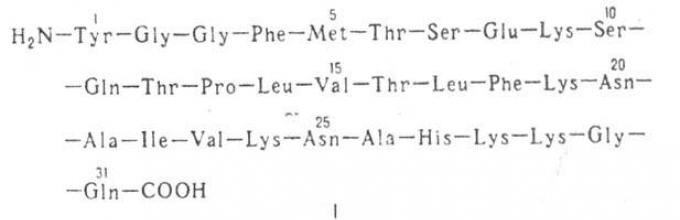
ΟΠΙΟΕΙΔΗ ΠΕΠΤΙΔΙΑ, ομάδα φυσικών και συνθετικών. πεπτίδια παρόμοια με τα οπιούχα (μορφίνη, κωδεΐνη κ.λπ.) ως προς την ικανότητά τους να συνδέονται με υποδοχείς οπιούχων του σώματος (δομές του νευρικού συστήματος, σε σχέση με τα οποία τα οπιούχα είναι εξωγενείς συνδέτες). Φυσικά ΟΠΙΟΕΙΔΗ ΠΕΠΤΙΔΙΑ (ενδογενείς συνδέτες υποδοχέων οπιούχων) απομονώθηκαν για πρώτη φορά το 1975 από τον εγκέφαλο των θηλαστικών. Αυτές ήταν οι λεγόμενες εγκεφαλίνες - λευκίνη-εγκεφαλίνη H 2 N-Tyr-Gly-Gly- Phe-Leu-COOH (μοριακό βάρος 556, βλ. ονομασίες γραμμάτων στο άρθρο Αμινοξέα) και μεθειονίνη-εγκεφαλίνη H 2 N-Tur - Gly -Gly-Phe-Met-COOH (μοριακό βάρος 574), τα οποία είναι πενταπεντίδια που διαφέρουν μόνο στο C-τελικό υπόλειμμα αμινοξέος. Η αλληλουχία αμινοξέων της μεθειονίνης-εγκεφαλίνης είναι πανομοιότυπη με το θραύσμα 61-65 της -λιποτροπίνης.
Άλλα ΟΠΙΟΕΙΔΗ ΠΕΠΤΙΔΙΑ, τα οποία έλαβαν την ονομασία της ομάδας ενδορφίνες, απομονώθηκαν επίσης από εκχυλίσματα ιστών της υπόφυσης και του υποθαλάμου των θηλαστικών. Όλα αυτά περιέχουν συνήθως ένα υπόλειμμα εγκεφαλίνης στη Ν-τελική περιοχή του μορίου. Υπάρχουν -ενδορφίνη (τύπος Ι), -ενδορφίνη (το μόριο της είναι πανομοιότυπο με το θραύσμα 1-16 (3-ενδορφίνη), -ενδορφίνη (αποτελείται από ένα υπόλειμμα -ενδορφίνης που συνδέεται στο C-άκρο με ένα υπόλειμμα λευκίνης) και -ενδορφίνη (-ενδορφίνη συνδεδεμένη στο C-άκρο με Phe-Lys). -νεο-ενδορφίνη (διαφέρει από την προηγούμενη απουσία υπολείμματος λυσίνης στο C-άκρο), δυνορφίνη Α (III), δυνορφίνη Β (IV) , δυνορφίνη-32 (αποτελείται από υπολείμματα δυνορφίνης Α και δυνορφίνης Β που συνδέονται με το διπεπτίδιο Lys-Arg, αντίστοιχα στα C- και Ν-άκρα), δερμορφίνη (V), κυοτορφίνη (H 2N-Tyr-Arg-COOH), casomorphin-5 (H2N-Tyr-Pro-Phe-Pro-Gly-COOH), casomorphin-7 (διαφέρει από την προηγούμενη στην παρουσία στο C-άκρο δύο επιπλέον υπολειμμάτων αμινοξέων Pro-Ile και μερικά άλλα πεπτίδια.

Όλα τα ενδογενή ΟΠΙΟΕΙΔΗ ΠΕΠΤΙΔΙΑ συντίθενται στον οργανισμό με τη μορφή μεγάλων πρόδρομων πρωτεϊνών, από τις οποίες απελευθερώνονται ως αποτέλεσμα της πρωτεόλυσης. Τρεις διαφορετικές πρόδρομες πρωτεΐνες των OPIOID PEPTIDES είναι γνωστές: η προεγκεφαλίνη, η προοπιομελανοκορτίνη και η προδυνορφίνη. Χώρος η δομή των εγκεφαλινών και της μορφίνης είναι παρόμοια. Οι εγκεφαλίνες και οι ενδορφίνες έχουν αναλγητική δράση (όταν εγχέονται απευθείας στον εγκέφαλο) και μειώνουν την κινητική δραστηριότητα. δραστηριότητα του γαστρεντερικού σωλήνα, επηρεάζουν τη συναισθηματική κατάσταση. Η επίδραση των OPIOID PEPTIDES εξαφανίζεται λίγα δευτερόλεπτα μετά τη χορήγηση της ναλοξόνης (VI), ενός ανταγωνιστή της μορφίνης.

Ταυτόχρονα με την υλοποίηση της πλήρους χημικής σύνθεσης των φυσικών ΟΠΙΟΕΙΔΩΝ ΠΕΠΤΙΔΙΩΝ, μελετώνται εντατικά οι διάφορες συνθετικές ενώσεις τους. ανάλογα. Ιδιαίτερη προσοχή δίνεται στη σύνθεση αναλόγων των OPIOID PEPTIDES, τα οποία έχουν αυξηθεί. αντίσταση στην πρωτεολυτική δράση. ένζυμα. Κάποια συνθετικά ανάλογα ΟΠΙΟΕΙΔΗ ΠΕΠΤΙΔΙΑ σελ. παρουσιάζουν δράση παρόμοια με τη μορφίνη σε περιφερική. χορηγείται.
Λογοτεχνία: Ενδορφίνες, εκδ. E. Costa, M. Trabucchi, μτφρ. from English, Μ., 1981; Jakubke H.-D., Eshkait X., Αμινοξέα, πεπτίδια, πρωτεΐνες, trans. from German, M., 1985, p. 289-95. Yu. P. Shvachkin.
Χημική εγκυκλοπαίδεια. Τόμος 3 >>
480 τρίψτε. | 150 UAH | $7,5 ", MOUSEOFF, FGCOLOR, "#FFFFCC",BGCOLOR, "#393939");" onMouseOut="return nd();"> Διατριβή - 480 RUR, παράδοση 10 λεπτά, όλο το εικοσιτετράωρο, επτά ημέρες την εβδομάδα και αργίες
240 τρίψτε. | 75 UAH | $3,75 ", MOUSEOFF, FGCOLOR, "#FFFFCC",BGCOLOR, "#393939");" onMouseOut="return nd();"> Περίληψη - 240 ρούβλια, παράδοση 1-3 ώρες, από 10-19 (ώρα Μόσχας), εκτός Κυριακής
Damirov Aslan Hasan ogly. Θεωρητική ανάλυση διαμόρφωσης λευκίνης-εγκεφαλίνης, Ν-τερματικού τριδεκαπεπτιδίου dynorphin και των αναλόγων τους: IL RGB OD 61:85-1/1277
Εισαγωγή
ΚΕΦΑΛΑΙΟ I. Ανασκόπηση βιβλιογραφίας 8
1.1. Πειραματικές και θεωρητικές μέθοδοι διαμορφωτικής ανάλυσης πεπτιδίων. 9
1.3. Δομική και λειτουργική οργάνωση των πεπτιδίων 29
1.4. Αντίστροφο δομικό πρόβλημα 39
1.5. Μέθοδος θεωρητικής διαμορφωτικής ανάλυσης "50
1.6. Στόχοι έρευνας 56
ΚΕΦΑΛΑΙΟ II. Θεωρητική διαμορφωτική ανάλυση Λευκίνης-Εγκεφαλίνης 57
2.1. Βιολογικές λειτουργίες λευκίνης-εγκεφαλίνης 57
2.2. Δυνατότητες διαμόρφωσης λευκίνης-εγκεφαλίνης 58
ΚΕΦΑΛΑΙΟ III. Θεωρητική ανάλυση συρροής αναλόγων λευκίνης-εγκεφαλίνης 69
3.1. Αναλογικό Tyr1- Gly2- Gly3- Phe4- Leu5- Arg6 70
3.2. Αναλογικό Ty^-D-Ala2-Gly3~ Phe4-Leu5 » 72
3.3. Αναλογικό Tyi^-D- Ala2- Gly3- Phe4- Leu5- - Arg6 79
3.4. Αναλογικό Tyr*-D- Ala2- Gly3- Phe4- Ala5- - Arg6 84
3.5. Αναλογικό TyrX-D~ Ala2- Gly3- Phe4- Gly5- - Arg6 88
3.6. Αναλογικό Tyr1-])- Ala2- Gly3- Phe4- Ala5- - Lys6 91
3.7. Αναλογικό Tyr ^D- Ala2- Gly3- Phe4- Ala5- - Asn6 93
3,8* Αναλογικό Tyg ^D*- Ala2- Gly3- Phe4-Aln5~ - Gin6 99
3.9. Αναλογικό Tyr -^- D- Ala2- Gly3- Phe4- Ala5- - His6 103
3,10* Αναλογικό Tyr -^-D- Ala2- Glu3-N- MePhe4- - Leu5- Arg6 106
ΚΕΦΑΛΑΙΟ ІУ. Θεωρητική ανάλυση διαμόρφωσης του πεπτιδίου ν-τερματικής δυνορφίνης και των αναλόγων του
4.1 Λειτουργίες δυνορφίνης και σχήμα για τον υπολογισμό της χωρικής της δομής IZ
4.2. Θραύσματα Arg - Arg7, Arg - Arg9, Arg -
Bys11, bys11-Lys13 116
4.3. Θραύσματα Phe 4-Arg7 και Arg 6-Arg9 121
4.4. Fragment Arg 9-Lys13 126
4.5. Fragment Tyr I*-Arg 9 129
4.6. Ν-τερματικό μόριο τριδεκαπεπτιδίου Dynorphin 132
4.7. - Ανάλογο Δυνορφίνης I - IЗ 137
4.8. - Ανάλογο Δυνορφίνης I - IZ 137
Λογοτεχνία 144
Εισαγωγή στην εργασία
Γενικά χαρακτηριστικά της εργασίας.Η διατριβή είναι αφιερωμένη στη μελέτη της χωρικής δομής της λευκίνης-εγκεφαλίνης, του Ν-τερματικού τριδεκαπεπτιδίου dynorphin και ορισμένων αναλόγων τους χρησιμοποιώντας τη μέθοδο της θεωρητικής διαμορφωτικής ανάλυσης. Κατά τον υπολογισμό της διαμόρφωσης των μορίων, η δυναμική ενέργεια λαμβάνεται ως το άθροισμα των ενεργειών των αλληλεπιδράσεων των ατόμων χωρίς σθένος, των ηλεκτροστατικών, των σπειροειδών αλληλεπιδράσεων και των δεσμών υδρογόνου. Για την αναζήτηση της ελάχιστης δυναμικής ενέργειας χρησιμοποιήθηκε η μέθοδος συζευγμένης κλίσης Η εργασία αναπτύχθηκε στο εργαστήριο προβλημάτων της «Μοριακής Βιοφυσικής» του Πανεπιστημίου του Αζερμπαϊτζάν Κρατικό Πανεπιστήμιοπου πήρε το όνομά του από τον S.M. Kirov, ένα καθολικό πρόγραμμα γραμμένο στην αλγοριθμική γλώσσα "FORTRAN", Οι υπολογισμοί πραγματοποιήθηκαν σε ηλεκτρονικούς υπολογιστές EC-I033, EC-I022 και BESM-6,
Συνάφεια του θέματος.Πολλές βιολογικές διεργασίες πραγματοποιούνται με την απαραίτητη συμμετοχή πρωτεϊνών και ορμονών. Η γνώση χωροταξική οργάνωσηκαι οι δυναμικές διαμορφωτικές ιδιότητες των φυσικών ολιγοπεπτιδίων είναι απαραίτητες για τη μελέτη των μοριακών μηχανισμών αναγνώρισης, διέγερσης, ρύθμισης και συντονισμού βιολογικές διεργασίες, φαινόμενα δομικών-λειτουργικών σχέσεων και λύσεις σε πολλά άλλα θέματα μεγάλης επιστημονικής και πρακτικής σημασίας. Η μέθοδος ανάλυσης περίθλασης ακτίνων Χ μπορεί να καθορίσει μόνο τις στατικές καταστάσεις των πρωτεϊνών και των ορμονών. Η θεωρητική διαμορφωτική ανάλυση δίνει μια ιδέα όχι μόνο για τη χωρική δομή, αλλά και για τις δυνατότητές της για αλλαγή. Επομένως, η εργασία προς αυτή την κατεύθυνση μπορεί να οδηγήσει σε πολύτιμες πληροφορίες σχετικά με τις λεπτές λεπτομέρειες των βιολογικών διεργασιών σε ατομικό-μοριακό επίπεδο.
Στόχος της εργασίας.Η εργασία της διατριβής είναι αφιερωμένη στη μελέτη της χωρικής δομής και των δυναμικών δομικών ιδιοτήτων των μορίων των ενδογενών πεπτιδικών ορμονών λευκίνη-εγκεφαλίνη, dynorphy-yaa, καθώς και μεγάλου αριθμού αναλόγων τους.
Δοκιμή μιας θεωρητικής προσέγγισης που καθιστά δυνατή την πρόβλεψη των δομών των αναλόγων που έχουν ορισμένες λειτουργίες φυσικών ορμονών ακόμη και πριν από τη σύνθεση και τις βιολογικές δοκιμές. Ήταν απαραίτητο να βρεθούν τέτοιες χημικές τροποποιήσεις ορμονών που θα παρατείνουν τη δράση τους ή θα αναπαράγουν επιλεκτικά μία από αυτές τις λειτουργίες. Αυτό απαιτούσε, πρώτον, να αποσαφηνιστούν τα γεωμετρικά και ενεργειακά χαρακτηριστικά των προτιμώμενων διαμορφωτικών καταστάσεων των φυσικών ορμονών (άμεσο δομικό πρόβλημα) και, δεύτερον, να τροποποιηθεί σκόπιμα η χημική δομή των φυσικών ορμονών με τέτοιο τρόπο ώστε η χωρική τους δομή να αντιστοιχεί σε καλά -καθορισμένες διαμορφώσεις χαμηλής ενέργειας του φυσικού πεπτιδίου (αντίστροφο δομικό πρόβλημα).
Επιστημονική καινοτομία.Για πρώτη φορά, οι χωρικές δομές και οι διαμορφωτικές ικανότητες της λευκίνης-εγκεφαλίνης, του ν-τερματικού τριδεκαπεπτιδίου diyaorphin και ορισμένων αναλόγων τους έχουν καθιερωθεί. Διαπιστώθηκε ότι η λευκίνη-εγκεφαλίνη μπορεί να έχει μόνο λίγες δομές χαμηλής ενέργειας ΔΙΑΦΟΡΕΤΙΚΟΙ ΤΥΠΟΙ, ο αριθμός των οποίων για τα ανάλογα του μειώνεται σημαντικά.
Οι διαμορφωτικές καταστάσεις των αναλόγων λευκίνης-εγκεφαλίνης μπορούν να χωριστούν σε δύο ομάδες: ανάλογα που υφίστανται σημαντικές δομικές αλλαγές λόγω αντικατάστασης και ανάλογα στα οποία η αντικατάσταση αμινοξέων οδηγεί σε αισθητή μετατόπιση της διαμορφωτικής ισορροπίας προς ορισμένες μορφές της φυσικής ορμόνης . Βρέθηκε ότι το Ν-τερματικό τριδεκαπεπτίδιο της δυνορφίνης έχει ένα περιορισμένο σύνολο δομικών τύπων χαμηλής ενέργειας, ο αριθμός των οποίων εξαρτάται από
φύση περιβάλλον. Έχουν βρεθεί ανάλογα που μοντελοποιούν ορισμένες λειτουργικές ιδιότητες της λευκίνης-εγκεφαλίνης και της δυνορφίνης.
