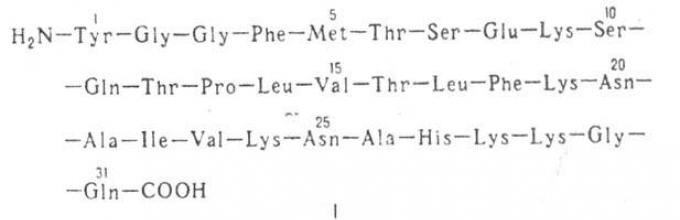
ОПИОИДНЫЕ ПЕПТИДЫ
, группа
природные и синтетич. пеп-тидов, сходных с опиатами (морфин, кодеин и др.) по способности
связываться с опиагными рецепторами организма (структуры нервной системы, по
отношению к к-рым опиаты являются экзогенными лигандами). Природные ОПИОИДНЫЕ ПЕПТИДЫ п. (эндогенные
лиганды опиатных рецепторов) выделены впервые в 1975 из мозга млекопитающих.
Это были так называемой энкефалины - лейцин-энкефалин H 2 N-Туг-Gly-Gly- Phe-Leu-COOH
(молекулярная масса 556; букв. обозначения см. в ст. Аминокислоты)и метионин-энкефалин
H 2 N-Туr -
Gly-Gly-Phe-Met-COOH (молекулярная масса 574), представляющие собой пентапентиды, различающиеся
лишь С-конце-вым аминокислотным остатком. Аминокислотная последовательность
метионин-энкефалина идентична фрагменту 61-65 -липотропина.
Из экстрактов тканей гипофиза
и гипоталамуса млекопитающих выделены и другие ОПИОИДНЫЕ ПЕПТИДЫ п., получившие групповое название
эндорфины. Все они в N-концевой области молекулы содержат обычно остаток энкефалина.
Различают
-эндорфин (формула I),-эндорфин
(его молекула тождественна фрагменту
1-16 (3-эндорфина), -эндорфин
(состоит из остатка -эндорфина,
связанного на С-конце с остатком лейцина) и-эндорфин
(-эндорфин,
связанный на С-конце с Phe-Lys).
и -Эндорфины
тождественны соответственно фрагментам 61-76, 61-91, 61-77 и 61-79-липотропина.
К природным ОПИОИДНЫЕ ПЕПТИДЫ п. относятся также -неоэндорфин
(II), (b-нео-эндорфин (отличается от предыдущего отсутствием на С-конце остатка
лизина), динорфин А (III), динорфин Б (IV), динорфин-32 (состоит из остатков
динорфина А и динор-фина Б, связанных дипептидом Lys-Arg соответственно по С- и N-концам),
дерморфин (V), киоторфин (H 2 N-Туг-Arg -COOH),
казоморфин-5 (H 2 N-Туг -Pro-Phe-Pro -Gly-COOH),
казоморфин-7 (отличается от предыдущего наличием на С-конце дополнительно двух
аминокислотных остатков Pro-Ilе и некоторые др. пептиды.

Все эндогенные ОПИОИДНЫЕ ПЕПТИДЫ п. синтезируются
в организме в виде крупных белков-предшественников, из которых они освобождаются
в результате протеолиза. Известны три различные белка-предшественника ОПИОИДНЫЕ ПЕПТИДЫ п.: проэнкефалин,
проопиомеланокор-тин и продинорфин. Пространств. строение энкефалинов и морфина
сходно. Энкефалины и эндорфины обладают обезболивающим действием (при их введении
непосредственно в мозг), снижают двигат. активность желудочно-кишечного тракта,
влияют на эмоциональное состояние. Действие ОПИОИДНЫЕ ПЕПТИДЫ п. исчезает через несколько секунд
после введения налоксона (VI)-антагониста морфина.

Одновременно с осуществлением
полного химический синтеза природных ОПИОИДНЫЕ ПЕПТИДЫ п. интенсивно изучаются их разнообразные синтетич.
аналоги. Особое внимание уделяется синтезу аналогов ОПИОИДНЫЕ ПЕПТИДЫ п., обладающих повыш.
устойчивостью к действию протеолитич. ферментов. Некоторые синтетич. аналоги ОПИОИДНЫЕ ПЕПТИДЫ
п. проявляют мор-финоподобную активность при периферич. введении.
Литература: Эндорфины,
под ред. Э. Коста, М. Трабукки, пер. с англ., М., 1981; Якубке Х.-Д.,
Ешкайт X., Аминокислоты, пептиды, белки, пер. с нем., М., 1985, с. 289-95. Ю.
П. Швачкин.
Химическая энциклопедия. Том 3 >>
480 руб. | 150 грн. | 7,5 долл. ", MOUSEOFF, FGCOLOR, "#FFFFCC",BGCOLOR, "#393939");" onMouseOut="return nd();"> Диссертация - 480 руб., доставка 10 минут , круглосуточно, без выходных и праздников
240 руб. | 75 грн. | 3,75 долл. ", MOUSEOFF, FGCOLOR, "#FFFFCC",BGCOLOR, "#393939");" onMouseOut="return nd();"> Автореферат - 240 руб., доставка 1-3 часа, с 10-19 (Московское время), кроме воскресенья
Дамиров Аслан Гасан оглы. Теоретический конформационный анализ лейцин-энкефалина, N-концевого тридекапептида динорфина и их аналогов: ил РГБ ОД 61:85-1/1277
Введение
ГЛАВА I. Литературный обзор 8
1.1. Экспериментальные и теоретические методы конформационного анализа пептидов. 9
1.3. Структурно-функциональная организация пептидов 29
1.4. Обратная отруктурная задача 39
1.5. Метод теоретического конформационного анализа " 50
1.6. Задачи исследовании 56
ГЛАВА II. Теоретический конформационный анализ Лейцин-Энкефалина 57
2.1. Биологические функции лейцин-энкефалина 57
2.2. Конформационные возможности лейцин-энкефалина 58
ГЛАВА III. Теоретический конфоешщионный анализ аналогов лейцин-энкефалина 69
3.1. Аналог Туг1- Gly2- Gly3- Phe4- Leu5- Arg6 70
3.2. Аналог Ty^-D- Ala2- Gly3~ Phe4- Leu5 » 72
3.3. Аналог Tyi^-D- Ala2- Gly3- Phe4- Leu5- - Arg6 79
3.4. Аналог Tyr*-D- Ala2- Gly3- Phe4- Ala5- - Arg6 84
3.5. Аналог TyrX-D~ Ala2- Gly3- Phe4- Gly5- - Arg6 88
3.6. Аналог Туг1-])- Ala2- Gly3- Phe4- Ala5- - Lys6 91
3.7. Аналог Туг ^D- Ala2- Gly3- Phe4- Ala5- - Asn6 93
3,8* Аналог Туг ^D*- Ala2- Gly3- Phe4-Aln5~ - Gin6 99
3,9. Аналог Туг -^- D- Ala2- Gly3- Phe4- Ala5- - His6 103
3,10* Аналог Туг -^-D- Ala2- Glu3-N- MePhe4- - Leu5- Arg6 106
ГЛАВА ІУ. Теоретшеский конформационный анализ n -концевого трщшкапептйда динорфина и его аналогов
4.1 Функции динорфина и схема расчета его про странственного строения ИЗ
4.2. Фрагменты Arg - Arg7, Arg - Arg9, Arg -
Bys11, bys11- Lys13 116
4.3. Фрагменты Phe 4- Arg7 и Arg 6- Arg9 121
4.4. Фрагмент Arg 9- Lys13 126
4.5. Фрагмент Туг I*-Arg 9 129
4.6. Молекула N-концевого тридекапептида динорфина 132
4.7. Аналог -динорфина I - ІЗ 137
4.8. Аналог -динорфина І - ІЗ 137
Литература 144
Введение к работе
Обшая характеристика работы. Диссертационная работа посвящена исследованию пространственного строения лейцин-энкефалина, N -концевого тридекапептида динорфина и целого ряда их аналогов методом теоретического конформационного анализа. Потенциальная энергия при расчете конформации молекул взята в виде суммы энергий взаимодействий валентно несвязанных атомов, электростатических, тороион-ных взаимодействий и водородной связи. Для поиска минимума потенциальной энергии применялся метод сопряженных градиентов, В работе была использована разработанная в проблемной лаборатории "Молекулярной биофизики" Азербайджанского Государственного университета им,С.М.Кирова универсальная программа, написанная на алгоритмическом языке "ФОРТРАН", Расчеты проводились на электронно-вычислительных машинах EC-I033, EC-I022 и БЭСМ-6,
Актуальность темы. Многие биологические процессы осуществляются при непременном участии белков и гормонов. Знание пространственной организации и динамических конформационных свойств природных олигопептидов необходимо для изучения молекулярных механизмов узнавания, стимулирования, регуляции и координации биологических процессов, явления структурно-функциональных соотношений и решения многих других вопросов, имеющих большое научное и практическое значение. Методом рентгеноструктурного анализа можно лишь определять статические состояния белков и гормонов. Теоретический кон-формационяый анализ дает представление не только о пространствен** ной структуре, но и о её потенциальных возможностях к изменению. Поэтому работы в этом направлении могут привести к получению ценной информации о тонких деталях биологических процессов на атомно-молекулярном уровне.
Цель работы. Диссертационная работа посвящена изучению пространственного строения и динамических конформационных свойств молекул эндогенных пептидных гормонов лейцин-энкефалина, динорфи-яа, а также большого числа их аналогов.
