экватору двухполюсного веретена. В зависимости от комбинации указанных свойств у водорослей различают следующие типы митоза (рис. 7, 8):
Закрытый внеядерный митоз

Закрытый внутриядерный митоз


Полузакрытый митоз


Открытый митоз

Рис. 7. Схема основных типов митозов у водорослей (по: С.А. Карпов, год). Линии внутри или вне ядра – микротрубочки веретена деления
Центрами организации микротрубочек митотического веретена при полузакрытом ортомитозе могут быть кинетосомы и другие структуры:
– открытый ортомитоз, встречается у криптофитовых, золотистых, харовых;
– полузакрытый ортомитоз, встречается у зеленых, красных, бурых и др.;
– закрытый ортомитоз, встречается у евгленовых;
– закрытый плевромитоз внутриядерный или внеядерный встречается у некоторых динофитовых;
– полузакрытый митоз, во время метафазы центриоли находятся не на полюсах, а в области метафазной пластинки; можно наблюдать у требуксиевых зеленых.



Рис. 8. Диаграмма сравнения (А) закрытого, (Б) метацентрического и (В) открытого митозов (по: L.E. Graham, L.W. Wilcox, 2000)
При митозе варьирует также форма веретена и форма полюсов веретена, а аткже продолжительность существования межзонального веретена. Пик митозов приходится на темновой период суток. В многоядерных клетках деление ядер может происходить синхронно. асинхронно, волнообразно.
Контрольные вопросы
1. Назовите основные структурные элементы растительных клеток.
2. Отличие строения клеток водорослей от клеток высших растений.
3. Клеточные покровы водорослей.
4. Что такое тека? У каких водорослей она встречается?
5. Основные пигменты водорослей. Местонахождение пигментов в клетках водорослей.
6. Строение пластид.
7. Особенности строения пластид водорослей.
8. Строение митохондрий.
9. Особенности строения митохондрий водорослей.
10. Строение ядра и ядерных оболочек. Особенности ядерных оболочек в клетках водорослей.
11. Схема митоза. Характеристика фаз митоза.
12. Типы митоза в клетках водорослей.
13. Чем различаются плевромитоз и ортомитоз?
14. Типы псевдоподий водорослей.
2. ОБЩАЯ ХАРАКТЕРИСТИКА ВОДОРОСЛЕЙ
2.1. Типы питания
Основным типом питания у водорослей является фототрофный тип. Во всех отделах водорослей есть представители, являющиеся строгими (облигатными) фототрофами. Однако многие водоросли достаточно легко переключаются с фототрофного типа питания на ассимиляцию органических веществ, или гетеротрофный тип питания. Однако чаще всего переход на гетеротрофное питание у водорослей не ведет к полному прекращению фотосинтеза, то есть в таких случаях можно говорить о миксотрофном, или смешанном, типе питания.
Способность к росту на органических средах в темноте или на свету при отсутствии углекислого газа показана для многих синезеленых, зеленых, желтозеленых, диатомовых и др. Замечено, что у водорослей гетеротрофный рост идет медленнее, чем автотрофный на свету.
Многообразие и пластичность способов питания водорослей позволяет им иметь широкое распространение и занимать различные экологические ниши.
2.2. Типы талломов
Вегетативное тело водорослей представлено слоевищем , или талломом , не дифференцированным на органы – корень, стебель, лист. В пределах слоевищного строения водоросли отличаются очень большим морфологическим разнообразием (рис. 9). Они представлены одноклеточными, многоклеточными, неклеточными организмы. Размеры их колеблются в широких пределах: от мельчайших одноклеточных до гигантских многометровых организмов. Форма тела водорослей также многообразна: от простейших шаровидных до сложно расчлененных, напоминающих высшие растения, форм.
Огромное разнообразие водорослей можно свести к нескольким типам морфологической структуры: монадный , ризоподиальный , пальмеллоидный , коккоидный, трихальный, гетеротрихальный, паренхиматозный, сифональный, сифонокладальный .
Монадный (жгутиковый) тип структуры таллома
Наиболее характерным признаком, определяющим этот тип структуры, является наличие жгутиков, с помощью которых монадные организмы активно передвигаются в водной среде (рис. 9, А ). Подвижные жгутиковые формы широко распространены у водорослей. Жгутиковые формы доминируют среди многих групп водорослей: евгленовых, динофитовых, криптофитовых, рафидовых, золотистых, встречаются у желтозеленых и зеленых водорослей. У бурых водорослей монадный тип структуры в вегетативном состоянии отсутствует, однако монадные стадии образуются во время репродукции (размножения). Количество жгутиков, их длина, характер размещения и движения разнообразны и имеют важное систематическое значение.

Рис. 9. Морфологические типы строения талломов у водорослей (по:): А – монадный (Хламидомонас ); Б – амебоидный (Ризохризис ); В – пальмеллоидный (Гидрурус ); Г – коккоидный (Педиаструм ); Д – сарциноидный (Хлоросарцина ); Е – нитчатый (Улотрикс ); Ж – разнонитчатый (Фритчиела ); З, И – тканевой (Фурцелярия, Ламинария ); К – сифональный (Каулерпа ); Л – сифонокладальный (Кладофора )
Подвижность монадных водорослей обуславливает полярность строения их клеток и колоний. Обычно на переднем полюсе клетки или вблизи от него прикрепляются жгутики. Основная форма клетки – каплевидная с более или менее суженным передним жгутиковым полюсом. Однако нередко монадные организмы отклоняются от этой основной формы и могут быть ассиметричными, спиралевидными, иметь суженный задний конец и т. д.
Форма клетки в значительной степени зависит от клеточных покровов, отличающихся большим разнообразием (плазмалемма; пелликула; тека; состоящая из органических, кремнеземных или известковых чешуек; домик; клеточная оболочка). Причудливые очертания клеток некоторых золотистых водорослей формирует своеобразный внутриклеточный скелет, состоящий из полых внутри кремнеземных трубочек. Клеточная оболочка обычно гладкая, иногда несет разнообразные выросты либо инкрустирована солями железа или кальция и тогда напоминает домик. В оболочке образуются лишь маленькие отверстия для выхода жгутиков.
Полярность монадных организмов проявляется и в расположении внутриклеточных структур. На переднем конце клетки нередко расположена разнообразно устроенная глотка , обычно выполняющая экскреторную функцию. Лишь у немногих фаготрофных жгутиковых глотка функционирует как клеточный рот - цитостом .
Своеобразными органеллами, свойственными водорослям, имеющим монадную структуру, являются сократительные вакуоли , выполняющие осморегуляторную функцию, слизистые тельца и стрекательные структуры . Стрекательные капсулы встречаются у динофитовых, евгленовых, золотистых, рафидофитовых, криптофитовых водорослей и выполняют защитную функцию. Единственное ядро занимает в клетках центральное положение. Хлоропласты, разнообразные по форме и окраске, могут быть осевыми или пристенными.
Тенденция к увеличению размеров тела проявляется в образовании разнообразных колоний. В простейших случаях колонии образуются вследствии нерасхождения делящихся клеток. Наблюдаются колонии кольцевидной, кустистой, древовидной, шаровидной формы. Зеленым монадным организмам большей частью свойственны колонии типа ценобиев с постоянным для каждого вида числом клеток.
В неблагоприятных условиях монадные организмы сбрасывают или втягивают свои жгутики, теряя при этом подвижность, и окружают себя обильной слизью.
Монадный тип структуры оказался перспективным. На его основе развились другие, более сложные структуры.
Ризоподиальный (амебоидный) тип структуры
Наибоее существенные признаки амебоидого типа структуры – отсутствие прочных клеточных покровов и способность к амебоидному движению, с помощью временно образующихся на поверхности клетки цитоплазматических выростов – псевдоподий . Различают несколько разновидностей псевдоподий, из которых у водорослей чаще всего наблюдаются ризоподии и лодоподии , реже аксоподии (рис. 9, Б ).
Принципиальных различий в строении и механизме действия сократительных систем, обусловливающих подвижность монадных и амебоидных организмов, на молекулярном уровне не существует. Амебоидное движение возникло, вероятно, в результате приспособления жгутиковых клеток к упрощенным условиям жизни, что привело к упрощению строения тела.
В клетках амебоидных водорослей имеются ядра, пластиды, митохондрии и другие органеллы, свойственные эукариотам: нередко наблюдаются сократительные вакуоли, стигма и базальные тела, способные образовывать жгутики.
Многие амебоидные организмы ведут прикрепленный образ жизни. Они могут строить домики различной формы и структуры: тонкие, нежные, или грубые, толстостенные.
Амебоидный тип тела распространен не столь широко, как монадный. Он наблюдается лишь у золотистых и желтозеленых водорослей.
Пальмеллоидный (гемимонадный) тип структуры
Характерным для этого типа структуры является сочетание неподвижного растительного образа жизни с наличием клеточных органелл, свойственных монадным организмам: сократительные вакуоли, стигма, жгутики. Так, вегетативные клетки могут иметь жгутики, с помощью которых они ограниченно передвигаются в пределах колониальной слизи, либо жгутики сохраняются в неподвижных клетках в сильно редуцированном виде.
Для клеток с пальмеллоидным (гемимонадным) типом характерно полярное строение. Иногда клетки находятся в домиках.
Гемимонадные водоросли нередко образуют колонии. В простейшем случае слизь бесструктурна, и клетки располагаются внутри нее без определенного порядка. Дальнейшее усложнение таких колоний проявляется как в дифференциации слизи, так и в более упорядоченном расположении клеток внутри слизи. Наиболее сложно устроены колонии дендровидного типа (род Гидрурус ) (рис. 9, В ).
Пальмеллоидный (гемимонадный) тип структуры был важным этапом на пути морфологической эволюции водорослей в направлении от подвижных монадных к типично растительным неподвижным формам.
Коккоидный тип структуры
Этот тип объединяет одноклеточные и колониальные водоросли, неподвижные в вегетативном состоянии. Клетки коккоидного типа одеты оболочкой и имеют протопласт растительного типа (тонопласт без сократительных вакуолей, стигм, жгутиков). Утрата признаков монадной структуры в строении клетки у организмов, ведущих растительный неподвижный образ жизни, приобретение новых структур, свойственных растительным клеткам – следующий крупный шаг в эволюции водорослей по растительному типу.
Огромное разнообразие водорослей коккоидного типа структуры связано с наличием клеточных покровов. Покровы обуславливают наличие различных клеток: шаровидных, яйцевидных, веретеновидных, эллипсоидных, цилиндрических, звездчато-лопастных, спиралевидных, грушевидных и др. Многообразие форм также умножается благодаря скульптурным украшениям клеточных покровов – шипам, шипикам, щетинкам, роговым отросткам.
Коккоидные водоросли образуют колонии разнообразной формы, в которых клетки объединены с помощью слизи или без нее.
Коккоидный тип структуры широко распространен почти во всех отделах эукариотических водорослей (за исключением евгленовых).
В эволюционном плане кокоидную структуру можно рассматривать как исходную для возникновения многоклеточных слоевищ, а также сифонального и сифонокладального типов структуры (рис. 9, Г, Д ).
Трихальный (нитчатый) тип структуры
Характерной особенностью нитчатого типа структуры является нитевидное расположение неподвижных клеток, которые образуются вегетативным путем в результате клеточного деления, происходящего преимущественно в одной плоскости. Клетки нитей имеют полярное строение и могут расти лишь в одном направлении, совпадающем с осью ядерного веретена.
В простейших случаях слоевища нитчатой структуры слагаются из клеток, морфологически подобных друг другу. Вместе с тем у многих водорослей на участках нитей, утончающихся или расширяющихся к концам, клетки отличаются по форме от остальных. При этом нередко нижняя клетка, лишенная хлоропластов, превращается в бесцветный ризоид или стопу. Нити могут быть простыми или ветвящимися, одно- или многорядными, свободноживущими или прикрепленными.
Нитчатый тип структуры представлен среди зеленых, красных, желтозеленых, золотистых водорослей (рис. 9, Е ).
Гетеротрихальный (разнонитчатый) тип структуры
Разнонитчатый тип возник на базе нитчатого типа. Разнонитчатое слоевище состоит большей частью из горизонтальных, стелящихся по субстрату нитей, выполняющих функцию прикрепления, и вертикальных, поднимающихся над субстратом нитей, выполняющих ассимиляционную функцию. Последние несут на себе органы размножения.
У некоторых водорослей вертикальные нити дифференцированы на междоузлия и узлы , от которых отходят мутовки боковых ветвей, имеющих также членистое строение. От узлов, кроме того, могут отрастать дополнительные нити, образующие коровое покрытие междоузлий. Функцию прикрепления к субстрату выполняют бесцветные ризоиды. Такое строение можно встретить у харовых, зеленых, бурых, красных, некоторых желтозеленых и золотистых водорослей (рис. 9, Ж ).
Паренхиматозный (тканевой) тип структуры
Одно из направлений эволюции разнонитчатого слоевища было связано с возникновением паренхиматозных слоевищ. Способность к неограниченному росту и делению клеток в разных направлениях привела к образованию объемных макроскопических слоевищ с морфофункциональной дифференцировкой клеток в зависимости от положения в слоевище (коровой, промежуточный слой, сердцевина).
В пределах данного типа наблюдается постепенное усложнение слоевищ от простых пластин до сложно дифференцированных слоевищ с примитивными тканями и органами. Паренхиматозный тип структуры - наивысшая эволюционная ступень морфологической дифференциации тела водорослей. Он широко представлен у крупных водорослей: бурых, красных и зеленых - так называемых водорослей-макрофитов (рис. 10).

Рис. 10. Поперечный разрез слоевища бурой водоросли (по:): 1 – наружная кора; 2 – внутренняя кора; 3 – сердцевина
Сифональный тип структуры
Сифональный (неклеточный) тип структуры характеризуется отсутствием внутри слоевища, достигающего сравнительно крупных, обычно макроскопических размеров и определенной степени дифференцировки, клеточных перегородок при наличии большого количества органелл. Перегородки в таком слоевище могут появляться лишь случайно, при его повреждении, или при образовании репродуктивных органов. В обоих случаях процесс образования перегородок отличается от формирования многоклеточного организма.
Сифональный тип структуры представлен у некоторых зеленых и желтозеленых водорослей. Однако это направление морфологической эволюции оказалось тупиковым.
Сифонокладальный тип структуры
Основным признаком сифонокладального типа структуры является способность к образованию из первичного неклеточного слоевища сложно устроенных слоевищ, состоящих из первично многоядерных сегментов. В основе формирования такого слоевища лежит сегрегативное деление , при котором митоз не всегда заканчивается цитокинезом.
Сифонокладальный тип структуры известен лишь у небольшой группы морских зеленых водорослей.
2.3. Размножение водорослей
Размножение является основным свойством живых существ. Его сущность заключается в воспроизведении себе подобных. У водорослей размножение может осуществляться бесполым, вегетативным и половым путем.
Бесполое размножение
Бесполое размножение водорослей осуществляется с помощью специализированных клеток – спор . Спорообразование обычно сопровождается делением протопласта на части и выходом продуктов деления из оболочки материнской клетки. При этом перед делением протопласта в нем происходят процессы, ведущие к его омоложению. Выход продуктов деления из оболочки материнской клетки является наиболее существенным отличием настоящего бесполого размножения от вегетативного. Иногда в клетке образуется только одна спора, но и тогда она покидает материнскую оболочку.
Споры обычно образуются в особых клетках, называемых спорангиями , отличающихся от обычных вегетативных клеток размерами и формой. Они возникают как выросты обычных клеток и выполняют только функцию образования спор. Иногда споры образуются в клетках, не отличающихся формой и размерами от обычных вегетативных клеток. Споры также отличаются от вегетативных клеток формой и более мелкими размерами. Количество спор в спорангии колеблется от одной до нескольких сотен. Споры представляют собой расселительную стадию в жизненном цикле водорослей.
В зависимости от строения различают:
зооспоры – подвижные споры зеленых и бурых водорослей, могут иметь один, два, четыре или много жгутиков, в последнем случае жгутики располагаются венчиком у переднего конца споры или парами по всей поверхности;
гемизооспоры – зооспоры, утратившие жгутики, но сохранившие сократительные вакуоли и стигму;
апланоспоры – неподвижные споры, которые одеваются оболочкой внутри материнской клетки;
автоспоры – апланоспоры, имеющие форму материнской клетки;
гипноспоры – неподвижные споры с утолщенными оболочками, предназначенные для переживания неблагоприятных условий среды.
У красных водорослей бесполое размножение осуществляется при помощи моноспор , биспор, тетраспор или полиспор . Моноспоры не имеют жгутика и оболочки. После выхода из материнской клетки, они способны к амебоидному движению. От вегетативных клеток моноспоры отличаются яйцевидной или шаровидной формой, богатым содержанием питательных веществ и интенсивной окраской.
Строение спор и типы спороношения имеют большое значение для систематики водорослей, так как отражают различия в организации предковых форм различных групп водорослей.
Вегетативное размножение
Вегетативное размножение у водорослей может осуществляться несколькими путями: простым делением надвое, множественным делением, почкованием, фрагментацией слоевища, столонами, выводковыми почками, параспорами, клубеньками, акинетами.
Простое деление.
Этот способ размножения встречается только у одноклеточных форм водорослей. Наиболее просто происходит деление у клеток, имеющих амебоидный тип строения тела.
Деление амебоидных форм . Деление амебоидов возможно в любом направлении. Оно начинается с вытягивания тела амебы, и далее на экваторе намечается перегородка, которая делит тело на две более или менее равные части. Деление цитоплазмы сопровождается делением ядра. Иногда делению предшествует переход в неподвижное состояние за счет втягивания ножек, при этом клетка приобретает шаровидную форму. Одновременно протоплазма теряет прозрачность, сократительная вакуоль исчезает. К концу деления происходит вытягивание клетки, ее перешнуровка, затем появляются ложноножки.