Πρακτική αξία.Με βάση την αλληλουχία αμινοξέων λευκίνης-εγκεφαλίνης και δυνορφίνης Ι-Ι3, καθορίστηκαν οι διαμορφωτικές ικανότητες και οι τρισδιάστατες δομές των μορίων, η γνώση των οποίων είναι απαραίτητη για τη μελέτη της βιολογικής δραστηριότητας και του μηχανισμού λειτουργίας αυτών των μορίων. ως στοχευμένη αναζήτηση αναλόγων.
Δοκιμασμένο θεωρητική προσέγγισηκαι μελετήθηκε η χωρική δομή των αναλόγων. Η έρευνα είναι χρήσιμη για τη στοχευμένη σύνθεση αναλόγων αυτών των μορίων με συγκεκριμένες λειτουργίες. Οι υπολογισμοί έδειξαν τη δυνατότητα πρόβλεψης της γεωμετρίας και των διαμορφωτικών ικανοτήτων των ορμονών και της χημικής δομής των αναλόγων τους με γνωστές ιδιότητες χρησιμοποιώντας τη μέθοδο της θεωρητικής διαμορφωτικής ανάλυσης.
Οι κύριες διατάξεις για την άμυνα: I. Αποτελέσματα υπολογισμού της χωρικής δομής λευκίνης-εγκεφαλίνης H - Tyr 1 - Gly 2 - Gly 5 - Phe^~ Leu^-OH Και τα ακόλουθα ανάλογα IGO: H-Tyr 1 -Gly 2 -Gly 5 -Phe 4 -Leu 5 -Arg 6-OH,
H-Tyr 1 -D-Ala 2 -Gly 5 -Phe 4 -Leu 5 -OH,
H-Tyr 1 -D-Ala 2 -Gly 5 -Phe 4 -Leu 5 -Arg 6 -OH,
H-Tyr 1 -D-Al a 2 -Gly 3 -Phe 4 -Al a 5 -Arg 6 -OH,
H-Tyr 1 -D-Ala 2 -Gly 3 -Phe 4 -Ala 5 -Ly s 6 ~0H,
H-Tyr 1 -D-Ala 2 -Gly 5 -Phe 4 -Ala 5 -Asn 6 -OH,
H-Tyr 1 -D-Al a 2 -Gly 5 -Phe Z| "-Gly 5 -Arg 6 -OH,
H-Tyr 1 -D-Ala 2 -Gly 5 -Phe 4 -Ala 5 -Gln 6 -OH,
H-Tyr 1 -D-Ala 2 -Gly5-phe^-Ala5-His 6-OH,
H-Tyr 1 -D-Ala 2 -Gly 5 -N-MePhe 4 -Leu 5 -Arg 6 -OH.
2. Αποτελέσματα υπολογισμού της χωρικής δομής της δυνορφίνης
I-IЗ N -Tyr 1 -Gly 2 -Gly 5 -Phe 4 -Leu 5 -Arg 6 -Arg 7 -lie 8 -Arg 9 -Pro 10 -Lys 11 -Leu 12 -Lys 15 -on και τα ακόλουθα ανάλογα: [ъ-
Ala 2] - και (b -Ala 12 ] -δυνορφίνη.
Συζήτηση της σχέσης μεταξύ χημικής δομής και χωρικής διάταξης των θεωρούμενων φυσικών και συνθετικών αλληλουχιών αμινοξέων,
Στοχευμένη αναζήτηση για χημικές τροποποιήσεις των μορίων λευκίνης-εγκεφαλίνης και δυνορφίνης, πιο σταθερές και αναπαράγοντας τις βιολογικές λειτουργίες των φυσικών πεπτιδίων.
Έγκριση εργασιών.Τα αποτελέσματα της εργασίας παρουσιάστηκαν στο 1st All-Union Symposium on Intermolecular Interactions and Conformations of Molecules (Βίλνιους, 1982), στο 1ο All-Union Biophysical Congress (Μόσχα, 1982), σε ένα επιστημονικό συνέδριο αφιερωμένο στα αποτελέσματα του ερευνητική εργασία για το 1983 ( Baku, 1984"), στο Ρεπουμπλικανικό Συνέδριο Νέων Επιστημόνων για τη Φυσική και Χημική Βιολογία (Baku, I98I), καθώς και σε σεμινάρια του Τμήματος Οπτικής και μοριακή φυσικήκαι το εργαστήριο προβλημάτων «Μοριακής Βιοφυσικής».
Δημοσίευση.Το κύριο περιεχόμενο της διατριβής παρουσιάζεται σε επτά δημοσιεύσεις.
Δομή και αντικείμενο της διπλωματικής εργασίας.Η διατριβή παρουσιάζεται σε 159 σελίδες δακτυλόγραφου κειμένου, αποτελείται από μια εισαγωγή, τέσσερα κεφάλαια και συμπεράσματα, περιέχει 10 σχήματα, 35 πίνακες και κατάλογο της αναφερόμενης βιβλιογραφίας, συμπεριλαμβανομένων 135 τίτλων.
Άμεσο δομικό πρόβλημα
Με τον όρο άμεσο δομικό πρόβλημα εννοούμε την καθιέρωση πληροφοριακών χαρακτηριστικών ενός μορίου με βάση αυτό χημική δομή. Η λύση σε αυτό το πρόβλημα για τα φυσικά ολιγοπεπτίδια δίνεται στον πίνακα. Φτιάχτηκα με βάση τις ακόλουθες υποθέσεις.
Η αλληλουχία αμινοξέων καθορίζει πλήρως τη χωρική δομή του μορίου του ολιγοπεπτιδίου. 2. Οι διαμορφωτικές καταστάσεις των ολιγοπεπτιδίων υπό φυσιολογικές συνθήκες έχουν ελάχιστες δυναμική ενέργεια. 3.Χωρική δομήτα φυσικά ολιγοπεπτίδια αντιστοιχούν στην αυστηρότερη συσκευασία της αλληλουχίας αμινοξέων. 4. Στις συμπαγείς δομές των ολιγοπεπτιδίων υπάρχει συνέπεια μεταξύ όλων των ενδομοριακών αλληλεπιδράσεων μικρής, μεσαίας και μεγάλης εμβέλειας.
Με τον όρο βραχυπρόθεσμες εννοούμε τις αλληλεπιδράσεις της πλευρικής αλυσίδας ενός δεδομένου υπολείμματος με δύο γειτονικές πεπτιδικές ομάδες. κάτω από τον μέσο όρο - αλληλεπιδράσεις του υπολείμματος με τέσσερις γείτονες και στις δύο πλευρές στην ακολουθία. κάτω από απομακρυσμένες - αλληλεπιδράσεις με υπολείμματα πιο απομακρυσμένα κατά μήκος της αλυσίδας (Εικ. Ι). Οι αλληλεπιδράσεις μικρής εμβέλειας καθορίζουν τις διαμορφωτικές ικανότητες των ελεύθερων μονοπεπτιδίων. Μια ποσοτική ιδέα αυτού του συστατικού μπορεί να ληφθεί με ανάλυση διαμόρφωσης των αντίστοιχων μεθυλαμιδίων Ν-ακετυλ-1-αμινοξέων. Επί του παρόντος, και για τα 20 τυπικά α-αμινοξέα, έχουν ληφθεί σύνολα βέλτιστων διαμορφώσεων που προτιμώνται για αλληλεπιδράσεις μικρής εμβέλειας, κατάλληλες για τον υπολογισμό οποιασδήποτε αλληλουχίας αμινοξέων. Το Σχήμα 2α δείχνει το δυναμικό εμβαδόν επιφάνειας του μορίου μεθυλαμιδίου Ν-ακετυλ-1-αλανίνης ως συνάρτηση των διεδρικών γωνιών y και t/g. Η επιφάνεια έχει τέσσερις περιοχές χαμηλής ενέργειας B, H, b και P. Οι χάρτες μη σχηματισμού του tr-\jr y b 20 τυπικών αμινοξέων, οι πλευρικές αλυσίδες των οποίων δεν περιέχουν κλάδο στο άτομο C, είναι πολύ κοντά. Οι πιθανές επιφάνειες των μεθυλαμιδίων Ν-ακετυλο-L-βαλίνης και β-ισολευκίνης έχουν τις ίδιες, αλλά ελαφρώς πιο στενές, περιοχές χαμηλής ενέργειας Ο χάρτης -t του μεθυλαμιδίου W-ακετυλο-γλυκίνης είναι κεντροσυμμετρικός και περιέχει εκτεταμένες και πρακτικά ισοενεργητικές περιοχές R(b) και B (P), εικ. 3. Η πιθανή επιφάνεια του μορίου μεθυλαμιδίου Ν-ακετυλ-β-προλίνης έχει δύο ελάχιστα, που αντιστοιχούν στις τιμές της γωνίας "r -60 (R.) και -140 (B), Εικ. 4.
Θεωρητικές προβλέψεις σχετικά με τις δυνατότητες πληροφόρησης και τη θέση της διαμορφωτικής ισορροπίας των μεθυλαμιδίων Ν-ακετυλ-ο-αμινοξέων σε πολικά και μη πολικά μέσα αντιστοιχούν σε πειραματικά δεδομένα που ελήφθησαν για έναν αριθμό από αυτές τις ενώσεις χρησιμοποιώντας φασματοσκοπία NMR, ODS, CD και IR μεθόδους, καθώς και διπολικές ροπές και ωσμομετρία αερίου-υγρού \p! 65]. Ταυτόχρονα, η σύγκριση των αποτελεσμάτων υπολογισμού αυτών των μορίων με τη γεωμετρία των υπολειμμάτων αμινοξέων σε πρωτεΐνες γνωστής τρισδιάστατης δομής έδειξε ότι οι πειραματικές τιμές των γωνιών tp, J\T της κύριας αλυσίδας όλων των υπολειμμάτων βρίσκονται μόνο σε περιοχές χαμηλής ενέργειας (0 - 4,0 kcad/mol) των χαρτών διαμόρφωσης t / - t/g ελεύθερων μονοπεπτιδίων. Υπάρχει επίσης μια ικανοποιητική αντιστοιχία μεταξύ των διατομών των μεθυλαμιδίων της δυνητικής επιφάνειας Y f , Y "I των Ν-ακετυλο-ο-αμινοξέων και των καταστάσεων διαμόρφωσης των πλευρικών αλυσίδων στην τρισδιάστατη δομή της πρωτεΐνης [bb - 68]. Ανίχνευση συσχέτισης μεταξύ της γεωμετρίας όλων των υπολειμμάτων αμινοξέων σε φυσικές αλληλουχίες και των καταστάσεων χαμηλής ενέργειας ελεύθερων μονοπεπτιδίων Η παρουσία συσχέτισης έδειξε ότι οι αλληλεπιδράσεις μικρής εμβέλειας καθορίζουν για όλα τα τυπικά υπολείμματα τα σύνολα των επιτρεπόμενων χωρικών μορφών. αναδιπλώνοντας την πεπτιδική αλυσίδα στη φυσική δομή, μόνο από τέτοια σύνολα εμφανίζεται μια επιλογή (πρώτα υπό την επίδραση αλληλεπιδράσεων μεσαίου εμβέλειας και μετά μεγάλης εμβέλειας) διαμορφωτικές καταστάσεις όλων των υπολειμμάτων. Αποδεικνύονται μόνο εκείνες οι διαμορφώσεις υπολειμμάτων που είναι προτιμότερες για αλληλεπιδράσεις μικρής εμβέλειας να είναι πραγματικό, ο συνδυασμός των οποίων σε μια δεδομένη αλληλουχία αμινοξέων εξασφαλίζει το σχηματισμό των πιο ευνοϊκών σταθεροποιητικών επαφών υπολειμμάτων.Έτσι, η αλληλουχία των καταστάσεων διαμόρφωσης των υπολειμμάτων και όχι οι ίδιες οι καταστάσεις. Η κατάσταση εδώ είναι, κατ' αρχήν, παρόμοια με τη χημική δομή των φυσικών πεπτιδίων.
Λόγω του τεράστιου αριθμού αρχικών παραλλαγών που μπορούν να αποτελούνται από διαμορφώσεις υπολειμμάτων χαμηλής ενέργειας, μια άμεση λύση στο άμεσο δομικό πρόβλημα είναι δυνατή μόνο για σύντομες ακολουθίες που δεν περιέχουν περισσότερες από 4-5 μονάδες. Για πεπτίδια υψηλότερου μοριακού βάρους, ο στόχος μπορεί να επιτευχθεί μόνο με τη διαίρεση ολόκληρου του προβλήματος σε έναν αριθμό λιγότερο επαχθών εργασιών και τη διαδοχική επίλυσή τους. Η ικανότητα χρήσης μιας προσέγγισης βήμα προς βήμα οφείλεται στην ύπαρξη συνέπειας μεταξύ όλων των αλληλεπιδράσεων μικρής, μεσαίας και μεγάλης εμβέλειας. Σε αυτή τη βάση, ελήφθησαν λύσεις σε μεγάλο αριθμό άμεσων δομικών προβλημάτων για πολύ πολύπλοκα ολιγοπεπτίδια και πρωτεΐνες χαμηλού μοριακού βάρους.
Δυνατότητες διαμόρφωσης λευκίνης-εγκεφαλίνης
Κατά την αντικατάσταση του τέταρτου υπολείμματος των πεπτιδίων BPPk και N -MeAla 4 BPPc OOOTBeTCTBeHHO Ha D -Ala - I D -N CHieAIA - BPPg ΟΛΕΣ οι περιηγήσεις CTpyK που επιτρέπονται για μόρια με τη διαμόρφωση b θα απαγορεύονται για μόρια με διαφορετική διαμόρφωση. Επομένως, τα συνθετικά ανάλογα του D-Ala4-BPPg και του D-N-MeAia 4 BPPg δεν θα έχουν τις φυσιολογικές ιδιότητες της φυσικής ένωσης. Ένας εναλλακτικός περιορισμός στις δομές ισχύει όταν το β-κατάλοιπο που αντικαθίσταται προηγείται μιας προλίνης. Αλλά ακόμα κι αν δεν πληρούται αυτός ο κανόνας, η συμπερίληψη ενός D-αμινοξέος, ειδικά ενός υποκατεστημένου σε άζωτο, στην β-αλληλουχία μπορεί να είναι η πιο αποτελεσματικός τρόποςεπίλυση του αντίστροφου δομικού προβλήματος. Αυτό θα εξαρτηθεί από τη συγκεκριμένη αλληλουχία, τις διαμορφωτικές ικανότητες του φυσικού πεπτιδίου και τον σκοπό της μελέτης. Έτσι, η αντικατάσταση του υπολείμματος β που βρίσκεται στην κατάσταση Β στη δομή ενδιαφέροντος με ένα υπόλειμμα D θα οδηγεί πάντα στην απαγόρευσή του. Εάν το αντικατασταθέν υπόλειμμα έχει τη μορφή L της κύριας αλυσίδας, τότε με τη συμπερίληψη ενός υπολείμματος D, η πιθανότητα πραγματοποίησης της αντίστοιχης διαμορφωτικής κατάστασης αυξάνεται, καθώς αυτό μειώνει την ενέργεια των αλληλεπιδράσεων μικρής εμβέλειας αυτού του υπολείμματος και επεκτείνει το εύρος των επιτρεπόμενων τιμών των γωνιών του Y, Tz (Εικ. 2), που μπορεί να οδηγήσουν στη δημιουργία πιο αποτελεσματικών επαφών μεταξύ υπολειμμάτων. Μεγάλες ευκαιρίες για στοχευμένο προσδιορισμό της δομής ιόντων ανοίγονται κατά την αντικατάσταση της γλυκίνης με β- και D-αλανίνη. Κατά την πρώτη αντικατάσταση, όλες οι διαμορφώσεις στις οποίες οι γωνίες p, ar της γλυκίνης πέφτουν στην περιοχή P θα είναι απαγορευμένες και οι διαμορφώσεις με τις γωνίες p, y γλυκίνης στην περιοχή b θα γίνουν λιγότερο πιθανές. Εάν το Giy αντικατασταθεί από το D-Ala, οι διαμορφώσεις με τη μορφή Β της γλυκίνης θα είναι αδύνατες και λιγότερο πιθανές με τη μορφή Β. Επιπλέον, η συμπερίληψη ενός υπολείμματος D στην αλυσίδα θα επηρεάσει τη σχέση του πεπτιδίου με τις πεπτιδάσες και θα οδηγήσει σε παράταση της δράσης του.