Апробация теоретического подхода, позволяющего ещё до синтеза и биологических испытаний предсказывать структуры аналогов, обладающих теми или иными функциями природных гормонов. Необходимо было найти такие химические модификации гормонов, которые сделали бы пролонгированным их действие или воспроизводили бы избирательно одну из этих функций. Для этого требовалось, во-первых, выяснить геометрические и энергетические характеристики предпочтительных конформационных состояний природных гормонов (прямая структурная задача) и, во-вторых, целенаправленно модифицировать химическое строение природных гормонов таким образом, чтобы их пространственное строение отвечало вполне определенным низкоэнергетическим конформациям природного пептида (обратная отруктурная задача).
Научная новизна. Впервые установлены пространственные структуры и конформационные возможности лейцин-энкефалина, n -концевого тридекапептида дияорфина и ряда их аналогов. Получено, что лейцин-энкефалин может иметь всего несколько низкоэнергетических структур разных типов, число которых для его аналогов значительно сокращается.
Конформационные состояния аналогов лейцин-энкефалина можно разделить на две группы: аналоги, претерпевающие вследствие замены значительные структурные изменения, и аналоги, в которых замена аминокислот приводит к заметному смещению конформационного равновесия в сторону некоторых форм природного гормона. Найдено, что N -концевой тридекапептид динорфина имеет ограниченный набор низкоэнергетических структурных типов, число которых зависит от
природы окружающей среды. Найдены аналоги, моделирующие те или иные функциональные свойства лейцин-энкефалина и динорфина.
Практическая ценность. На основе аминокислотной последовательности лейцин-энкефалина и динорфина I-I3 определены конформа-ционяые возможности и трехмерные структуры молекул, знание которых необходимо для изучения биологической активности и механизма функционирования этих молекул, а также целенаправленного поиска аналогов.
Апробирован теоретический подход и изучена пространственная структура аналогов. Исследование шляется полезным для целенаправленного синтеза аналогов этих молекул со специфическими функциями. Проведенный расчет показал возможность предсказания геометрии и конформационных возможностей гормонов и химического строения их аналогов с заведомо известными свойствами с помощью метода теоретического конформационного анализа.
Основные положения, выносимые на зашиту: I. Результаты расчета пространственной структуры лейцин-энкефалина Н - Туг 1 - Gly 2 - Gly 5 - Phe^~ Leu^-OH И следующих ЄГО аналогов: H-Tyr 1 -Gly 2 -Gly 5 -Phe 4 -Leu 5 -Arg 6 -OH,
H-Tyr 1 -D-Ala 2 -Gly 5 -Phe 4 -Leu 5 -OH,
H-Tyr 1 -D-Ala 2 -Gly 5 -Phe 4 -Leu 5 -Arg 6 -OH,
H-Tyr 1 -D-Al a 2 -Gly 3 -Phe 4 -Al a 5 -Arg 6 -OH,
H-Tyr 1 -D-Ala 2 -Gly 3 -Phe 4 -Ala 5 -Ly s 6 ~0H,
H-Tyr 1 -D-Ala 2 -Gly 5 -Phe 4 -Ala 5 -Asn 6 -OH,
H-Tyr 1 -D-Al a 2 -Gly 5 -Phe Z| "-Gly 5 -Arg 6 -OH,
H-Tyr 1 -D-Ala 2 -Gly 5 -Phe 4 -Ala 5 -Gln 6 -OH,
H-Tyr 1 -D-Ala 2 -Gly5-phe^-Ala5-His 6 -OH,
H-Tyr 1 -D-Ala 2 -Gly 5 -N-MePhe 4 -Leu 5 -Arg 6 -OH.
2. Результаты расчета пространственной структуры динорфина
І-ІЗ Н -Туг 1 -Gly 2 -Gly 5 -Phe 4 -Leu 5 -Arg 6 -Arg 7 -lie 8 -Arg 9 -Pro 10 -Lys 11 -Leu 12 -Lys 15 -он и следующих его аналогов: [ъ -
Ala 2 ] - и (Ъ -Ala 12 ] -динорфина.
Обсуждение зависимости между химической структурой и пространственным отроением рассмотренных природных и синтетических аминокислотных последовательностей,
Целенаправленный поиск химических модификаций молекул лейцин-энкефалина и динорфина, более устойчивых и воспроизводшцих биологические функции природных пептидов.
Апробация работы. Результаты работы докладывались на УІ Всесоюзном симпозиуме по межмолекулярному взаимодействию и конформа-циям молекул (Вильнюс, 1982 г.), на I Всесоюзном биофизическом съезде (Москва, 1982 г.), на научной конференции, посвященной итогам научно-исследовательских работ за 1983 год (Баку, 1984 г»), на Республиканской конференции молодых ученых по физико-химической биологии (Баку, I98I г.), а также на семинарах кафедры оптики и молекулярной физики и проблемной лаборатории "Молекулярной биофизики" .
Публикация. Основное содержание диссертации изложено в семи публикациях.
Структура и объем диссертации. Диссертационная работа изложена на 159 страницах машинописного текста, состоит из введения, четырех глав и выводов, содержит 10 рисунков, 35 таблиц и список цитированной литературы, включающий 135 наименований.
Прямая структурная задача
Под прямой структурной задачей будем подразумевать установление кинформационных особенностей молекулы по её химическому строению. Решение такой задачи для природных олигопептидов, приведенных в табл. I строилось на основе следующих предположений .
Аминокислотная последовательность полностью определяет пространственное строение олРігопептидной молекулы. 2.Конформацион-ные состояния олигопептидов в физиологических условиях обладают минимальной потенциальной энергией. 3.Пространственное строение природных олигопептидов отвечает шютнеышей упаковке аминокислотной последовательности. 4.В компактных структурах олигопептидов шеет место согласованность между всеми ближними, среднши и дальніми внутримолекулярными взаимодействиями.
Под ближними подразумеваются взаимодействия боковой цепи данного остатка с двумя прилегающими пептидными группами; под среднши - взаимодействия остатка с четырьмя соседями по обе стороны в последовательности; под дальніми - взаимодействия с более удаленными по цепи остатками (рис. I). Ближние взаимодействия определяют конформационные возможности свободных монопептидов. Количественное представление об этой компоненте может дать конфор-мационный анализ соответствующих метиламидов N -ацетил-л-аминокислот. В настоящее время для всех 20 стандартных -аминокислот получены предпочтительные по ближним взаимодействиям наборы оптимальных конформаций, пригодные для расчета любой аминокислотной последовательности. На рис, 2а представлена потенциальная поверхность молекулы метиламида N -ацетил-ь -аланина как функция двугранных углов у и т/г . Поверхность имеет четыре области низкой энергии В, Н, ь и Р. Очень близки нонформационные карты тр- \jr у іб из 20 стандартных аминокислот, боковые цепи которых не содержат разветвление при С атоме. Потенциальные поверхности метиламидов N -ацетил-L -валина и ь -изолейцина имеют те же, но несколько более суженные области низкой энергии.Карта -тг ме-тиламида W -ацетил-глицина центросимметрична и содержит обширные и практически изоэнергетические области R(ь) иВ(Р), рис. 3. Потенциальная поверхность молекулы метиламида N -ацетил-ь -проли-на имеет два минимума, отвечающих значениям угла " г -60 (R.) и- 140 (В), рис. 4.
Теоретические предсказания относительно информационных возможностей и положения конформационного равновесия метиламидов N -ацетил-ос-аминокислот в полярных и неполярных средах соответствуют экспериментальным данным, полученным для целого ряда этих сое-динений с помощью методов ЯМР, ДОВ, КД и ИК-спектроскопии, а также дипольных моментов и газожидкостной осмометрии \р! 65]. В то же время, сопоставление результатов расчета этих молекул с геометрией аминокислотных остатков в белках известной трехмерной структуры показало, что опытные значения углов тр, J\T основной цепи всех остатков находятся только в низкоэнергетических областях (0 т 4.0 ккад/моль) конформационных карт т/ - т/г свободных монопептидов. Удовлетворительное соответствие имеется и между т о о Я сечениями потенциальной поверхности У f , У "Ї ме тиламидов N -ацетил-ot-аминокислот и конформационными состояниями боковых цепей в трехмерной структуре белка [бб - 68]. Обнаружение корреляции между геометрией всех аминокислотных остатков в природных последовательностях и низкоэнергетическими состояниями свободных монопептидов . Наличие корреляции показало, что ближние взаимодействия определяют для. всех стандартных остатков наборы дозволенных пространственных форм. При свертывании пептидной цепи в нативную структуру только из таких наборов происходит выбор (вначале под действием средних, а затем дальних взаимодействий) конформационных состояний всех остатков. Реальными оказываются лишь те предпочтительные по ближним взаимодействиям кон-формации остатков, сочетание которых в данной аминокислотной последовательности обеспечивает образование самых выгодных стабилизирующих межостаточных контактов. Такім образом, специфичной для нативной структуры любого природного пептида оказывается последовательность конформационных состояніш остатков, а не сами состояния. Ситуация здесь в принципе аналогична химическому строению природных пептидов.