Деление жгутиковых форм . У жгутиковых форм встречаются наиболее сложные типы вегетативного размножения. Типы размножения определяются уровнем организации и степенью полярности клеток. У некоторых криптофитовых, золотистых и зеленых водорослей размножение простым делением надвое происходит в подвижном состоянии только по продольной оси и начинается с переднего полюса клетки. Жгутики при этом могут достаться только одной клетке или поровну разделиться между новыми клетками. Клетка, которой не досталось жгутика, образует его сама. У большинства вольвоксовых и евгленовых водорослей во время размножения оболочка клетки ослизняется и деление происходит в неподвижном состоянии. У всех жгутиковых форм, имеющих панцирь, клетки делятся на две равные или неравные части. После разделения старый панцирь сбрасывается и образуется новый.
Деление коккоидных форм . У водорослей с коккоидным типом структуры клеток вегетативное размножение приобретает типичные черты деления неподвижной растительной клетки с хорошо выраженной клеточной оболочкой. По своей простоте оно приближается к амебоидному типу вегетативного размножения и осуществляется простым делением клетки надвое.
Почкование.
Клеткам нитчатых разветвленных водорослей свойственны два пути вегетативного размножения: простым делением надвое и почкованием. Сочетание этих способов размножения обуславливает боковое ветвление нитчатых водорослей.
Фрагментация.
Фрагментация присуща всем группам многоклеточных водорослей и проявляется в разных формах: образование гормогониев, регенерация оторвавшихся частей слоевища, спонтанное отпадание ветвей, отрастание ризоидов. Причиной фрагментации могут быть механические факторы (волны, течение, погрызы животных), отмирание части клеток. Примером последнего способа фрагментации может быть образование у синезеленых водорослей гормогониев. Каждый гормогоний может дать начало новой особи. Размножение частями слоевищ, характерное для красных и бурых водорослей, не всегда приводит к возобновлению нормальных растений. Морские водоросли, растущие на камнях и скалах, нередко частично или полностью разрушаются под действием волн. Их оторвавшиеся фрагменты или целые слоевища не способны снова закрепиться на твердых грунтах из-за постоянного движения воды. Кроме того, органы прикрепления вновь не образуются. Если такие слоевища попадают в спокойные места с илистым или песчаным дном, они продолжают расти, лежа на грунте. Со временем более старые части отмирают и отходящие от них ветви превращаются в самостоятельные слоевища, в подобных случаях говорят о неприкрепленных, или свободноживущих, формах соответствующих видов. Водоросли сильно видоизменяются: ветви их становятся более тонкими, узкими и слабее разветвляются. Неприкрепленные формы водорослей не образуют органов полового и бесполого размножения и могут размножаться только вегетативно.
Размножение побегами, столонами, выводковыми почками, клубеньками, акинетами.
У тканевых форм зеленых, бурых и красных водорослей вегетативное размножение приобретает свою законченную форму, которая мало отличается от вегетативного размножения высших растений. Сохраняя способность к регенерации частей слоевища, тканевые формы приобретают специализированные образования, которые выполняют функцию вегетативного размножения. У многих видов бурых, красных, зеленых и харовых водорослей имеются побеги, на которых вырастают новые слоевища. На слоевищах некоторых бурых и красных водорослей развиваются выводковые почки (пропагулы), которые отпадают и прорастают в новые слоевища.
С помощью одноклеточных или многоклеточных зимующих клубеньков происходит сезонное возобновление харовых водорослей. Некоторые нитчатые водоросли (например, зеленые улотриксовые водоросли) размножаются акинетами – специализированными клетками с утолщенной оболочкой и большим количеством запасных питательных веществ. Они способны переживать неблагоприятные условия.
Половое размножение
Половое размножение у водорослей связано с половым процессом, который заключается в слиянии двух клеток, в результате чего образуется зигота, вырастающая в новую особь или дающая зооспоры.
Половое размножение у водорослей бывает нескольких типов:
гологамия (коньюгация) – без образования специализированных клеток;
гаметогамия – с помощью специализированных клеток – гамет.
Гологамия . В простейшем случае процесс происходит путем слияния двух неподвижных, лишенных клеточных оболочек вегетативных клеток. У одноклеточных жгутиковых форм водорослей половой процесс осуществляется путем слияния двух особей.
При слиянии содержимого двух безжгутиковых вегетативных клеток половой процесс называют коньюгацией . Во время коньюгации происходит слияние двух клеток, которые выполняют функцию половых клеток – гамет. Слияние содержимого клеток происходит через специально образующийся коньюгационный канал, получается зигота, которая впоследствии покрывается толстой оболочкой и превращается в зигоспору. Если скорость перетекания содержимого клеток одинакова, зигота образуется в коньюгационном канале. В этом случае разделение клеток на мужскую и женскую условно.
Гаметогамия . Половое размножение у водорослей, в том числе и одноклеточных, чаще происходит путем деления содержимого клеток и образования в них специализированных половых клеток – гамет. У всех зеленых и бурых водорослей мужские гаметы имеют жгутики, а женские гаметы имеют их не всегда. У примитивных водорослей гаметы образуются в вегетативных клетках. У более высокоорганизованных форм гаметы находятся в особых клетках, называемых гаметангиями. В вегетативной клетке или гаметангии может быть от одной до нескольких сотен гамет. В зависимости от размеров сливающихся гамет различают несколько типов гаметогамии: изогамия, гетерогамия, оогамия.
Если сливающиеся гаметы имеют одинаковую форму и размеры, такой половой процесс называют изогамией .
Если сливающиеся гаметы имеют одинаковую форму, но разные размеры (женская гамета крупнее мужской), то говорят о гетерогамии .
Половой процесс, при котором сливаются неподвижная крупная клетка – яйцеклетка и подвижная мелкая мужская клетка – сперматозоид , называется оогамией . Гаметангии с яйцеклетками называют архегониями или оогониями , а со сперматозоидами – антеридиями . Мужские и женские гаметы могут развиваться на одной особи (однодомные) или на разных особях (двудомные). Образующаяся в результате слияния гамет зигота после некоторых изменений превращается в зигоспору. Последняя обычно одета плотной оболочкой. Зигоспора может находиться длительное время в покое (до нескольких месяцев) или прорастать без периода покоя.
Автогамия . Особый тип полового процесса. Он заключается в том, что ядро клетки делится мейотически, из четырех образовавшихся ядер, два разрушаются, а оставшиеся два ядра сливаются, образуя зиготу, которая без периода покоя увеличивается в размерах и превращается в ауксоспору. Так происходит омоложение особей.
2.4. Жизненные циклы водорослей
Жизненный цикл, или цикл развития , - это совокупность всех этапов развития организмов, в результате прохождения которых из определенных особей или их зачатков образуются новые, сходные с ними особи и зачатки. Этап старения, ведущий к отмиранию особи, и периоды покоя выходят за пределы жизненного цикла. Цикл развития бывает простым или сложным, что связано с соотношением диплоидных и гаплоидных ядерных фаз, или форм развития (рис. 11).

Рис. 11. Жизненные циклы водорослей (по:): I – гаплобионтный с зиготической редукцией; II – гаплодиплобионтный со спорической редукцией; III – диплобионтный с гаметической редукцией; IV – гаплодиплобионтный с соматической редукцией. Доминирующая фаза в случаях I и III бывает многоклеточной; если она одноклеточная, то долговременна и способна к митотическому воспроизведению; 1 – гаплоидная фаза; 2 – диплоидная фаза
Понятие жизненного цикла связано с чередованием поколений. Под поколением понимают совокупность особей, рассматриваемых по отношению к предкам и потомкам, живущим в близкое время, и генетически с ним связанных.
Простой жизненный цикл характерен для цианобактерий, у которых не обнаружено половое размножение. У них жизненные циклы бывают полными (большими ) и малыми. Малый жизненный цикл соответствует определенным ответвлениям большого цикла и приводит к повторному образованию промежуточных возрастных состояний индивидов цианобактерий. Цикл развития цианбактерий, таким образом, включает определенные отрезки развития одного или ряда последовательных поколения конкретной систематической формы: от зачатка особи до возникновения новых зачатков такого же типа.
У большинства водорослей с половым процессом, в зависимости от времени года и внешних условий, наблюдаются разные формы размножения (половое и бесполое), при этом происходит смена гаплоидной и диплоидной ядерных фаз. Изменения, претерпеваемые индивидом между одноименными фазами развития, и составляют его жизненный цикл.
Органы полового и бесполого размножения могут развиваться на одной и той же особи или на разных особях. Растения, образующие споры, называются спорофитами , а образующие гаметы – гаметофитами . Растения, способные производить и споры и гаметы, называют гаметоспорофитами . Гаметоспорофиты характерны для многих водорослей: зеленых (Ульвовые), бурых (Эктокарповые) и красных (Бангиевые). Развитие органов размножения того или иного типа определяется температурой среды. Например, на пластинчатых слоевищах красной водоросли Порфира Тенера при температуре ниже 15–17 °C образуются органы полового размножения, а при более высокой температуре – органы бесполого размножения. В общем, у многих водорослей гаметы развиваются при более низкой температуре, чем споры. На развитие тех или иных органов размножения влияют и другие факторы: интенсивность света, продолжительность дня, химический состав воды, в том числе ее соленость.
Гаметофиты, гаметоспорофиты и спорофиты водорослей могут внешне не отличаться или иметь хорошо выраженные морфологические отличия. Различают изоморфную (сходную) и гетероморфную (разную) смену форм развития, которую отождествляют с чередованием поколений. У большинства гаметоспорофитов чередование поколений не происходит. Иногда гаметофиты и спорофиты, не различаясь морфологически, существуют в разных экологических условиях; в некоторых случаях они отличаются и морфологически. Например, у красной водоросли Порфира Тенера спорофиты имеют вид ветвящихся однорядных нитей, которые внедряются в известковый субстрат (раковины моллюсков, скалы). Они растут предпочтительно при слабом освещении и проникают в субстрат на большую глубину. Гаметофиты этой водоросли имеют вид пластинок, растут при хорошем освещении вблизи уреза воды и в приливно-отливной зоне.
При гетероморфном чередовании поколений строение спорофитов и гаметофитов различается в некоторых случаях довольно значительно. Так, у зеленых водорослей из родов Акросифония и Спонгоморфа гаметофит многоклеточный, высотой несколько сантиметров, а спорофит одноклеточный, микроскопический. Возможны и другие соотношения размеров гаметофита и спорофита. У бурой водоросли Сахарины гаметофит микроскопический, а спорофит длиной до 12 м. У большинства водорослей гаметофиты и спорофиты представляют собой самостоятельные растения. У ряда видов красных водорослей спорофиты растут на гаметофитах, а у некоторых бурых водорослей гаметофиты развиваются внутри слоевища спорофита.
Гетероморфная смена форм развития, когда наблюдается четко выраженное обособление спорофита от гаметофита, характерна для более высокоорганизованных групп водорослей. При этом одна из форм, чаще всего гаметофит, бывает микроскопической. Считают, что гетероморфный цикл развития водорослей возник из изоморфного. Способы развития гаметофита и спорофита имеют большое значение в систематике водорослей. Наиболее сложные и многообразные циклы развития, не встречающиеся у других водорослей, характерны для красных водорослей.
Смена ядерных фаз.
При половом процессе в результате слияния гамет и их ядер происходит удвоение числа хромосом в ядре. На определенном этапе цикла развития, при мейозе, происходит редукция числа хромосом, в результате которой образующиеся ядра получают одинарный набор хромосом. Спорофиты многих водорослей диплоидны, и мейоз в цикле их развития совпадает с моментом образования спор, из которых развиваются гаплоидные гаметоспорофиты или гаметофиты. Такой мейоз называют спорической редукцией . Спорофиты более примитивных красных водорослей (родов Кладофора, Эктокарпус и многие другие) наряду с гаплоидными спорами образуют диплоидные споры, которые снова развиваются в спорофиты. Споры, появляющиеся на гаметоспорофитах, служат для самовозобновления материнских растений. Спорофиты и гаметофиты водорослей, стоящих на высших ступенях эволюции, строго чередуются без самовозобновления.
У ряда водорослей мейоз происходит в зиготе. Такой мейоз называют зиготической редукцией и встречается у ряда видов зеленых и харовых водорослей. У пресноводных вольвоксовых и улотриксовых водорослей спорофит представлен одноклеточной зиготой, которая производит до 32 зооспор, по массе во много раз превышающих родительские гаметы, т.е. по существу наблюдается спорическая редукция.
Некоторые группы водорослей имеют гаметическую редукцию , которая характерна для животных, а не для растительных организмов. У этих водорослей мейоз происходит при образовании гамет, при этом остальные клетки слоевища остаются диплоидными. Такая смена ядерных фаз присуща широко распространенным по всему земному шару диатомовым водорослям и бурым фукусовым (которые включают наиболее массовые морские виды водорослей), а из зеленых – крупному роду Кладофора. Развитие с гаметической редукцией ядра, как полагают, дает указанным водорослям определенные преимущества по сравнению с другими.
Если редукционное деление происходит в спорангиях перед образованием спор бесполого размножения (спорическая редукция), то имеет место чередование поколений – диплоидного спорофита и гаплоидного гаметофита. Этот тип жизненного цикла называется гаплобионтным со спорической редукцией . Он характерен для некоторых зеленых водорослей, многих бурых и красных водорослей.
Наконец, у немногих водорослей мейоз происходит в вегетативных клетках диплоидного таллома (соматическая редукция), из которых затем развиваются гаплоидные талломы. Такой жизненный цикл с соматической редукцией известен у красных и зеленых водорослей.
Контрольные вопросы
Типы питания водорослей.
Типы таллома водорослей.
Характеристика монадной морфологической структуры.
Характеристика ризоподиальной морфологической структуры. Типы цитоплазматических выростов.
Характеристика пальмеллоидной морфологической структуры.
Характеристика коккоидной морфологической структуры.
Характеристика трихальной морфологической структуры.
Характеристика гетеротрихальной морфологической структуры.
Характеристика паренхиматозной морфологической структуры.
Характеристика сифональной морфологической структуры.
Характеристика сифонокладальной морфологической структуры.
12. Бесполое размножение. Типы спор.
13. Типы вегетативного размножения водорослей.
14. Типы полового размножения водорослей.
15. Чем различаются спорофит и гаметофит?
16. Что такое гетероморфная и изоморфная смена генераций?
17. Смена ядерных фаз в жизненном цикле водорослей. Спорическая, зиготическая и гаметическая редукция.
3. ЭКОЛОГИЧЕСКИЕ ГРУППЫ ВОДОРОСЛЕЙ
Водоросли распространены по всему земному шару и встречаются в различных водных, наземных и почвенных биотопах. Известны разнообразные экологические группы: водоросли водных мест обитания, наземные водоросли, почвенные водоросли, водоросли горячих источников, водоросли снега и льда, водоросли гиперсоленых источников.
3.1. Водоросли водных местообитаний
3.1.1. Фитопланктон
Термин «фитопланктон» означает совокупность растительных организмов, парящих в толще воды. Планктонные водоросли – основной, а в некоторых случаях и единственный, продуцент первичного органического вещества, на базе которого существует все живое в водоеме. Продуктивность фитопланктона зависит от комплекса разнообразных факторов.
Планктонные водоросли обитают в разнообразных водоемах – от океана до лужи. Причем большее разнообразие экологических условий во внутренних водоемах по сравнению с морями определяет и значительно большее разнообразие видового состава и экологических комплексов пресноводного планктона.
Фитопланктон пресноводных экосистем характеризуется четко выраженной сезонностью. В каждый сезон в водоеме преобладает одна или несколько групп водорослей, а в периоды интенсивного развития господствует нередко всего один вид. Так зимой подо льдом (особенно когда лед покрыт снегом) фитопланктон очень беден или почти отсутствует из-за недостатка света. Вегетационное развитие водорослей планктона как сообщества начинается в марте – апреле, когда уровень солнечного света становится достаточным для фотосинтеза водорослей даже подо льдом. В это время появляются довольно многочисленные мелкие жгутиковые – евгленовые, динофитовые, золотистые, а также холоднолюбивые диатомовые. В период вскрытия льда до установления температурной стратификации, что обычно бывает при прогреве верхнего слоя воды до 10–12C°, начинается бурное развитие холодолюбивого комплекса диатомовых водорослей. Летом, при температуре воды выше 15C°, наблюдается максимум продуктивности синезеленых, евгленовых и зеленых водорослей. В зависимости от трофического и лимнологического типа водоема в это время может происходить «цветение» воды, вызванное развитием синезеленых и зеленых водорослей.
Одной из существенных особенностей пресноводного фитопланктона является обилие в нем временно планктонных водорослей. Ряд видов, которые принято считать типично планктонными, в прудах и озерах имеют донную или перифитонную (прикрепленную к какому-либо предмету) фазу в своем развитии.
Морской фитопланктон состоит в основном из диатомовых и динофитовых водорослей. Из диатомовых особенно многочисленны представители родов Хетоцерос, Ризосоления, Талассиозира и некоторые другие, отсутствующие в пресноводном планктоне. Очень разнообразен в морском фитопланктоне состав жгутиковых форм динофитовых водорослей. Весьма многочисленны в морском фитопланктоне представители примнезиофитовых, представленные в пресных водах лишь несколькими видами. Хотя морская среда на значительных пространствах относительно однородна, в распределении морского фитопланктона аналогичной однородности не наблюдается. Различия по видовому составу и численности нередко выражены даже на сравнительно небольших акваториях морских вод, но особенно четко они отражаются в крупномасштабной географической зональности распределения. Здесь проявляется экологическое действие основных факторов среды: солености воды, температуры, освещенности и содержания питательных веществ.