Η λύση στο αντίστροφο δομικό πρόβλημα της γιαοναπεπτιδικής ορμόνης ύπνου θα βασίζεται επίσης στο γνωστό σύνολο όλων των διαμορφώσεων χαμηλής ενέργειας ενός φυσικού μορίου και στις απλούστερες και φυσικά απαντώμενες χημικές τροποποιήσεις των αλληλουχιών αμινοξέων που αναφέρονται παραπάνω.
Μέχρι τώρα, κατά την επίλυση του αντίστροφου δομικού προβλήματος, χρησιμοποιήθηκαν υποκαταστάσεις αμινοξέων των οποίων η επίδραση στις διαμορφωτικές ικανότητες του μορίου μπορούσε εύκολα να εκτιμηθεί a priori, τουλάχιστον στο επίπεδο του σχήματος της κύριας αλυσίδας και του σχήματος. Απαιτήθηκε αυστηρή ποσοτική ανάλυση μόνο για τον προσδιορισμό της σειράς διάταξης των καταστάσεων διαμόρφωσης σύμφωνα με τις ενεργειακές τιμές σε ένα στενό εύρος επιλεγμένων επιλογών. Ωστόσο, όλες οι υποκαταστάσεις που δεν περιλαμβάνουν υπολείμματα γλυκίνης και προλίνης δεν οδηγούν σε εκ των προτέρων αποκλεισμό οποιωνδήποτε μορφών και σχημάτων του πεπτιδικού οκελέτου. Η επίδραση τέτοιων αντικαταστάσεων στις δυνατότητες διαμόρφωσης δεν εξαρτάται τόσο από αλληλεπιδράσεις μικρής εμβέλειας, αλλά από αλληλεπιδράσεις μεσαίου και μεγάλου βεληνεκούς, δηλ. από ένα πολύ περίπλοκο σύστημα ενδομοριακής επαφής, που προσδιορίζεται όχι από μεμονωμένα υπολείμματα, αλλά από ολόκληρη την αλληλουχία. Δεδομένου ότι σε κάθε περίπτωση η κατάσταση είναι μοναδική, είναι ακόμα δυνατό να εντοπιστούν οι συνέπειες των αντικαταστάσεων ακόμη και σε σχέση με μορφές και σχήματα μόνο με υπολογισμό. Το επόμενο αντικείμενο εξέτασης είναι ένα ανάλογο ενός α-πεπτιδίου στο οποίο τα υπολείμματα Ala 2 και Gly 8 αντικαθίστανται από Vai 8. Θα μπορούσε να αναμένεται ότι η συμπερίληψη στην αλληλουχία δύο υπολειμμάτων με ογκώδεις και άκαμπτες πλευρικές αλυσίδες θα αλλάξει σημαντικά την κατανομή ενέργειας της διαμόρφωσης του φυσικού εννεαπεπτιδίου. Ωστόσο, πριν από τον υπολογισμό ήταν δύσκολο να το υποθέσουμε παγκόσμια δομήΗ τροποποιημένη αλληλουχία θα είναι η διαμόρφωση HS, η οποία έχει την υψηλότερη ενέργεια στη σειρά που εκχωρείται για το 8-πεπτίδιο. Αποδείχθηκε ότι και τα δύο υπολείμματα βαλίνης ταιριάζουν καλά μόνο στη δομή των ΗΠΑ, χωρίς να αλλάξουν τη γεωμετρία τους και χωρίς να καταστρέψουν το υπάρχον σύστημα αλληλεπιδράσεων. Η σταθερότητα αυτής της δομής στο ανάλογο Vai » αυξάνεται σε σύγκριση με το πεπτίδιο 8 κατά 8,0 kcal/mol.
Η επόμενη τροποποίηση της &-πεπτιδικής αλληλουχίας - σημείωση από Asp σε Asn, είναι η πιο ασήμαντη στερικά, αλλά πολύ σημαντική ηλεκτρονικά. Καθιστά πραγματικές δομές μόνο ενός τύπου eefeefe για το πεπτίδιο Asn 5 -S, μεταξύ των οποίων η διαμόρφωση Pa είναι κυρίαρχη. Όπως δείχνουν οι υπολογισμοί, σε αυτή την περίπτωση οι αλλαγές αφορούν μόνο δύο αλληλεπιδράσεις μεταξύ υπολειμμάτων με το ΥΤ, δηλαδή τις επαφές του Asn με το Gin και με το Trp. Η απουσία αρνητικού φορτίου στο υπόλειμμα στην πέμπτη θέση, η οποία είναι κεντρική στη δομή Pa, αφαιρεί την ηλεκτροστατική απώθηση μεταξύ των υπολειμμάτων του πρώτου ζεύγους (περίπου +6,8 kcal/mol για το φυσικό πεπτίδιο έως +0,2 για το Asn-αναλογικό). Ταυτόχρονα, όμως, εξαφανίζεται η σταθεροποιητική πολική αλληλεπίδραση μεταξύ των υπολειμμάτων του δεύτερου ζεύγους, η οποία, ωστόσο, στο S-πεπτίδιο στη διαμόρφωση PA είναι πολύ λιγότερο αποτελεσματική (-3,0 kcal/mol).
Αναλογικό Tyi^-D- Ala2- Gly3- Phe4- Leu5- - Arg6
Οι διαμορφώσεις χαμηλής ενέργειας του [p-Ala 2]-LEK που λάβαμε δίνονται στον Πίνακα. 9, τα ενεργειακά χαρακτηριστικά της πιο σταθερής διαμόρφωσης κάθε μορφής της κύριας αλυσίδας -LEK φαίνονται στον πίνακα. 10. Τα αποτελέσματα του υπολογισμού έδειξαν ότι στην ακολουθία LEC, η αντικατάσταση του Gly fc με το D-Alar οδηγεί σε ισχυρή ενεργειακή διαφοροποίηση της διαμόρφωσης, των μορφών και των σχημάτων της κύριας αλυσίδας. Από τους 16 πιθανούς τύπους δομών, μόνο τρεις διαμορφώσεις, δηλαδή ffff, fffe και efff, αποδείχθηκαν χαμηλής ενέργειας (Πίνακας 9). Σε αντίθεση με το μόριο LEC, οι διαμορφώσεις του σχήματος effe για το ανάλογό του είναι υψηλής ενέργειας. Η σχετική τους ενέργεια αυξήθηκε κατά μέσο όρο 3,0 kcal/mol σε σύγκριση με τις διαμορφώσεις του ίδιου σχήματος του μορίου LEK. Αυτό οφείλεται προφανώς στο γεγονός ότι σε διαμορφώσεις χαμηλής ενέργειας του σχήματος efff, το δεύτερο υπόλειμμα είναι στη μορφή ραχοκοκαλιάς Β, η οποία είναι λιγότερο ευνοημένη για το D-στερεοϊσομερές.
Η συνολική δομή του αναλόγου LEC είναι σε LP % L32 του σχήματος ffff σε αυτή τη διαμόρφωση, οι αλληλεπιδράσεις διπεπτιδίου συνεισφέρουν -9,2 kcal/mol, το τριπεπτίδιο -1,7 kcal/mol, το τετραπεπτίδιο-nne -3,3 kcal/mol και το πενταπεπτίδιο -10,1 kcal/mol mol. Είναι το τελευταίο στοιχείο που κάνει αυτή τη διαμόρφωση την πιο συμφέρουσα. Η ενέργεια της διαμόρφωσης B2PHB2B32 του σχήματος fffe είναι μόνο 0,1 kcal/mol υψηλότερη από την ενέργεια της συνολικής διαμόρφωσης. Εδώ, η αλληλεπίδραση πενταπεπτιδίου μεταξύ των υπολειμμάτων Tyr και Leu είναι πιο εξασθενημένη και οι αλληλεπιδράσεις τρι- και τετραπεπτιδίου είναι ισχυρότερες. Πλέον τεράστια συμβολήαλληλεπιδράσεις διασποράς (-20,7 kcal/mol) συμβάλλουν στη σταθεροποίηση αυτής της διαμόρφωσης. της οποίας η ονομαστική ενέργεια είναι 3,0 kcal/mol. Με βάση τα αποτελέσματα υπολογισμού, μπορεί να υποτεθεί ότι, υπό φυσιολογικές συνθήκες, το ανάλογο D-Ala-LEK δεν θα εκτελεί όλες τις λειτουργίες που είναι εγγενείς στο φυσικό μόριο LEC. Η λειτουργία του μορίου, η υλοποίηση του οποίου απαιτεί μια δομή LEC του τύπου effe, θα γίνει απραγματοποίητη. Έτσι, τα αποτελέσματα της διαμορφωτικής ανάλυσης του αναλόγου Leutsch-εγκεφαλίνης καθιστούν δυνατό να γίνουν οι ακόλουθες υποθέσεις - πρώτον, αυτό το ανάλογο δεν θα μπορεί να εκτελέσει όλες τις λειτουργίες του LEC, αλλά μόνο εκείνες στις οποίες η χωρική του δομή θα πρέπει να είναι ffff , τύπους fffe και εν μέρει efff, ΚΑΙ δεύτερον, λόγω αντικατάστασης; 2 του υπολείμματος αμινοξέος Gly στο D - Ala, το ανάλογο μόριο γίνεται πιο ανθεκτικό στη δράση των αποικοδομητικών ενζύμων, γεγονός που επιβραδύνει την ενζυματική υδρόλυση αυτού του αναλόγου στο σώμα και, ως εκ τούτου, παρατείνει τη δράση του." Analogue Tyr I- D- Ala2- Gly3- Phe4- Leu5-Arg 6.
Η χωρική δομή αυτού του αναλόγου του μορίου LEC μελετήθηκε με βάση τις βέλτιστες διαμορφώσεις των δύο άλλων αναλόγων του - και -LEK, που συζητήθηκαν παραπάνω. Οι προκύπτουσες διαμορφώσεις χαμηλής ενέργειας του αναλόγου -LEK φαίνονται στον Πίνακα. II, και τα ενεργειακά χαρακτηριστικά της πιο σταθερής διαμόρφωσης κάθε μορφής της κύριας αλυσίδας είναι στον Πίνακα. 12.
Η σχετική ενέργεια των 28 διαμορφώσεων αυτού του αναλόγου είναι κάτω από 5,0 kcal/mol. Όλα ανήκουν σε επτά διαφορετικές μορφές της κύριας αλυσίδας, πέντε σχήματα. Οι παραλλαγές χαμηλής ενέργειας του αναλόγου -LEK που ελήφθησαν ως βάση για τον υπολογισμό αποδείχθηκαν ελπιδοφόρες και διατηρήθηκαν για το ανάλογο p-Ala2, Arg 6]. Τα σχήματα χαμηλής ενέργειας της κύριας αλυσίδας του αναλόγου -LEK, που ελήφθησαν ως αποτέλεσμα της διαμορφωτικής του ανάλυσης, φαίνονται στο Σχ. 8.
Η σχετική ενέργεια της διαμόρφωσης B2PRR2B21B5522 του σχήματος ffffe είναι 0,9 kcal/mol Σε αυτή τη διαμόρφωση, η συμβολή των αλληλεπιδράσεων δι-τρι-, τετρα- και πενταπεπτιδίου, αντίστοιχα, είναι -10,0, -3,9, -5,1 και -7,0 kcal / ΕΛΙΑ δερματος. Όπως φαίνεται, οι δύο πρώτες αλληλεπιδράσεις σε αυτή τη διαμόρφωση είναι πιο αδύναμες από αυτές της παγκόσμιας και η αλληλεπίδραση εξα-πεπτιδίου είναι πολύ μικρή,
Στη διαμόρφωση %REB3B32%22 του σχήματος fffee, η σχετική ενέργεια είναι 3,2 kcal/mol.Σε αυτή τη διαμόρφωση έχω ένα σταθεροποιητικό αποτέλεσμα! κυρίως αλληλεπιδράσεις δι- και πενταπεπτιδίου. Είναι ίσα με -8,4 και -9,3 kcal/mol, αντίστοιχα. Γενικά, οι μη σθένες αλληλεπιδράσεις έχουν ισχυρό σταθεροποιητικό αποτέλεσμα, περίπου -29,0 kcal/mol,
Έτσι, μπορεί να υποτεθεί ότι το μόριο -LEK μπορεί να διατηρήσει μόνο μερικές από τις βιολογικές λειτουργίες της φυσικής ορμόνης. Αλλά η επίδρασή του στο σώμα μάλλον θα είναι παρατεταμένη.Το ανάλογο του Tyr είναι το D-Ala 2-Gly 3-Phe 4-A1a 5-Arg 6.
Δεδομένου ότι η χωρική δομή αυτού του αναλόγου μελετήθηκε με βάση τα αποτελέσματα των υπολογισμών του μορίου [n-A1a 2t Arg 6]-LEK που συζητήθηκε παραπάνω, οι διαμορφώσεις μόνο των πέντε καλύτερων σχημάτων του πεπτιδικού σκελετού υπολογίστηκαν για αυτό το μόριο, συγκεκριμένα : fffef, ffffe, effff, efffe και fffee ,Οι προτιμώμενες διαμορφώσεις που βρέθηκαν φαίνονται στον Πίνακα. 13, και τα ενεργειακά χαρακτηριστικά των πιο σταθερών διαμορφώσεων κάθε θεωρούμενης μορφής της κύριας αλυσίδας δίνονται στον Πίνακα. 14
Θραύσματα Phe 4-Arg7 και Arg 6-Arg9
Αναγνωρίστηκε για πρώτη φορά από τους Goldstein et al. [І27І από εκχύλισμα υπόφυσης χοίρου, το Ν-τερματικό τριδεκαπεπτίδιο της δυνορφίνης έχει τη βιολογική δραστηριότητα ολόκληρου του πεπτιδίου, η αλληλουχία της δυνορφίνης (I-IZ) περιλαμβάνει λευκίνη-εγκεφαλίνη. Η αλληλουχία αμινοξέων της δυνορφίνης προσδιορίστηκε με συνδυασμό της αυτόματης μεθόδου Edman και υδρόλυσης με καρβοξυπεπτιδάσες Α και Υ\l28j. Υποτίθεται ότι η δυνορφίνη είναι ο πρόδρομος των δύο πεπτιδίων ΙΑ (Ν-τερματικό 17-μελές θραύσμα) και IB (C-τερματικό τεμάχιο 13-μελών), που σχηματίζονται ως αποτέλεσμα ενζυματικής υδρόλυσης πεπτιδικός δεσμός Lyz - Arg 19.
Η Dynorphin διαφέρει από άλλα πεπτίδια οπιούχων στο ότι αλληλεπιδρά με τους υποδοχείς οπιούχων στο παρασκεύασμα του ειλεού του ινδικού χοιριδίου, 50 φορές πιο ισχυρή από την π-ενδορφίνη, 200 φορές πιο ισχυρή από τη μορφίνη και 700 φορές πιο ισχυρή από την LEC. Η δυνορφίνη μπορεί να παίζει ρόλο στη ρύθμιση των αποκρίσεων σε αλλαγές στον μεταβολισμό νερού-αλατιού, καθώς και να συμμετέχει σε διαδικασίες που σχετίζονται με τον τοκετό και τη σίτιση Η βιολογική δραστηριότητα της δυνορφίνης ήταν ανθεκτική στην καταστροφή από το βρωμιούχο κυανογόνο.