Из-за огромного количества исходных вариантов, которые могут быть составлены из низкоэнергетических конформаций остатков, непосредственное решение прямой структурной задачи возможно лишь для коротких последовательностей, включающих не более 4-5 звеньев. Для более высокомолекулярных пептидов цель может быть достигнута только при разделении всей задачи на ряд менее громоздких задач и их последовательном решении. Возможность использования поэтапного подхода обусловлена наличием согласованности между всеми ближними, средними и дальними взаимодействиями. На этой основе получено решение большого числа прямых структурных задач для весьма сложных олигопептидов и низкомолекулярных белко
Конформационные возможности лейцин-энкефалина
При замене четвертого остатка пептидов ВРРк и N -MeAla 4 ВРРс OOOTBeTCTBeHHO Ha D -Ala - И D -N ЧИеАІа - BPPg ВСЄ CTpyK туры, разрешенные для молекул с ь -конфигурацией, окажутся запрещенными для молекул с другой конфигурацией. Поэтому синтетические аналоги D - Ala4 - BPPg и D -N -MeAia 4 BPPg не будут обладать физиологическими свойствами природного соединения. Альтернативный запрет на структуры действует при условии, когда заменяемый ь -остаток предшествует пролину. Но даже, если это уоловие не соблюдается, включение в ь -последовательность D -аминокислоты, особенно замещенной по азоту, может оказаться наиболее эффективным способом решения обратной структурной задачи. Это завиоит от конкретной последовательности, конформационных возможностей природного пептида и цели исследования. Так, замена ь -остатка, находящегося в интересующей структуре в состоянии В, на D -остаток всегда приведет к её запрету. Если же заменяемый остаток имеет L форму основной цепи, то с включением D -остатка возрастает вероятность реализации соответствующего конформационного состояния, поскольку при этом понижается энергия ближних взаимодействий данного остатка и, расширяется область разрешенных значений его углов Y , Цг (рис. 2), что может привести к установлению более эффективных межостаточных контактов. Большие возможности для целенаправленной детерминации ион омой структуры открываются при замещении глицина на ь - и D -аланин. При первой замене окажутся запрещенными все конформации, в которых углы р, яг глицина попадают в область Р, а конформации с углами тр, у глицина в области ь станут менее вероятными. В случае замены Giy на D -Ala окажутся невозможными конформации о В формой глицина, а менее вероятными с В. формой. Кроме того, включение в цепь D -остатка скажется на взаимоотношении пептида с пептидазами и приведет к пролонгированию его действия.
Решение обратной структурной задачи яонапептидного горюна сна также отроится на основе известного набора всех низкоэнергетических конформации природной молекулы и упоминавшихся выше простейших и встречающихся в природе химических модификаций аминокислотных последовательностей.
До сих пор, при решении обратной структурной задачи были использованы такие аминокислотные замены, воздействие которых на конформационные возможности молекулы легко поддавались априорной оценке, во всяком случае на уровне формы основной цепи и шейпа. Строгий количественный анализ требовался лишь для определения порядка расположения конформационных состояний по величинам энергии в узком ряду отобранных вариантов. Однако, все замены, не касающиеся остатков глицина и пролина, не ведут к априорному исключению каких-либо форм и шейпов пептидного окелета. Влияние таких замен на конформационные возможности зависит не столько от ближних, сколько от средних и дальних взаимодействий, т.е. от очень сложной внутримолекулярной контактной системы, определяемой не отдельными остатками, а всей последовательностью. Поскольку в каждом случае ситуация уникальна, то выявить последствия замен даже в отношении форм и шейпов пока можно только расчетным путем. Следующий объект рассмотрения - аналог -пептида, в котором оотатки Ala 2 и Gly 8 заменены на Vai 8. Можно было ожидать, что включение в последовательность сразу двух остатков с объемными и негибкими боковыми цепями существенно изменит энергетическое распределение конформацйи природного нонапептида. Однако до расчета трудно было предположить, что глобальной структурой модифицированной последовательности станет конформация УШ, имеющая в выделенном для 8 -пептида ряду самую высокую энергию. Оказалось, что оба остатка валина хорошо вписываются только в структуру УШ, не изменяя её геометрию и не разрушая сложившуюся систему взаимодей о о ствий; стабильность этой структуры у Vai » -аналога возрастает по сравнению с 8 -пептидом на 8.0 ккал/моль.
Следующая модификация &-пептидной последовательности - заметі я на Asp на Asn , самая незначительная в стерическом отношении, но весьма существенная в электронном. Она делает реальными для Asn 5 -S-пептида структуры лишь одного типа eefeefe , среди которых доминирует конформация Па. Как показывает расчет, в этом случае изменения касаются только двух межостаточных взаимодейст с Я т вий, а именно контактов Asn с Gin и с Trp . Отсутствие отрицательного заряда у остатка в пятом положении, являющегося центральным в структуре Па снимает электростатическое отталкивание между остатками первой пары (о +6.8 ккал/моль у природного -пептида до +0.2 у Asn -аналога). Одновременно,правда, иочезает стабилизирующее полярное взаимодействие между остатками второй пары, которое, однако, у S -пептида в конформацйи Па значительно менее эффективно (-3.0 ккал/моль).
Аналог Tyi^-D- Ala2- Gly3- Phe4- Leu5- - Arg6
Полученные нами низкоэнергетические конформации [р -Ala 2]-ЛЭК приведены в табл. 9, энергетические характеристики самой стабильной конформации каждой формы основной цепи -ЛЭК показаны в табл. 10, Результаты расчета показали, что в по-следовательнооти ЛЭК замена Gly fc на D -Alar приводит к сильной энергетической дифференциации конформации, форм основной цепи и шейпов. Из 16 возможных типов структур только конформации трех, а именно ffff , fffe и efff оказались низкоэнергетическими (табл. 9). В отличие от молекулы ЛЭК, конформации шейпа effe для его аналога являются высокоэнергетичными. Их относительная энергия повысилась в среднем на 3.0 ккал/моль по сравнению с конформация-ми того же шейпа молекулы ЛЭК. Это, по-видимому, связано с тем, что в низкоэнергетических конформациях шейпа efff второй остаток находится в В форме основной цепи, которая менее предпочтительна для D-стереоизомера,
Глобальной структурой аналога ЛЭК является В LP % L32 шейпа ffff в этой конформации дипептидные взаимодействия вносят -9,2 ккал/моль, трипептидные -1.7 ккал/моль, тетрапептид-нне -3,3 ккал/моль и пентапептидные -10.1 ккал/моль. именно последняя составляющая и делает эту конформацию самой выгодной. Энергия конформации В2РНВ2В32 шейпа fffe всего на 0.1 ккал/моль выше энергии глобальной конформации. Здесь более ослаблено пента-пептидное взаимодействие между остатками Туг и Leu , а три- и тетрапептидные взаимодейотвия сильнее. Самый большой вклад в стабилизацию этой конформации вносят дисперсионные взаимодействия (-20.7 ккал/моль). нооительная энергия которой равна 3.0 ккал/моль. На основе результатов расчета можно предположить, что в физиологических условиях о аналог D - Ala -ЛЭК не будет выполнять все функции, присущие природной молекуле ЛЭК. Неосуществимой отанет функция молекулы, для реализации которой необходима структура ЛЭК типа effe . Т.о» результаты конформационного анализа аналога лейцш-энкефалина дают возможность сделать следующие предположения - во-первых, этот аналог сможет осуществлять не все функции ЛЭК, а лишь те, при которых его пространственное строение должно быть ffff , fffe и отчасти efff типов, И во-вторых, вследствие заме? 2 ны аминокислотного остатка Gly на D - Ala молекула аналога приобретает большую устойчивость к действию раощепляющих ферментов, что замедляет ферментативный гидролиз этого аналога в организме и, следовательно, пролонгирует его действие» Аналог Туг I- D- Ala2- Gly3- Phe4- Leu5-Arg 6.
Пространственная структура этого аналога молекулы ЛЭК изучена на основе оптимальных конформаций двух его других аналогов -- и -ЛЭК, рассмотренных выше. Полученные при этом низкоэнергетические конформаций аналога -ЛЭК показаны в табл. II, а энергетические характеристики самой стабильной конформаций каждой формы основной цепи - в табл. 12.
Относительная энергия 28 конформаций этого аналога ниже 5.0 ккал/моль. Все они принадлежат семи разным формам основной цепи, пяти шейпам. Взятые за основу расчета низкоэнергетические варианты аналога -ЛЭК оказались перспективными и сохранены для р- Ala2, Arg 6]-аналога. Низкоэнергетичеокие шейпы основной цепи аналога -ЛЭК, полученные в результате его конформационного анализа, показаны на рис 8.
Относительная энергия конформации B2PRR2B21B5522 шейпа ffffe равна 0,9 ккал/моль, В этой конформации вклад ди- три-, тетра- и пентапептидных взаимодействий соответственно составляет --10,0, -3,9, -5.1 и -7,0 ккал/моль. Как видно, первые два взаимодействия в этой конформации слабее, чем у глобальной, а гекса-пептидное взаимодействие очень мало,
У конформации %РЕВ3В32%22 шейпа fffее относительная энергия составляет 3,2 ккал/моль, В этой конформации стабилизирующее действие оказываю! в основном ди- и пентапептидные взаимодействия. Они равны -8,4 и -9.3 ккал/моль соответственно, В целом сильное стабилизирующее действие оказывают невалентные взаимодействия, порядка -29,0 ккал/моль,
Т,о, можно предположить, что молекула -ЛЭК может сохранять только некоторые биологические функции природного гормона. Но её действие в организме вероятно будет пролонгированным, Аналог Туг - D-Ala 2-Gly 3-Phe 4-А1а 5-Arg 6.