Планктонные водоросли обычно имеют специальные приспособления к обитанию во взвешенном состоянии. Одни имеют разного рода выросты и придатки тела – шипы, щетинки, роговые выросты, перепонки. Другие образуют колонии, обильно выделяющие слизь. Третьи накапливают в теле вещества, повышающие их плавучесть (капли жира у диатомовых, газовые вакуоли у синезеленых). Эти образования гораздо сильнее развиты у морских фитопланктеров, чем у пресноводных. Одним из приспособлений к существованию в толще воды во взвешенном состоянии являются мелкие размеры тела планктонных водорослей.
3.1.2. Фитобентос
К фитобентосу относится совокупность растительных организмов, которые приспособлены к существованию в прикрепленном или неприкрепленном состоянии на дне водоемов и на разнообразных предметах, живых и мертвых организмах, находящихся в воде.
Возможность произрастания бентосных водорослей в конкретных местообитаниях определяется как абиотическими, так и биотическими факторами. Среди биотических факторов существенную роль играет конкуренция с другими водорослями и присутствие консументов. Это приводит к тому, что отдельные виды бентосных водорослей растут далеко не на всякой глубине и не во всех водоемах с подходящим световым и гидрохимическим режимом. Для роста бентосных водорослей как фотосинтезирующих организмов особенно важен свет. Но на степень его использования влияют другие экологические факторы: температуры, содержания биогенных и биологически активных веществ, кислорода и неорганических источников углерода, а главное – темпы поступления этих веществ в слоевище, что находится в зависимости от концентрации веществ и скорости движения воды. Как правило, места с интенсивным движением воды отличаются пышным развитием бентосных водорослей.
Бентосные водоросли, растущие в условиях активного движения воды , получают преимущества по сравнению с водорослями, растущими в малоподвижных водах. Один и тот же уровень фотосинтеза может быть достигнут организмами фитобентоса в условиях течения при меньшей освещенности, что способствует росту более крупных слоевищ. Движение воды, к тому же, предотвращает оседание на скалы и камни илистых частиц, которые мешают закреплению зачатков водорослей, благоприятствуют росту донных водорослей, смывая с поверхности грунта животных, питающихся водорослями. Наконец, хотя при сильном течении или сильном прибое происходит повреждение слоевищ водорослей или отрыв их от грунта, движение воды все же не препятствует поселению микроскопических видов водорослей или микроскопических стадий водорослей-макрофитов.
Влияние движения воды на развитие бентосных водорослей особенно ощутимо в реках, ручьях и горных потоках. В этих водоемах выделяется группа бентосных организмов, предпочитающих места с сильным течением. В озерах, где не бывает сильных течений, основное значение приобретает волновое движение. В морях волны также оказывают значительное влияние на жизнь бентосных водорослей, в частности на их вертикальное распределение.
В северных морях на распространение и численность бентосных водорослей оказывает влияние лед . В зависимости от его толщины, движения и торошения заросли водорослей могут быть уничтожены (стерты) до глубины в несколько метров. Поэтому, например, в Арктике, многолетние бурые водоросли (Фукус, Ламинария ) легче всего найти у берега среди валунов и выступов скал, препятствующих движению льда.
Многообразное влияние на жизнь бентосных водорослей оказывает температура . Наряду с другими факторами она определяет их скорость роста, темп и направление развития, момент закладки у них органов размножения, географическую зональность распространения.
Интенсивному развитию водорослей способствует также умеренное содержание в воде биогенных элементов . В пресных водах такие условия создаются в неглубоких прудах, в прибрежной зоне озер, в речных заводях, в морях – в мелких заливах.
Если в таких местах существует достаточное освещение, твердые грунты и слабое движение воды, то создаются оптимальные условия для жизни фитобентоса. При отсутствии движения воды и ее недостаточном обогащении биогенными веществами, бентосные водоросли растут плохо. Такие условия существуют в скалистых бухтах с большим уклоном дна и значительными глубинами в центре, так как биогенные вещества из донных отложений не выносятся в верхние горизонты. К тому же макроскопические морские водоросли, служащие субстратом для многих мелких форм бентосных водорослей, в подобных местообитаниях могут отсутствовать.
Источниками биогенных веществ в воде служат береговые стоки и донные отложения. Особенно велика роль последних как аккумуляторов органических остатков. В донных отложениях в результате жизнедеятельности бактерий и грибов происходит минерализация органических остатков; сложные органические вещества переходят в простые неорганические соединения, доступные для использования фотосинтезирующими растениями.
Кроме света, движения воды, температуры и содержания биогенных веществ, произрастание бентосных водорослей зависит от присутствия растительноядных водных животных – морских ежей, брюхоногих моллюсков, ракообразных, рыб. Это особенно заметно по зарослям ламинариевых водорослей, отличающихся крупными размерами. В тропических морях в некоторых местах рыбы полностью выедают зеленые, бурые и красные водоросли с мягким слоевищем. Брюхоногие моллюски, ползая по дну, поедают микроскопические водоросли и мелкие проростки макроскопических видов.
Преобладающими донными водорослями континентальных водоемов являются диатомовые, зеленые, синезеленые и желтозеленые нитчатые водоросли, прикрепленные или не прикрепленные к субстрату.
Основные бентосные водоросли морей и океанов – бурые и красные, иногда зеленые макроскопические прикрепленные слоевищные формы. Все они могут обрастать мелкими диатомовыми, синезелеными и другими водорослями.
В зависимости от места произрастания среди бентосных водорослей различают следующие экологические группы:
эпилиты – растут на поверхности твердого грунта (скалах, камнях);
эпипелиты – населяют поверхность рыхлых грунтов (песок, ил);
эпифиты – эпизоиты – живут на поверхности растений/ животных;
эндофиты – эндозоиты, или эндосимбионты – живут внутри тела растений/ животных, но питаются самостоятельно (имеют хлоропласты и фотосинтезируют);
эндолиты – живут в известковом субстрате (скалы, раковины моллюсков, панцири ракообразных).
Иногда выделяют группу организмов обрастания , или перифитон . Организмы, входящие в эту группу, живут на предметах большей частью движущихся или обтекаемых водой. Кроме того, они удалены от дна и находятся в условиях иного светового, пищевого и температурного режима, чем организмы истинно донных обитаний.
В состав обрастания входят микроводоросли и водоросли-макрофиты. Микроскопические водоросли (синезеленые и диатомовые) формируют слизистую бактериально-водорослево-детритную пленку на внесенном в водную среду субстрате. Затем на первичной микропленке вместе с животными поселяются макроводоросли (красные, бурые и зеленые). Это создает серьезные помехи в хозяйственной деятельности человека. Из-за обрастателей уменьшается скорость движения судов и эффективность работы гидроакустических приборов, увеличивается расход топлива, происходит утяжеление и коррозия подводных конструкций. Кроме того, слизистая пленка, образуемая обрастателями, может нарушать эксплуатацию водопроводных труб, засорять отверстия водозаборов и трубопроводов, нарушать процессы теплообмена в холодильных установках.
Прикрепленные организмы обрастания, обитающие на подводных сооружениях в приливно-отливной зоне и на глубине до 1 м, в зимний период обычно элиминируют под воздействием длительного осушения и истирания льдом. Поэтому ежегодно в весенне-летний период здесь формируются сообщества обрастания, характерные для пионерной стадии биологической сукцессии. Доминирующими видами таких сообществ, наряду с усоногими раками и моллюсками, часто являются водоросли-макрофиты. В сублиторальной зоне подводных сооружений - с глубины 0,7-0,9 м до их основания (6-12 м) - развивается многолетнее обрастание. В его составе преобладают бурые водоросли из родов Сахарина и Костария . Биомасса этих крупных водорослей в умеренных широтах может быть очень значительной, и исчисляться десятками килограммов на квадратный метр.
Водоросли-обрастатели могут существовать и в воздушной среде (аэрофитон ). Из них преобладают зеленые и синезеленые водоросли. При определенных условиях водоросли аэрофитона могут повреждать промышленные и строительные материалы, памятники архитектуры, картины и т.п., если они не защищены ядовитыми покрытиями. Причиной повреждений являются продукты метаболизма обрастателей, в основном – органические кислоты. Водоросли аэрофитона особенно распространены во влажных тропиках, где достаточно тепла, влаги и пыли органического происхождения, являющейся питательной средой для их развития. Биоповреждения от них могут быть значительными.
Эпилиты . К этой группе относятся прикрепленные водоросли. Они заселяют поверхность камней, образуя корковидные покрытия или плоские подушечки, либо обладают особыми органами прикрепления – ризоидами. Интенсивное развитие эпилитов наблюдается в водоемах с твердым дном и быстрым течение воды. Типичными эпилитами являются представителями золотистых водорослей из рода Гидрурус , бурые водоросли из родов Сахарина, Ламинария, Костария и др.
Эпипелиты . Неприкрепленные водоросли, расстилающиеся по дну, связывающие и укрпеляющие субстрат. Часто они представлены свободно ползающими по субстрату диатомовыми, золотистыми, евгленовыми, криптофитовыми, динофитовыми водорослями. Органом прикрепления эпипелитов иногда являются короткие ризоиды, которые не могут глубоко укореняться. Лишь харовые водоросли с их длинными ризоидами хорошо развиваются на илистом дне.
Обычно органами прикрепления эпилитов и эпипелитов служат специальные образования – подошва, ножка, стопа, слизистый тяж или слизистая подушечка, валик и т.п.
Эпифиты/ эпизоиты. Водоросли используют живые организмы как субстрат. Эпизоиты – это водоросли, которые поселяются на животных. На поверхности раковин моллюсков встречаются мелкие зеленые (Эдогониум, Кладофора, Ульва ) и красные (Гелидиум, Пальмария ,) водоросли; на губках – зеленые, синезеленые и диатомовые водоросли. Эпизоиты обитают на ракообразных, коловратках, реже на водных неасекомых или личинках, червях и даже на более крупных животных. К эпизоитам относятся виды зеленых и харовых водорослей из родов Хлорангиелла, Харациохлорис, Коржиковиелла, Хлорангиопсис и др. Большинство эпизоитов не может существовать изолированно от субстрата. На мертвых животных или на их оболочках, сброшенных во время линьки, водоросли обычно гибнут.
Эпифитами называют водоросли, поселяющиеся на растениях. Между растением-субстратом (базифитом) и растением-эпифитом возникают кратковременные связи. Сложное и интересное явление эпифитизма до сих пор слабо изучено. Нередки случаи двойного или даже тройного эпифитизма, когда одни водоросли, поселяющиеся на других, более крупных формах, сами являются субстратом для других, более мелких или микроскопических видов. Иногда для развития эпифитов имеет значение физиологическое состояние растения-субстрата. Количество эпифитов, как правило, возрастает по мере старения водоросли-базифита. Например, наибольшее видовое богатство эпифитных эдогониевых водорослей отмечается на отмерших водных растениях (Манник, Тростник, Осока ).
Эндофиты/ эндозоиты, или эндосимбионты
Эндосимбионты , или внутриклеточные симбионты – водоросли, живущие в тканях или клетках других организмов (беспозвоночных животных или водорослей). Они образуют своеобразную экологическую группу. Внутриклеточные симбионты не теряют способности к фотосинтезу и размножению внутри клеток хозяина. Эндосимбионтами могут быть самые разные водоросли, но наиболее многочисленными являются эндосимбиозы одноклеточных зеленых и желтозеленых водорослей с одноклеточными животными. Водоросли-участники таких симбиозов соответственно называются зоохлореллами и зооксантеллами . Зеленые и желтозеленые водоросли образуют эндосимбиозы и с многоклеточными организмами: губками, гидрами и др. Эндосимбиозы синезеленых водорослей с простейшими называют синцианозами . Нередко в слизи одних видов синезеленых могут поселяться другие виды цианобактерий. Обычно они используют уже готовые органические соединения, образующиеся в изобилии при распаде слизи колонии растения-хозяина, и интенсивно размножаются.
Наиболее распространенными эндофитами являются представители золотистых (виды родов Хромулина, Миксохлорис ) и зеленых (роды Хлорохитриум, Хламидомикса ) водорослей, поселяющиеся в теле ряски и сфагновых мхов. Зеленая водоросль рода Картерия поселяется в эпидермальных клетках ресничного червя Конволюта , один вид рода Хлорелла – в вакуолях простейших, а виды рода Хлорококкум – в клетках криптофитовой водоросли Цианофора .
3.1.3. Водоросли экстремальных водных экосистем
Водоросли горячих источников . Водоросли, которые вегетируют при температуре 35–85 °C, называют термофильными. Нередко высокая температура среды сочетается с повышенным содержанием минеральных солей или органических веществ (сильно загрязненные горячие сточные воды заводов, фабрик, электростанций или атомных станций). Типичные обитатели горячих вод – синезеленые водоросли и, в меньшей степени, диатомовые и зеленые водоросли.
Водоросли снега и льда . Водоросли, развивающиеся на поверхности льда и снега, называют криофильными . Развиваясь в большом количестве, они могут вызывать зеленое, желтое, голубое, красное, коричневое или черное «цветение» снега или льда. Среди криофильных водорослей преобладают зеленые, синезеленые и диатомовые водоросли. Лишь немногие из этих водорослей имеют стадии покоя, большинство лишены каких-либо специальных морфологических приспособлений для перенесения низких температур.
Водоросли соленых водоемов получили название галофильные или галобионты . Вегетируют такие водоросли при повышенной концентрации в воде солей, достигающей 285 г/л, в озерах с преобладанием поваренной соли и 347 г/л в глауберовых озерах. По мере увеличения солености количество видов водорослей уменьшается; очень высокую соленость переносят лишь немногие их них. В пересоленных (гипергалинных) водоемах преобладают одноклеточные подвижные зеленые водоросли – гипергалобы , клетки которых лишены оболочки и окружены плазмалеммой (Астеромонас, Пединомонас) . Они отличаются повышенным содержанием хлорида натрия в протоплазме, высоким внутриклеточным осмотическим давлением, накоплением в клетках каротиноидов и глицерина. В некоторых селенных водоемах такие водоросли могут вызывать красное или зеленое «цветение» воды. Дно гипергалинных водоемов иногда сплошь покрыто синезелеными водорослями; среди них преобладают виды из родов Осциллатория, Спирулина и др. При уменьшении солености наблюдается увеличение видового разнообразия водорослей: помимо синезеленых появляются диатомовые водоросли (виды родов Навикула , Ницшия ).
3.2. Водоросли вневодных местообитаний
Хотя для большинства водорослей основной жизненной средой служит вода, в силу эвритопности этой группы организмов они успешно осваивают и разнообразные вневодные местообитания. При наличии хотя бы периодического увлажнения многие из них развиваются на различных наземных предметах – скалах, коре деревьев, заборах и т. д. Благоприятной средой для обитания водорослей является почва. Кроме того, известны и такие сообщества водорослей-эндолитов, основной жизненной средой которых является окружающий их известковый субстрат.
Сообщества, образуемые водорослями вневодных местообитаний, подразделяют на аэрофильные, эдафофильные и литофильные.
3.2.1. Аэрофильные водоросли
Основной жизненной средой аэрофильных водорослей является окружающий их воздух. Типичные местообитания – поверхность различных внепочвенных твердых субстратов (скалы, камни, кора деревьев, стены домов и т. д.). В зависимости от степени увлажнения их подразделяют на две группы: воздушные и водно-воздушные. Воздушные водоросли обитают в условиях только атмосферного увлажнения и испытывают постоянную смену влажности и высыхания. Водно-воздушные водоросли подвергаются действию постоянного орошения водой (под брызгами водопадов, в зоне прибоя и т. д.).
Условия существования этих водорослей весьма своеобразны и характеризуются, прежде всего, частой сменой двух факторов – влажности и температуры. Водоросли, обитающие в условиях исключительно атмосферного увлажнения, вынуждены часто переходить из состояния переизбыточного увлажнения (например, после ливня) в состояние минимальной влажности в засушливые периоды, когда они высыхают настолько, что могут быть растерты в порошок. Водно-воздушные водоросли живут в условиях относительно постоянного увлажнения, однако и они испытывают значительные колебания этого фактора. Например, водоросли, обитающие на скала, орошаемых брызгами водопадов, в летнее время, когда сток существенно уменьшается, испытывают дефицит влаги. Подвержены аэрофильные сообщества и постоянным колебаниям температуры. Днем они сильно прогреваются, ночью охлаждаются, зимой промерзают. Правда, некоторые аэрофильные водоросли обитают в достаточно постоянных условиях (на стенках оранжерей). Но в целом, к неблагоприятным условиям существования этой группировки приспособились сравнительно немногие водоросли, представленные микроскопическими одноклеточными, колониальными и нитчатыми формами синезеленых и зеленых водорослей и, в значительно меньшей степени, диатомовых водорослей. Аэрофильные формы известны и среди красных водорослей рода Порфиридиум и др.; они встречаются на камнях, старых стенах оранжерей. Число видов, обнаруженных в аэрофильных группировках, приближается к 300. При развитии аэрофильных водорослей в массовом количестве они обычно имеют вид порошкообразных или слизистых налетов, войлокообразных масс, мягких или твердых пленок и корочек.
На коре деревьев обычными поселенцами являются повсеместно распространенные зеленые водоросли из родов Плеврококкус , Хлорелла , Хлорококкус. Синезеленые и диатомовые водоросли находятся на деревьях значительно реже. Есть сведения, что на голосеменных растениях растут преимущественно зеленые водоросли.