Η δυνορφίνη έχει μοναδική επίδραση στην αναλγησία που προκαλείται σε ποντίκια από οπιοειδή όπως η μορφίνη, η β-ενδορφίνη (BER) και τα ανάλογα λευκίνης-ανκεφαλίνης. Αποδυναμώνει την αναλυτική δράση της μορφίνης, ενισχύει την αναλγητική δράση μεγάλων δόσεων BER, χωρίς να αποδυναμώνει την επίδραση μικρών δόσεων BER, η Dynorphin (I - 13) δεν προκαλεί αναλγησία. Υποτίθεται ότι η δυνορφίνη έχει μια ρυθμιστική επίδραση στην αναλυτική επίδραση της μορφίνης, του BER και ορισμένων άλλων ενδογενών οπιοειδών.
Η δυνορφίνη επηρεάζει επίσης τη συμπεριφορά των ποντικών, αυξάνοντας το αίσθημα πείνας όταν χορηγείται κεντρικά στον εγκέφαλο, ωστόσο, αυτές οι επιδράσεις δεν εξαλείφονται με το nalakoon # Η δράση της δυνορφίνης (I-I3) εμποδίζεται πλήρως από το nalakoon, αλλά χρειάζεται 13 φορές περισσότερο για την καταστολή της δραστηριότητας της δυνορφίνης (I-I3) της υψηλότερης συγκέντρωσής της παρά για την καταστολή της δράσης της LEC ή της νορμορφίνης. Υψηλά καθαρισμένες μεμβράνες εγκεφάλου ποντικού αποικοδομούν γρήγορα τη δυνορφίνη (I-I3), υποδεικνύοντας την παρουσία μιας αντίστοιχης πρωτεάσης στις μεμβράνες του εγκεφάλου».
Kazarossian et al. Ένας εξαιρετικά ευαίσθητος ειδικός ποσοτικός αντιορός (Ac) ελήφθη στο σύζευγμα του βιολογικά ενεργού θραύσματος Ι-Ι3 της δυνορφίνης.» Το φάρμακο Ac δεν αντιδρά με το LEC που περιλαμβάνεται στη δομή της δυνορφίνης (I-I3). Μια μελέτη της διασταυρούμενης αντίδρασης του Ac με περικομμένα θραύσματα δυνορφίνης (Ι-Ι3) έδειξε ότι το καρβοξυτελικό υπόλειμμα λυσίνης-13 και η ελεύθερη αμινομάδα Tyr δεν είναι απαραίτητα για την ανοσοαντιδραστικότητα της δυνορφίνης (Ι-Ι3). Ταυτόχρονα, η διαδοχική βράχυνση της δυνορφίνης (I-I3) από το C-άκρο, ξεκινώντας από το 12ο υπόλειμμα, οδηγεί σε σταδιακή μείωση της ανοσοαντιδραστικότητας, η οποία γίνεται εξαιρετικά χαμηλή μετά την αφαίρεση του 9ου υπολείμματος και πρακτικά απουσιάζει δυνορφίνη (1-5), εκείνα. λευκίνη-εγκεφαλίνη Η ίδια εργασία έδειξε ότι η αντικατάσταση του Gly με β-Ala2 στη δυνορφίνη (I-I3) οδηγεί σε μείωση της ανοσοαντιδραστικότητας.
Οι Chavkin και Goldstein [іЗЗ] μελέτησαν διαφορετικά μέρη του μορίου της δυνορφίνης (I-ІЗ) στην αλληλεπίδρασή του με τους υποδοχείς του νευρικού πλέγματος των μυών του χοίρου. Η αφαίρεση αμινοξέων από το C-άκρο της δυνορφίνης (Ι-Ι3) έδειξε ότι η λυσίνη-13, η λυσίνη-Η και η αργινίνη-7 απαιτούνται προφανώς για να διατηρηθεί ένα υψηλό επίπεδο αλληλεπίδρασης του πεπτιδίου με τους υποδοχείς. Η αφαίρεση της Ν-τερματικής τυροσίνης οδήγησε στην απώλεια της βιολογικής δραστηριότητας του πεπτιδίου." Οι συγγραφείς \_133" προτείνουν ότι τα TT 7 Lys και Arg απαιτούνται προφανώς για την ειδικότητα της αλληλεπίδρασης με τους υποδοχείς δυνορφίνης (I-I3). που διαφέρουν από τους υποδοχείς mu στον ίδιο ιστό Wüster et al. χορηγήθηκε σε ποντίκια ταυτόχρονα για 6 ημέρες -LEK (I), ως αγωνιστής υποδοχέα δέλτα-οπιοειδών (-0P) και σουφεντανίλη (P) ως αγωνιστής ja-0P Κατά τη σύγκριση των οπιοειδών δράσεων της δυνορφίνης (I-I3), -LEK, LEC, σουφεντανίλης κ.λπ. στο φάρμακο vas deferens σε συγκεντρώσεις 140 - 2500 φορές μεγαλύτερες για ποντίκια ανεκτικά στο I + P από ό,τι για τα ποντίκια ελέγχου, βρέθηκε ότι η δραστηριότητα της δυνορφίνης (I-I3) μειώθηκε ελαφρά. Σε αυτή τη βάση, οι συγγραφείς προτείνουν ότι υπάρχουν υποδοχείς οπιούχων ειδικών για τη δυνορφίνη (I-I3) και λειτουργούν ανεξάρτητα από τα f-- και o 0P. Έτσι, η δυνορφίνη (I- I3) και LEC αλληλεπιδρούν με διαφορετικούς πληθυσμούς υποδοχέων οπιούχων.
Η δομή της δυνορφίνης (I-13) στο νερό και σε άλλες ουσίες μελετήθηκε χρησιμοποιώντας φασματοσκοπικές μεθόδους Φάσματα CD υδατικά διαλύματαΗ δυνορφίνη υποδηλώνει τη διαταραχή και την πολύ ασταθή δομή της. Μαζί με την πολυλειτουργικότητα, η δυνορφίνη (I-I3) έχει υψηλή ευαισθησία και ειδικότητα κάθε δράσης της. Και αυτό προφανώς οφείλεται στο γεγονός ότι το μόριο της δυνορφίνης (I-I3) υπό φυσιολογικές συνθήκες δεν είναι στατιστικά κυμαινόμενο πηνίο, αλλά έχει ένα πολύ συγκεκριμένο σύνολο ισοενεργειακών διαμορφώσεων διάφοροι τύποιΘεωρώντας μεγάλος αριθμόςλειτουργίες που εκτελούνται από τη δινορφίνη, μπορεί κανείς να αναμένει ότι αυτό το σετ είναι πολύ αντιπροσωπευτικό. Για τη μελέτη της δομικής και λειτουργικής οργάνωσης ενός ολιγοπεπτιδίου σε ατομικό-μοριακό επίπεδο, είναι απαραίτητο να γνωρίζουμε το πλήρες σύνολο των χαμηλής ενέργειας και φυσιολογικά «ενεργών» διαμορφωτικών καταστάσεων του μορίου. Λάβαμε μια λύση σε αυτό το πρόβλημα για το Ν-τερματικό τριδεκαπεπτίδιο dynorphin και έναν αριθμό από τα ανάλογα του χρησιμοποιώντας θεωρητική ανάλυση διαμόρφωσης σε πολικά και μη πολικά περιβάλλοντα.
Πρακτική εργασία στην ενότητα
"Αναπαραγωγή γενετικές πληροφορίες»
Είναι γνωστό ότι σύγχρονη πορείαΗ γενική βιολογία για τα σχολεία περιέχει ανεπαρκές υλικό για πρακτικά μαθήματα. Επιπλέον, η ανεπάρκεια ή η απουσία υλικών πόρων, η έλλειψη εξοπλισμού και αναλώσιμων στα σχολικά χημικά και βιολογικά εργαστήρια προκαλούν μια δύσκολη κατάσταση με τα εργαστηριακά και πρακτικά μαθήματα στο μάθημα. γενική βιολογία. Ωστόσο, ένα τμήμα του μαθήματος όπως η «Αναπαραγωγή γενετικής πληροφορίας» παρέχει αρκετές ευκαιρίες για πρακτική εξάσκηση προκειμένου να αναπτυχθούν δεξιότητες στην επεξεργασία και λειτουργία γενετικών πληροφοριών.
Αυτή η εργασία αντιπροσωπεύει την ανάπτυξη ενός πρακτικού μαθήματος που μπορεί να χρησιμοποιηθεί για τη διεξαγωγή ανεξάρτητων και δοκιμέςσε αυτό το θέμα χρησιμοποιώντας υλικά για τη χημεία των κυττάρων.
Κατά τη διεξαγωγή ενός μαθήματος, μπορούν να επιτευχθούν οι ακόλουθοι στόχοι.
1. Εμπέδωση γνώσεων για τη δομή και τις ιδιότητες γενετικός κώδικας.
2. Εμπέδωση γνώσεων σχετικά με τη διαδικασία αναδιπλασιασμού - αντιγραφή προτύπου DNA και την αρχή της συμπληρωματικότητας.
3. Εμπέδωση γνώσεων για τη μεταγραφή και μετάφραση γενετικής πληροφορίας - η διαδικασία μετάδοσης.
4. Διατύπωση της θεμελιώδους αρχής της βιολογίας σχετικά με τη μεταφορά γενετικών πληροφοριών σε ένα κύτταρο:
DNA--->mRNA--->πρωτεΐνη.
5. Επεξήγηση της δυνατότητας μετάδοσης πληροφοριών από ιούς RNA σύμφωνα με το σχήμα:
ιικό RNA--->cDNA--->mRNA--->ιική πρωτεΐνη.
7. Γνωριμία με τις μεθόδους της σύγχρονης βιοτεχνολογίας.
Φυσικά, αυτό κάθε άλλο παρά εξαντλεί τους στόχους της προτεινόμενης εργασίας, αλλά καλύπτουν τις πιο σημαντικές ενότητες του θέματος «Αναπαραγωγή γενετικών πληροφοριών».
Για τη διεξαγωγή του μαθήματος, πρέπει να έχετε καλή γνώση του υλικού σχετικά με τις ιδιότητες και τη δομή του γενετικού κώδικα, τις διαδικασίες αναπαραγωγής γενετικών πληροφοριών (ανατύπωση, μεταγραφή και μετάφραση), την αρχή της συμπληρωματικότητας, τον κανόνα του Chargaff, ο οποίος πρέπει να επαναληφθεί πριν από την εκτέλεση της εργασίας.
Η μεταφορά της γενετικής πληροφορίας συμβαίνει πάντα με έναν ορισμένο τρόπο, που αντικατοπτρίζεται στο λεγόμενο «κεντρικό δόγμα της βιολογίας», δηλαδή, μόνο προς την κατεύθυνση από το DNA στο mRNA και μετά στην πρωτεΐνη.
Το πρώτο στάδιο αναπαραγωγής της γενετικής πληροφορίας, που ονομάζεται μεταγραφή, εμφανίζεται με τη βοήθεια της RNA πολυμεράσης, η οποία δημιουργεί ένα συμπληρωματικό αντίγραφο του γονιδίου με τη μορφή mRNA.
Στο δεύτερο στάδιο, που ονομάζεται αναμετάδοση, οι πληροφορίες μεταφράζονται από τη γλώσσα των νουκλεοτιδίων (RNA) στη γλώσσα των αμινοξέων (πρωτεΐνη). Έτσι, πραγματοποιείται γενετική πληροφορία για την κατασκευή λειτουργικών μονάδων - μορίων πρωτεΐνης με συγκεκριμένες λειτουργίες, οι οποίες είναι και γενετικά σταθεροποιημένες.
Όταν οι ιοί που περιέχουν RNA εισέρχονται σε ένα κύτταρο, πληροφορίες μπορούν να μεταδοθούν κατά μήκος της αλυσίδας: ιικό RNA--->cDNA--->DNA--->mRNA--->πρωτεΐνη ιού. Αυτή η διαδικασία υλοποιείται χρησιμοποιώντας αντίστροφη μεταγραφάση, η οποία στο πρώτο στάδιο αναπαραγωγής της γενετικής πληροφορίας του ιού δημιουργεί κωδικοποιητικό DNA (cDNA) με βάση το εκμαγείο ιικού RNA. Αυτό το cDNA στη συνέχεια εισάγεται στο DNA του κυττάρου ξενιστή. Ωστόσο, αυτό συμβαίνει μόνο όταν χρησιμοποιούνται οι πόροι του κυττάρου στο οποίο έχει διεισδύσει ο ιός.
Ένα τέτοιο σχήμα για τη μεταφορά γενετικών πληροφοριών θεωρείται αταβισμός. Αυτό οφείλεται στο γεγονός ότι το RNA, προφανώς, κατά τη διάρκεια της χημικής εξέλιξης άρχισε να παίζει το ρόλο ενός μορίου πληροφοριών νωρίτερα από το DNA. Το κύριο επιχείρημα υπέρ αυτής της δήλωσης είναι η παρουσία ενζυματικής δραστηριότητας Μόρια RNA, που ανακαλύφθηκε από τον Thomas Cech, και την ικανότητα των μορίων RNA να αναπαράγονται. Ο συγγραφέας αυτής της ανακάλυψης τιμήθηκε με το βραβείο Νόμπελ.
Ωστόσο, η ριβοενζυμική δραστηριότητα του RNA είναι δεκάδες χιλιάδες φορές χαμηλότερη από εκείνη της RNA πολυμεράσης και μόνο μικρά θραύσματα RNA -ολιγονουκλεοτίδια μήκους έως 50-100 βάσεων- την έχουν. Από την άλλη πλευρά, υπάρχει η άποψη ότι η δραστηριότητα της ριβοζύμης είναι δευτερεύουσα και δεν έχει σχέση με τη χημική εξέλιξη.
Ένας μοναδικός γενετικός κώδικας χρησιμοποιείται για την καταγραφή γενετικών πληροφοριών. Εάν σε ένα εργαστήριο γίνει γνωστή η αλληλουχία αμινοξέων μιας πρωτεΐνης, τότε σε άλλο εργαστήριο μπορούν να γράψουν τις αντίστοιχες νουκλεοτιδικές αλληλουχίες DNA (ή RNA) και το αντίστροφο.
Για δραστηριότητες στην τάξη, μπορείτε να προσφέρετε διάφορες μορφές εργασίας που βασίζονται στη συμπλήρωση χαρτών νουκλεοτιδίων και χαρτών αμινοξέων των αντίστοιχων πολυπεπτιδίων (Παραρτήματα 1-4). Αυτή μπορεί να είναι ατομική ή ομαδική εργασία. Η εργασία σε ομάδες μπορεί να θεωρηθεί ως η εργασία χωριστών εργαστηρίων βιοτεχνολογίας, καθένα από τα μέλη των οποίων εκτελεί μια συγκεκριμένη λειτουργία. Μεμονωμένοι μαθητές ή ομάδες ανταλλάσσουν κάρτες, συμπληρώνοντάς τις σταδιακά. Στο τέλος της εργασίας, μια ομάδα ειδικών ή ένας ειδικός (αυτός θα μπορούσε να είναι δάσκαλος) ελέγχει τις κάρτες, εντοπίζοντας σφάλματα μετάλλαξης.
Η πολυπλοκότητα της εργασίας θα εξαρτηθεί από την ικανότητα χρήσης εκπαιδευτικό υλικό: πίνακες του γενετικού κώδικα, σχήματα αναδιπλασιασμού, μεταγραφή και μετάφραση, πίνακες συμπληρωματικότητας, ιδιότητες του γενετικού κώδικα κ.λπ. Στο μάθημα μπορεί να δοθεί ο χαρακτήρας εργαστηριακής, πρακτικής, ανεξάρτητης ή ελέγχου.
Για να καθορίσετε εργασίες, είναι καλύτερο να χρησιμοποιείτε χάρτες μικρών πολυπεπτιδίων, για παράδειγμα, ορισμένων πεπτιδικών ορμονών. Για το σκοπό αυτό, είναι βολικό να χρησιμοποιούνται ολιγοπεπτίδια των ορμονών βαζοπρεσίνη και ωκυτοκίνη, καθώς και εγκεφαλίνες μεθειονίνης και λευκίνης - φυσικές ενδορφίνες που παράγονται στο σώμα των ζώων και των ανθρώπων (Παραρτήματα 1-4). Η βαζοπρεσίνη και η ωκυτοκίνη έχουν ευρύ φάσμα δράσης και οι ενδογενείς ουσίες που μοιάζουν με μορφίνη έχουν τραβήξει την προσοχή σε σχέση με το πρόβλημα του εθισμού στα ναρκωτικά και την εξήγηση της ναρκωτικής επίδρασης.