Поскольку пространственная структура этого аналога исследована на основе результатов расчета рассмотренной выше молекулы [п-А1а 2t Arg 6]-ЛЭК, то для этой молекулы были рассчитаны конформации только пяти лучших шейпов пептидного скелета, а именно: fffef , ffffe , effff , efffe и fffee , Найденные предпочтительные конформации показаны в табл. 13, а энергетические характеристики самых стабильных конформации каждой рассмотренной формы основной цепи даны в табл. 14
Фрагменты Phe 4- Arg7 и Arg 6- Arg9
Впервые выделенный Голдштейном и сотр. [І27І из экстракта гипофиза свиньи N-концевой тридекапептид динорфина обладает биологической активностью целого пептида, последовательность динорфина (І-ІЗ) включает лейцин-энкефалин. Аминокислотную последовательность динорфина определяли, сочетая автоматический метод Эд-мана и гидролиз карбоксипептидазами А и Y \l28j. Предполагают, что динорфин является предшественником двух пептидов ІА (N-концевого 17 членного фрагмента) и IB (С-концевого 13 членного фрагмента), образующихся в результате ферментативного гидролиза пептидной связи Ьуз - Arg 19 .
Динорфин отличается от других опиатных пептидов тем, что он взаимодействует с опиатными рецепторами препарата подвздошной кишки морской свинки в 50 раз сильнее р -эндорфина, в 200 раз лучше морфина и в 700 раз сильнее ЛЭК . Динорфин может играть роль в регуляции ответов на изменение водно-солевого обмена, а также участвовать в процессах, связанных с деторождением и кормлением , Биологическая активность динорфина была резистентной к разрушению цианогенбромидом.
Динорфин обладает своеобразным действием на аналгезию, вызываемую у мышей опиоидами, такими как морфин, р -эндорфин (БЭР) и аналоги лейцин-анкефалина. Он ослабляет аналитический эффект морфина, усиливает обезболивающее действие больших доз БЭР, не ослабляя при этом эффект малых доз БЭР, Динорфин (I - 13) не вы зывает аналгезию. Предполагают, что динорфин оказывает модулирующее действие на аналитический эффект морфина, БЭР и некоторых других эндогенных опиоидов «
Динорфин также влияет на поведение мышей, усиливая при центральном введении в мозг чувство голода, однако, эти эффекты не устраняются налакооном # Действие динорфина (I-I3) полностью блокируется налакооном, но для подавления активности динорфина (I-I3) требуется в 13 раз более высокая его концентрация, чем для подавления действия ЛЭК или норморфина . Высокоочищеиные мембраны мозга мышей быстро деградируют динорфин (I-I3), что указывает на присутствие соответствующей протеазы в мембранах мозга»
Казароссианом и сотр. получена высокочувсвительная специфическая количья антисыворотка (Ас) к конъюгату биологически активного фрагмента I-I3 динорфина» Препарат Ас не реагирует о ЛЭК, входящим в структуру динорфина (I-I3). Исследование перекрестной реакции Ас с укороченными фрагментами динорфина (I-I3) показало, что С-концевой остаток лизина-13 и свободная аминогруппа Туг не существенны для иммунореактивности динорфина (I-I3). В то же время, последовательное укорачивание динорфина (I-I3) с С-конца, начиная с 12 го остатка ведет к постепенному снижению иммунореактивности, которая становится крайне низкой после удаления 9-го остатка и практически отсутствует в динорфине (1-5), т.е. лейцин-энкефалине В этой же работе было показано, что замещение Gly на в- Ala2 в динорфине (I-I3) приводит к снижению иммунореактивности .
Чавкин и Голдштейн [іЗЗ] изучали различные участки молекулы динорфина (І-ІЗ) в её взаимодействии с рецепторами нервного сплетения мышц морокой свинки. Удаление аминокислот с С-конца динорфина (І-ІЗ) показало, что лизин-13, лизин-Н и аргинин-7, по-види 115 мому, нужны для сохранения выоокого уровня взаимодействия пептида с рецепторами. Удаление N-концевого тирозина приводило к утрате биологической активности пептида» Авторы \_133"\ предполагают, что ТТ 7 Lys и Arg , по-видимому, нужны для проявления специфичности взаимодействия с рецепторами динорфина (I-I3), отличающимися от мю-рецепторов в той же ткани. Вюстер и др. вводили мышам одновременно в течение 6 дней -ЛЭК (I), как агонист дельта-опиатных рецепторов (-0Р), и суфентанил (П) как агонист j«.-0P. При сравнении опиоидных активностей динорфина (I-I3), -ЛЭК, ЛЭК, суфентанила и др. на препарате vas deferens в концентрациях в 140 - 2500 раз больших для толерантных к I + П мышей, чем для контрольных мышей,получили, что активность динорфина (I-I3) уменьшалась незначительно. На этом основании авторы предполагают, что существуют опиатные рецепторы, специфичные к динорфину (I-I3) и функционирующие независимо от f-- и о 0Р, Таким образом, динорфин (I-I3) и ЛЭК взаимодействуют с различными популяциями опиатных рецепторов.
Спектроскопическими методами изучена структура динорфина (I-13) в воде и других оредах Спектры КД водных растворов динорфина свидительствуют о его неупорядоченности и весьма лабильной структуре Наряду с полифункциональностью,динорфин (I-I3) обладает высокой чувствительностью,специфичностью каждого своего действия А это по-видимому, связано о тем,что молекула динорфина (I-I3) в физиологических условиях не является статистически флуктуирующим клубком, а имеет вполне определенный набор изоэнергетических кон-формаций различных типов Учитывая большое число выполняемых ди норфином функций, можно ожидать, что этот набор является весьма представительным. Для изучения структурно-функциональной организации олигопептида на атомно-молекулярном уровне необходимо знание полного набора низкоэнергетических и фйзиологически"активных" конформационных состояний молекулы. Решение этой задачи получено нами для N-концевого тридекапептида динорфина и ряда его аналогов методом теоретического конформационного анализа в полярной и неполярной средах.
Практические работы по разделу
«Воспроизведение генетической информации»
Известно, что современный курс общей биологии для школ содержит недостаточно материалов для практических занятий. Кроме того, недостаточность или отсутствие материальной базы, дефицит оборудования и расходных материалов в школьных химических и биологических лабораториях обусловливают нелегкое положение с лабораторными и практическими занятиями по курсу общей биологии. Однако такой раздел курса, как «Воспроизведение генетической информации», предоставляет достаточно возможностей для проведения практических занятий с целью выработки навыков по обработке и оперированию генетической информацией.
Настоящая работа представляет собой разработку практического занятия, которое может быть использовано для проведения самостоятельных и контрольных работ по этой теме с привлечением материалов по химии клетки.
При проведении занятия могут быть реализованы следующие цели.
1. Закрепление знаний по структуре и свойствам генетического кода.
2. Закрепление знаний о процессе редупликации – матричном копировании ДНК и принципе комплементарности.
3. Закрепление знаний о транскрипции и трансляции генетической информации – процессе передачи.
4. Формулирование основополагающего
принципа биологии о передаче генетической
информации в клетке:
ДНК--->иРНК--->белок.
5. Объяснение возможности передачи
информации РНК-содержащими вирусами по схеме:
вирусная РНК--->кДНК--->иРНК--->вирусный белок.
7. Знакомство с методами современной биотехнологии.
Разумеется, этим далеко не исчерпываются цели предлагаемого задания, однако они охватывают важнейшие разделы темы «Воспроизведение генетической информации».
Для проведения занятия необходимо хорошее владение материалом о свойствах и строении генетического кода, процессах воспроизведения генетической информации (редупликации, транскрипции и трансляции), принципе комплементарности, правиле Чаргаффа, которые следует повторить перед проведением работы.
Передача генетической информации происходит всегда определенным образом, нашедшим отражение в так называемой «центральной догме биологии», а именно, только в направлении от ДНК к иРНК и далее к белку.
Первый этап воспроизведения генетической информации, называемый транскрипцией , происходит при помощи РНК-полимеразы, которая строит комплементарную копию гена в виде иРНК.
На втором этапе, который называется трансляцией , происходит перевод информации с языка нуклеотидов (РНК) на язык аминокислот (белка). Таким образом, происходит реализация генетической информации для построения функциональных единиц – белковых молекул со специфическими функциями, которые также генетически закреплены.
При попадании РНК-содержащих вирусов в клетку возможна передача информации по цепочке: вирусная РНК--->кДНК--->ДНК--->иРНК--->белок вируса. Этот процесс реализуется с помощью обратной транскриптазы, которая на первом этапе воспроизведения генетической информации вируса строит кодирующую ДНК (кДНК) по матрице вирусной РНК. Затем эта кДНК встраивается в ДНК клетки-хозяина. Однако это происходит только при использовании ресурсов клетки, в которую проник вирус.
Подобную схему переноса генетической информации считают атавизмом. Это связано с тем, что РНК, по-видимому, в ходе химической эволюции стала играть роль информационной молекулы раньше, чем ДНК. Главным аргументом в пользу этого утверждения считают наличие ферментативной активности у молекул РНК, открытой Томасом Чехом, и способность молекул РНК к самовоспроизведению. Автор этого открытия был удостоен Нобелевской премии.