Иной систематический состав у группировок водорослей, обитающих на поверхности обнаженных скал. Здесь развиваются диатомовые и некоторые, преимущественно одноклеточные, зеленые водоросли, но наиболее обычны для этих местообитаний представители синезеленых водорослей. Водоросли и сопутствующие им бактерии образуют «горный загар» (наскальные пленки и корочки) на кристаллических породах различных горных массивов. В рухляке, скапливающемся в выемках скал, обитают обычно одноклеточные зеленые водоросли и синезеленые. Особенно обильны разрастания водорослей на поверхности влажных скал. Они образуют пленки и наросты различного цвета. Как правило, здесь обитают виды, снабженные толстыми слизистыми обвертками. В зависимости от интенсивности освещения, слизь бывает окрашена более или менее интенсивно, что определяет цвет разрастаний. Они могут быть ярко-зеленые, золотистые, бурые, лиловые, почти черные в зависимости от образующих их видов. Особенно характерны для орошаемых скал такие представители синезеленых водорослей, как виды родов Глеокапса , Толипотрикс , Спирогира и др. В разрастаниях на влажных скалах можно встретить и диатомовые водоросли из родов Фрустулия , Ахнантес и др.
Таким образом, аэрофильные сообщества водорослей очень разнообразны и возникают, как при вполне благоприятных, так и при экстремальных условиях. Внешние и внутренние приспособления к такому образу жизни разнообразны и сходны с адаптациями почвенных водорослей, особенно развивающихся на поверхности почвы.
3.2.2. Эдафофильные водоросли
Основной жизненной средой эдафофильных водорослей является почва. Типичные местообитания – поверхность и толщина почвенного слоя, оказывающая на бионтов физико-химическое воздействие. В зависимости от местонахождения водорослей и их образа жизни в пределах этого типа различают три группы: наземные водоросли , массово развивающиеся на поверхности почвы в условиях атмосферного увлажнения; водно-наземные водоросли , массово развивающиеся на поверхности почвы, постоянно пропитанной водой; почвенные водоросли , населяющие толщу почвенного слоя.
Почва как биотоп имеет сходство и с водными и с воздушными местообитаниями: в ней есть воздух, но насыщенный водяными парами, что обеспечивает дыхание атмосферным воздухом без угрозы высыхания. Свойством. которое кардинально отличает почву от вышеназванных биотопов, является ее непрозрачность. Этот фактор оказывает решающее воздействие на развитие водорослей. Однако в толще почвы, куда не проникает свет, жизнеспособные водоросли обнаруживаются на глубине до 2 м в целинных и до 2,7 м в пахотных. Это объясняется способностью некоторых водорослей переходить в темноте к гетеротрофному питанию.
В глубоких слоях почвы обнаруживается небольшое число водорослей. Для поддержания своей жизнеспособности почвенные водоросли должны иметь способность к перенесению неустойчивой влажности, резких колебаний температуры и сильной инсоляции. Эти свойства обеспечиваются у них рядом морфологических и физиологических особенностей. Например, отмечено, что почвенные водоросли имеют относительно небольшие размеры в сравнении с соответствующими водными формами тех же видов. С уменьшением размеров клеток возрастает их водоудерживающая способность и устойчивость против засухи. Важную роль в засухоустойчивости почвенных водорослей играет способность к обильному образованию слизи – слизистых колоний, чехлов и оберток, состоящих из гидрофильных полисахаридов. Благодаря наличию слизи, водоросли быстро поглощают воду при увлажнении и запасают ее, замедляя высыхание. Поразительную жизнеспособность демонстрируют почвенные водоросли, хранящиеся в воздушно-сухом состоянии в почвенных образцах. Если такую почву после десятков лет поместить на питательную среду, то можно будет наблюдать развитие водорослей.
Характерной чертой почвенных водорослей является «эфемерность» вегетации – способность быстро переходить из состояния покоя к активной жизнедеятельности и наоборот. Они также способны переносить колебания температуры в очень широком диапазоне: от -200 до +84°C. Почвенные водоросли (преимущественно синезеленые) обладают устойчивостью против ультрафиолетового и радиоактивного излучения.
Подавляющее большинство почвенных водорослей являются микроскопическими формами, но их можно нередко увидеть на поверхности почвы невооруженным глазом. Массовое развитие таких водорослей может вызывать позеленение склонов оврагов и обочин лесных дорог.
По систематическому составу почвенные водоросли довольно разнообразны. Среди них в примерно равных соотношениях представлены синезеленые и зеленые водоросли. Менее разнообразно в почвах представлены желтозеленые и диатомовые водоросли.
3.2.3. Литофильные водоросли
Основной жизненной средой литофильных водорослей служит окружающий их непрозрачный плотный известковый субстрат. Типичные местообитания – в глубине твердых пород определенного химического состава, окруженных воздухом или погруженных в воду. Различают две группы литофильных водорослей: сверлящие водоросли , которые активно внедряются в известковый субстрат; туфообразующие водоросли , откладывающие вокруг своего тела известь и обитающие в периферических слоях отлагаемой ими среды, в пределах доступных для воды и света. По мере нарастания отложений они отмирают.
Контрольные вопросы
1. Охарактеризуйте основные экологические группы водорослей водных местообитаний: фитопланктон и фитобентос.
2. Отличия пресноводного и морского фитоплактона. Представители морского и пресноводного фитопланктона.
3. Морфологические приспособления водорослей к планктонному образу жизни.
4. Сезонные изменения качественных и количественных показателей пресноводного фитопланктона.
5. Отличия пресноводного и морского фитобентоса. Систематический состав морского и пресноводного фитобентоса.
6. Экологические группы фитобентоса по отношению к субстрату (эпилиты, эпипелиты, эпифиты, эндофиты).
7. Что такое обрастание? Какие водоросли могут составлять данную экологическую группу?
8. Аэрофильные водоросли. Приспособления к экстремальным условиям среды. Систематический состав воздушных водорослей.
9. Эдафофильные водоросли. Приспособления к условиям среды. Систематический состав почвенных водорослей.
10. Литофильные водоросли.
4. РОЛЬ ВОДОРОСЛЕЙ В ПРИРОДЕ И ПРАКТИЧЕСКОЕ ЗНАЧЕНИЕ
Роль водорослей в природных экосистемах . В водных биоценозах водоросли играют роль продуцентов. Используя энергию света, они способны синтезировать органические вещества из неорганических. По данным радиоуглеродного анализа, средняя первичная продукция океанов, обусловленная жизнедеятельностью водорослей, составляет 550 кг углерода на 1 гектар в год. Суммарная величина его первичной продукции составляет 550,2 млрд. тонн (в сырой биомассе) в год и, по мнениям ученых, вклад водорослей в общую продукцию органического углерода на нашей планете составляет от 26 до 90%. Важную роль выполняют водоросли в круговороте азота. Они способны использовать как органические (мочевина, аминокислоты, амиды), так и неорганические (ионы аммония и нитратов) источники азота. Уникальную группу представляют собой синезеленые водоросли, которые способны фиксировать газообразный азот, переводя его в соединения, доступные для других растений.
Водоросли – продуценты кислорода . Водоросли в процессе своей жизнедеятельности выделяют кислород, необходимый для дыхания водных организмов. В водной среде (особенно в морях и океанах) водоросли являются практически единственными продуцентами свободного кислорода. Кроме того, они играют большую роль в общем балансе кислорода на Земле, так как океаны служат главным регулятором баланса кислорода атмосферы Земли.
Водоросли – среда для других водных организмов . Образуя подводные леса, водоросли-макрофиты создают высокопродуктивные экосистемы, которые дают пищу, пристанище и защиту многим другим живым организмам. Установлено, что столб воды объемом 5 л с одним экземпляром бурой водоросли Цистозиры содержит до 60 тыс. особей различных беспозвоночных животных, включая моллюсков, клещей и ракообразных.
Водоросли – пионеры растительности . Наземные водоросли могут поселяться на голых скалах, песке и других бесплодных местах. После их отмирания образуется первый слой будущей почвы. Почвенные водоросли участвуют в процессах формирования структуры и плодородия почв.
Водоросли как геологический фактор. Развитие водорослей в прошедшие геологические эпохи привело к образованию ряда горных пород. Вместе с животными водоросли принимали участие в образовании рифов в океанах. Поселяясь ближе к поверхности воды, они образовывали гребни этих рифов. Рифовые постройки красных водорослей известны в Крыму как вершины Яйлы и др. Синезеленые водоросли участвовали в образовании известняков-строматолитов, харовые водоросли – в образовании известняков-хароцитов (подобные отложения обнаружены в Туве). В образовании меловых пород принимают участие коколитофориды (меловые породы на 95% состоят из остатков панцирей этих водорослей). Массовое скопление панцирей диатомовых водорослей привело к образованию диатомита (горной муки), крупные залежи которого обнаружены в Приморском крае, на Урале и Сахалине. Водоросли явились исходным материалом для жидких и твердых нефтеподобных соединений – сапропелей, горячих сланцев, углей.
Активная деятельность водорослей в образовании горных пород отмечена в некоторых регионах и в настоящее время. Они поглощают карбонат кальция и образуют минерализованные продукты. Эти процессы особенно активны в тропических водах с высокими температурами и низким парциальным давлением.
В разрушении горных пород наибольшее значение имеют сверлящие водоросли. Они медленно и упорно разрыхляют известковые субстраты, делая их доступными для выветривания, крошения и размывания.
Симбиотические отношения с другими организмами . Водоросли образуют несколько важных симбиозов. Во-первых, с грибами образуют лишайники, во-вторых, как зооксанты обитают совместно с некоторыми беспозвоночными животными, такими, как губки, асцидии, рифовые кораллы. Ряд цианофит образует ассоциации с высшими растениями.
Водоросли имеют большое практическое значение в повседневной жизни и хозяйственной деятельности человека, принося как пользу, так и вред. Крупные, главным образом морские, водоросли были известны с древнейших времен и издавна использовались в хозяйстве человека.
Водоросли как пищевой продукт . Человек употребляет в пищу преимущественно морские водоросли, особенно широко используют их жители юго-восточной Азии и островов Тихого океана. В Китае использование водорослей в пищевом рационе известно с 9 века до н. э. Среди водорослей-макрофитов (многоклеточных зеленых, бурых и красных) нет ядовитых видов, потому что они не содержат алкалоидов – веществ с наркотическим и отравляющим эффектом. В пищу используют около 160 видов различных водорослей. По своим пищевым качествам водоросли не уступают многим сельскохозяйственным культурам. В них содержится большой процент белка, углеводов и жиров. Водоросли – прекрасный источник витаминов C, A, D, группы B, рибофлавина, пантотеновой и фолиевой кислот, микроэлементов.
Из микроскопических водорослей в пищу используют синезеленые наземные виды рода Носток, которые служат пищей в Китае и Южной Америке. В Японии употребляют ячменный хлеб «тенгу» – это мощные пласты плотной студенистой массы на склонах некоторых вулканов, состоящей из синезеленых водорослей из родов Глеокапса, Геотеке, Микроцистис с примесью бактерий. Спирулину использовали ацтеки еще в 16 веке, приготовляя из высушенной водоросли лепешки, а население района озера Чад в Северной Америке до сих пор готовят из этой водоросли продукт под названием дихе. Спирулина содержит большое количество белка, и ее широко культивируют в ряде стран.
Водоросли как удобрение . Водоросли содержат достаточное количество органических и минеральных веществ, поэтому издавна использовались в качестве удобрений. Преимущества таких удобрений в том, что они не содержат семена сорняков и споры фитопатогенных грибов, а по содержанию калия превосходят почти все типы применяемых удобрений. Азотфиксирующие синезеленые водоросли широко используют на рисовых полях вместо азотных удобрений. Показано, что удобрения из водорослей могут повышать всхожесть семян, урожайность, устойчивость к болезням.
Лечебные свойства водорослей . Водоросли широко применяются в народной медицине как глистогонное средство и для лечения ряда заболеваний, таких как зоб, нервные расстройства, склероз, ревматизм, рахит и др. Показано, что экстракты водорослей многих видов содержат антибиотические вещества, могут снижать давление. Экстракты из Саргассума, Ламинарии и Сахарины в экспериментах на мышах подавляли рост саркомы и лейкемических клеток. В США и Японии из них получены препараты, способствующие выведению из организма радионуклидов. Эффективность таких сорбентов достигает 90–95%.
Водоросли как источник промышленного сырья . Еще с прошлого века водоросли использовались для получения соды и йода. В настоящее время из водорослей получают альгиновую кислоту и ее соли – альгинаты, а также каррагинаны и агар.
Из бурых водорослей получают спирт маннит – необходимое сырье для фармакологической и пищевой промышленности при изготовлении лекарств и продуктов питания для диабетиков.
Отрицательная роль водорослей . Ряд водорослей (синезеленые, динофитовые, золотистые, зеленые) выделяют токсины, которые могут вызывать различные заболевания животных, растений и человека, причем некоторые из них могут оказаться смертельными. Среди динофитовых водорослей, вызывающих "красные приливы" на огромных морских акваториях, токсическими являются виды родов Гимнодиниум, Ноктилюка, Амфидиниум и др. Наибольшее число токсических видов выявлено среди синезеленых водорослей. По своему действию токсины синезеленых водорослей в несколько раз превосходят такие яды, как кураре и ботулин. Токсичность водорослей проявляется в массовой гибели гидробионтов, водоплавающей птицы, в отравлениях и других заболеваниях людей, возникающих при вдыхании, использовании воды, употреблении в пищу моллюсков, рыбы и т.д.
При сильном развитии – «цветении водоемов» некоторые водоросли (золотистые, желтозеленые, синезеление) могут придавать воде неприятный запах и вкус, делая воду непригодной для питья.
Чрезмерный рост водорослей может препятствовать прохождению воды через фильтры водозаборных сооружений. Известно, что обрастание водорослями судов заметно увеличивает эксплуатационные расходы. Макрофиты могут способствовать коррозии материалов на нефтяных платформах и других подводных морских сооруженях.
Проблема обрастания – едва ли не самая древняя при освоении океана. Любой предмет, оказавшийся в контакте с морской средой, вскоре покрывается массой прикрепившихся к нему организмов: животных и водорослей. Общая площадь погруженных в море субстратов составляет около 20% от площади поверхности верхних отделов шельфа. Общая биомасса обрастания исчисляется миллионами тонн, ущерб от него – миллиардами долларов (Звягинцев, 2005). В биологическом аспекте – это естественный процесс, составляющий неотъемлемую часть жизни гидросферы. В то же время явление обрастания подсказало человеку идею выращивания в морских хохяйствах в промышленных масштабах ряда ценных видов моллюсков (Устрицы, Мидии, Гребешка, Жемчужницы ) и водорослей (Сахарины, Порфиры, Грацилярии, Эухемы и др.). Водоросли являются пионерными организмами обрастания. Микроводоросли вместе с бактериями образуют на поверхности искусственных субстратов, внесенных в воду, первичную микропленку, служащую субстратом для оседания других гидробионтов. Макроводоросли вместе с ракообразными, моллюсками, гидроидами и другими животными нередко формируют первоначальные стадии многолетних сообществ обрастания.
Контрольные вопросы
1. Роль водорослей в повышении плодородия земель.
2. Роль водорослей в водных экосистемах.
3. Роль водорослей в наземных экосистемах.
4. Значение водорослей в геологических процессах.
5. Пищевая и биологическая ценность водорослей. Какие водоросли можно употреблять в пищу?
6. Лечебные свойства водорослей.
7. Почему нежелательно разрастание золотистых и желтозеленых водорослей в водохранилищах? Что такое «цветение» водоемов?
8. Водоросли, которые вызывают отравления животных и человека.
9. Явление обрастания. Роль водорослей в сообществах обрастания.
5. СОВРЕМЕННАЯ СИСТЕМАТИКА ВОДОРОСЛЕЙ
Классификация живых организмов занимала умы людей со времен Аристотеля. Шведский ботаник Карл Линней был первым, кто в 18 веке применил название Algae к одной из групп растений и положил начало фикологии (от греч. phycos – водоросль и logos – учение) как науке. Среди водорослей Линней различал всего четыре рода: Хара, Фукус, Ульва и Конферва. В 19 веке было описано большинство (несколько тысяч) современных родов водорослей. Большое количество новых родов вызвало необходимость группирования их в таксоны более высокого ранга. Начальные попытки классификации базировались исключительно на внешних признаках таллома. Первым, кто предложил окраску таллома водорослей в качестве фундаментального признака для установления больших таксономических групп, или мегатаксонов, был английский ученый В. Гарвей (Harvey,1836). Он выделил крупные ряды: Chlorospermeae – зеленые водоросли, Melanospermeae – бурые водоросли и Rhodospermeae – красные водоросли. Позднее они были переименованы соответственно в Chlorophyceae, Phaeophyceae и Rhodophyceae.
Основы современной систематики водорослей были заложены в первой половине 20 века чешским ученым А. Пашером. Он установил 10 классов водорослей: Синезеленые, Красные, Зеленые, Золотистые, Желтозеленые, Диатомовые, Бурые, Динофитовые, Криптофитовые и Евгленовые. Каждый класс характеризуется специфическим набором пигментов, запасных продуктов и строением жгутиков. Эти постоянные различия между крупными таксонами побудили рассматривать их как самостоятельные филогенетические группы, не связанные родством, и отказаться от понятия водоросли – Algae как определенной таксономической единицы.
Таким образом, слово "водоросли" фактически является не систематическим, а экологическим понятием и дословно означает – "то, что растет в воде". Водоросли – низшие растения, которые содержат в подавляющем большинстве хлорофилл, способны к фототрофному питанию и живут преимущественно в воде. Все водоросли, кроме харофитовых, в отличие от высших растений, не имеют многоклеточных половых органов с покровами из стерильных клеток.
Современные системы различаются главным образом по числу и объему мегатаксонов – отделов и царств. Число отделов варьирует от 4 до 10-12. В русской фикологической литературе почти каждому из указанных выше классов соответствует отдел. В зарубежной литературе наблюдается тенденция к укрупнению отделов и, соответственно, уменьшению их количества.