Οι κάρτες μπορεί να περιλαμβάνουν υλικό από την ενότητα "Cell Chemistry", δηλαδή τους τύπους και τις ιδιότητες των αμινοξέων. Τα ολιγοπεπτίδια της βαζοπρεσσίνης και της ωκυτοκίνης περιέχουν αμινοξέα που περιέχουν SH (κυστεΐνη), τα οποία σχηματίζουν δισουλφιδικές γέφυρες στο δευτερεύουσα δομήπεπτίδιο, το οποίο μπορεί να αντικατοπτρίζεται στον βαθμό δυσκολίας της εργασίας.
Οι κάρτες περιλαμβάνουν κωδικόνια τερματισμού, τα οποία πρέπει να γράφονται σε αντίστοιχες τριπλέτες στις αλυσίδες DNA ή RNA. Περιλαμβάνεται επίσης το κωδικόνιο εκκίνησης για το αμινοξύ μεθειονίνη, το οποίο στην περίπτωση αυτή βρίσκεται στην αρχή της αλυσίδας.
Τα νουκλεοτίδια της κύριας αλληλουχίας μετά το κωδικόνιο εκκίνησης (και τα αντίστοιχα αμινοξέα) δεν περιλαμβάνονται στο περιεχόμενο των χαρτών, καθώς δεν έχουν θεμελιώδη σημασία για την επεξεργασία της γενετικής πληροφορίας και αφαιρούνται από την αλληλουχία αμινοξέων κατά την επεξεργασία (πρωτεόλυση).
Η προτεινόμενη εργασία των μαθητών με κάρτες και τη συμπλήρωση πινάκων για μετάφραση γενετικών πληροφοριών (αναδιπλασιασμός, μεταγραφή, μετάφραση), γραφή τύπων και συμβόλων αμινοξέων μπορεί να σχεδιαστεί για 1-2 μαθήματα, ανάλογα με την πολυπλοκότητα και τη φύση της εργασίας.
Στο τέλος του μαθήματος οι μαθητές βαθμολογούνται και διατυπώνονται τα ακόλουθα συμπεράσματα.
Η γενετική πληροφορία είναι καθολική. Μορφές ζωής με άλλους γενετικούς κώδικες δεν έχουν ανακαλυφθεί, δηλ. Ο γενετικός κώδικας είναι ίδιος για όλους τους οργανισμούς και δεν υπάρχει άλλος γενετικός κώδικας. Αυτός ο κώδικας έχει αρκετές δυνατότητες για να περιγράψει ολόκληρη την ποικιλία των πρωτεϊνικών μορίων.
Οι χάρτες χρησιμοποιούν γενικά αποδεκτές συντομογραφίες: mRNA – messenger RNA; cDNA – Κωδικός κλώνος DNA. συνθ. Το DNA είναι ο συμπληρωματικός κλώνος του DNA. Το κωδικόνιο αμινοξέος επιλέχθηκε αυθαίρετα, ως ένα από τα πιθανά, που επιτρέπεται στην εργασία των μαθητών.
Για τη διεξαγωγή του μαθήματος, χρησιμοποιούνται παραλλαγές καρτών στις οποίες λείπει οποιαδήποτε γραμμή, δηλ. Υπάρχουν 5 παραλλαγές κάθε κάρτας. Αντίστοιχα, η εργασία μπορεί να διανεμηθεί σε συγκεκριμένο αριθμό μαθητών και ομάδων. Μπορείτε να προσφέρετε εργασία σε άλλους χάρτες για άλλα πεπτίδια, ο αριθμός των οποίων είναι πρακτικά απεριόριστος.
Παράρτημα 1
Η μεθειονίνη-εγκεφαλίνη είναι μια ορμόνη των πυρήνων του εγκεφαλικού φλοιού, ένα ενδογενές οπιοειδές πεπτίδιο, που αποτελείται από 5 αμινοξέα
Αμινοξέων Comp. DNA |
Παράρτημα 2
Η λευκίνη-εγκεφαλίνη είναι μια ορμόνη των πυρήνων του εγκεφαλικού φλοιού, ένα ενδογενές οπιοειδές πεπτίδιο, που αποτελείται από 5 αμινοξέα
Χημική φόρμουλαρίζα αμινοξέων |
|||||||
Αμινοξέων Comp. DNA |
Παράρτημα 3
Βαζοπρεσίνη - αντιδιουρητική ορμόνη - που παράγεται από την υπόφυση, προκαλεί σύσπαση των λείων μυών, μειώνει την απέκκριση νερού, αποτελείται από 9 αμινοξέα με έναν δισουλφιδικό δεσμό
Χημικός τύπος ρίζας αμινοξέος |
Ενδορφίνες(ενδογενείς μορφίνες (από το όνομα του αρχαίου Έλληνα θεού Μορφέα - «αυτός που διαμορφώνει τα όνειρα») - μια ομάδα πολυπεπτιδικών χημικών ενώσεων παρόμοιας δομής με τα οπιούχα (ενώσεις που μοιάζουν με μορφίνη), που παράγονται φυσικά στους νευρώνες του εγκεφάλου και έχουν την ικανότητα να μειώνουν τον πόνο παρόμοιο με τα οπιούχα και να επηρεάζουν τη συναισθηματική κατάσταση.
Οι ενδορφίνες σχηματίζονται από λιποτροπίνες στον εγκεφαλικό ιστό και στον ενδιάμεσο λοβό της υπόφυσης. Γενικού τύπουΗ δομή για αυτές τις ενώσεις είναι μια τετραπεπτιδική αλληλουχία στο Ν-άκρο. Η βήτα-ενδορφίνη σχηματίζεται από τη βήτα-λιποτροπίνη με πρωτεόλυση. Η βήτα-λιποτροπίνη σχηματίζεται από την πρόδρομη προορμόνη προοπικορτίνη (μοριακό βάρος 29 kDa, 134 υπολείμματα αμινοξέων).
Στον πρόσθιο λοβό της υπόφυσης, το πρόδρομο μόριο διασπάται σε ACTH και β-λιποτροπίνη, τα οποία εκκρίνονται στο πλάσμα. Ένα μικρό μέρος (περίπου 15%) της β-λιποτροπίνης διασπάται για να σχηματίσει β-ενδορφίνη. Η βιοσύνθεση της προοπικορτίνης στην πρόσθια υπόφυση ρυθμίζεται από την κορτικολιμπερίνη στον υποθάλαμο. Τρεις διαφορετικές πρόδρομες πρωτεΐνες πεπτιδίων οπιοειδών είναι γνωστές: η προεγκεφαλίνη, η προοπιομελανοκορτίνη και η προδυνορφίνη.
Τα φυσικά οπιοειδή πεπτίδια απομονώθηκαν για πρώτη φορά το 1976 από τον εγκέφαλο των θηλαστικών. Αυτές ήταν οι λεγόμενες εγκεφαλίνες - λευκίνη-εγκεφαλίνη και μεθειονίνη-εγκεφαλίνη, που διαφέρουν μόνο στο τελικό C-κατάλοιπο.
Στις αρχές της δεκαετίας του '70, διάφορα εργαστήρια σε όλο τον κόσμο ανακάλυψαν ότι τα εγκεφαλικά κύτταρα έχουν υποδοχείς που δεσμεύουν τη μορφίνη και μόνο σε αυτή τη δεσμευμένη μορφή γίνεται ενεργή. Δεν υπήρχε λόγος να υποθέσουμε ότι ο εγκέφαλος προετοίμασε ειδικά αυτού του είδους τους υποδοχείς για ένα τόσο σπάνιο συστατικό όπως η μορφίνη. Προέκυψε μια υποψία ότι η λειτουργία αυτών των υποδοχέων ήταν να δεσμεύουν όχι τη μορφίνη, αλλά κάποια ουσία κοντά σε αυτήν, που παράγεται από τον ίδιο τον οργανισμό. Το 1976, ο Δρ. Hughes στη Σκωτία εξήγαγε αυτή τη μυστηριώδη ουσία από τον εγκέφαλο ενός ινδικού χοιριδίου, το οποίο αμέσως παρουσίασε μια απότομη μείωση της ευαισθησίας στον πόνο. Ο Χιουζ ονόμασε την ουσία εγκεφαλίνη, που στα ελληνικά σημαίνει «από τον εγκέφαλο». Και ο καθηγητής Cho Hao Lee στο Σαν Φρανσίσκο εξήγαγε ένα άλλο εσωτερικό φάρμακο από τον εγκέφαλο της καμήλας, και πιο συγκεκριμένα από την υπόφυση της καμήλας, η οποία αποδείχθηκε ότι ήταν 50 φορές ισχυρότερη από τη διάσημη μορφίνη. Ο Cho το ονόμασε ενδορφίνη - «εσωτερική μορφίνη». Το ίδιο 1976, δύο ακόμη εσωτερικά φάρμακα απομονώθηκαν από το αίμα των ζώων, τα οποία ήταν παρόμοια με τη μορφίνη στη σύνθεση, αλλά, σε αντίθεση με τη φυτική μορφίνη, δεν κατέστειλαν την αναπνοή και δεν οδήγησαν σε εθισμό στα ναρκωτικά. Και τέλος, ο Δρ Πλες στην Ελβετία συνέθεσε ενδορφίνη, δηλαδή την παρήγαγε στο εργαστήριο, σε δοκιμαστικό σωλήνα, γνωρίζοντας ακριβώς χημική σύνθεσηκαι τη δομή αυτής της μυστηριώδους ουσίας. Άλλα οπιοειδή πεπτίδια, οι ενδορφίνες, έχουν επίσης απομονωθεί από εκχυλίσματα ιστών της υπόφυσης και του υποθαλάμου των θηλαστικών. Όλα αυτά περιέχουν συνήθως ένα υπόλειμμα εγκεφαλίνης στη Ν-τελική περιοχή. Όλα τα ενδογενή πεπτίδια οπιοειδών συντίθενται στο σώμα ως μεγάλες πρόδρομες πρωτεΐνες με πρωτεόλυση. Η χωρική δομή των εγκεφαλινών είναι παρόμοια με τη μορφίνη. Οι εγκεφαλίνες και οι ενδορφίνες έχουν αναλγητική δράση, μειώνουν την κινητική δραστηριότητα του γαστρεντερικού σωλήνα και επηρεάζουν τη συναισθηματική κατάσταση.
· MSH - ορμόνη διέγερσης μελανοκυττάρων.
· LPG - λιποτροπική ορμόνη.
· CPPP - ενδιάμεσο πεπτίδιο παρόμοιο με κορτικοτροπίνη.
· Η ACTH είναι αδρενοκορτικοτροπική ορμόνη.
Ρύθμιση της έκκρισης
Όλα τα προϊόντα διάσπασης POMC παράγονται σε ισομοριακές ποσότητες και εκκρίνονται στο αίμα ταυτόχρονα. Έτσι, είναι αδύνατο να αυξηθεί η έκκριση της αδρενοκορτικοτροπικής ορμόνης χωρίς ταυτόχρονη αύξηση της έκκρισης της βήτα-λιποτροπικής ορμόνης.Η παραγωγή της POMC ρυθμίζεται από παράγοντες που σχηματίζονται στον υποθάλαμο και τον παρακοιλιακό πυρήνα του εγκεφάλου: κορτικολιμπερίνη, αργινίνη αγγειοπιεσίνη - ενεργοποιήστε τη σύνθεση της ACTH, της κορτιζόλης - του κύριου αναστολέα της σύνθεσης της κορτικοτροπικής ορμόνης και του σχηματισμού POMC, επομένως, η κορτικολιμπερίνη, η αργινίνη αγγειοπιεσίνη και η κορτιζόλη θα επηρεάσουν τη σύνθεση και την έκκριση της β-ενδορφίνης.
Η σύνθεση της β-ενδορφίνης μειώνεται σε ενδοκρινικές, λοιμώδεις και ιογενείς ασθένειες, στο σύνδρομο χρόνιας κόπωσης· η σύνθεση μπορεί να αυξηθεί μέσω της σωματικής δραστηριότητας.
Μεταφορά και περιφερικός μεταβολισμός
Οι ενδορφίνες συντίθενται «για μελλοντική χρήση» και απελευθερώνονται στο αίμα σε ορισμένες δόσεις λόγω της εκκένωσης των εκκριτικών κυστιδίων. Το επίπεδό τους στο αίμα αυξάνεται με την αύξηση της συχνότητας απελευθέρωσης ορμονών από τα αδενικά κύτταρα. Εισερχόμενοι στο αίμα, οι ορμόνες συνδέονται με τις πρωτεΐνες του πλάσματος. Τυπικά, μόνο το 5-10% των μορίων της ορμόνης βρίσκονται σε ελεύθερη κατάσταση στο αίμα και μόνο αυτά μπορούν να αλληλεπιδράσουν με τους υποδοχείς.
Η αποικοδόμηση των πεπτιδικών ορμονών αρχίζει συχνά στο αίμα ή στα τοιχώματα των αιμοφόρων αγγείων· αυτή η διαδικασία είναι ιδιαίτερα έντονη στα νεφρά. Οι πρωτεϊνοπεπτιδικές ορμόνες υδρολύονται από πρωτεϊνάσες, δηλαδή εξω- (στα άκρα της αλυσίδας) και ενδοπεπτιδάσες. Η πρωτεόλυση έχει ως αποτέλεσμα το σχηματισμό πολλών θραυσμάτων, μερικά από τα οποία μπορεί να παρουσιάζουν βιολογική δραστηριότητα. Πολλές ορμόνες πρωτεΐνης-πεπτιδίου απομακρύνονται από την κυκλοφορία μέσω της δέσμευσης σε υποδοχείς της μεμβράνης και της επακόλουθης ενδοκυττάρωσης του συμπλέγματος ορμόνης-υποδοχέα. Η αποικοδόμηση τέτοιων συμπλοκών συμβαίνει στα λυσοσώματα· το τελικό προϊόν αποικοδόμησης είναι τα αμινοξέα, τα οποία χρησιμοποιούνται και πάλι ως υποστρώματα σε αναβολικές και καταβολικές διεργασίες.
Ο κύριος στόχος των ενδορφινών είναι το λεγόμενο σύστημα οπιοειδών (ο κύριος σκοπός του είναι η προστασία από βλάβες στο στρες, η ανακούφιση από τον πόνο και ο συντονισμός των συστημάτων οργάνων και ιστών στο επίπεδο του σώματος συνολικά) του σώματος, και ειδικότερα οι υποδοχείς οπιοειδών . Η ενδορφίνη είναι υπεύθυνη για τη ρύθμιση της δραστηριότητας όλων των εσωτερικών αδένων, για την εργασία ανοσοποιητικό σύστημα, πέρα από το επίπεδο πίεσης, η ενδορφίνη επηρεάζει και το νευρικό σύστημα. Ειδικοί υποδοχείς μορφίνης έχουν ανακαλυφθεί στον εγκέφαλο. Αυτοί οι υποδοχείς βρίσκονται σε συναπτικές μεμβράνες. Το μεταιχμιακό σύστημα είναι πιο πλούσιο σε αυτά, από το οποίο εξαρτάται η συναισθηματική απόκριση. Στη συνέχεια, απομονώθηκαν ενδογενή πεπτίδια από τον εγκεφαλικό ιστό που μιμούνται τις διάφορες επιδράσεις της μορφίνης όταν χορηγείται με ένεση. Αυτά τα πεπτίδια, τα οποία έχουν την ικανότητα να συνδέονται ειδικά με υποδοχείς οπιούχων, ονομάζονται ενδορφίνες και εγκεφαλίνες.