Однако рибозимная активность РНК в десятки тысяч раз ниже, чем у РНК-полимеразы, и обладают ею только короткие фрагменты РНК – олигонуклеотиды длиной до 50–100 оснований. С другой стороны, существует мнение, что рибозимная активность является вторичной и не имеет отношения к химической эволюции.
Для записи генетической информации используется единый генетический код. Если в одной лаборатории становится известной последовательность аминокислот белка, то в другой лаборатории могут написать соответствующие последовательности нуклеотидов ДНК (или РНК), и наоборот.
Для занятия в классе можно предложить несколько форм работы, основанных на заполнении нуклеотидных карт и аминокислотных карт соответствующих им полипептидов (Приложения 1–4). Это может быть индивидуальная и групповая работа. Работу в группах можно представить как работу отдельных биотехнологических лабораторий, каждый из членов которых выполняет конкретную операцию. Отдельные ученики или группы обмениваются картами, постепенно заполняя их. Группа экспертов или один эксперт (это может быть учитель) в конце работы проводят сверку карт, выявляя ошибки-мутации.
Сложность работы будет зависеть от умения использовать учебный материал: таблицы генетического кода, схемы редупликации, транскрипции и трансляции, таблицы комплементарности, свойств генетического кода и др. Занятию можно придать характер лабораторной, практической, самостоятельной или контрольной работы.
Для конкретизации заданий лучше использовать карты небольших полипептидов, например, некоторых пептидных гормонов. С этой целью удобно использовать олигопептиды гормонов вазопрессина и окситоцина, а также метионин- и лейцин-энкефалинов – природных эндорфинов, вырабатывающихся в организме животных и человека (Приложения 1–4). Вазопрессин и окситоцин имеют широкий спектр действия, а эндогенные морфиноподобные вещества привлекают внимание в связи с проблемой наркомании и объяснением наркотического эффекта.
В карты может быть включен материал из раздела «Химия клетки», а именно формулы и свойства аминокислот. В олигопептидах вазопрессина и окситоцина имеются SH-содержащие аминокислоты (цистеин), образующие дисульфидные мостики во вторичной структуре пептида, что может быть отражено в степени сложности задания.
В карты включены кодоны-терминаторы, которые надо записать соответствующими триплетами в цепях ДНК или РНК. Включен также кодон-инициатор аминокислоты метионина, который стоит в этом случае в начале цепи.
Нуклеотиды лидирующей последовательности после кодона-инициатора (и соответствующие аминокислоты) не включены в содержание карт, поскольку не имеют принципиального значения для обработки генетической информации и удаляются из аминокислотной последовательности при процессинге (протеолизе).
Предлагаемая работа учащихся с карточками и заполнением таблиц перевода генетической информации (редупликации, транскрипции, трансляции), написанием формул и обозначений аминокислот может быть рассчитана на 1–2 урока в зависимости от сложности и характера задания.
В заключение занятия выставлются оценки учащимся и формулируются следующие выводы.
Генетическая информация универсальна. Формы жизни с другими генетическими кодами не обнаружены, т.е. генетический код един для всех организмов, и другого генетического кода нет. Этот код имеет достаточно возможностей для описания всего разнообразия белковых молекул.
На картах использованы общепринятые сокращения: иРНК – информационная РНК; кДНК – кодирующая цепь ДНК; комп. ДНК – комплементарная цепь ДНК. Кодон аминокислот выбран произвольно, как один из возможных, что допускается в работе учащихся.
Для проведения занятия используются варианты карт, в которых отсутствует какая-либо одна строка, т.е. имеется 5 вариантов каждой карты. Соответственно, работу можно распределить на конкретное число учащихся и групп. Можно предложить работу и по другим картам для других пептидов, количество которых практически не ограничено.
Приложение 1
Метионин-энкефалин – гормон ядер коры мозга, эндогенный опиоидный пептид, состоит из 5 аминокислот
Аминокислота Комп. ДНК |
Приложение 2
Лейцин-энкефалин – гормон ядер коры мозга, эндогенный опиоидный пептид, состоит из 5 аминокислот
Химическая формула радикала аминокислоты |
|||||||
Аминокислота Комп. ДНК |
Приложение 3
Вазопрессин – антидиуретический гормон – продуцируемый гипофизом, вызывает сокращение гладких мышц, снижает экскрецию воды, состоит из 9 аминокислот с одной дисульфидной связью
Химическая формула радикала аминокислоты |
Эндорфины (эндогенные морфины (от имени древнегреческого бога Морфей -«тот, кто формирует сны»)-группа полипептидных химических соединений по структуре сходных с опиатами (морфиноподобными соединениями),которые естественным путем вырабатываются в нейронах головного мозга и обладают способностью уменьшать боль аналогично опиатам и влиять на эмоциональное состояние.
Эндорфины образуются из липотропинов в ткани мозга и в промежуточной доле гипофиза. Общим типом структуры для этих соединений является тетра-пептидная последовательность на N-конце. Бета-эндорфин образуется из бета-липотропина путем протеолиза. Бета-липотропин образуется из предшественника-прогормона проопикортина (молекулярная масса 29 кДа, 134 аминокислотных остатка).
В передней доле гипофиза молекула предшественника расщепляется на АКТГ и b-липотропин, которые секретируются в плазму. Небольшая часть (около 15%) b-липотропина расщепляется с образованием b-эндорфина. Биосинтез проопикортина в передней доле гипофиза регулируется кортиколиберином гипоталамуса. Известны три различных белка-предшественника опиоидных пептидов: проэнкефалин, проопиомеланокортин и продинорфин.
Природные опиоидные пептиды выделены впервые в 1976 году из мозга млекопитающих. Это были так называемые энкефалины - лейцин-энкефалин и метионин-энкефалин, различающиеся лишь концевым С-остатком.
В начале 70-х годов в разных лабораториях мира обнаружили, что клетки головного мозга имеют рецепторы, связывающие морфин, и только в таком связанном виде он становится активным. Предполагать, что мозг специально заготовил такого рода рецепторы под столь редкий ингредиент как морфин, не было оснований. Возникло подозрение, что функция этих рецепторов состояла в связывании не морфина, но какого-то близкого к нему вещества, вырабатываемого самим организмом. В 1976 году доктор Хьюз в Шотландии извлек это таинственное вещество из мозга морской свинки, у которой сразу же резко снизилась болевая чувствительность. Хьюз назвал вещество энкефалином, что по-гречески означает «из мозга». А профессор Чо Хао Ли в Сан-Франциско извлек из мозга верблюда, а конкретнее из верблюжьего гипофиза еще один внутренний наркотик, оказавшийся в 50 раз сильнее известного морфия. Чо и назвал его эндорфином – «внутренним морфином». В том же 1976 году выделили из крови животных еще два внутренних наркотика, которые были сходны с морфином по составу, но в отличие от растительного морфина не угнетали дыхания и не приводили к наркозависимости. И, наконец, доктор Плесс в Швейцарии синтезировал эндорфин, то есть изготовил его в лаборатории, в пробирке, точно зная химический состав и строение этого загадочного вещества. Из экстрактов тканей гипофиза и гипоталамуса млекопитающих выделены и другие опиоидные пептиды - эндорфины. Все они в N-концевой области обычно содержат остаток энкефалина. Все эндогенные опиоидные пептиды синтезируются в организме в виде крупных белков-предшественников путем протеолиза. Пространственное строение энкефалинов сходно с морфином. Энкефалины и эндорфины обладают обезболивающим действием, снижают двигательную активность ЖКТ, влияют на эмоциональное состояние.
· МСГ - меланоцитстимулирующий гормон;
· ЛПГ - липотропный гормон;
· КППП - кортикотропиноподобный промежуточный пептид;
· АКТГ - адренокортикотропный гормон.
Регуляция секреции
Все продукты расщепления ПОМК производятся в эквимолярных количествах и секретируются в кровь одновременно. Таким образом, невозможно увеличение секреции адренокортикотропного гормона без сопутствующего увеличения секреции бета-липотропного гормона.Продукция ПОМК регулируется факторами, которые образуются в гипоталамусе и паравентрикулярном ядре головного мозга: кортиколиберин, аргининвазопрессин – активируют синтез АКТГ, кортизол - главным ингибитором синтеза кортиколиберина и образования ПОМК, следовательно кортиколиберин, аргининвазопрессин, кортизол будут оказывать влияние на синтез и секрецию β-эндорфина.
Синтез β-эндорфина уменьшается при эндокринных, инфекционных и вирусных заболеваниях, синдроме хронической усталости, усилить синтез можно с помощью физической нагрузки.
Транспорт и периферический метаболизм
Эндорфины синтезируются «впрок» и выделяются в кровь определенными порциями за счет опустошения секреторных везикул. Их уровень в крови возрастает при увеличении частоты выброса гормона из железистых клеток. Поступая в кровь, гормоны связываются с белками плазмы. Обычно лишь 5-10% молекул гормонов находится в крови в свободном состоянии, и только они могут взаимодействовать с рецепторами.