Наиболее распространена в классификационных построениях схема Паркера (Parker, 1982). Она признает разделение между прокариотными и эукариотными формами. У прокариотных форм в клетках отсутствуют окруженные мембранами органеллы. К прокариотам относят Bacteria и Cyanophyta (Cyanobacteria). Эукариотные формы включают все другие водоросли и растения. Подразделение водорослей долгое время является предметом споров. Гарвей (Harvey, 1836) разделял водоросли прежде всего по окраске. Хотя отделов в настоящее время признается намного больше, составу пигментов, биохимическим и структурным особенностям строения клетки придается огромное значение. П. Силва (Silva, 1982) различает 16 основных классов. Классы различаются пигментацией, запасными продуктами, особенностями клеточной стенки и ультраструктурой жгутиков, ядра, хлоропластов, пиреноидов и глазков.
Новая информация по ультраструктуре водорослей, полученная в последние десятилетия методами электронной микроскопии, генетики и молекулярной биологии, позволяет исследовать мельчайшие детали строения клетки. "Взрывы" информации периодически побуждают ученых пересматривать устоявшиеся традиционные представления о систематике водорослей. Постоянное поступление новой информации стимулирует новые подходы к классификации, и каждая предлагаемая схема неизбежно остается приблизительной. По современным данным организмы, традиционно рассматриваемые среди низших растений, выходят за рамки Царства Растений. Они включены в состав большого числа самостоятельно эволюционирующих групп. В таблице приведены в разных трактовках мегатаксоны, в состав которых входят водоросли. Как видно, различные таксоны водорослей могут находиться в разных филах; одни и те же филы могут объединять разные эколого-трофические группы организмов (таблица).
Более 100 лет назад К.А. Тимирязев прозорливо заметил, что «нет ни растения, ни животного, а есть один нераздельный органический мир. Растение и животное – только средние величины, только типические представления, которые мы слагаем, отвлекаясь от известных признаков организмов, придавая исключительное значение одним, пренебрегая другими». Сейчас мы не можем не восхититься его потрясающей биологической интуицией.
Современная система водорослей, изложенная в данном учебном пособии, включает 9 отделов: Синезеленые, Красные, Диатомовые, Гетероконтные, Гаптофитовые, Криптофитовые, Динофитовые, Зеленые, Харофитовые и Евгленовые водоросли. Сходство состава пигментов, строения фотосинтетического аппарата и жгутиков послужило основанием для объединения классов водорослей, имеющих золотисто-бурую окраску, в одну большую группу – Гетероконтные, или Разножгутиковые водоросли (Охрофитовые).
Мегасистема относимых к низшим растениям организмов
|
Империя |
Царство |
Отдел (Тип) |
Трофогруппа |
|
Eubacteria/ Prokaryota |
Cyanobacteria/Bacteria |
Cyanophyta/ Cyanobacteria |
Водоросли |
|
Excavates/ Eukaryota |
Euglenobiontes/ Protozoa |
Euglenophyta/ Euglenozoa Acrasiomycota |
Водоросли Миксомицеты |
|
Rhizaria/ Eukaryota |
Cercozoa/ Plantae |
Chlorarachniophyta Plasmodiophoromy-cota |
Водоросли Миксомицеты |
|
Rhizaria/ Eukaryota |
Myxogasteromycota Dictyosteliomycota |
Миксомицеты Миксомицеты |
|
|
Choromalveola-tes/ Eukaryota |
Straminopilae/ Chromista/ He-terokontobiontes |
Labyrinthulomycota -Oomycota Heterokontophyta |
Миксомицеты Грибы Водоросли |
|
Choromalveola-tes/ Eukaryota |
Haptophytes/ Chromista |
Prymnesiophyta/ Haptophyta |
Водоросли |
|
Choromalveola-tes/ Eukaryota |
Cryptophytes/ Chromista |
Водоросли |
|
|
Choromalveola-tes/ Eukaryota |
Alveolates/ Protozoa |
Dinophyta/ Myzozoa |
Водоросли |
|
Plantae/ Eukaryota |
Glaucophytes/ Plantae |
Glaucocystophyta/ Glaucophyta |
Водоросли |
|
Plantae/ Eukaryota |
Rhodobiontes/ Plantae |
Cyanidiophyta Rhodophyta |
Водоросли Водоросли |
|
Plantae/ Eukaryota |
Chlorobiontes/ Plantae |
Chlorophyta Charophyta |
Водоросли Водоросли |
ВОДОРОСЛИ
Общая характеристика
Водоросли представляют собой большую группу низших расте ний — от микроскопически малых, одноклеточных организмов до многоклеточных гигантов. Термин «водоросли» не является систе матической единицей. Под названием «водоросли» объединяется несколько систематических отделов низших растений, имеющих различное происхождение, характеризующихся сходным образом жизни и автотрофным типом питания. Как показывает название, для водорослей обычно характерен водный образ жизни.
Тело водорослей, как и других низших растений, представляет собой слоевище — таллом, не расчлененное на корень, стебель и лист, и у большинства видов имеет простое анатомическое строение. Водоросли содержат в своих клетках хлорофилл, способны фотосинтезировать. Строение клетки, тела, способы размножения различны у отдельных предста вителей этой группы растительных отделов.
Клетки большинства водорослей имеют целлюлозную оболочку; встречаются клетки, содержимое которых окружено лишь тонкой пограничной мембраной. Клеточные оболочки водорослей разнообразны по строению и химическому составу. Основой оболочки является белково - углеводный комплекс. Оболочке свойственны неоднородность, слоистость. Слои отличаются один от другого по толщине, плотности и химическому составу. Нередко оболочки пропитываются органическими соединениями (лигнином и кутином).
В оболочке имеются особые отверстия — поры. Помимо пор, оболочки клеток многих водорослей снабжены различного рода выро стами — щетинками, шипиками и чешуйками.
В протопласте клетки различают ядро и цитоплазму. У боль шинства водорослей в клетке имеется только одно ядро, но иногда их бывает 2, 3 и больше. Клетки сине-зеленых водорослей лишены оформленного ядра. Форма, размеры и местоположение ядра в клетке у разных водорослей сильно варьируют. В ядре у водорослей име ются те же структуры, что и в ядрах других растений: оболочка, ядерный сок, ядрышко, включения хроматина.
Цитоплазма состоит из основного вещества (стромы) и погруженных в него телец (органелл). Отличительной особенностью кле ток водорослей является слабое развитие эндоплазматическои сети.
В специфических органеллах — хлоропластах (названных хро матофорами) находятся тельца, богатые белковыми веществами, которые называются пиреноидами. Пиреноид окружен обкладкой в виде кольца или отдельных пластинок обычно крахмальной природы. В хлоропластах содержится зеленый пигмент хлорофилл, существующий в нескольких формах .
Кроме хлорофилла, водоросли содержат и другие пигменты, ко торые часто своим присутствием маскируют зеленую окраску водо рослей. Наиболее характерными и часто встречающимися пигмен тами водорослей, кроме хлорофилла (зеленый), каротина (желто- оранжевый) и ксантофилла (желтый), являются фикоциан, фико- эритрин и фукоксантин. Пигмент фикоциан, растворимый в воде, окрашивает водоросли в синий цвет (сине-зеленые водоросли); фикоэритрин, тоже растворимый в воде, придает красную окраску (красные водоросли); фукоксантин обусловливает бурую окраску (бурые водоросли).
Обладая пигментами, водоросли придают субстрату, на котором они поселяются, различную окраску. В Антарктиде, например, советскими учеными были открыты 3 озера с различной окраской воды: синей, зеленой и красной. Цвет воды этих озер был обусловлен наличием в них очень мелких водорослей соответствующей окраски. Ледяные берега Гренландии во многих местах с наступлением весен него потепления окрашиваются в красный (кровяной) цвет, что объясняется массовым размножением водорослей с красной окрас- кой.
Название Красного моря обязано произрастанию в большом количестве в этом море водорослей, имеющих красноватый оттенок. Различная окраска водорослей имеет приспособительное значе ние. Дневной свет к водорослям, погруженным глубоко в воду, доходит всегда в измененном составе. Вода сравнительно хорошо пропускает синие и зеленые и сильно поглощает красные и желтые лучи. На больших глубинах задержание зеленых лучей одним хлорофиллом происходит плохо, ему на помощь приходит красный пигмент, который легко поглощает зеленые лучи. Поэтому красные, или багряные, водоросли распространены на большой глубине морей.
По форме и размерам водоросли очень разнообразны. Встре- чаются микроскопически малые и достигающие огромных размеров (до 50 м и более) одноклеточные и многоклеточные формы. Переходную ступень между ними занимают колониальные водоросли. Колонии состоят из нескольких неплотно соединенных однородных клеток. С увеличением размеров водорослей происходит и некото рая дифференциация их тела. Например, у бурых водорослей, дости гающих крупных размеров, таллом сильно расчленен. Такие водоросли прикрепляются к субстрату тонкими бесцветными нитями, которые называются ризоидами.
Существуют одноклеточные водоросли, у которых тело достигает больших размеров 0,5 м и больше. Они имеют большое количество ядер и хроматофоров. Внешне тело такой водоросли имеет расчле нение, но внутри перегородок нет, т. е. является одноклеточной. Примером такой гигантской одноклеточной водоросли может слу жить морская каулерпа.
Размножение водорослей может быть вегетативное, бесполое и половое. Вегетативное размножение осуществляется частями тела водорослей. Бесполое размножение происходит за счет образования зооспор, реже спор. Половой процесс у различных типов водорослей представлен: изогамией, гетерогамией, оогамией и автогамией.
У некоторых более высокоорганизованных водорослей (красных, бурых) наблюдается чередование полового и бесполого поколений.
Водоросли характеризуются большим разнообразием. Общее количество их видов свыше 20 тыс. Классификация водорослей очень сложна, и в настоящее время она еще не закончена. Виды объединяются в более крупные систематические единицы — роды, семей ства, порядки, классы, отделы (типы).
Все разнообразие водорослей объединяют обычно в 6...10 крупных отделов (типов), представители которых отличаются по строе нию, способу размножения, происхождению, но чаще всего пред ставители этих отделов различаются между собой по окраске.
В данном учебнике рассмотрено 6 отделов водорослей: сине- зеленые, разножгутиковые, диатомовые, зеленые, бурые и крас ные.
Водоросли имеют очень древнее происхождение, в котором много еще неясного. Нет единой точки зрения на родственные отношения между отделами водорослей. Некоторые представители их произошли, по-видимому, от более простых организмов типа жгутиковых, а сами водоросли являются предками некоторых более раз витых растений (грибы, мхи).
Представители современного разнообразного мира водорослей имеют различную древность по своему происхождению, они появились в разные геологические эры. Наиболее древними считаются сине-зеленые водоросли.
Эволюция водорослей шла от подвижных форм к неподвижным. Более примитивными и древними группами водорослей считаются те, которые проводят жизнь в подвижном состоянии; более органи зованным водорослям свойствен неподвижный образ жизни. Путь эволюции водорослей шел от простейших — одноклеточных микро скопически малых форм к многоклеточным сложным формам.
Как уже говорилось, в подавляющем большинстве водоросли живут в водной среде. Одни из них растут в соленой воде океанов и морей, другие — в реках, прудах и пресноводных озерах. Боль шое влияние на распространение водорослей по глубинам водоемов оказывает свет, поэтому в поверхностных слоях воды количество водорослей всегда больше. Морские водоросли красные и бурые часто образуют огромные подводные заросли, которые занимают десятки километров. В зависимости от местообитания водоросли делятся на 2 большие группы: бентосные и планктонные.
Бентосные, или донные, водоросли обитают, прикрепившись ко дну или подводным предметам, и образуют заросли главным образом в прибрежной полосе. Планктонные водоросли живут в воде во взвешенном состоянии, не прикрепляясь ко дну, они могут перено ситься движением воды.
Водоросли живут не только в воде, они встречаются на поверх ности почвы, в почве, на деревьях, на сваях, но всегда на увлаж ненных местах. Часто, особенно ранней весной, почвы «зацветают», или «зеленеют», что объясняется развитием колоссального коли чества микроскопически малых водорослей. «Зацветание» почвы от водорослей происходит в различных районах Советского Союза - в северных, степных и даже пустынных. Разрастаясь в большом количестве, они создают условия для развития бактерий и других микроорганизмов.
Водоросли имеют большое значение в природе и широко исполь зуются человеком.
Как автотрофные растения они перерабатывают огромное коли чество минеральных веществ и углекислого газа в органическую массу. Водоросли создают огромные запасы пищи для животного мира морей, океана и пресноводных водоемов. Так, 1 га зарослей водорослей может дать урожай, равный 100 т сырой или 10 т сухой массы.
Водоросли вырабатывают колоссальное количество кислорода.
Развитие рыбного хозяйства неразрывно связано с водорослями. Поглощая много углекислого газа и выделяя кислород, водоросли очищают водоемы.
В приморских странах (Англия, Франция, Норвегия, Ирландия и др.), особенно в Японии, водоросли широко употребляются в пищу (морская капуста и др.) и на корм скоту (в сыром, сухом и силосо ванном виде). Некоторые водоросли используют для удобрения полей.
Многие водоросли накапливают большое количество йода и брома. Йода содержится в золе водорослей около 0,2% от сухой массы. Часто водоросли являются основным источником йода. Вываривая, красные и бурые водоросли, получают ценное вещество — агар-агар, который употребляется в микробиологии как питательная среда для выращивания микроорганизмов, а также в кондитерской промышленности при изготовлении мармелада и пр.
В древние геологические эпохи диатомовые водоросли вместе с кремнеземом образовали осадочную породу — трепел, который употребляется в производстве динамита, кирпича, для полировки предметов и др.
В пресных водоемах водоросли участвуют в образовании сапропе ля, или органического ила. Сапропель содержит большое количест во органических веществ и часто используется для грязелечения. Сапропель, который содержит большое количество солей кальц ия, фосфора, железа, может быть использован на корм сельско хозяйственным животным.
Водоросли, особенно морские, могут принести и вред, когда в большом количестве покрывают подводные части корабля. При массовом отмирании водорослей происходит порча воды и, как следствие, гибель рыбы.
Отдел сине-зеленые водоросли
Сине-зеленые водоросли, или цианеи, являются наиболее примитивными и наиболее древними по происхождению организмами. На основании палеоботанических данных установ лено, что современные сине-зеленые водоросли мало чем отличаются от своих ископаемых предков. В большинстве случаев они представлены одноклеточными формами, хотя имеются и многоклеточ ные формы, собранные в колонии.
Как показывает само название этого отдела водорослей, для них характерна сине-зеленая окраска различных оттенков в зависи мости от соотношения пигментов — хлорофилла, каротина, фико- цианина и фикоэритрина.
Оболочка клетки сине-зеленых водорослей состоит из пектино- вых веществ и покрыта снаружи слизистым веществом. В их клет- ках отсутствуют морфологически обособленное ядро и хроматофоры, цитоплазма пропитана пигментами, и поэтому называется хромато плазмой.
В процессе ассимиляции вместо обычного растительного крах- мала образуется углевод гликоген (животный крахмал).
Размножаются сине-зеленые водоросли очень быстро, простым делением клеток пополам. Половой процесс размножения у них отсутствует.
По строению и характеру деления клетки сине-зеленых водо- рослей сходны с бактериями. Они, так же как и бактерии, не имеют ясно выраженного ядра, жгутиковые стадии развития у них отсут ствуют. У некоторых сине-зеленых водорослей (нитчатых) размно жение происходит участками, на которые распадаются эти водоросли. Такие участки называются гормогониями. При неблаго приятных условиях из обычных клеток образуются споры, которые покрываются утолщенной оболочкой. Она предохраняет содержимое от неблагоприятных условий, благодаря чему водоросль сохраняет жизнеспособность в течение длительного времени. При наступлении благоприятных условий споры прорастают и дают начало новой клетке.
Обитают сине-зеленые водоросли преимущественно в пресных водах — прудах, озерах, реках, но встречаются также в морях, на поверхности почвы, на скалах. Сине-зеленые водоросли могут жить как при низких температурах на снегу и льду, так и при высо ких температурах (до 80 °С) в горячих ключах. После отмирания клеток масса водорослей в виде хлопьев грязно-зеленоватого цвета всплывает на поверхность воды.
Древнее происхождение, недифференцированное строение кле ток, отсутствие оформленного ядра и полового процесса, размноже ние простым делением клетки, способность образовывать споры — все эти особенности указывают на примитивность сине-зеленых водорослей. По своему упрощен ному строению они значительно отличаются от других водоросл ей и ближе всего стоят к бактериям.
Сине-зеленые водоросли отличаются хорошей приспособ ляемостью к различным услови ям окружающей среды, что и способствовало сохранению их до наших дней без особых из менений.
Отдел сине-зеленые водоро сли объединяет около 1400 ви дов. Представителями этого отдела могут служить такие водо росли, как хроококк, осциллатория, носток и др.
Хроококк — одноклеточная водоросль шаро видной формы, иногда эти водоросли образуют как бы колонии. Часто такие водоросли собраны в группы по 2...4, разделенные тонкой перегородкой и окруженные общим довольно толстым слизи стым слоем.
Хроококк широко распространен на болотах среди водных рас тений, между кочками и среди тины.
Осциллатория — нитчатая сине-зеленая водоросль, распространенная в водоемах со стоячей водой, часто образует темно-зеленую пленку на поверхности воды или на илистом дне. Клетки этой водоросли имеют цилиндрическую форму, плотно соединены в одну нить.
Носток — нитчатая сине-зеленая водоросль, нити или цепочки ее соединены в колонии, часто шаровидной формы, размером с плод сливы. Снаружи эти колонии покрыты студенистой массой. Обитает носток по берегам прудов и озер, на влажной почве и на дне водоемов.
Некоторые сине-зеленые водоросли вместе с грибами образуют различные виды лишайников.
Отдел разножгутнковые водоросли
Разножгутиковые, или желто-зеленые, водоросли характеризуются тем, что их зооспоры имеют 2 неравных жгутика, причем короткий жгутик гладкий, а длинный — перистый. Хроматофоры желто-зеленого цвета, дисковидной формы. Одноклеточные, колониальные, нитчатые и неклеточные организмы. Примером может служить ботридиум.