Επειδή Οι υποδοχείς ορμονών οπιούχων βρίσκονται στην εξωτερική επιφάνεια της πλασματικής μεμβράνης, η ορμόνη δεν διεισδύει στο κύτταρο. Οι ορμόνες (οι πρώτοι αγγελιοφόροι σήματος) μεταδίδουν το σήμα μέσω του δεύτερου αγγελιοφόρου, τον ρόλο του οποίου παίζουν τα cAMP, cGMP, τριφωσφορική ινοσοτόλη και ιόντα ασβεστίου. Αφού προσκολληθεί η ορμόνη στον υποδοχέα, ακολουθεί μια αλυσίδα γεγονότων που αλλάζει τον μεταβολισμό του κυττάρου.
Φυσιολογικά, οι ενδορφίνες και οι εγκεφαλίνες έχουν ισχυρή αναλγητική, αντι-σοκ και αντιστρες δράση, μειώνουν την όρεξη και μειώνουν την ευαισθησία ορισμένων τμημάτων του κεντρικού νευρικού συστήματος. Οι ενδορφίνες ομαλοποιούν την αρτηριακή πίεση, τον ρυθμό αναπνοής, επιταχύνουν την επούλωση των κατεστραμμένων ιστών και το σχηματισμό κάλων στα κατάγματα.
Οι ενδορφίνες εμφανίζονται συχνά σε συνδυασμό με την απελευθέρωση αδρεναλίνης. Κατά τη διάρκεια των μακρών προπονήσεων, απελευθερώνεται αδρεναλίνη στο σώμα, αυξάνεται ο μυϊκός πόνος και αρχίζουν να παράγονται ενδορφίνες, οι οποίες μειώνουν τον πόνο και αυξάνουν την αντίδραση του σώματος και την ταχύτητα προσαρμογής στο στρες.
Τι επηρεάζουν τα συστήματα ενδορφινών:
· - αναλγητικά αποτελέσματα
· - επιβράδυνση της αναπνοής, καρδιακός παλμός – δράση κατά του στρες
· - ενίσχυση της ανοσίας
· - ρύθμιση της νεφρικής αιματικής ροής
· - ρύθμιση της εντερικής δραστηριότητας
· - συμμετοχή στις διαδικασίες διέγερσης και αναστολής στο νευρικό σύστημα
· - συμμετοχή στις διαδικασίες ανάπτυξης συνειρμικών-διαχωριστικών συνδέσεων στο νευρικό σύστημα - ρύθμιση του μεταβολικού ρυθμού
· - αίσθημα ευφορίας
· - επιταχύνει την επούλωση των κατεστραμμένων ιστών
· -σχηματισμός κάλων κατά τη διάρκεια καταγμάτων
Επιπλέον, οι ενδορφίνες σχετίζονται με τη θερμορύθμιση, τη μνήμη, τη λιπόλυση, την αναπαραγωγή, την εμπειρία ευχαρίστησης, την καταστροφή του σωματικού λίπους, την αντιδιούρηση, την καταστολή του υπεραερισμού ως απόκριση στο αυξημένο διοξείδιο του άνθρακα και την αναστολή της σύνθεσης θυρεοτροπίνης και γοναδοτροπίνης.
Παθολογία
Έλλειψη ενδορφίνης παρατηρείται στην κατάθλιψη και σε καταστάσεις συνεχούς συναισθηματικού στρες, επιδεινώνει χρόνιες παθήσεις και μπορεί να προκαλέσει σύνδρομο χρόνιας κόπωσης. Εξ ου και η συνοδευτική κατάθλιψη στη διάθεση και η αυξημένη ευαισθησία σε μολυσματικές ασθένειες.
Η παραγωγή ενδορφίνης μειώνεται σε ορισμένες παθολογίες. Ως αποτέλεσμα της έλλειψης ενδορφινών στον οργανισμό, αυξάνεται ο κίνδυνος χρόνιων ασθενειών, οι λεγόμενες «ασθένειες του τρόπου ζωής», οι οποίες έχουν γίνει πρόσφατα η κύρια αιτία θνησιμότητας. Οι ασθένειες του τρόπου ζωής περιλαμβάνουν διαβήτη, καρδιαγγειακές παθήσεις, χρόνιες παθήσεις του αναπνευστικού, καρκίνο και παχυσαρκία.
Η έλλειψη ενδορφινών εκφράζεται με απάθεια, πολύ κακή διάθεση και τελικά οδηγεί τον άνθρωπο σε κατάθλιψη. Κάθε άτομο θέλει να μάθει πώς να απολαμβάνει τη ζωή. Το αίσθημα ευχαρίστησης ενός ατόμου εμφανίζεται όταν αυξάνεται το επίπεδο των ενδορφινών, οι οποίες παράγονται από τον εγκέφαλο και αυτό χημική ένωσηπαρόμοια με το φάρμακο μορφίνη. Ως εκ τούτου, η ενδορφίνη έλαβε ένα τέτοιο όνομα - ενδογενής μορφίνη, δηλαδή, που παράγεται από το ίδιο το σώμα.
Η πιο σοβαρή εκδήλωση είναι η ανηδονία, μια ασθένεια στην οποία ένα άτομο δεν μπορεί να βιώσει ευχαρίστηση.
Νευροορμόνες
Οι νευροορμόνες είναι ουσίες με υψηλή φυσιολογική δραστηριότητα που παράγονται σε νευροεκκριτικά κύτταρα του νευρικού συστήματος (νευρώνες).
Σύμφωνα με τον μηχανισμό δράσης τους, έχουν πολλά κοινά με τους νευροδιαβιβαστές, αλλά οι νευροορμόνες, σε αντίθεση με αυτές, εισέρχονται στο αίμα και σε άλλα βιολογικά υγρά του σώματος (λέμφος, εγκεφαλονωτιαίο υγρό και υγρό ιστών) και έχουν μακροχρόνια, απομακρυσμένη ρυθμιστική δράση. .
Με χημική δομήΟι νευροορμόνες είναι πεπτίδια (περιέχουν αμινοξέα) ή κατεχολαμίνες (βιογενείς αμίνες), το υποχρεωτικό τμήμα τους είναι η 3,4-διυδροξυφαινυλαλανίνη (κατεχόλη).
Οι νευροορμόνες διατηρούν την ομοιόσταση νερού-αλατιού, ρυθμίζουν τον τόνο των λείων μυών και τις μεταβολικές διεργασίες και επίσης συμμετέχουν στη ρύθμιση της δραστηριότητας των ενδοκρινών αδένων. Γενικά, η λειτουργία αυτών των ουσιών είναι να διατηρούν τις προστατευτικές και προσαρμοστικές λειτουργίες του σώματος.
Η σύνθεση των νευροορμονών λαμβάνει χώρα στα νευροεκκριτικά κύτταρα του υποθαλάμου (ντοπαμίνη, βαζοπρεσίνη, ωκυτοκίνη, νορεπινεφρίνη, σεροτονίνη και παράγοντες απελευθέρωσης), νωτιαίος μυελός, επίφυση, επινεφρίδια (χρωμαφινικός ιστός του μυελού) Συντίθενται επίσης στα γάγγλια, τα παραγάγγλια και τους νευρικούς κορμούς του αυτόνομου νευρικού συστήματος (σύνθεση αδρεναλίνης και νορεπινεφρίνης).
Η διαδικασία της βιοσύνθεσης των πεπτιδικών νευροορμονών λαμβάνει χώρα στο σώμα του νευρώνα, σε μια δομή που ονομάζεται ενδοπλασματικό δίκτυο. στη συνέχεια, στο σύμπλεγμα Golgi, συσκευάζονται σε κόκκους και από εκεί μεταφέρονται κατά μήκος του άξονα στις νευρικές απολήξεις.
Νευροφυσιολογία του ύπνου
Νευροφυσιολογικοί μηχανισμοί ύπνου και τα χαρακτηριστικά του που σχετίζονται με την ηλικία
Ο ύπνος είναι μια φυσιολογική κατάσταση που χαρακτηρίζεται από την απώλεια ενεργών νοητικών συνδέσεων του υποκειμένου με τον κόσμο γύρω του. Ο ύπνος είναι ζωτικής σημασίας για τα ανώτερα ζώα και τους ανθρώπους. Για πολύ καιρό πίστευαν ότι ο ύπνος είναι μια ανάπαυση απαραίτητη για την αποκατάσταση της ενέργειας των εγκεφαλικών κυττάρων μετά την ενεργό εγρήγορση. Ωστόσο, αποδεικνύεται ότι η εγκεφαλική δραστηριότητα κατά τη διάρκεια του ύπνου είναι συχνά υψηλότερη από ό,τι κατά τη διάρκεια της εγρήγορσης. Διαπιστώθηκε ότι η δραστηριότητα των νευρώνων σε μια σειρά εγκεφαλικών δομών αυξάνεται σημαντικά κατά τη διάρκεια του ύπνου, δηλ. Ο ύπνος είναι μια ενεργή φυσιολογική διαδικασία.
Στάδια ύπνου
Οι αντανακλαστικές αντιδράσεις κατά τον ύπνο μειώνονται. Ένα άτομο που κοιμάται δεν αντιδρά σε πολλές εξωτερικές επιρροές εκτός και αν είναι υπερβολικά ισχυρό.
Θεωρίες ύπνου:
Χιουμοριστική θεωρία, θεωρεί ως αιτία ύπνου ουσίες που εμφανίζονται στο αίμα κατά την παρατεταμένη εγρήγορση. Η απόδειξη αυτής της θεωρίας είναι ένα πείραμα στο οποίο ένας ξύπνιος σκύλος μεταγγίστηκε με το αίμα ενός ζώου που είχε στερηθεί τον ύπνο για 24 ώρες. Το ζώο παραλήπτης αποκοιμήθηκε αμέσως. Όμως οι χυμικοί παράγοντες δεν μπορούν να θεωρηθούν ως η απόλυτη αιτία ύπνου. Αυτό αποδεικνύεται από τις παρατηρήσεις της συμπεριφοράς δύο ζευγών αχώριστων διδύμων. Το νευρικό τους σύστημα ήταν εντελώς διαχωρισμένο και το κυκλοφορικό τους σύστημα είχε πολλές αναστομώσεις. Αυτά τα δίδυμα θα μπορούσαν να κοιμηθούν διαφορετική ώρα: το ένα κορίτσι, για παράδειγμα, μπορούσε να κοιμάται, ενώ το άλλο ξύπνιο.
Υποφλοιώδεις και φλοιώδεις θεωρίες ύπνου.Με διάφορους όγκους ή μολυσματικές αλλοιώσεις του υποφλοιώδους, ειδικά του εγκεφαλικού στελέχους, των σχηματισμών, οι ασθενείς εμφανίζουν διάφορες διαταραχές ύπνου - από αϋπνία έως παρατεταμένο ληθαργικό ύπνο, που υποδηλώνει την παρουσία κέντρων υποφλοιώδους ύπνου. Όταν οι οπίσθιες δομές του υποθαλάμου και του υποθαλάμου ερεθίστηκαν, τα ζώα αποκοιμήθηκαν και μετά τη διακοπή του ερεθισμού ξύπνησαν, γεγονός που υποδηλώνει την παρουσία κέντρων ύπνου σε αυτές τις δομές.
Χημική θεωρία.Σύμφωνα με αυτή τη θεωρία, κατά τη διάρκεια της εγρήγορσης, εύκολα οξειδωμένα προϊόντα συσσωρεύονται στα κύτταρα του σώματος, με αποτέλεσμα την έλλειψη οξυγόνου, και το άτομο αποκοιμιέται. Πέφτουμε για ύπνο όχι επειδή είμαστε δηλητηριασμένοι ή κουρασμένοι, αλλά για να μην δηλητηριαζόμαστε και κουραζόμαστε.
Λειτουργίες ύπνου
o παρέχει ανάπαυση στο σώμα.
o παίζει σημαντικό ρόλο στις μεταβολικές διεργασίες. Κατά τη διάρκεια του ύπνου βραδέων κυμάτων, απελευθερώνεται αυξητική ορμόνη. Ύπνος REM: αποκατάσταση της πλαστικότητας των νευρώνων και εμπλουτισμός τους με οξυγόνο. βιοσύνθεση πρωτεϊνών και RNA νευρώνων.
o προωθεί την επεξεργασία και αποθήκευση πληροφοριών. Ο ύπνος (ειδικά ο αργός ύπνος) διευκολύνει την εδραίωση του μελετημένου υλικού, ενώ ο ύπνος REM υλοποιεί υποσυνείδητα μοντέλα αναμενόμενων γεγονότων. Η τελευταία περίσταση μπορεί να χρησιμεύσει ως ένας από τους λόγους για το φαινόμενο του déjà vu.
o αυτή είναι η προσαρμογή του σώματος στις αλλαγές του φωτισμού (μέρα-νύχτα).
o αποκαθιστά την ανοσία ενεργοποιώντας τα Τ-λεμφοκύτταρα που καταπολεμούν το κρυολόγημα και τις ιογενείς ασθένειες.
Τύποι ύπνου
Μετά από περαιτέρω λεπτομερή μελέτη, αποδείχθηκε ότι ο ύπνος είναι ετερογενής στις φυσιολογικές του εκδηλώσεις και έχει δύο ποικιλίες: αργό (ήρεμο ή ορθόδοξο) και γρήγορο (ενεργητικό ή παράδοξο).
Με τον ύπνο αργών κυμάτων, η αναπνοή και ο καρδιακός ρυθμός μειώνονται, οι μύες χαλαρώνουν και οι κινήσεις των ματιών επιβραδύνονται. Καθώς ο ύπνος βραδέων κυμάτων βαθαίνει, ο συνολικός αριθμός κινήσεων του ατόμου που κοιμάται γίνεται ελάχιστος. Αυτή τη στιγμή είναι δύσκολο να τον ξυπνήσεις. Ο ύπνος NREM συνήθως καταλαμβάνει 75 - 80%.
Κατά τη διάρκεια του ύπνου REM, αντίθετα, ενεργοποιούνται φυσιολογικές λειτουργίες: η αναπνοή και ο καρδιακός ρυθμός αυξάνονται, σωματική δραστηριότηταΚατά τη διάρκεια του ύπνου, οι κινήσεις των βολβών των ματιών γίνονται γρήγορες (γι' αυτό και αυτό το είδος ύπνου ονομάζεται «γρήγορος»). Οι γρήγορες κινήσεις των ματιών δείχνουν ότι ο κοιμισμένος ονειρεύεται αυτή τη στιγμή. Και αν τον ξυπνήσετε 10 - 15 λεπτά μετά το τέλος των γρήγορων κινήσεων των ματιών, θα σας πει για αυτό που είδε στο όνειρό του. Όταν ξυπνά κανείς κατά τη διάρκεια του ύπνου αργών κυμάτων, συνήθως δεν θυμάται όνειρα. Παρά τη σχετικά μεγαλύτερη ενεργοποίηση των φυσιολογικών λειτουργιών στον ύπνο REM, οι μύες του σώματος είναι χαλαροί κατά τη διάρκεια αυτής της περιόδου και είναι πολύ πιο δύσκολο να ξυπνήσει ο κοιμισμένος. Ο ύπνος REM είναι σημαντικός για τη λειτουργία του σώματος. Εάν ένα άτομο στερηθεί τεχνητά τον ύπνο REM (ξύπνησε σε περιόδους γρήγορων κινήσεων των ματιών), τότε, παρά την αρκετά επαρκή συνολική διάρκεια ύπνου, μετά από πέντε έως επτά ημέρες θα αναπτύξει ψυχικές διαταραχές.
Ο εναλλασσόμενος γρήγορος και αργός ύπνος είναι χαρακτηριστικός για υγιείς ανθρώπους και το άτομο αισθάνεται ξεκούραστο και σε εγρήγορση.
Υπάρχει μια άλλη ταξινόμηση των σταδίων ύπνου:
1. Φάση εξισορρόπησης: χαρακτηρίζεται από επίδραση τόσο σε ισχυρά όσο και σε αδύναμα ερεθίσματα.
2. Παράδοξη φάση: τα ισχυρά ερεθίσματα προκαλούν πιο αδύναμες αποκρίσεις από τα αδύναμα ερεθίσματα.
3. Φάση Ultradox: ένα θετικό ερέθισμα αναστέλλει, και ένα αρνητικό προκαλεί ένα εξαρτημένο αντανακλαστικό.