Деградация пептидных гормонов часто начинается уже в крови или на стенках кровеносных сосудов, особенно интенсивно этот процесс идет в почках. Белково-пептидные гормоны гидролизуются протеиназами, а именно экзо- (по концам цепи) и эндопептидазами. Протеолиз приводит к образованию множества фрагментов, некоторые из которых могут проявлять биологическую активность. Многие белково-пептидные гормоны удаляются из системы циркуляции за счет связывания с мембранными рецепторами и последующего эндоцитоза гормон-рецепторного комплекса. Деградация таких комплексов происходит в лизосомах, конечным продуктом деградации являются аминокислоты, которые вновь используются в качестве субстратов в анаболических и катаболических процессах.
Основная мишень эндорфинов - это так называемая опиоидная система (главное ее назначение - защита от стрессорных повреждений, обезболивание и координация работы систем органов и тканей на уровне организма в целом) организма, и опиоидные рецепторы в частности. Эндорфин отвечает за регуляцию деятельность всех внутренних желез, за работу иммунной системы, за уровнем давления, также эндорфин влияет на нервную систему. В головном мозге были обнаружены специфические рецепторы морфина. Эти рецепторы сосредоточены на синаптических мембранах. Наиболее богата ими лимбическая система, от которой зависит эмоциональный ответ. В дальнейшем из мозговой ткани выделили эндогенные пептиды, имитирующие при инъекциях различные эффекты морфина. Эти пептиды, обладающие способностью специфически связываться с опиатными рецепторами, получили название эндорфинов и энкефалинов.
Т.к. рецепторы опиатных гормонов располагаются на наружной поверхности плазматической мембраны, то гормон внутрь клетки не проникает. Гормоны (первые вестники сигнала) передают сигнал посредством второго вестника, роль которого выполняют цАМФ, цГМФ, инозотолтрифосфат, ионы Ca. После присоединения гормона к рецептору следует цепь событий, изменяющих метаболизм клетки.
Физиологически, эндорфины и энкефалины обладают сильнейшим обезболивающим, противошоковым и антистрессовым действием, они понижают аппетит и уменьшают чувствительность отдельных отделов центральной нервной системы. Эндорфины нормализуют артериальное давление, частоту дыхания, ускоряют заживление поврежденных тканей, образование костной мозоли при переломах.
Эндорфины часто возникают в связке с выделением адреналина. При долгих тренировках в организме выделяется адреналин, усиливается боль в мышцах и начинают вырабатываться эндорфины, уменьшающие боль, повышающие реакцию и скорость адаптации организма к нагрузкам.
На что влияют системы эндорфинов:
· - противоболевые эффекты
· -замедление дыхания, сердцебиения–противострессовые эффекты
· - усиление иммунитета
· - регуляция почечного кровотока
· - регуляция деятельности кишечника
· - участие в процессах возбуждения и торможения в нервной системе
· - участие в процессах выработки ассоциативно-диссоциативных связей в нервной системе - регуляция интенсивности обмена веществ
· - чувство эйфории
· - ускоряют заживление поврежденных тканей
· -образование костной мозоли при переломах
Вдобавок к этому эндорфины связаны с терморегуляцией, памятью, липолизом, репродукцией, переживанием удовольствия, деструкцией жиров в организме, антидиурезом, подавлением гипервентиляции в ответ на повышение содержания углекислого газа, и ингибированием синтеза тиротропина и гонадотропина.
Патология
Недостаток эндорфина отмечается при депрессии, в ситуации постоянного эмоционального стресса, он обостряет хронические заболевания, может вызвать синдром хронической усталости. Отсюда и сопровождающее этим состояниям понижение настроение и повышенная восприимчивость к инфекционным заболеваниям.
Выработка эндорфина снижается при некоторых патологиях. В следствие недостатка эндорфинов в организме повышается риск хронических заболеваний, так называемыми «болезнями образа жизни», которые в последнее время являются основной причиной смертности. Болезни образа жизни – это диабет, сердечно-сосудистые, хронические респираторные заболевания, рак и ожирение.
Недостаток эндорфинов выражается в апатии, очень плохом настроении и в конечном итоге приводит человека к депрессии. Любой человек хочет знать: как получать удовольствие от жизни. Ощущение удовольствия у человека появляется при повышении уровня эндорфинов, которые вырабатываются головным мозгом и это химическое соединение похоже на наркотик морфин. Поэтому эндорфин получил такое имя – эндогенный морфин, то есть вырабатываемый самим организмом.
Наиболее тяжелым проявлением является ангедония – болезнь, при которой человек не способен испытывать удовольствие.
Нейрогормоны
Нейрогормоны - вещества, обладающие высокой физиологической активностью, которые вырабатываются в нейросекреторных клетках нервной системы (нейронах).
По механизму действия у них много общего с нейромедиаторами, но нейрогормоны в отличие от них поступают в кровь и другие биологические жидкости организма (лимфа, спинномозговая и тканевая жидкость) и оказывают продолжительное дистанционное действие регулирующего характера.
По химической структуре нейрогормоны являются пептидами (содержат аминокислоты) или катехоламинами (биогенными аминами), их обязательным фрагментом является 3,4-дигидроксифенилаланин (катехол).
Нейрогормоны поддерживают водно-солевой гомеостаз, регулируют тонус гладкой мускулатуры и процессы метаболизма, а также участвуют в регуляции деятельности эндокринных желез. В обшем и целом функция этих веществ - поддержание защитных и приспособительных функций организма.
Синтез нейрогормонов происходит в нейросекреторных клетках гипоталамуса (дофамин, вазопрессин, окситоцин, норадреналин, серотонин и рилизинг-факторы), спинного мозга, эпифиза, надпочечников(хромаффинная ткань мозгового вещества).Синтезируются они также в ганглиях, параганглиях и нервных стволах вегетативной нервной системы(синтез адреналина и норадреналина).
Процесс биосинтеза пептидных нейрогормонов происходит в теле нейрона, в структуре, называемой эндоплазматической сетью; затем в комплексе Гольджи происходит их упаковка в гранулы и оттуда по аксону происходит транспортировка к нервным окончаниям.
Нейрофизиология сна
Нейрофизиологические механизмы сна и его возрастные особенности
Сон - физиологическое состояние, которое характеризуется потерей активных психических связей субъекта с окружающим его миром. Сон является жизненно необходимым для высших животных и человека. Длительное время считали, что сон представляет собой отдых, необходимый для восстановления энергии клеток мозга после активного бодрствования. Однако оказалось, что активность мозга во время сна часто выше, чем во время бодрствования. Было установлено, что активность нейронов ряда структур мозга во время сна существенно возрастает, т.е. сон - это активный физиологический процесс.
Стадии сна
Рефлекторные реакции во время сна снижены. Спящий человек не реагирует на многие внешние воздействия, если они не имеют чрезмерной силы.
Теории сна:
Гуморальная теория , в качестве причины сна рассматривает вещества, появляющиеся в крови при длительном бодрствовании. Доказательством этой теории служит эксперимент, при котором бодрствующей собаке переливали кровь животного, лишенного сна в течение суток. Животное-реципиент немедленно засыпало. Но гуморальные факторы не могут рассматриваться как абсолютная причина возникновения сна. Об этом свидетельствуют наблюдения за поведением двух пар неразделившихся близнецов. У них разделение нервной системы произошло полностью, а системы кровообращения имели множество анастомозов. Эти близнецы могли спать в разное время: одна девочка, например, могла спать, а другая бодрствовала.
Подкорковая и корковая теории сна. При различных опухолевых или инфекционных поражениях подкорковых, особенно стволовых, образований мозга, у больных отмечаются различные нарушения сна - от бессонницы до длительного летаргического сна, что указывает на наличие подкорковых центров сна. При раздражении задних структур субталамуса и гипоталамуса животные засыпали, а после прекращения раздражения они просыпались, что указывает на наличие в этих структурах центров сна.
Химическая теория. По этой теории во время бодрствования в клетках тела накапливаются легко окисляющиеся продукты, в результате возникает дефицит кислорода, и человек засыпает. Мы засыпаем не от того, что отравлены или устали, а чтобы не отравиться и не устать.
Функции сна
o обеспечивает отдых организма.
o играет важную роль в процессах метаболизма. Во время медленного сна высвобождается гормон роста. Быстрый сон: восстановление пластичности нейронов, и обогащение их кислородом; биосинтез белков и РНК нейронов.
o способствует переработке и хранению информации. Сон (особенно медленный) облегчает закрепление изученного материала, быстрый сон реализует подсознательные модели ожидаемых событий. Последнее обстоятельство может служить одной из причин феномена дежавю.
o это приспособление организма к изменению освещённости (день-ночь).
o восстанавливает иммунитет путём активизации T-лимфоцитов, борющимися с простудными и вирусными заболеваниями.
Разновидности сна
При дальнейшем детальном исследовании оказалось, что по своим физиологическим проявлениям сон неоднороден и имеет две разновидности: медленный (спокойный или ортодоксальный) и быстрый (активный или парадоксальный).
При медленном сне наступает уменьшение частоты дыхания и ритма сердцебиений, расслабление мышц и замедление движений глаз. По мере углубления медленного сна общее количество движений спящего становится минимальным. В это время его трудно разбудить. Медленный сон обычно занимает 75 - 80%.