Ботридиум — наиболее характерный пред ставитель разножгутиковых водорослей. Таллом представляет собой зеленый пузырек 1...2 мм в диаметре, с нижней стороны пузырька расположены бесцветные ветвистые выросты — ризоиды, которыми водоросль внедряется в почву. Это одноклеточная многоядериая водоросль. Середина пузырька заполнена клеточным соком, цитоплазма расположена постенно. Хроматофоры содержат много каро тиноидов, поэтому ботридиум имеет желтовато-зеленый цвет. Пиреноиды отсутствуют.
Размножается главным образом зооспорами, которые образуются в колоссальном количестве в середине пузырька. Обитает на сырой земле, по краям луж, образует на почве налеты темно-зеленого цвета.
Отдел диатомовые водоросли
Диатомовые, диатомеи, или кремнеземные, или бациллариевые, водоросли — чрезвы чайно разнообразные, микроскопически малые, в большинстве случаев одноклеточные организмы. Этот отдел водорослей характе ризуется своеобразным строением клеток. Клетка покрыта сплошной оболочкой в виде пектиновой, студнеобразной пленки, снаружи она охвачена кремнистым панцирем, который состоит из двух само стоятельных половинок, так называемых створок. Одна из этих ство рок покрывает другую, как крышка коробочку. Вдоль половинок панциря с обеих сторон имеется щелевидное отверстие. Через это отверстие цитоплазма клетки сообщается с внешней средой. Створ ки обладают исключительной прочностью. Они не переваривают ся животными и птицами, не разрушаются даже при накалива нии на огне. Существует свыше 5000 видов. Отличительными видо выми признаками служат форма клеток и различные утолщения на оболочках в виде рубчиков, сеток и пр.
Форма клеток отдельных видов диатомовых водорослей бывает удлиненно-квадратная, эллиптическая, круглая, звездчатая, в виде лент, спиралей и др. Клетки содержат цитоплазму, ядро и один или несколько хроматофоров. Кроме хлорофилла и фукоксантина, Хроматофоры содержат и другие пигменты желто-бурого цвета, поэтому Хроматофоры имеют желтый цвет. Крахмал в клетках диатомовых водорослей отсутствует, запасные вещества представ лены маслом.
Размножаются главным образом прямым делением, которое про ходит у них своеобразно. При делении каждая дочерняя клетка получает ядро, один хроматофор и только одну из створок оболочки, вторая створка образуется заново. Кроме прямого деления, диато мовые водоросли размножаются половым путем, когда сливаются две клетки, предварительно сбросившие оболочки.
Обитают диатомовые водоросли в морских и пресных водах, часто они являются основной составной частью планктона и служат ценной пищей для животных. Створки отмерших клеток водорослей опускаются на дно и постепенно образуют огромные отложения, известные, под названием "горная мука, диатомит, трепел.
Большие отложения диатомовых водорослей в виде диатомового и ла сосредоточены в приполярных областях океанов, около Аляски, Алеутских островов, в Охотском и Беринговом морях. Имеются они и в Балтийском море.
К диатомовым относятся пресноводная водоросль пиниуля рия, фрагилярия и табелля рия; эти водоросли образуют колонии в виде лент или цепочек; навикула растет кустовндной коло нией.
Отдел зеленые водоросли
Зеленые водоросли являются одним из наиболее разнообразных отделов водорослей", он объединяет около 5000 видов. Для представителей, этого отдела характерна зеленая окраска, которая обусловливается хлорофиллом и не маскируется какими- либо-другими пигментами. Зеленые водоросли представлены одно-клеточными, многоклеточными, колониальными формами. Зеленые водоросли имеют чаще всего нитчатое строение, нити состоят из одного ряда клеток. Клетка имеет целлюлозную оболочку, цито плазму, ядро и хроматофоры. Размножение происходит бесполым, вегетативным и половым путем зеленых водорослей обитает в воде, но некоторые из них живут и на суше, на тем.
Большинство представит илей живут на деревьях, на кам нях в сырых, затененных местах. В прудах и реках образуют тину.
Отдел зеленые водоросли делится на несколько классов, из кото рых рассмотрим равножгутиковые, или собственно зеленые, водоросли сцеплянки, или конъюгаты , и харовые, или лучицы Класс равножгутиковые Среди зеленых водо рослей этот класс наиболее обширный. Представители разнообразны по внешнему виду и внутреннему строению. Для них характерно наличие двух одинаковых жгутиков Этот класс объединяет одно клеточные и колониальные, подвижные и неподвижные формы. Класс разделяют на 8 порядков. Рассмотрим представителей 4 по рядков.
Порядок вольвоксовые . Наиболее характерными представителями порядка вольвоксоиые являются хламидомонада и вольвокс.
Хламидомонада — одноклеточная, подвижная водоросль. В большом количестве обитает в пресных мелководных водоемах — лужах, прудах, канавах вдоль дорог. При обильном размножении окрашивает воду в зеленый цвет. Представляет собой микроскопически малую клетку овальной или округлой формы. На одном (переднем) конце клетка вытянута в виде носика, на этом же конце имеются 2 равных жгутика, которые способствуют передвижению водоросли. Вся полость клетки запол нена цитоплазмой, в которой находится ядро, ближе к носику
Вольво кс, или волчок , является характерным примером колониальных форм микроскопических водорослей. Колония этой водоросли заметна невооруженным глазом, она достигает величины булавочной головки и имеет форму шара. Такая шарообразная колония вольвокса состоит из огромного количества клеток (до 50 тыс.), расположенных в один слой по периферии шара. Каждая клетка несет по 2 жгутика. Все жгутики расположены по периферии и способствуют передвижению всей колонии. Движение жгутиков всех клеток всегда согласованно. Полость шара запол нена жидкой слизью. Размножается вольвокс вегетативным и половым путем. При вегетативном размножении внутри материнской колонии (шара) образуется 8...15 дочерних колоний (шаров). При созревании дочерних колоний стенки взрослого шара разрываются и молодые колонии выходят наружу, после чего материнская коло ния погибает.
Половое размножение — оогамия — происходит только в наибо лее крупных клетках. При этом в клетках колонии вольвокса об разуются голые двухжгутиковые гаметы, сливаются лишь гаметы из разных особей водоросли.
Существует много видов вольвоксов. Все они распространены главным образом в непроточных хорошо прогреваемых пресно водных водоемах со стоячей водой — прудах, озерах, реже в реках
Хлорелла микроскопическая одноклеточная водоросль шаровидной формы. Для хлореллы характерны быстрое размножение и очень активный процесс фотосинтеза. Благодаря наличию большого количества пластид хлорелла отличается эффек тивным использованием солнечной энергии, обычные культурные растения потребляют всего 0,1% солнечной энергии при биохими ческих превращениях, а хлорелла — 2,5%, т. е. в 25 раз больше. Другое положительное качество хлореллы — очень быстрое размно жение.
Улотрикс нитчатая неветвящаяся водоросль, состоящая из одного ряда клеток. Нижняя бесцветная клетка нити имеет своеобразную удлиненную форму Этой клеткой водоросль прикрепляется к подводным предметам.
Нить улотрикса удлиняется за счет поперечного деления клеток и может неограниченно нарастать в длину. Клетки однородные, короткие, каждая из них цитоплазму, ядро, хроматоформ с пиреноидами. Размножается улотрикс главным образом бесполым путем, образуя 4-жгутиковые зооспоры. Эти зооспоры некоторое время двигаются, а затем оседают на какой-либо подводный предмет и прорастают в новую нить Половой процесс — изогамия. Причем мужские и женские гаметы внешне не различимы, но физио логически они разные и выходят из разных нитей, поэтому разные гаметы обозначаются знаком + и знаком —.
Улотрикс обитает в пресных водоемах, где им обрастают под водные предметы (камни, сваи), которые приобретают ярко-зеленый цвет.
Кладофора нитчатая зеленая водоросль. Слоевище состоит из ветвящихся нитей, образованных одним рядом многоядерных клеток, отдельные растения имеют вид подводных кустиков Клетки крупные, многоядерные, с толстой, слоистой облчкой. Встречается в пресных и соленых водах
Отдел красные водоросли, или багрянки насчи тывает более 600 родов и около 4000 видов. От других водорослей отличаются краской окраской, которая обусловливается наличием у них, кроме хлорофилла, еще двух пигментов — фикоэритрина (красного цвета) и фикоциана (сине-зеленого цвета). От соотноше ния этих пигментов зависит окраска водорослей (изменяется от красной до почти черной ). Представители отдела в подавляющем большинстве многоклеточ ные организмы сложного строения, и только наиболее примитивные имеют одноклеточное или колониальное слоевище. Чаще слоевище имеет форму кустиков, пластинок, у некоторых оно очень сильно расчленено.
Красные водоросли обитают в морях на больших глубинах, чем зеленые и бурые. Это объясняется тем, что красный пигмент способ ствует улавливанию на больших глубинах зеленых и синих лучей спектра и тем самым улучшает процесс фотосинтеза. В красных водорослях откладывается не обычный крахмал . В отличие от обычного крахмала он йодом окрашивается не в синий, а в красно- бурый цвет.
У красных водорослей отсутствуют зооспоры и сперматозоиды. Бесполое размножение у них происходит при помощи неподвижных спор. Половое размножение — оогамия, но вместо сперматозоидов образуются мужские клетки — спермации, они переносятся к яйцеклеткам водой. Яйцеклетки образуются в специальных органах — карпогонах.
_К красным водорослям относятся делессерия с талломом в виде кустика; полисифония у которой таллом имеет вид ветвящейся нити.
Отдел бурые водоросли
Для представителей отдела бурые водоросли харак терна бурая окраска, обусловленная присутствием в хроматофорах коричневого пигмента — фукоксантина. Наличие фукоксантина маскирует зеленый цвет и придает этим водорослям бурую окраску различных оттенков. Кроме фукоксантина, они содержат ксанто филл и каротин. Отдел бурых водорослей объединяет свыше 900 видов.
Бурые водоросли отличаются обычно крупным многоклеточным талломом. Самые крупные представители водорослей встречаются именно среди бурых водорослей. Некоторые из них, например мак роцистис, достигают 60 м длины, но имеются и мелкие формы вели чиной в несколько миллиметров.
Вместо крахмала в клетках бурых водорослей имеются глюко за и сахаристые вещества — манит и ламинарии, которые придают этим водорослям в вареном виде сладковатый вкус. В качестве запасного вещества они часто откладывают масла.
Таллом бурых водорослей многолетний, но листообразные пластинки отмирают ежегодно, а ранней весной нарастают вновь.
Сложное внешнее строение бурых водорослей обусловливает у них и дифференциацию в анатомическом строении (они имеют различной формы клетки). Некоторые ученые считают, что у этих водорослей имеются даже разные ткани.
Размножаются бурые водоросли различными способами. Неко торые из них размножаются примитивным половым путем — изога мией, когда сливаются 2 одинаковые по форме гаметы. У других, более развитых водорослей (ламинария) наблюдается более сложный половой процесс — оогамия, при котором крупная яйцеклетка сливается с мелкой подвижной мужской гаметой — сперматозои дом.
Бесполое размножение у бурых водорослей осуществляется зооспорами, которые образуются в большом количестве в зооспо рангиях. У бурых водорослей довольно четко выражено чередование поколений; бесполого и полового. На листовидных пластинках у этих водорослей образуются одноклеточные зооспорангии, собран ные в группы, среди которых располагаются бесплодные нити. В каждом зооспорангии образуется 16...64 и более зооспор. Зооспоры внешне одинаковы, а физиологически разные. Одни из них прорастают и образуют микроскопически малые жен ские, а другие — мужские гаметофиты. На мужских гаметофитах в дальнейшем образуются антеридии и в них по одному спермато зоиду, а на женских гаметофитах формируются оогонии, несущие по одной яйцеклетке. После слияния сперматозоида с яйцеклеткой образуется зигота, из которой развивается бесполое поколение — спорофит.
Бурые водоросли являются морскими обитателями, многие из них чаще встречаются в северных морях, нередко они образуют в морях и океанах огромные заросли. В северной части Атланти ческого океана, в Саргассовом море, в огромном количестве встре- чается род бурых водорослей — саргассум. Эти водоросли чаще всего находятся в плавающем состоянии благодаря наличию у них особых пузырей, наполненных воздухом.
Бурые водоросли считаются древней группой растений, у них обнаруживается более высокая степень дифференциации не только внешней, но и внутренней части таллома. Внешне они сходны с выс шими растениями, поэтому некоторые ботаники считают, что эти водоросли, возможно, дали начало высшим растениям.
Отдел бурые водоросли состоит из 4 порядков. Рассмотрим пред ставителей двух порядков: ламинариевые и фукусовые.
Порядок ламинариевые. Это очень крупные водоросли, достигающие иногда 60 м и больше. Таллом их сильно рассечен и, кроме того, имеет хорошо развитые разветвленные ризоиды, которыми водоросли прочно прикрепляются ко дну моря. Обитают ламинариевые в прибрежной полосе морей па глубине 5...10 м и часто образуют огромные заросли подводных «лесов».
К ламинариевым относятся род ламинария (включает 30 видов), род лессония (насчитывает 5 видов) и род макроцистис. Эти водоросли — многолетние растения, отличающиеся одно от другого строением таллома.
Отдел слизевики
Плазмодий образуется в результате слияния голых амебооб разных клеток слизевика и у некоторых видов слизевиков достигает размеров ладони человека. Он обычно окрашен в ярко-желтый цвет и обладает способностью очень медленно амебообразно пере двигаться (0,1 мм/мин). При передвижении плазмодий стремится скрыться от света и направиться к источнику влаги. Встречаются слизевики обычно в тенистых лесах; на гнилых растениях, между корой и древесиной, в трещинах пней, под опавшими листьями. Размножаются спорами.
Положение слизевиков в филогенетической системе неясно, по-видимому, они произошли от каких-то жгутиковых. Из этого отдела рассмотрим плазмодиофору.
Болезнь распространяется через почву и особенно быстро разви вается на кислых почвах.
Зеленые водоросли - самый обширный из всех отделов водорослей, насчитывающий по разным оценкам от 4 до 13 - 20 тысяч видов. Все они имеют зеленый цвет слоевищ, что обусловлено преобладанием в хлоропластах хлорофилла a и b над другими пигментами. Клетки некоторых представителей зеленых водорослей (Хламидомонас, Трентеполия, Гематококкус ) окрашены в красный или оранжевый цвета, что связано с накоплением вне хлоропласта каротиноидных пигментов и их производных.
В морфологическом отношении они отличаются большим разнообразием. Среди зеленых водорослей встречаются одноклеточные, колониальные, многоклеточные и неклеточные представители, активно подвижные и неподвижные, прикрепленные и свободноживущие. Чрезвычайно велик и диапазон их размеров - от нескольких микрометров (что сравнимо по размерам с бактериальными клетками) до 1–2 метров.
Клетки одноядерные или многоядерные, с одним или несколькими хроматофорами, содержащими хлорофилл и каротиноиды. Хлоропласты покрыты двумя мембранами и обычно имеют стигму, или глазок, - фильтр, проводящий синий и зеленый свет к фоторецептору. Глазок состоит из нескольких рядов липидных глобул. Тилакоиды - структуры, где локализованы фотосинтетические пигменты - собраны в стопки (ламеллы) по 2–6. В переходной зоне жгутиков есть звездчатое образование. Жгутиков чаще всего два. Основной компонент клеточной стенки – целлюлоза.
У хлорофит встречаются различные типы питания: фототрофное, миксотрофное и гетеротрофное. Запасной полисахарид зеленых водорослей – крахмал – откладывается внутри хлоропласта. Хлорофиты также могут накапливать липиды, которые откладываются в виде капель в строме хлоропласта и в цитоплазме.
Многоклеточные слоевища нитевидные, трубчатые, пластинчатые, кустистые или иного строения и разнообразной формы. Из известных типов организации таллома у зеленых водорослей отсутствует только амебоидный.
Они широко распространены в пресных и морских водах, в почве и в наземных местообитаниях (на почве, скалах, коре деревьев, стенах домов и пр.). В морях распространено около 1/10 от общего количества видов, которые растут обычно в верхних слоях воды до 20 м. Среди них есть планктонные, перифитонные и бентосные формы. Иначе говоря, зеленые водоросли освоили три основных среды обитания живых организмов: воду – землю – воздух.
Зеленые водоросли обладают положительным (движение к источнику света) и отрицательным (движение от яркого источника света) фототаксисом. Кроме интенсивности освещения, на фототаксис влияет температура. Положительным фототаксисом при температуре 160°С обладают зооспоры видов родов Гематококкус, Улотрикс, Ульва , а также отдельные виды десмидиевых водорослей, у которых движение клеток осуществляется за счет выделения слизи через поры в оболочке.
Размножение. Для зеленых водорослей характерно наличие всех известных способов размножения: вегетативное, бесполое и половое.
Вегетативное размножение у одноклеточных форм происходит делением клетки пополам. Колониальные и многоклеточные формы хлорофит размножаются частями тела (слоевища, или таллома).
Бесполое размножение у зеленых водорослей представлено широко. Осуществляется чаще подвижными зооспорами, реже неподвижными апланоспорами и гипноспорами. Клетки, в которых образуются споры (спорангии), в большинстве случаев ничем не отличаются от остальных вегетативных клеток таллома, реже они имеют иную форму и более крупные размеры. Формирующиеся зооспоры могут быть голыми или покрытыми жесткой клеточной стенкой. Количество жгутиков у зооспор варьирует от 2 до 120. Зооспоры разнообразной формы: шаровидные, эллипсоидные или грушевидные, одноядерные, лишенные обособленной оболочки, с 2–4 жгутиками на переднем, более заостренном конце и хлоропластом в расширенном заднем конце. Обычно они имеют пульсирующие вакуоли и стигму. Зооспоры образуются одиночно или, чаще, в числе нескольких из внутреннего содержимого материнской клетки, выходят наружу через образующееся в оболочке круглое или щелевидное отверстие, реже вследствие ее общего ослизнения. В момент выхода из материнской клетки зооспоры иногда окружены тонким слизистым пузыре, в скором времени расплывающимся (род Улотрикс).