4. Ναρκωτική φάση: γενική μείωση της εξαρτημένης αντανακλαστικής δραστηριότητας με σημαντικά ισχυρότερη μείωση των αντανακλαστικών σε αδύναμα ερεθίσματα παρά σε ισχυρά.
5. Ανασταλτική φάση: πλήρης αναστολή ρυθμισμένων αντανακλαστικών
Ηλικιακά χαρακτηριστικά:
Ο ύπνος των παιδιών είναι επιφανειακός και ευαίσθητος. Κοιμούνται πολλές φορές την ημέρα.
Στα νεογέννητα, ο ύπνος καταλαμβάνει το μεγαλύτερο μέρος της ημέρας και ο ενεργοποιημένος ύπνος ή ο ύπνος με συσπάσεις (ανάλογος με τον παράδοξο ύπνο στους ενήλικες), αποτελεί την πλειοψηφία του ύπνου. Τους πρώτους μήνες μετά τη γέννηση, ο χρόνος εγρήγορσης αυξάνεται γρήγορα, το ποσοστό του παράδοξου ύπνου μειώνεται και ο αργός ύπνος αυξάνεται.
Υγιεινή ύπνου:
Ο ύπνος πρέπει να έχει επαρκή διάρκεια και βάθος για την ηλικία. Τα παιδιά με κακή υγεία, όσα αναρρώνουν από οξείες μολυσματικές ασθένειες, αυξημένη διεγερσιμότητα του νευρικού συστήματος και παιδιά που κουράζονται γρήγορα, υποτίθεται ότι κοιμούνται για μεγαλύτερο χρονικό διάστημα. Πριν πάτε για ύπνο θα πρέπει να αποκλείονται τα διεγερτικά παιχνίδια και η έντονη διανοητική εργασία. Το δείπνο πρέπει να είναι ελαφρύ, το αργότερο 2-1,5 ώρες πριν τον ύπνο. Καλό για ύπνο:
· φρέσκος, δροσερός αέρας στο δωμάτιο (15-16)
· το κρεβάτι δεν πρέπει να είναι ούτε μαλακό ούτε σκληρό.
· καθαρά, απαλά κλινοσκεπάσματα χωρίς ρυτίδες ή σημάδια
· Είναι προτιμότερο να ξαπλώνετε στη δεξιά πλευρά ή στην πλάτη, κάτι που εξασφαλίζει πιο ελεύθερη αναπνοή και δεν δυσκολεύει το έργο της καρδιάς.
Τα παιδιά πρέπει να διδάσκονται να σηκώνονται και να πάνε για ύπνο την ίδια ώρα. Ένα παιδί αναπτύσσεται αρκετά εύκολα εξαρτημένα αντανακλαστικάστο περιβάλλον ύπνου. Το εξαρτημένο ερέθισμα σε αυτή την περίπτωση είναι η ώρα του ύπνου.
Νευροφυσιολογία του ΑΝΣ
Η έννοια του αυτόνομου νευρικού συστήματος εισήχθη για πρώτη φορά το 1801 από τον Γάλλο γιατρό A. Besha. Αυτό το τμήμα του κεντρικού νευρικού συστήματος παρέχει τις αυτόνομες λειτουργίες του σώματος και περιλαμβάνει τρία συστατικά:
1) συμπαθητικός?
2) παρασυμπαθητικός?
3) μετασυμπαθητικός.
Οι βλαστικές λειτουργίες περιλαμβάνουν εκείνες τις λειτουργίες που διασφαλίζουν το μεταβολισμό στο σώμα μας (πέψη, κυκλοφορία του αίματος, αναπνοή, απέκκριση κ.λπ.). Αυτά περιλαμβάνουν επίσης τη διασφάλιση της ανάπτυξης και ανάπτυξης του σώματος, την αναπαραγωγή και την προετοιμασία του σώματος για δυσμενείς επιπτώσεις. Το αυτόνομο σύστημα παρέχει ρύθμιση της δραστηριότητας των εσωτερικών οργάνων, των αιμοφόρων αγγείων, των ιδρωτοποιών αδένων και άλλων παρόμοιων λειτουργιών. Ρυθμίζει το μεταβολισμό, τη διεγερσιμότητα και αυτόνομη λειτουργίαεσωτερικά όργανα, καθώς και τη φυσιολογική κατάσταση των ιστών και των μεμονωμένων οργάνων (συμπεριλαμβανομένου του εγκεφάλου και του νωτιαίου μυελού), προσαρμόζοντας τη δραστηριότητά τους στις περιβαλλοντικές συνθήκες.
Το συμπαθητικό τμήμα του νευρικού συστήματος διασφαλίζει την κινητοποίηση των διαθέσιμων πόρων του σώματος (ενέργεια και πνευματική) για την εκτέλεση επείγουσας εργασίας.Είναι σαφές ότι αυτό μπορεί να οδηγήσει σε ανισορροπίες στο σώμα.Αποκατάσταση ισορροπίας και σταθερότητας εσωτερικό περιβάλλοντο σώμα είναι καθήκον του παρασυμπαθητικού νευρικού συστήματος. Για να γίνει αυτό, είναι απαραίτητο να διορθώνονται συνεχώς οι μετατοπίσεις που προκαλούνται από τις επιρροές του συμπαθητικού τμήματος, για την αποκατάσταση και διατήρηση της ομοιόστασης. Υπό αυτή την έννοια, η δραστηριότητα αυτών των τμημάτων του νευρικού αυτόνομο σύστημα σε έναν αριθμό αντιδράσεων εκδηλώνεται ως ανταγωνιστικό.
Στη φυσιολογία, η υποομοιόσταση νοείται ως η διατήρηση σταθερών εσωτερικών περιβαλλοντικών παραμέτρων στο σώμα. Αυτά περιλαμβάνουν τη διατήρηση σταθερής σύνθεσης αίματος, θερμοκρασίας σώματος κ.λπ.
Τα κέντρα του αυτόνομου νευρικού συστήματος βρίσκονται στο εγκεφαλικό στέλεχος και στο νωτιαίο μυελό. Τα κέντρα του παρασυμπαθητικού νευρικού συστήματος βρίσκονται στο εγκεφαλικό στέλεχος και στο ιερό τμήμα του νωτιαίου μυελού.Στον μεσεγκέφαλο υπάρχουν κέντρα που ρυθμίζουν τη διαστολή της κόρης και την προσαρμογή του ματιού. Στον προμήκη μυελό υπάρχουν κέντρα του νευρικού παρασυμπαθητικού συστήματος, από το οποίο αναχωρούν ίνες ως μέρος του πνευμονογαστρικού, του προσώπου και των γλωσσοφαρυγγικών νεύρων. Αυτά τα κέντρα εμπλέκονται σε μια σειρά από λειτουργίες, συμπεριλαμβανομένης της ρύθμισης της δραστηριότητας ενός αριθμού εσωτερικών οργάνων (καρδιά, στομάχι, έντερα, ήπαρ κ.λπ.) και «πυροδοτούν» την έκκριση σάλιου, δακρυϊκού υγρού κ.λπ. αυτές οι λειτουργίες εκτελούνται σύμφωνα με την αρχή του αντανακλαστικού (από τον τύπο της απόκρισης σε ένα ερέθισμα). Μερικά από αυτά τα αντανακλαστικά θα περιγραφούν παρακάτω.
Τα κέντρα του νευρικού παρασυμπαθητικού αυτόνομου συστήματος βρίσκονται επίσης στα ιερά τμήματα του νωτιαίου μυελού. Οι ίνες από αυτά πηγαίνουν ως μέρος των πυελικών νεύρων, που νευρώνουν τα πυελικά όργανα (παχύ έντερο, κύστη, γεννητικά όργανα κ.λπ.).
Τα κέντρα του συμπαθητικού νευρικού συστήματος βρίσκονται στο θωρακικό και οσφυϊκό τμήμα του νωτιαίου μυελού. Οι αυτόνομες ίνες από αυτά τα κέντρα αναχωρούν ως μέρος των πρόσθιων ριζών του νωτιαίου μυελού μαζί με τα κινητικά νεύρα.
Όλα τα παραπάνω κέντρα του συμπαθητικού και του νευρικού παρασυμπαθητικού συστήματος είναι υποδεέστερα στο ανώτερο αυτόνομο κέντρο - τον υποθάλαμο. Ο υποθάλαμος, με τη σειρά του, επηρεάζεται από διάφορα άλλα κέντρα του εγκεφάλου. Όλα αυτά τα κέντρα αποτελούν το μεταιχμιακό σύστημα. Μια πλήρης περιγραφή του συστήματος θα δοθεί στο αντίστοιχο θέμα, αλλά τώρα ας δούμε το «έργο» των περιφερειακών τμημάτων του νευρικού αυτόνομου συστήματος.
Και στις δύο πλευρές της σπονδυλικής στήλης στην κοιλιακή πλευρά υπάρχουν δύο κορμοί του συμπαθητικού νευρικού συστήματος. Ονομάζονται επίσης συμπαθητικές αλυσίδες. Η αλυσίδα αποτελείται από μεμονωμένα γάγγλια που συνδέονται μεταξύ τους και το νωτιαίο μυελό με πολυάριθμες νευρικές ίνες. Κάθε ίνα που φτάνει στο γάγγλιο νευρώνει έως και αρκετές δεκάδες νευρώνες στο γάγγλιο (απόκλιση). Χάρη σε αυτή τη διάταξη, οι συμπαθητικές επιρροές έχουν συνήθως διάχυτο, γενικευμένο χαρακτήρα. Με τη σειρά τους, τα νεύρα απομακρύνονται από αυτά τα γάγγλια, τα οποία πηγαίνουν στα τοιχώματα των αιμοφόρων αγγείων, στους ιδρωτοποιούς αδένες και στα εσωτερικά όργανα. Εκτός από τα γάγγλια του οριακού κορμού, σε κάποια απόσταση από αυτά υπάρχουν τα λεγόμενα προσπονδυλικά γάγγλια, τα μεγαλύτερα από αυτά είναι το ηλιακό πλέγμα και οι μεσεντερικοί κόμβοι.
Τα επινεφρίδια παίζουν σημαντικό ρόλο στη δραστηριότητα του συμπαθητικού νευρικού συστήματος.Σχηματίζονται στον άνθρωπο κατά την προγεννητική περίοδο λόγω της μετανάστευσης νευροβλαστών (μη διαφοροποιημένων νευρώνων) από τον νευρικό σωλήνα στην περιοχή των νεφρών. Εκεί, αυτά τα κύτταρα σχηματίζουν ένα ειδικό όργανο στις κορυφές και των δύο νεφρών - τα επινεφρίδια. Τα επινεφρίδια νευρώνονται από τα συμπαθητικά νεύρα. Επιπλέον, μπορούν να ενεργοποιηθούν από την αδρενοκορτικοτροπική ορμόνη, η οποία απελευθερώνεται ως απάντηση στο στρες από την υπόφυση και φτάνει στα επινεφρίδια μαζί με το αίμα. Υπό την επίδραση αυτής της ορμόνης, ένα μείγμα αδρεναλίνης και ρεπινεφρίνης απελευθερώνεται από τα επινεφρίδια στο αίμα, τα οποία μεταφέρονται μέσω της κυκλοφορίας του αίματος και προκαλούν μια σειρά από συμπαθητικές αντιδράσεις (αύξηση καρδιακού ρυθμού, εφίδρωση, αυξημένη παροχή αίματος στους μύες, ερυθρότητα του δέρματος και πολλά άλλα).
Οι άξονες των συμπαθητικών νευρώνων στις περιφερειακές συνάψεις απελευθερώνουν τον μεσολαβητή αδρεναλίνη Τα μόρια της αδρεναλίνης και της νορεπινεφρίνης αλληλεπιδρούν με τους αντίστοιχους υποδοχείς. Υπάρχουν δύο τύποι τέτοιων υποδοχέων: οι άλφα και οι βήτα αδρενεργικοί υποδοχείς. Ορισμένα εσωτερικά όργανα έχουν μόνο έναν από αυτούς τους υποδοχείς, άλλα έχουν και τους δύο. Έτσι, στα τοιχώματα των αιμοφόρων αγγείων υπάρχουν τόσο άλφα όσο και βήτα αδρενεργικοί υποδοχείς. Η σύνδεση του συμπαθητικού πομπού με τον άλφα-αδρενεργικό υποδοχέα προκαλεί στένωση των αρτηριολίων και η σύνδεση με τον βήτα-αδρενεργικό υποδοχέα προκαλεί τη διαστολή των αρτηριολίων. Στο έντερο, όπου υπάρχουν και οι δύο τύποι αδρενεργικών υποδοχέων, ο μεσολαβητής αναστέλλει τη δραστηριότητά του. Στον καρδιακό μυ και στα τοιχώματα των βρόγχων υπάρχουν μόνο β-αδρενεργικοί υποδοχείς - ο συμπαθητικός πομπός προκαλεί διαστολή των βρόγχων και αυξημένο καρδιακό ρυθμό.
Τα γάγγλια της παρασυμπαθητικής διαίρεσης του νευρικού αυτόνομου συστήματος, σε αντίθεση με τα συμπαθητικά, βρίσκονται στα τοιχώματα των εσωτερικών οργάνων ή κοντά σε αυτά. Η νευρική ίνα (άξονας νευρώνα) από το αντίστοιχο παρασυμπαθητικό κέντρο στο εγκεφαλικό στέλεχος ή στον ιερό νωτιαίο μυελό φτάνει στο νευρωμένο όργανο χωρίς διακοπή και καταλήγει στους νευρώνες του παρασυμπαθητικού γαγγλίου. Ο επόμενος παρασυμπαθητικός νευρώνας βρίσκεται είτε μέσα στο όργανο είτε σε κοντινή απόσταση από αυτό. Οι ενδοοργανικές ίνες και τα γάγγλια σχηματίζουν πλέγματα πλούσια σε νευρώνες στα τοιχώματα πολλών εσωτερικών οργάνων της καρδιάς, των πνευμόνων, του οισοφάγου, του στομάχου κ.λπ., καθώς και στους εξωκρινείς και αδένες εσωτερικής έκκρισης. Η ανατομική δομή του παρασυμπαθητικού τμήματος του νευρικού αυτόνομου συστήματος δείχνει ότι η επίδρασή του στα όργανα είναι πιο τοπική από αυτή του συμπαθητικού νευρικού συστήματος.
Ο μεσολαβητής στις περιφερικές συνάψεις του παρασυμπαθητικού νευρικού συστήματος είναι η ακετυλοχολίνη, για την οποία υπάρχουν δύο τύποι υποδοχέων: οι Μ- και οι Ν-χολινεργικοί υποδοχείς. Αυτή η διαίρεση βασίζεται στο γεγονός ότι οι Μ-χολινεργικοί υποδοχείς χάνουν την ευαισθησία τους στην ακετυλοχολίνη υπό την επίδραση της ατροπίνης (που απομονώνεται από έναν μύκητα του γένους Muscaris) και στους Η-χολινεργικούς υποδοχείς - υπό την επίδραση της νικοτίνης.
Η επίδραση του συμπαθητικού και του παρασυμπαθητικού αυτόνομου συστήματος στις λειτουργίες του σώματος. Στα περισσότερα όργανα, η διέγερση του συμπαθητικού και του νευρικού παρασυμπαθητικού αυτόνομου συστήματος προκαλεί αντίθετα αποτελέσματα. Ωστόσο, πρέπει να έχουμε κατά νου ότι αυτές οι αλληλεπιδράσεις δεν είναι απλές. Για παράδειγμα, τα παρασυμπαθητικά νεύρα προκαλούν χαλάρωση των σφιγκτήρων της κύστης και ταυτόχρονα συστολή των μυών της ουροδόχου κύστης. Τα συμπαθητικά νεύρα συσπούν τον σφιγκτήρα και ταυτόχρονα χαλαρώνουν τους μύες. Ένα άλλο παράδειγμα: η διέγερση των συμπαθητικών νεύρων αυξάνει τον ρυθμό και τη δύναμη των καρδιακών συσπάσεων και ο ερεθισμός του πνευμονογαστρικού (παρασυμπαθητικού) νεύρου μειώνει τον ρυθμό και τη δύναμη των καρδιακών συσπάσεων. Επιπλέον, μελέτες έχουν δείξει ότι μεταξύ αυτών των τμημάτων του νευρικού αυτόνομου συστήματος δεν υπάρχει μόνο ανταγωνισμός (πολυκατευθυντική), αλλά και συνέργεια (μονοκατευθυντική). Η αύξηση του τόνου ενός μέρους του νευρικού αυτόνομου συστήματος, κατά κανόνα, οδηγεί σε αύξηση του τόνου ενός άλλου τμήματος. Επιπλέον, αποδείχθηκε ότι υπάρχουν όργανα και ιστοί με έναν μόνο τύπο νεύρωσης. Για παράδειγμα, τα αγγεία του δέρματος, ο μυελός των επινεφριδίων, η μήτρα, οι σκελετικοί μύες και μερικά άλλα έχουν μόνο συμπαθητική νεύρωση και οι σιελογόνοι αδένες νευρώνονται μόνο από παρασυμπαθητικές ίνες.