При быстром сне физиологические функции, наоборот, активизируются: учащаются дыхание и ритм сердца, повышается двигательная активность спящего, движения глазных яблок становятся быстрыми (в связи с чем этот вид сна и получил название " быстрый"). Быстрые движения глаз свидетельствуют о том, что спящий в этот момент видит сновидения. И если его разбудить спустя 10 - 15 минут после окончания быстрых движений глаз, он расскажет об увиденном во сне. При пробуждении в период медленного сна человек, как правило, не помнит сновидений. Несмотря на относительно большую активизацию физиологических функций в быстром сне, мышцы тела в этот период бывают расслабленными, и разбудить спящего значительно труднее. Быстрый сон имеет важное значение для жизнедеятельности организма. Если человека искусственно лишить быстрого сна (будить в периоды появления быстрых движений глаз), то, несмотря на вполне достаточную общую продолжительность сна, через пять - семь дней у него наступают психические расстройства.
Чередование быстрого и медленного сна характерно для здоровых людей, при этом человек чувствует себя выспавшимся и бодрым.
Существует еще одна классификация стадий сна:
1. Уравнительная фаза: характеризуется эффектом как на сильные, так и на слабые раздражители.
2. Парадоксальная фаза: сильные раздражители вызывают более слабые ответные реакции, чем слабые раздражители.
3. Ультрадоксальная фаза: положительный раздражитель тормозит, а отрицательный - вызывает условный рефлекс.
4. Наркотическая фаза: общее снижение условно рефлекторной деятельности со значительно более сильным уменьшением рефлексов на слабые раздражители, чем на сильные.
5. Тормозная фаза: полное торможение условных рефлексов
Возрастные особенности:
Детский сон поверхностный и чуткий. Они спят несколько раз в день.
У новорожденных сон занимает большую часть суток, а активированный сон, или сон с подергиваниями (аналог парадоксального сна взрослых), составляет большую часть сна. В первые месяцы после рождения быстро увеличивается время бодрствования, доля парадоксального сна снижается, а медленного увеличивается.
Гигиена сна:
Сон должен иметь достаточную для возраста продолжительность и глубину. Более продолжительное время полагается спать детям с ослабленным здоровьем, выздоравливающим после острых инфекционных заболеваний, повышенной возбудимостью нервной системы, быстро утомляющимся детям. Перед сном следует исключить возбуждающие игры, усиленную умственную работу. Ужин должен быть легким, не позднее 2-1,5 часа до сна. Для сна благоприятны:
· свежий, прохладный воздух в помещении (15-16)
· постель должна быть не мягкой и не жесткой.
· чистое, мягкое без складок и рубцов постельное белье
· лежать лучше на правом боку или спине, что обеспечивает более свободное дыхание, не затрудняет работу сердца.
Следует приучать детей вставать и ложиться в одно и то же время. У ребенка довольно легко образуются условные рефлексы на обстановку сна. Условным раздражителем при этом является время отхода ко сну.
Нейрофизиология ВНС
Впервые понятие вегетативная нервная система было введено в 1801 г. французским врачом А. Беша. Этот отдел ЦНС обеспечивает вегетативные функции организма и включает в себя три компонента:
1) симпатический;
2) парасимпатический;
3) метасимпатический.
К вегетативным относят те функции, которые обеспечивают обмен веществ в нашем организме (пищеварение, кровообращение, дыхание, выделение и др.). К ним относят также обеспечение роста и развития организма, размножения, подготовку организма к неблагоприятным воздействиям. Вегетативная система обеспечивает регуляцию деятельности внутренних органов, сосудов, потовых желез и другие подобные функции. Она регулирует обмен веществ, возбудимость и автономную работу внутренних органов, а также физиологическое состояние тканей и отдельных органов (в том числе головного и спинного мозга), приспосабливая их деятельность к условиям окружающей среды.
Симпатический отдел нервной системы обеспечивает мобилизацию имеющихся у организма ресурсов (энергетических и интеллектуальных) для выполнения срочной работы.Ясно, что это может приводить к нарушениям равновесия в организме.Восстановление равновесия и постоянства внутренней среды организма является задачей нервной парасимпатической системы.Для этого необходимо непрерывно подправлять сдвиги, вызванные влияниями симпатического отдела, восстанавливать и поддерживатьгомеостаз.В этом смысле деятельность этих отделов нервной вегетативной системы в рядереакций проявляется как антагонистическая.
Подгомеостазом в физиологии понимают поддержание в организме постоянства параметров внутренней среды. К ним относится поддержание постоянства состава крови, температуры тела и т.д.
Центры вегетативной нервной системы находятся в мозговом стволе и спинном мозге. В мозговом стволе и в крестцовом отделе спинного мозга расположены центры парасимпатической нервной системы.В среднем мозге находятся центры, регулирующие расширение зрачка и аккомодацию глаза. В продолговатом мозге имеются центры нервной парасимпатической системы, от которых отходят волокна в составе блуждающего, лицевого и языкоглоточного нервов. Эти центры участвуют в осуществлении целого ряда функций, в том числе регулируют деятельность ряда внутренних органов (сердца, желудка, кишечника, печени и т. д.), являются «запускающими» для выделения слюны, слезной жидкости и т. д. Все эти функции осуществляются по рефлекторному принципу (по типу ответной реакции на раздражитель). Ниже будут описаны некоторые из этих рефлексов.
В крестцовых сегментах спинного мозга также находятся центры нервной парасимпатической вегетативной системы. Волокна от них идут в составе тазовых нервов, которые иннервируют органы таза (толстый кишечник, мочевой пузырь, половые органы и пр.).
Центры симпатической нервной системы находятся в грудных и поясничных сегментах спинного мозга. Вегетативные волокна от этих центров отходят в составе передних корешков спинного мозга вместе с двигательными нервами.
Все перечисленные выше центры симпатической и нервной парасимпатической системы подчинены высшему вегетативному центру - гипоталамусу. Гипоталамус, в свою очередь, подвержен влиянию ряда других центров головного мозга. Все эти центры образуют лимбическую систему.Полное описание системы будет приведено в соответствующей теме, а сейчас рассмотрим «работу» периферических отделов нервной вегетативной системы.
По обе стороны позвоночника с брюшной стороны расположены два ствола симпатической нервной системы. Их называют также симпатическими цепочками. Цепочка состоит из отдельных ганглиев, соединенных между собой и спинным мозгом многочисленными нервными волокнами. Каждое волокно, пришедшее к ганглию, иннервирует в ганглии до нескольких десятков нейронов (дивергенция). Благодаря такому устройству симпатические влияния обычно имеют разлитой, генерализованный характер. В свою очередь от этих ганглиев отходят нервы, которые направляются к стенкам сосудов, потовым железам и внутренним органам. Кроме ганглиев пограничного ствола, на некотором удалении от них находятся так называемыепревертебральные ганглии.Самые крупные из них -солнечное сплетениеибрыжеечные узлы.
Большую роль в деятельности симпатической нервной системы играют надпочечники.Они формируются у человека во внутриутробный период за счет миграции нейробластов (еще не дифференцированных нейронов) из нервной трубки в район почек. Там эти клетки образуют на вершинах обеих почек специальный орган - надпочечники. Надпочечники иннервируются симпатическими нервами. Кроме того, они могут активироваться адренокортикотропным гормоном, который выделяется в ответ на стресс из гипофиза и вместе с кровью достигает надпочечников. Под действием этого гормона из надпочечников выбрасывается в кровь смесьадреналинаинорадреналина,которые разносятся по кровяному руслу и вызывают целый ряд симпатических реакций (учащение ритмики сокращений сердца, выделение пота, усиленное кровоснабжение мышц, покраснение кожи и многое другое).
Аксоны симпатических нейронов в периферических синапсах выделяют медиатор адреналин.Молекулы адреналина и норадреналина взаимодействуют с соответствующими рецепторами. Известно два типа таких рецепторов: альфа - и бета-адренорецепторы. В некоторых внутренних органах имеется только один из этих рецепторов, в других - оба. Так, в стенках кровеносных сосудов имеются и альфа-, и бета-адренорецепторы. Соединение симпатического медиатора с альфа-адренорецептором вызывает сужение артериол, а соединение с бета-адренорецептором - расширение артериол. В кишечнике, где имеются оба типа адренорецепторов, медиатор тормозит его деятельность. В сердечной мышце и стенках бронхов находятся только бета-адренорецепторы - симпатический медиатор вызывает расширение бронхов и учащение сердечных сокращений.
Ганглии парасимпатического отдела нервной вегетативной системы в отличие от симпатических расположены в стенках внутренних органов или вблизи них. Нервное волокно(аксон нейрона) от соответствующего парасимпатического центра в мозговом стволе или крестцовом отделе спинного мозга доходит до иннервируемого органа, не прерываясь, и заканчивается на нейронах парасимпатического ганглия. Следующий парасимпатический нейрон находится или внутри органа, или в непосредственной близости от него. Внутриорганные волокна и ганглии образуют сплетения, богатые нейронами, в стенках многих внутренних органов сердца, легких, пищевода, желудка и т.д., а также в железах внешней и внутренней секреции. Анатомическая конструкция парасимпатической части нервной вегетативной системы указывает на то, что влияния на органы с ее стороны носят более локальный характер, чем со стороны нервной симпатической системы.
Медиатором в периферических синапсах нервной парасимпатической системы служит ацетилхолин,к которому имеется два типа рецепторов: М- и Н-холинорецепторы. Это разделение основано на том, что М-холинорецепторы теряют чувствительность к ацетилхолину под влиянием атропина (выделен из гриба родаMuscaris), Н-холинорецепторы - под влиянием никотина.