У многих видов вместо зооспор или наряду с ними образуются неподвижные споры – апланоспоры. Апланоспоры - споры бесполого размножения, у которых отсутствуют жгутики, но имеются сократительные вакуоли. Апланоспоры рассматривают как клетки, у которых приостановлено дальнейшее развитие в зооспоры. Они также возникают из протопласта клетки в числе одной или нескольких, но не вырабатывают жгутиков, а, приняв шаровидную форму, одеваются собственной оболочкой, в образовании которой оболочка материнской клетки не участвует. Апланоспоры освобождаются вследствие разрыва или ослизнения оболочек материнских клеток и прорастают после некоторого периода покоя. Апланоспоры с очень толстыми оболочками называются гипноспорами. Они обычно принимают на себя функцию покоящейся стадии. У автоспор, которые представляют собой уменьшенные копии неподвижных вегетативных клеток, отсутствуют сократительные вакуоли. Образование автоспор коррелирует с завоеванием наземных условий, в которых вода не может всегда присутствовать в достаточном количестве.
Половое размножение осуществляется гаметами, возникающих в неизмененных, слегка измененных или значительно преобразованных клетках – гаметангиях. Подвижные гаметы монадного строения, двужгутиковые. Половой процесс у зеленых водорослей представлен различными формами: гологамия, конъюгация, изогамия, гетерогамия, оогамия. При изогамии гаметы морфологически совершенно подобны друг другу и различия между ними являются чисто физиологическими. Зигота одевается толстой оболочкой, нередко со скульптурными выростами, содержит большое количество запасных веществ и прорастает сразу или после некоторого периода покоя. При прорастании содержимое зиготы у большинства видов делится на четыре части, которые выходят из оболочки и прорастают в новые особи. Значительно реже гаметы развиваются в новый организм без слияния, сами по себе, без образования зиготы. Такое размножение называется партеногенезом , а споры, образующиеся из отдельных гамет, – партеноспорами .
При гетерогамии обе гаметы различаются между собой по величине и иногда по форме. Более крупные гаметы, часто менее подвижные, принято считать женскими, меньшие по величине и более подвижные – мужскими. В одних случаях различия эти невелики, и тогда говорят просто о гетерогамии, в других весьма значительны.
Если женская гамета неподвижна и напоминает больше яйцеклетку, то подвижная мужская становится сперматозоидом, а половой процесс получает название оогамии. Гаметангии, в которых возникают яйцеклетки, называются оогониями, от вегетативных клеток они отличаются как по форме, так и по величине. Гаметангии, в которых образуются сперматозоиды, называются антеридиями . Зигота, получившаяся в результате оплодотворения яйцеклетки сперматозоидом, формирует толстую оболочку и называется ооспорой .
При типичной оогамии яйцеклетки крупные, неподвижные и развиваются чаще всего по одной в оогонии, сперматозоиды мелкие, подвижные, образуются в антеридии в большом количестве. Оогонии и антеридии могут развиваться на одной особи, в этом случае водоросли однодомные; если они развиваются на разных особях – двудомные. Оплодотворенная яйцеклетка одевается толстой бурой оболочкой; нередко соседние с ней клетки дают короткие веточки, которые обрастают ооспору, оплетая ее однослойной корой.
Жизненные циклы . У большинства представителей зеленых водорослей жизненный цикл гаплобионтный с зиготической редукцией. У таких видов диплоидной стадией является только зигота – клетка, получающаяся в результате оплодотворения яйцеклетки сперматозоидом. Другой тип жизненного цикла – гаплодиплобионтный со спорической редукцией – встречается у Ульвовых, Кладофоровых и некоторых Трентеполиевых. Для этих водорослей характерно чередование диплоидного спорофита и гаплоидного гаметофита. Гаплодиплобионтный жизненный цикл с соматической редукцией известен только у Празиолы . Наличие диплобионтного жизненного цикла у Бриопсидовых и Дазикладиевых подвергается сомнению.
У некоторых Улотриксовых одна и та же особь может давать начало как зооспорам, так и гаметам. В других случаях зооспоры и гаметы образуются на разных особях, т.е. жизненный цикл водорослей включает в себя как половую (гаметофит), так и бесполую (спорофит) форму развития. Спорофит обычно диплоидный, т.е. имеет в клетках двойной набор хромосом, гаметофит гаплоидный, т.е. имеет одинарный набор хромосом. Это наблюдается в тех случаях, когда мейоз происходит при образовании спор (спорическая редукция) и часть жизненного цикла водоросли от зиготы до образования спор проходит в диплофазе, а часть от споры до образования гамет в гаплофазе. Такой цикл развития характерен для видов рода Ульва.
В пределах Улотриксовых водорослей широко распространена зиготическая редукция, когда мейоз происходит при прорастании зиготы. Диплоидной в этом случае оказывается только зигота, весь остальной жизненный цикл протекает в гаплофазе. Значительно реже встречается гаметическая редукция, когда мейоз происходит при образовании гамет. В этом случае гаплоидными являются только гаметы, а весь остальной цикл диплоидный.
Систематика
До сих пор отсутствует единая устоявшаяся система зеленых водорослей, особенно в отношении группировки порядков в различные предлагаемые классы. Очень долго типу дифференциации таллома придавали основное значение при выделении порядков у зеленых водорослей. Однако в последнее время в связи с накоплением данных об ультраструктурных особенностях жгутиковых клеток, типе митоза и цитокинеза и др. очевидна гетерогенность многих таких порядков.
Отдел включает 5 классов: Ульвофициевые– Ulvophyceae, Брипсодовые – Bryopsidophyceae, Хлорофициевые – Chlorophyceae , Требуксиевые –Trebouxiophyceae , Празиновые – Prasinophyceae .
Класс Ульвофициевые – Ulvophyceae
Известно около 1 тысячи видов. Название класса происходит от типового рода Ulva . Включает виды с нитчатым и пластинчатым талломом. Жизненные циклы разнообразны. Виды преимущественно морские, реже пресноводные и наземные. Некоторые входят в состав лишайников. У морских представителей в клеточных стенках может откладываться известь.
Порядок Улотриксовые – Ulotrichales .
Род Улотрикс (рис. 54). Виды Улотрикса обитают чаще в пресных, реже в морских, солоноватых водоемах и в почве. Они прикрепляются к подводным предметам, формируя ярко-зеленые кустики размером до 10 см и более. Неразветвленные нити Улотрикса , состоящие из одного ряда цилиндрических клеток с толстыми целлюлозными оболочками, прикрепляются к субстрату бесцветной конической базальной клеткой, выполняющей функции ризоида. Характерным является строение хроматофора, который имеет вид постенной пластинки, образующей незамкнутый поясок или кольцо (цилиндр).
Рис. 54. Улотрик c (по:): 1 – нитчатый таллом, 2 – зооспора, 3 – гамета, 4 – копуляция гамет
Бесполое размножение Улотрикса осуществляется 2 следующими способами: распадением нити на короткие участки, развивающиеся в новую нить, или образованием в клетках четырехжгутиковых зооспор. Зооспоры выходят из материнской клетки, сбрасывают один за другим жгутики, прикрепляются боком к субстрату, покрываются тонкой целлюлозной оболочкой и прорастают в новую нить. Половой процесс изогамный. После оплодотворения зигота вначале плавает, затем оседает на дно, теряет жгутики, вырабатывает плотную оболочку и слизистую ножку, которой прикрепляется к субстрату. Это покоящийся спорофит. После периода покоя происходит редукционное деление ядра и зигота прорастает зооспорами. Так в жизненном цикле Улотрикса происходит чередование поколений, или смена половой и бесполой форм развития: нитчатый многоклеточный гаметофит (поколение, формирующее гаметы) сменяется одноклеточным спорофитом – поколением, которое представлено своеобразной зиготой на ножке и способно образовывать споры.
Порядок Ульвовые - Ulvales . Имеют пластинчатое, мешковидное, трубчатое или, редко, нитчатое слоевище всевозможных оттенков зеленого цвета. По краю пластины могут быть волнистыми либо складчатыми, для прикрепления к субстрату снабжены короткой ножкой или основанием с небольшим базальным диском. Морские и пресноводные виды. Наиболее распространены в прибрежных водах дальневосточных морей виды родов Ульва, Монострома, Корнманния и Ульвария.
Род Ульва (рис. 55). Таллом представляет собой светло-зеленую или ярко-зеленую, тонкую двуслойную, нередко перфорированную пластину либо однослойную полую трубку, прикрепленную к субстрату суженным в короткий черешок основанием.
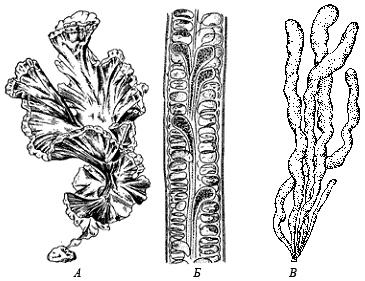
Рис. 55. Ульва : А – внешний вид Ульвы окончатой , Б – поперечный срез таллома, В – внешний вид Ульвы кишечницы
Смена форм развития в жизненном цикле Ульвы сводится к изоморфной, когда бесполая стадия (спорофит) и половая стадия (гаметофит) морфологически подобны друг другу, и гетероморфной, когда они морфологически различны. Гаметофит многоклеточный, пластинчатый, спорофит одноклеточный. На гаметофитах образуются двужгутиковые гаметы, на спорофитах - четырехжгутиковые зооспоры.
Виды рода встречаются в морях всех климатических зон, хотя предпочитает теплые воды. Например, на мелководье Черного и Японского морей Ульва - один из самых массовых родов водорослей. Многие виды Ульвы выносят опреснение воды; их часто можно встретить в устьях рек.
Класс Бриопсидовые –Bryopsidophyceae
Известно около 500 видов. Слоевище неклеточное. Образовано простыми или переплетенными сифонными нитями, образующими сложные структуры. Таллом в виде пузырей, кустиков, губчатых, дихотомически разветвленных кустов. Слоевище сегментированное, имитирующее многоклеточное, из нескольких или многих ядерных клеток. Нити и кустики всех оттенков зеленого или буроватого цвета.
Порядок Бриопсидовые – Bryopsidales
Большинство видов встречается в пресных и солоноватых водоемах. Некоторые из них растут на почве, на камнях, песке и иногда на солончаках.
Род Бриопсис – нитевидные кустики до 6-8 см высоты, перисто или неправильно разветвленные, верхние веточки с перетяжками у основания. Слоевище сифонного неклеточного строения. Растет единичными кустиками или небольшими куртинами в в прибрежной зоне, обитает в теплых и умеренных морях (приложение, 7Б).
Род Кодиум – шнуровидные дихотомически разветвленные кустики 10–20 см высоты, губчатые. мягкие, прикрепляются дисковидной подошвой. Внутренняя часть слоевища образована сложно переплетенными сифонными нитями. Растет на мягких и твердых грунтах в сублиторальной зоне до глубины 20 м одиночными растениями или небольшими группами (приложение, 7А, Б).
Род Каулерпа включает около 60 видов морских водорослей, ползучие, распростертые на грунте части слоевища которых имеют вид ветвящихся цилиндров, достигающих в длину нескольких десятков сантиметров. Через определенные интервалы вниз от них отходят обильно ветвящиеся ризоиды, закрепляющие растение в грунте, а вверх – плоские листообразные вертикальные побеги, в которых сосредоточены хлоропласты.

Рис. 56. Каулерпа: А – внешний вид таллома; Б – срез таллома с целлюлозными балками
Таллом каулерпы, несмотря на свои крупные размеры, не имеет клеточного строения – в нем полностью отсутствуют поперечные перегородки, и формально он представляет собой одну гигантскую клетку (рис.56). Такое строение таллома называют сифонным . Внутри таллома каулерпы располагается центральная вакуоль, окруженная слоем цитоплазмы, содержащей многочисленные ядра и хлоропласты. Различные части таллома растут у своих верхушек, где скапливается цитоплазма. Центральную полость во всех частях таллома пересекают цилиндрические скелетные тяжи – целлюлозные балки, придающие телу водоросли механическую прочность.
Каулерпа легко размножается вегетативно: при отмирании более старых частей таллома отдельные участки его с вертикальными побегами становятся независимыми растениями. Виды этого рода обитают главным образом в тропических морях, и лишь немногие заходят в субтропические широты, например, распространенная в Средиземном море Каулерпа прорастающая . Эта водоросль предпочитает мелководье со спокойной водой, например, лагуны, защищенные от действия постоянного прибоя коралловыми рифами, и поселяется как на различных твердых субстратах – камнях, рифах, скалах, на песчаном и илистом грунте.
Класс Хлорофициевые –Chlorophyceae
Известно около 2,5 тысяч видов. Слоевище одноклеточное или колониальное моннадное, свободно живущее.
Порядок Вольвоксовые - Volvocales .
Род Хламидомонада (рис. 57)включает свыше 500 видов одноклеточных водорослей, которые обитают в пресных, мелких, хорошо прогреваемых и загрязненных водоемах: прудах, лужах, канавах и т.п. При их массовом размножении вода приобретает зеленую окраску. Хламидомонада также обитает на почве и на снегу. Ее тело имеет овальную, грушевидную или шаровидную форму. Клетка одета плотной оболочкой, нередко отстающей от протопласта, с двумя одинаковыми жгутиками на переднем конце; с их помощью хламидомонада активно передвигается в воде. Протопласт содержит 1 ядро, чашевидный хроматофор, стигму и пульсирующие вакуоли.

Рис. 57. Строение и развитие Хламидомонады: А – вегетативная особь; Б – пальмеллевидная стадия; В – размножение (молодые особи внутри материнской клетки)
Хламидомонады размножаются преимущественно бесполым путем. При подсыхании водоема они размножаются делением клетки пополам. Клетки останавливаются, теряют жгутики, стенки их клеток ослизняются, и в таком неподвижном состоянии клетки переходят к делению. Стенки образующихся при этом дочерних клеток также ослизняются, так что в итоге образуется система вложенных друг в друга слизистых обверток, в которых группами располагаются неподвижные клетки. Это - пальмеллевидное состояние водоросли. При попадании в воду клетки снова образуют жгутики, покидают материнскую клетку в виде зооспор и переходят к одиночному монадному состоянию.
В благоприятных условиях хламидомонада интенсивно размножается другим путем – клетка останавливается, и ее протопласт, несколько отстав от стенки, последовательно делится продольно на две, четыре или восемь частей. Эти дочерние клетки образуют жгутики и выходят наружу в виде зооспор, которые вскоре снова приступают к размножению.
Половой процесс у хламидомонады изогамный или оогамный. Гаметы меньших размеров образуются внутри материнской клетки так же, как и зооспоры, но в большем количестве (16, 32 или 64). Оплодотворение происходит в воде. Оплодотворенная яйцеклетка покрывается многослойной оболочкой и оседает на дно водоема. После периода покоя зигота делится мейотически с образованием 4 гаплоидных дочерних особей хламидомонады.
Род Вольвокс – наиболее высокоорганизованные представители порядка, образуют гигантские колонии, состоящие из сотен и тысяч клеток. Колонии имеют вид слизистых, диаметром до 2 мм, шариков, в периферическом слое которых расположено до 50 тыс. клеток со жгутиками, сросшихся своими боковыми ослизненными стенками друг с другом и соединенных плазмодесмами (рис. 58). Внутренняя полость

Рис. 58. Внешний вид колоний Вольвокса
шара заполнена жидкой слизью. В колонии существует специализация клеток: периферическую ее часть составляют вегетативные клетки, а между ними разбросаны более крупные – репродуктивные.
Около десятка из клеток колонии – это гонидии, клетки бесполого размножения. В результате многократных делений они дают начало молодым, дочерним колониям, которые выпадают внутрь материнского шара и освобождаются лишь после его разрушения. Половой процесс – оогамия. Оогонии и антеридии возникают также из репродуктивных клеток. Колонии однодомные и двудомные. Виды рода встречаются в прудах и старицах рек, где в период интенсивного размножения вызывают «цветение» воды.
Класс Требуксиевые – Trebouxiophyceae
Класс назван по типовому роду Trebouxia . Включает в основном одноклеточные коккоидные формы. Встречаются сарциноидные и нитчатые представители. Пресноводные и наземные, реже морские формы, многие формируют симбиозы. Около 170 видов.
Порядок Хлорелловые - Chlorellales . Объединяет коккоидных автоспоровых представителей.
Род Хлорелла – одноклеточные водоросли в виде неподвижного шарика. Клетка одета гладкой оболочкой; содержит одно ядро и пристенный, цельный, рассеченный или лопастной хроматофор с пиреноидом. Клеточная стенка ряда видов наряду с целлюлозой содержит спорополленин – чрезвычайно устойчивое к действию различных ферментов вещество, встречающееся также в пыльцевых зернах и спорах высших растений. Размножается хлорелла бесполым путем, образуя до 64 неподвижных автоспор. Полового размножения нет. Хлорелла распространена в различных водоемах, встречается на сырой почве, коре деревьев, входит в состав лишайников.
Порядок Требуксиевые - Trebouxiales . Включает роды и виды, входящие в состав лишайников.
Род Требуксия – одноклеточная водоросль. Сферические клетки имеют единственный осевой звездчатый хлоропласт с одним пиреноидом. Бесполое размножение осуществляется голыми зооспорами. Встречается или в свободноживущем виде в наземных местообитаниях (на коре деревьев), или как фотобионт лишайников.