Αυτόνομα αντανακλαστικά. Αυτά τα αντανακλαστικά είναι πολλά. Συμμετέχουν σε πολλούς κανονισμούς του ανθρώπινου σώματος. Όταν εκτελούνται φυτικά αντανακλαστικά, οι επιρροές μεταδίδονται κατά μήκος των αντίστοιχων νεύρων (συμπαθητικών ή παρασυμπαθητικών) από το κεντρικό νευρικό σύστημα. Στην ιατρική πράξη υψηλότερη τιμήδίνουν σπλαχνικά-σπλαχνικά (από το ένα εσωτερικό όργανο στο άλλο), σπλαχνικό-δερματικό (από τα εσωτερικά όργανα στο δέρμα) και δερμο-σπλαχνικά (από το δέρμα στα εσωτερικά όργανα).
Τα σπλαχνικά περιλαμβάνουν αντανακλαστικές αλλαγές στην καρδιακή δραστηριότητα, τον αγγειακό τόνο, την παροχή αίματος στον σπλήνα με αύξηση ή μείωση της πίεσης στην αορτή, στον καρωτιδικό κόλπο ή στα πνευμονικά αγγεία. Για παράδειγμα, λόγω της συμπερίληψης ενός τέτοιου αντανακλαστικού, εμφανίζεται καρδιακή ανακοπή όταν ερεθίζονται τα κοιλιακά όργανα. Τα σπλάχνα-δερμικά αντανακλαστικά εμφανίζονται όταν τα εσωτερικά όργανα ερεθίζονται και εκδηλώνονται με αλλαγές στην ευαισθησία των αντίστοιχων περιοχών του δέρματος (σύμφωνα με το ποιο όργανο ερεθίζεται), εφίδρωση και αγγειακή αντίδραση. Τα δερμοσπλαχνικά αντανακλαστικά εκδηλώνονται στο γεγονός ότι όταν ορισμένες περιοχές του δέρματος ερεθίζονται, η λειτουργία των αντίστοιχων εσωτερικών οργάνων αλλάζει. Η πραγματική χρήση της θέρμανσης ή ψύξης ορισμένων περιοχών του δέρματος για ιατρικούς σκοπούς, για παράδειγμα για πόνο στα εσωτερικά όργανα, βασίζεται στον μηχανισμό αυτών των αντανακλαστικών.
Τα αυτόνομα αντανακλαστικά χρησιμοποιούνται συχνά από τους γιατρούς για να κρίνουν τη λειτουργική κατάσταση του νευρικού αυτόνομου συστήματος. Για παράδειγμα, στην κλινική, οι αντανακλαστικές αλλαγές στα αιμοφόρα αγγεία μελετώνται συχνά κατά τη διάρκεια μηχανικού ερεθισμού του δέρματος (για παράδειγμα, όταν ένα αμβλύ αντικείμενο περνά πάνω από το δέρμα). Σε ένα υγιές άτομο, αυτό προκαλεί βραχυπρόθεσμη λεύκανση της ερεθισμένης περιοχής του δέρματος (λευκός δερμογραφισμός, δερματοδερμία). Με υψηλή διεγερσιμότητα του νευρικού αυτόνομου συστήματος, εμφανίζεται μια κόκκινη λωρίδα στο σημείο του ερεθισμού του δέρματος, που συνορεύει με ωχρές λωρίδες στενωμένων αγγείων (κόκκινος δερμογραφισμός) και με ακόμη υψηλότερη ευαισθησία - πρήξιμο του δέρματος σε αυτό το σημείο. Συχνά στην κλινική χρησιμοποιούνται λειτουργικές αυτόνομες εξετάσεις για να κριθεί η κατάσταση του νευρικού αυτόνομου συστήματος. Για παράδειγμα, η ορθοστατική αντίδραση: όταν μετακινείται από μια ξαπλωμένη σε μια όρθια θέση, η αρτηριακή πίεση αυξάνεται και ο καρδιακός ρυθμός αυξάνεται. Το πρότυπο των αλλαγών στην αρτηριακή πίεση και την καρδιακή δραστηριότητα κατά τη διάρκεια αυτής της εξέτασης μπορεί να χρησιμεύσει ως διαγνωστικό σημάδι ασθένειας στο σύστημα ελέγχου της αρτηριακής πίεσης. Ένα άλλο παράδειγμα είναι η οφθαλμική-καρδιακή αντίδραση (αντανακλαστικό Aschner): όταν ασκείται πίεση στους βολβούς των ματιών, εμφανίζεται βραχυπρόθεσμη μείωση του καρδιακού ρυθμού.
Φυτικά κέντρα. Ο προμήκης μυελός περιέχει νευρικά κέντρα που αναστέλλουν τη δραστηριότητα της καρδιάς (πυρήνες του πνευμονογαστρικού νεύρου). Στον δικτυωτό σχηματισμό του προμήκη μυελού υπάρχει ένα αγγειοκινητικό κέντρο, που αποτελείται από δύο ζώνες: τον πιεστικό και τον καταθλιπτικό. Η διέγερση της πιεστικής ζώνης οδηγεί σε αγγειοσυστολή και η διέγερση της πιεστικής ζώνης οδηγεί στη διαστολή τους. Το αγγειοκινητικό κέντρο και οι πυρήνες του πνευμονογαστρικού νεύρου στέλνουν συνεχώς ώσεις, χάρη στις οποίες διατηρείται ένας σταθερός τόνος: οι αρτηρίες και τα αρτηρίδια στενεύουν συνεχώς κάπως και η καρδιακή δραστηριότητα επιβραδύνεται.
Ο προμήκης μυελός περιέχει το αναπνευστικό κέντρο, το οποίο, με τη σειρά του, αποτελείται από κέντρα εισπνοής και εκπνοής. Στο επίπεδο της γέφυρας υπάρχει ένα κέντρο αναπνοής υψηλότερου επιπέδου (πνευμοταξικό κέντρο), το οποίο προσαρμόζει την αναπνοή στις αλλαγές της φυσικής δραστηριότητας. Η αναπνοή στον άνθρωπο μπορεί επίσης να ελεγχθεί εκούσια από τον εγκεφαλικό φλοιό, για παράδειγμα κατά τη διάρκεια της ομιλίας.
Στον προμήκη μυελό υπάρχουν κέντρα που διεγείρουν την έκκριση των σιελογόνων, δακρυϊκών και γαστρικών αδένων, την έκκριση της χολής από τη χοληδόχο κύστη και την έκκριση του παγκρέατος. Στον μεσεγκέφαλο, κάτω από τους πρόσθιους φυμάτιους του τετραδύμου, υπάρχουν παρασυμπαθητικά κέντρα προσαρμογής του οφθαλμού και το αντανακλαστικό της κόρης. Όλα τα παραπάνω κέντρα του συμπαθητικού και του παρασυμπαθητικού συστήματος είναι υποδεέστερα στο ανώτερο αυτόνομο κέντρο - τον υποθάλαμο.
Ο ρόλος του υποθαλάμου στη ρύθμιση των αυτόνομων λειτουργιών. Η επίδραση στη ρύθμιση του συμπαθητικού και του παρασυμπαθητικού επιτρέπει στον υποθάλαμο να επηρεάζει τις αυτόνομες λειτουργίες του σώματος μέσω χυμικών και νευρικών οδών. Ήταν προηγουμένως κατανοητό ότι ο ερεθισμός των πυρήνων της πρόσθιας ομάδας συνοδεύεται από παρασυμπαθητικά αποτελέσματα. Ο ερεθισμός των πυρήνων της οπίσθιας ομάδας προκαλεί συμπαθητικές επιδράσεις στη λειτουργία των οργάνων. Πυρηνική διέγερση μεσαία ομάδαοδηγεί σε μείωση της επιρροής του συμπαθητικού τμήματος του αυτόνομου νευρικού συστήματος. Η καθορισμένη κατανομή των λειτουργιών του υποθαλάμου δεν είναι απόλυτη. Όλες οι δομές του υποθαλάμου είναι ικανές να παράγουν συμπαθητικά και παρασυμπαθητικά αποτελέσματα σε διάφορους βαθμούς. Κατά συνέπεια, υπάρχουν λειτουργικές συμπληρωματικές, αμοιβαία αντισταθμιστικές σχέσεις μεταξύ των δομών του υποθαλάμου.
Συνολικά λόγω μεγάλη ποσότητασυνδέσεις, πολυλειτουργικότητα δομών, ο υποθάλαμος εκτελεί μια ολοκληρωμένη λειτουργία αυτόνομης, σωματικής και ενδοκρινικής ρύθμισης, η οποία εκδηλώνεται επίσης στην οργάνωση ορισμένων ειδικών λειτουργιών από τους πυρήνες του. Έτσι, στον υποθάλαμο υπάρχουν κέντρα ομοιόστασης, θερμορύθμισης, πείνας και κορεσμού, δίψας και ικανοποίησης αυτής, σεξουαλική συμπεριφορά, φόβος, οργή, ρύθμιση του κύκλου εγρήγορσης-ύπνου. Όλα αυτά τα κέντρα πραγματοποιούν τις λειτουργίες τους ενεργοποιώντας ή αναστέλλοντας το αυτόνομο νευρικό σύστημα, το ενδοκρινικό σύστημα και τις δομές του εγκεφαλικού στελέχους και του πρόσθιου εγκεφάλου.
Ο υποθάλαμος, με τη σειρά του, επηρεάζεται από μια σειρά ανώτερων κέντρων του εγκεφάλου, συμπεριλαμβανομένου του φλοιού.
Ετσι, Το αυτόνομο νευρικό σύστημα έχει μια σειρά από ανατομικά και φυσιολογικά χαρακτηριστικά που καθορίζουν τους μηχανισμούς λειτουργίας του:
Ανατομικές ιδιότητες
1. Διάταξη τριών συστατικών νευρικών κέντρων. Το χαμηλότερο επίπεδο του συμπαθητικού τμήματος αντιπροσωπεύεται από τα πλάγια κέρατα από τον VII αυχενικό έως τον III-IV οσφυϊκό σπόνδυλο και το παρασυμπαθητικό τμήμα αντιπροσωπεύεται από τα ιερά τμήματα και το εγκεφαλικό στέλεχος. Τα ανώτερα υποφλοιώδη κέντρα βρίσκονται στο όριο των πυρήνων του υποθαλάμου (το συμπαθητικό τμήμα είναι η οπίσθια ομάδα και το παρασυμπαθητικό τμήμα είναι η πρόσθια ομάδα). Το επίπεδο του φλοιού βρίσκεται στην περιοχή της έκτης έως της όγδοης περιοχής Brodmann (κινητοαισθητηριακή περιοχή), στο οποίο σημείο επιτυγχάνεται εντοπισμός των εισερχόμενων νευρικών ερεθισμάτων. Λόγω της παρουσίας μιας τέτοιας δομής του αυτόνομου νευρικού συστήματος, το έργο των εσωτερικών οργάνων δεν φτάνει στο κατώφλι της συνείδησής μας.
2. Παρουσία αυτόνομων γαγγλίων. Στο συμπαθητικό τμήμα, βρίσκονται είτε και στις δύο πλευρές κατά μήκος της σπονδυλικής στήλης (αλυσίδα συμπαθητικού νεύρου), είτε αποτελούν μέρος των πλεγμάτων. Έτσι, το τόξο έχει μια σύντομη προγαγγλιακή και μια μακρά μεταγαγγλιακή διαδρομή. Οι νευρώνες της παρασυμπαθητικής διαίρεσης βρίσκονται στο γάγγλιο που βρίσκεται κοντά στο όργανο εργασίας ή στο τοίχωμά του, επομένως το τόξο έχει μακρά προγαγγλιακή και σύντομη μεταγαγγλιακή διαδρομή.
1. Χαρακτηριστικά της λειτουργίας των αυτόνομων γαγγλίων. Η παρουσία του φαινομένου του animation (η ταυτόχρονη εμφάνιση δύο αντίθετων διαδικασιών - απόκλισης και σύγκλισης). Η απόκλιση είναι η απόκλιση των νευρικών ερεθισμάτων από το σώμα ενός νευρώνα σε πολλές μεταγαγγλιακές ίνες ενός άλλου. Σύγκλιση είναι η σύγκλιση στο σώμα κάθε μεταγαγγλιακού νευρώνα παλμών από πολλούς προγαγγλιακούς. Αυτό εξασφαλίζει την αξιοπιστία της μεταφοράς πληροφοριών από το κεντρικό νευρικό σύστημα στο όργανο εργασίας. Η αύξηση της διάρκειας του μετασυναπτικού δυναμικού, η παρουσία ιχνών υπερπόλωσης και συναπτικής καθυστέρησης συμβάλλουν στη μετάδοση της διέγερσης με ταχύτητα 1,5-3,0 m/s. Ωστόσο, οι παρορμήσεις σβήνονται εν μέρει ή μπλοκάρονται πλήρως στα αυτόνομα γάγγλια. Με αυτόν τον τρόπο ρυθμίζουν τη ροή των πληροφοριών από το κεντρικό νευρικό σύστημα. Λόγω αυτής της ιδιότητας, ονομάζονται νευρικά κέντρα που βρίσκονται στην περιφέρεια και το αυτόνομο νευρικό σύστημα ονομάζεται αυτόνομο.
2. Χαρακτηριστικά των νευρικών ινών. Οι προγαγγλιακές νευρικές ίνες ανήκουν στην ομάδα Β και διεξάγουν διέγερση με ταχύτητα 3-18 m/s, οι μεταγαγγλιακές νευρικές ίνες ανήκουν στην ομάδα C. Διενεργούν διέγερση με ταχύτητα 0,5–3,0 m/s. Δεδομένου ότι η απαγωγική οδός του συμπαθητικού τμήματος αντιπροσωπεύεται από προγαγγλιακές ίνες και η παρασυμπαθητική αντιπροσωπεύεται από μεταγαγγλιακές ίνες, η ταχύτητα μετάδοσης των παλμών είναι υψηλότερη στο παρασυμπαθητικό νευρικό σύστημα.
Γενικά, το συμπαθητικό νευρικό σύστημα εκτελεί μια προσαρμοστική-τροφική λειτουργία, εμπλέκεται σε σωματική δραστηριότητα, συναισθηματικές αντιδράσεις, στρες, πόνο και απώλεια αίματος. Εξασφαλίζει την προσαρμογή του οργανισμού στις μεταβαλλόμενες περιβαλλοντικές συνθήκες.
Το παρασυμπαθητικό νευρικό σύστημα είναι ανταγωνιστής του συμπαθητικού νευρικού συστήματος και εκτελεί ομοιοστατικές και προστατευτικές λειτουργίες, ρυθμίζει την κένωση των κοίλων οργάνων. Ο ομοιοστατικός ρόλος έχει αποκαταστατικό χαρακτήρα και δρα σε κατάσταση ηρεμίας. Αυτό εκδηλώνεται με τη μορφή μείωσης της συχνότητας και της ισχύος των καρδιακών συσπάσεων, διέγερσης του γαστρεντερικού σωλήνα με μείωση των επιπέδων γλυκόζης στο αίμα κ.λπ.