Влияние симпатической и парасимпатической вегетативной системы на функции организма. В большинстве органов возбуждение симпатической и нервной парасимпатической вегетативной системы вызывает противоположные эффекты. Однако нужно иметь в виду, что эти взаимодействия непростые. Например, парасимпатические нервы вызывают расслабление сфинктеров мочевого пузыря и одновременно сокращение его мускулатуры. Симпатические нервы сокращают сфинктер и одновременно расслабляют мускулатуру. Другой пример: возбуждение симпатических нервов увеличивает ритм и силу сердечных сокращений, а раздражение блуждающего (парасимпатического) нерва снижает ритм и силу сердечных сокращений. Более того, исследования показали, что между этими отделами нервной вегетативной системы существуют не только антагонизм (разнонаправлено), но и синергизм (однонаправлено). Повышение тонуса одного отдела нервной вегетативной системы, как правило, приводит к повышению тонуса и другого отдела. Более того, выяснилось, что есть органы и ткани только с одним типом иннервации. Например, сосуды кожи, мозговой слой надпочечников, матка, скелетные мышцы и некоторые другие имеют только симпатическую иннервацию, а слюнные железы иннервируются лишь парасимпатическими волокнами.
Вегетативные рефлексы. Эти рефлексы многочисленны. Они участвуют во многих регуляциях организма человека. При осуществлении вегетативных рефлексов влияния передаются по соответствующим нервам (симпатическим или парасимпатическим) из ЦНС. В медицинской практике наибольшее значение придают висцеро-висцеральным (от одного внутреннего органа на другой), висцеро-дермальным (от внутренних органов на кожу) и дермо-висцеральным (от кожи на внутренние органы) рефлексам.
К числу висцеро-висцеральных относят рефлекторные изменения сердечной деятельности, тонуса сосудов, кровенаполнения селезенки при повышении или понижении давления в аорте, каротидном синусе или легочных сосудах. Например, благодаря включению такого рефлекса происходит остановка сердца при раздражении органов брюшной полости. Висцеро-дермальные рефлексы возникают при раздражении внутренних органов и проявляются в изменении чувствительности соответствующих участков кожи (в соответствии с тем, какой орган при этом раздражается), потоотделении, реакции сосудов. Дермо-висцеральные рефлексы проявляются в том, что при раздражении определенных участков кожи изменяется функционирование соответствующих внутренних органов. Собственно на механизме этих рефлексов основано применение в лечебных целях согревания или охлаждения определенных участков кожи, например при болях во внутренних органах.
Вегетативные рефлексы часто используются врачами для суждения о функциональном состоянии нервной вегетативной системы. Например, в клинике часто исследуют рефлекторные изменения сосудов при механическом раздражении кожи (например, при проведении по коже тупым предметом). У здорового человека при этом возникает кратковременное побледнение раздражаемого участка кожи (белый дермографизм, derma- кожа). При высокой возбудимости нервной вегетативной системы на месте раздражения кожи появляется красная полоса, окаймленная бледными полосами суженых сосудов (красный дермографизм), а при еще более высокой чувствительности - отек кожи в этом месте. Часто в клинике используют функциональные вегетативные пробы для суждения о состоянии нервной вегетативной системы. Например, ортостатическая реакция: при переходе из положения, лежа в положение стоя происходит повышение кровяного давления и учащение сердечных сокращений. Характер изменения кровяного давления и сердечной деятельности при этой пробе может служить диагностическим признаком заболевания системы управления кровяным давлением. Другой пример, глазо-сердечная реакция (рефлекс Ашнера): при надавливании на глазные яблоки происходит кратковременное урежение сердечных сокращений.
Вегетативные центры . В продолговатом мозге расположены нервные центры, тормозящие деятельность сердца (ядра блуждающего нерва). В ретикулярной формации продолговатого мозга находитсясосудодвигательный центр,состоящий из двух зон: прессорной и депрессорной. Возбуждение прессорной зоны приводит к сужению сосудов, а возбуждение депрессорной зоны - к их расширению. Сосудодвигательный центр и ядра блуждающего нерва постоянно посылают импульсы, благодаря которым поддерживается постоянный тонус: артерии и артериолы постоянно несколько сужены, а сердечная деятельность замедлена.
В продолговатом мозге находится дыхательный центр,который, в свою очередь, состоит из центров вдоха и выдоха. На уровне моста находится центр дыхания (пневмотаксический центр) более высокого уровня, который приспосабливает дыхание к изменениям физической нагрузки. Дыхание у человека может управляться также произвольно со стороны коры больших полушарий, например во время речи.
В продолговатом мозге находятся центры, возбуждающие секрецию слюнных, слезных и желудочных желез, выделение желчи из желчного пузыря, секрецию поджелудочной железы. В среднем мозге под передними буграми четверохолмия находятся парасимпатические центры аккомодации глаза и зрачкового рефлекса. Все перечисленные выше центры симпатической и парасимпатической системы подчинены высшему вегетативному центру - гипоталамусу.
Роль гипоталамуса в регуляции вегетативных функций. Влияние на симпатическую и парасимпатическую регуляцию позволяет гипоталамусу воздействовать на вегетативные функции организма гуморальным и нервным путями. Ранее уже разбиралось, что раздражение ядер передней группы сопровождается парасимпатическими эффектами. Раздражение ядер задней группы вызывает симпатические эффекты в работе органов. Стимуляция ядер средней группы приводит к снижению влияний симпатического отдела автономной нервной системы. Указанное распределение функций гипоталамуса не абсолютно. Все структуры гипоталамуса способны в разной степени вызывать симпатические и парасимпатические эффекты. Следовательно, между структурами гипоталамуса существуют функциональные взаимодополняющие, взаимокомпенсирующие отношения.
В целом за счет большого количества связей, полифункциональности структур гипоталамус выполняет интегрирующую функцию вегетативной, соматической и эндокринной регуляции, что проявляется и в организации его ядрами ряда конкретных функций. Так, в гипоталамусе располагаются центры гомеостаза, теплорегуляции, голода и насыщения, жажды и ее удовлетворения, полового поведения, страха, ярости, регуляции цикла бодрствование-сон. Все эти центры реализуют свои функции путем активации или торможения вегетативного отдела нервной системы, эндокринной системы, структур ствола и переднего мозга.
Гипоталамус, в свою очередь, подвержен влиянию ряда высших центров головного мозга, включая кору.
Таким образом, вегетативная нервная система обладает рядом анатомических и физиологических особенностей, которые определяют механизмы ее работы:
Анатомические свойства
1. Трехкомпонентное расположение нервных центров. Низший уровень симпатического отдела представлен боковыми рогами с VII шейного по III–IV поясничные позвонки, а парасимпатического – крестцовыми сегментами и стволом мозга. Высшие подкорковые центры находятся на границе ядер гипоталамуса (симпатический отдел – задняя группа, а парасимпатический – передняя). Корковый уровень лежит в области шестого-восьмого полей Бродмана (мотосенсорная зона), в которых достигается точечная локализация поступающих нервных импульсов. За счет наличия такой структуры вегетативной нервной системы работа внутренних органов не доходит до порога нашего сознания.
2. Наличие вегетативных ганглиев. В симпатическом отделе они расположены либо по обеим сторонам вдоль позвоночника (симпатическая нервная цепочка), либо входят в состав сплетений. Таким образом, дуга имеет короткий преганглионарный и длинный постганглионарный путь. Нейроны парасимпатического отдела находятся в ганглии, расположенном вблизи рабочего органа или в его стенке, поэтому дуга имеет длинный преганглионарный и короткий постганглионарный путь.
1. Особенности функционирования вегетативных ганглиев. Наличие феномена мультипликации (одновременного протекания двух противоположных процессов – дивергенции и конвергенции). Дивергенция – расхождение нервных импульсов от тела одного нейрона на несколько постганглионарных волокон другого. Конвергенция – схождение на теле каждого постганглионарного нейрона импульсов от нескольких преганглионарных. Это обеспечивает надежность передачи информации из ЦНС на рабочий орган. Увеличение продолжительности постсинаптического потенциала, наличие следовой гиперполяризации и синаптической задержки способствуют передаче возбуждения со скоростью 1,5–3,0 м/с. Однако импульсы частично гасятся или полностью блокируются в вегетативных ганглиях. Таким образом они регулируют поток информации из ЦНС. За счет этого свойства их называют вынесенными на периферию нервными центрами, а вегетативную нервную систему – автономной.
2. Особенности нервных волокон. Преганглионарные нервные волокна относятся к группе В и проводят возбуждение со скоростью 3-18 м/с, постганглионарные – к группе С. Они проводят возбуждение со скоростью 0,5–3,0 м/с. Так как эфферентный путь симпатического отдела представлен преганглионарными волокнами, а парасимпатического – постганглионарными, то скорость передачи импульсов выше у парасимпатической нервной системы.
В целом, симпатическая нервная система осуществляет адаптационно-трофическую функцию, включаясь в работу при физических нагрузках, эмоциональных реакциях, стрессах, болевых воздействий, кровопотерях. Она обеспечивает приспособление организма к меняющимся условиям среды существования.
Парасимпатическая нервная система является антагонистом симпатической и выполняет гомеостатическую и защитную функции, регулирует опорожнение полых органов. Гомеостатическая роль носит восстановительный характер и действует в состоянии покоя. Это проявляется в виде уменьшения частоты и силы сердечных сокращений, стимуляции деятельности желудочно-кишечного тракта при уменьшении уровня глюкозы в крови и т. д.