Класс Празиновые – Prasinophyceae
Название класса происходит от греч. prasinos – зеленый. Жгутиковые или, реже, коккоидные или пальмеллоидные одноклеточные организмы.
Порядок Пирамимонадовые - Pyramimonadales . Клетки несут 4 или больше жгутиков, три слоя чешуек. Митоз открытый, с веретеном, сохраняющимся в телофазе, цитокинез идет за счет образования борозды деления.
Род Пирамимонас – одноклеточные организмы (рис. 59). От переднего конца клетки отходит 4–16 жгутиков, которые могут быть в пять раз длиннее клетки. Хлоропласт обычно единственный, с одним пиреноидом и одним или больше глазками. Клетки и жгутики покрыты несколькими слоями чешуек. Широко распространены в пресных, солоноватых и морских водах. Встречаются в планктоне и бентосе, могут вызывать "цветение" воды.

Рис. 59. Внешний вид водоросли Пирамимонас
Порядок Хлородендровые – Chlorodendrales . Клетки сжатые, с четырьмя жгутиками, покрыты текой, митоз закрытый, цитокинез идет за счет образования борозды деления.
Род Тетраселмис может встречаться в виде подвижных четырехжгутиковых клеток или в виде неподвижных клеток, прикрепленных слизистыми ножками. Клетки покрыты текой. При делении клеток новая тека формируется вокруг каждой дочерней клетки внутри теки материнской. На переднем конце клетки через отверстие в теке выходят жгутики, которые покрыты волосками и чешуйками. Хлоропласт один, с базальным пиренодом. Клетки обычно зеленого цвета, но иногда приобретают красную окраску, что связано с накоплением каротиноидов. Морские представители, могут обитать в морских плоских червях.
Экология и значение
Зеленые водоросли широко распространены по всему миру. Большинство из них можно встретить в пресных водоемах, но немало солоноватоводных и морских форм. Нитчатые зеленые водоросли, прикрепленные или неприкрепленные, наряду с диатомовыми и синезелеными являются преобладающими бентосными водорослями континентальных водоемов. Они встречаются в водоемах различной трофности (от дистрофных до эвтрофных) и с различным содержанием органических веществ (от ксено- до полисапробных), водородных ионов (от щелочных до кислых), при различных температурах (термо-, мезо- и криофилы).
Среди зеленых водорослей имеются планктонные, перифитонные и бентосные формы. В группе морского пикопланктона празиновая водоросль Остреококкус считается самой маленькой эукариотной свободноживущей клеткой. Есть виды зеленых водорослей, которые приспособились к жизни в почве и наземных местообитаниях. Их можно встретить на коре деревьев, скалах, различных постройках, на поверхности почв и в толще воздуха. В этих местообитаниях особенно распространены представители родов Трентеполия и Требуксия . Зеленые водоросли вегетируют в горячих источниках при температуре 35–52°С, а в отдельных случаях до 84°С и выше, нередко при повышенном содержании минеральных солей или органических веществ (сильно загрязненные горячие сточные воды заводов, фабрик, электростанций или атомных станций). Они также преобладают среди криофильных видов водорослей. Они могут вызывать зеленое, желтое, голубое, красное, коричневое, бурое или черное «цветение» снега или льда. Эти водоросли находятся в поверхностных слоях снега или льда и интенсивно размножаются в талой воде при температуре около 0 °С. Лишь немногие виды имеют стадии покоя, тогда как большинство лишены каких-либо специальных морфологических приспособлений к низким температурам.
В пересоленных водоемах преобладают одноклеточные подвижные зеленые водоросли – гипергалобы, клетки которых лишены оболочки и окружены лишь плазмалеммой. Эти водоросли отличаются повышенным содержанием хлористого натрия в протоплазме, высоким внутриклеточным осмотическим давлением, накоплением в клетках каротиноидов и глицерина, большой лабильностью ферментных систем и обменных процессов. В соленых водоемах они нередко развиваются в массовом количестве, вызывая красное или зеленое «цветение» соленых водоемов.
Микроскопические одноклеточные, колониальные и нитчатые формы зеленых водорослей приспособились к неблагоприятным условиям существования в воздушной среде. В зависимости от степени увлажнения их подразделяют на 2 группы: воздушные водоросли, обитающие в условиях только атмосферного увлажнения, и, следовательно, испытывающие постоянную смену влажности и высыхания; водновоздушные водоросли, подвергающиеся действию постоянного орошения водой (под брызгами водопада, прибоя и т. д.). Условия существования водорослей аэрофильных сообществ очень своеобразны и характеризуются, прежде всего, частой и резкой сменой двух факторов - влажности и температуры.
Сотни видов зеленых водорослей обитают в почвенном слое. Почва как биотоп имеет сходство и с водными и с воздушными местообитаниями: в ней есть воздух, но насыщенный водяными парами, что обеспечивает дыхание атмосферным воздухом без угрозы высыхания. Интенсивное развитие водорослей как фототрофных организмов возможно только в пределах проникновения света. В целинных почвах это поверхностный слой почвы толщиной до 1 см, в обрабатываемых почвах он немного толще. Однако в толще почвы, куда не проникает свет, жизнеспособные водоросли обнаруживаются на глубине до 2 м в целинных почвах и до 3 м – в пахотных. Это объясняется способностью некоторых водорослей переходить в темноте к гетеротрофному питанию. Многие водоросли сохраняются в почве в покоящемся состоянии.
Для поддержания своей жизнедеятельности почвенные водоросли имеют некоторые морфологические и физиологические особенности. Это относительно мелкие размеры почвенных видов, а также способность к обильному образованию слизи – слизистых колоний, чехлов и обверток. Благодаря наличию слизи, водоросли быстро поглощают воду при увлажнении и запасают ее, замедляя высыхание. Характерной чертой почвенных водорослей является «эфемерность» их вегетации – способность быстро переходить из состояния покоя к активной жизнедеятельности и наоборот. Они также способны переносить разные колебания температуры почвы. Диапазон выживаемости ряда видов лежит в пределах от -200 до +84 °С и выше. Наземные водоросли составляют важную часть растительности Антарктиды. Они окрашены почти в черный цвет, поэтому температура их тела оказывается выше температуры окружающей среды. Почвенные водоросли являются также важными компонентами биоценозов аридной (засушливой) зоны, где почва в летнее время нагревается до 60–80°С. Защитой от избыточной инсоляции служат темные слизистые чехлы вокруг клеток.
Своеобразную группу представляют эндолитофильные водоросли, связанные с известковым субстратом. Во-первых, это – сверлящие водоросли. Например, водоросли из рода Гомонтия сверлят раковины перловиц и беззубок, внедряются в известковый субстрат в пресных водоемах. Они делают известковый субстрат рыхлым, легко поддающимся различным воздействиям химических и физических факторов. Во-вторых, ряд водорослей в пресных и морских водоемах способны переводить растворенные в воде соли кальция в нерастворимые и отлагающие их на своих талломах. Ряд тропических зеленых водорослей, в частности Галимеда , откладывает в талломе карбонат кальция. Они принимают активное участие в постройке рифов. Гигантские залежи останков Галимеды , иногда достигающие 50 м в высоту, встречаются в континентальных шельфовых водах, связанных с Большим Барьерным Рифом в Австралии и других регионах, на глубине от 12 до 100 м.
Зеленые требуксиевые водоросли, вступая в симбиотические отношения с грибами, входят в состав лишайников. Около 85% лишайников содержат в качестве фотобионта одноклеточные и нитчатые зеленые водоросли, 10% - цианобактерии и 4% (и более) содержат одновременно синезеленые и зеленые водоросли. В качестве эндосимбионтов они существуют в клетках простейших, криптофитовых водорослей, гидр, губок и некоторых плоских червей. Даже хлоропласты отдельных сифоновых водорослей, например Кодиума , становятся симбионтами для голожаберных моллюсков. Эти животные питаются водорослями, хлоропласты которых остаются жизнеспособными в клетках дыхательной полости, причем на свету они очень эффективно фотосинтезируют. Ряд зеленых водорослей развивается на шерсти млекопитающих. Эндосимбионты, претерпевая морфологические изменения по сравнению со свободноживущими представителями, не теряют способности фотосинтезировать и размножаться внутри клеток хозяина.
Хозяйственное значение . Повсеместное распространение зеленых водорослей определяет их огромное значение в биосфере и хозяйственной деятельности человека. Благодаря способности к фотосинтезу они являются основными продуцентами громадного количества органических веществ в водоемах , которые широко используются животными и человеком. Поглощая из воды углекислый газ, зеленые водоросли насыщают ее кислородом, необходимым всем живым организмам. Велика их роль в биологическом круговороте веществ. Быстрое размножение и очень высокая скорость ассимиляции (примерно в 3-5 раз выше, чем у наземных растений) приводят к тому, что за сутки масса водоросли увеличивается более чем в 10 раз. При этом в клетках хлореллы накапливаются углеводы (в селекционных штаммах их содержание достигает 60%), липиды (до 85%), витамины B, С и К. Белок хлореллы, на долю которого может приходиться до 50% сухой массы клетки, содержит все незаменимые аминокислоты. Уникальная способность видов Хлореллы ассимилировать от 10 до 18% световой энергии (против 1–2% у наземных растений) позволяет использовать эту зеленую водоросль для регенерации воздуха в замкнутых биологических системах жизнеобеспечения человека при длительных космических полетах и подводном плавании.
Ряд видов зеленых водорослей используют как индикаторные организмы в системе мониторинга водных экосистем. Наряду с фототрофным способом питания многие одноклеточные зеленые водоросли (хламидомонады) способны всасывать через оболочку растворенные в воде органические вещества, что способствует активному очищению загрязненных вод, в которых развиваются эти виды. Поэтому их применяют для очистки и доочистки загрязненных вод, а также как корм в рыбохозяйственных водоемах.
Некоторые виды зеленых водорослей используются населением ряда стран в пищу . Для пищевых целей, например, в Японии специально культивируют виды рода Ульва . Эти водоросли широко используют, особенно в странах Юго-Восточной Азии, под названием Морского салата. Ульвовые по содержанию белка (до 20%) заметно превосходят другие виды водорослей. Отдельные виды зеленых водорослей используют в качестве продуцентов физиологически активных веществ. Зеленые водоросли - хороший модельный объект для разнообразных биологических исследований. Виды Гематококкуса культивируют для получения астаксантина, Ботриококкус - для получения липидов. В то же время с «цветением» воды одного из озер на Тайване, вызванного Ботриококкусом, связывают гибель рыб.
Виды родов Хлорелла и Хламидомонас - модельные объекты для изучения фотосинтеза в растительных клетках. Хлорелла , благодаря очень высоким темпам размножения, является объектом массового культивирования для использования в различных областях
Поверхностные пленки зеленых водорослей имеют большое противоэрозионное значение . Скрепляющее значение имеют некоторые одноклеточные виды зеленых водорослей, выделяющие обильную слизь. Слизистые вещества клеточных оболочек склеивают частицы почвы. Развитие водорослей влияет на структурирование мелкозема, придавая ему водостойкость и препятствуя выносу с поверхностного слоя. Влажность почвы под водорослевыми пленками обычно выше, чем там, где они отсутствуют. Кроме того, пленки уменьшают водопроницаемость почвы и замедляют испарение воды, что оказывает влияние и на солевой режим почвы. Уменьшается вымывание из почвы легкорастворимых солей; их содержание под макроразрастаниями водорослей выше, чем на других участках. В то же время замедляется поступление солей из глубоких слоев почвы.
Почвенные водоросли оказывают влияние и на рост и развитие высших растений. Выделяя физиологически активные вещества, они ускоряют рост проростков, особенно их корней.
Среди зеленых водорослей, обитающих в загрязненных водоемах, доминируют обычно хлорококковые, устойчивые к длительному воздействию многих токсических веществ.
Клетки водорослей способны аккумулировать из воды различные химические элементы, причем коэффициенты их накопления достаточно высоки. Мощными концентраторами являются пресноводные зеленые водоросли, особенно нитчатые. При этом интенсивность накопления в них металлов гораздо выше, чем в других пресноводных гидробионтах. Немалый интерес представляет способность водорослей концентрировать в себе радиоактивные элементы. Отмершие клетки водорослей удерживают накопленные элементы не менее прочно, чем живые, а в некоторых случаях десорбция из мертвых клеток меньше, чем из живых. Способность ряда родов (Хлорелла, Сценедесмус и др.) концентрировать и прочно удерживать в своих клетках химические элементы и радионуклиды позволяет использовать их в специализированных системах очистки для дезактивации промышленных сточных вод, например для дополнительной очистки слабоактивных сточных вод АЭС.
Некоторые зеленые водоросли являются антагонистами вируса гриппа, полиовируса и др. Выделяемые водорослями биологически активные вещества играют важную роль в обеззараживании воды и подавлении жизнедеятельности патогенной микрофлоры.
В специальных биологических прудах сообщества водорослей и бактерий используют для разложения и детоксикации гербицидов . Доказана способность ряда зеленых водорослей гидролизовать гербицид пропанил, который быстрее разрушается бактериями.
Контрольные вопросы
Назовите характерные черты строения клетки зеленых водорослей.
Какие пигменты и типы питания известны у зеленых водорослей?
Как размножаются зеленые водоросли? Что такое зооспоры, апланоспоры, автоспоры?
Какие классы выделяют у зеленых водорослей?
Назовите характерные особенности зеленых водорослей класса Ульвофициевые.
Назовите характерные особенности зеленых водорослей класса Бриопсидовые.
Назовите характерные особенности зеленых водорослей класса Хлорофициевые.
Назовите характерные особенности зеленых водорослей класса Требуксиевые.
Назовите характерные особенности зеленых водорослей класса Празиновые.
В каких местообитаниях встречаются зеленые водоросли? Охарактеризуйте их основные экологические группы.
Роль и значение зеленых водорослей в природе.
Каково хозяйственное значение зеленых водорослей?
Что такое «цветение воды»? Участие зеленых водорослей в биологической очистке вод.
Зеленые водоросли как нетрадиционные источники энергии.
Водоросли, одноклеточные и многоклеточные формы бентосных водорослей. Здесь встречаются все морфологические типы слоевища , кроме ризоподиальных одноклеточных и крупных многоклеточных форм со сложным строением. Многие нитчатые зелёные водоросли крепятся к субстрату только на ранних стадиях развития, затем они становятся свободноживущими, формируя маты или шары.
| Зелёные водоросли | ||||||||||||
| Разнообразие сифоновых водорослей . Иллюстрация из книги Эрнста Геккеля Kunstformen der Natur , 1904 |
||||||||||||
| Научная классификация | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Международное научное название | ||||||||||||
|
Chlorophyta Pascher , 1914 |
||||||||||||
|
||||||||||||
Зелёные водоросли - самый обширный на данное время отдел водорослей: по приблизительным подсчётам сюда входит от 13 000 до 20 000 видов. Все они отличаются в первую очередь чисто-зелёным цветом своих слоевищ, сходным с окраской высших растений и вызванным преобладанием хлорофилла над другими пигментами .
Строение
Жгутиковые клетки зелёных водорослей являются изоконтами - жгутики имеют сходную структуру, хотя они могут различаться по длине. Обычно имеется два жгутика, но их может быть также четыре или много. Жгутики зелёных водорослей не имеют мастигонем (в отличие от гетероконт), но могут иметь изящные волоски или чешуйки.
Жизненные циклы
Жизненные циклы зелёных водорослей очень разнообразны. Здесь встречаются всевозможные типы.
Гаплобионтный с зиготической редукцией (Hydrodictyon reticulatum , Eudorina ). Двужгутиковые гаметы освобождаются из родительской клетки через пору в её оболочке, слияние гамет осуществляется с помощью трубки. Далее зигота превращается в покоящуюся зигоспору, а после периода физиологического покоя прорастает с образованием 4 зооспор (в результате мейотического деления). Каждая зооспора формирует полиэдр и прорастает, формируя небольшие сферические сеточки из слипшихся зооспор.
Гапло-диплобионтный со спорической редукцией (Ulva , Ulothrix , некоторые виды Cladophora ). Двужгутиковые изогаметы выходят из материнской клетки, после чего гаметы образованные разными нитями сливаются в воде. Образуется четырехжгутиковая зигота, которая активно парит в воде. После этого она опускается на какой либо субстрат и покрывается плотной оболочкой, превращаясь таким образом в дубинкообразную клетку (кодиолум), далее следует стадия физиологического покоя. При наступлении благоприятных условий прорастает в 4-16 зооспор или апланоспор, которые после непродолжительного периода плавания прикрепляются к субстрату и прорастают в новые нити. Активируют выход из покоящегося состояния разные факторы: повышение температуры, изменение среды и др.
Диплобионтный с гаметической редукцией (Bryopsis ). Планозигота оседает и прорастает в нитчатое слоевище с крупным ядром, ядро делится таким образом образуются стефаноконтные зооспоры прорастающие в вегетативный таллом.
Особенно много зелёных водорослей развивается весной, когда все камни на литорали бывают покрыты сплошным изумрудным налётом из зелёных водорослей, резко контрастирующим с белым снегом, лежащим на прибрежных камнях. Ворсистый зелёный ковер на камнях образуют развивающиеся нитчатки - улотрикс (Ulothrix ) и уроспора (Urospora ). Летом часто развивается много эгагропилы (Aegagropila linnaei ) (syn. Cladophora aegagropila ), которая нередко имеет вид зелёной слизистой массы. На открытом скалистом побережье ярко-зелёные разветвленные кустики образует акросифония (Acrosiphonia ).
Роль в природе и использование
Некоторые зелёные водоросли (например, ульва) широко употребляются в пищу . Хлореллу используют в качестве индикатора уровня загрязнения воды и содержат на космических кораблях, подводных лодках для очистки воздуха от углекислого газа.







