ЛЕКЦИЯ 15. ЭВОЛЮЦИЯ ОНТОГЕНЕЗА, СООТНОШЕНИЕ ОНТО - И
Вопросы для закрепления материала.
1. Что такое видообразование?
2. Основные пути и способы видообразования.
3. Принцип основателя, из чего вытекает его действие?
РАЗДЕЛ 4 ПРОБЛЕМЫ МАКРОЭВОЛЮЦИИ.
1 Понятие макроэволюции, сходства и различия микро - и макроэволюции.
2 Общие представления об онтогенезе и эволюции онтогенеза.
3 Биогенетический закон, рекапитуляция, учение о филэмбриогенезах.
4 Принципы преобразования органов и функций.
1 Понятие макроэволюции, сходства и различия микро- и макроэволюции. Во времена Ч. Дарвина и в последующую эпоху расцвета его эволюционного учения почти ничего не было известно о двух таких основных явлениях жизни и наиболее общих характеристиках живых организмов на Земле как наследственность и изменчивость. Явления наследственности и изменчивости живых организмов были известны людям, но научных представлений о характере и механизмах наследования признаков и их изменчивости не было. Лишь после развития современной генетики с начала XX столетия появилась возможность положить достаточно точные сведения об основных закономерностях наследования и изменчивости признаков и свойств организмов в основу нового – микроэволюционного этапа изучения эволюционного процесса. В эпоху развития классического дарвинизма построение эволюционной теории проводилось на основании результатов, полученных в самых разнообразных разделах биологии, исследователями, работавшими с применением лишь описательных и сравнительных методов. Это позволило создать достаточно развернутую картину основных этапов и явлений эволюционного процесса, а также создать в первом приближении общую схему филогенеза живых организмов. Такое классическое направление в развитии эволюционных идей является изучением процесса макроэволюции. Макроэволюционный процесс, в отличие от микроэволюционного, охватывает большие отрезки времени, обширные территории и все (включая высшие) таксоны живых организмов, а также все основные общие и специальные явления эволюции.
Данные систематики, палеонтологии, биогеографии, сравнительной анатомии, молекулярной биологии и других биологических дисциплин дают возможность с большой точностью восстанавливать ход эволюционного процесса на любых уровнях выше вида. Совокупность этих данных составляет основу филогенетики – дисциплины, посвященной выяснению особенностей эволюции крупных групп органического мира. Сопоставление хода эволюционного процесса в разных группах, при неодинаковых условиях внешней среды, в разном биотическом и абиотическом окружении и т.д. позволяет выделить общие, характерные для большинства групп особенности исторического развития. На макроэволюционном уровне внутри вновь возникших форм без какого-либо перерыва продолжается процесс микроэволюции. Нарушается лишь характер отношений между вновь возникшими видами. Теперь они могут вступать в межвиловые отношения. Эти отношения способны повлиять на эволюционное событие лишь изменяя давление и направление действия элементарных эволюционных факторов, то есть через микроэволюционный уровень. Макроэволюционные явления, имея огромные масштабы времени, исключают возможность их непосредственного экспериментального исследования. Значит, их результаты оказываются понятными лишь с позиций механизма осуществления эволюции – с позиций микроэволюции. На микроэволюционном (внутривидовом) уровне при изучении эволюции оказалось возможным применить точные экспериментальные подходы, которые помогли выяснить роль отдельных эволюционных факторов, сформулировать представления об элементарной эволюционной единице, элементарном эвалюционном материале и явлении.
В 30-е годы XX в. в результате интенсивного развития популяционной генетики возникла объективная возможность более глубокого познания механизма возникновения новых признаков (адаптаций) и механизма возникновения видов, чем это было возможно ранее, лишь на основе наблюдений в природе. Существенным моментом при этом оказалась возможность прямого эксперимента при изучении механизма эволюции: благодаря использованию быстро размножающихся видов организмов стало возможным моделировать эволюционные ситуации и наблюдать протекание эволюционного процесса. За короткое время в изучаемых совокупностях стало возможным наблюдать значимые эволюционные изменения – вплоть до возникновения репродуктивной изоляции исходной формы.
2 Общие представления об онтогенезе и эволюции онтогенеза. Онтогенез (гр. ontos - сущее, genesis - происхождение) – это индивидуальное развитие организмов, в процессе которого из оплодотворенного яйца (при партеногенезе из неоплодотворенного) развивается взрослый организм. У простейших онтогенез осуществляется в пределах клеточной организации. Термин ввел Э. Геккель в 1866 г. Онтогенез представляет собой неотъемлемое свойство жизни, как эволюция, и её продукт. Процесс онтогенеза представляет собой реализацию генетической информации. Онтогенез – это предопределенный процесс, и, в отличие от эволюции, является развитием по программе (ею служит генотип данной особи), развитием, направленным к определенной конечной цели, которой является достижение половозрелости и размножения. В то же время, усложнение организации в ряду поколений – есть результат процесса эволюции. Чем сложнее организация взрослого организма, а это является отражением эволюции, тем сложнее и длительнее процесс его онтогенеза. Так оказываются тесно взаимосвязанными индивидуальное развитие и эволюция (рисунок 4). Онтогенез состоит из этапов (этапность – еще одна особенность онтогенеза): эмбриональный этап, постэмбриональное развитие и жизнь взрослого организма. Крупные этапы (периоды) развития можно подразделить на более дробные стадии, как в эмбриональном развитии позвоночных – бластулы, гаструлы, нейрулы. Стадию дробления, в свою очередь, можно
разделить на стадии двух, четырех, восьми и более бластомеров. В результате представление об этапности онтогенеза теряется и вырисовывается вполне плавный процесс индивидуального развития. Как видно, онтогенез представляет собой упорядоченную последовательность процессов (А.С. Северцов, 1987 , 2005) .
Эволюционные изменения связаны не только с образованием и вымиранием видов, преобразованием органов, но и с перестройкой онтогенетического развития. Без изменения отдельных стадий в онтогенезе немыслим филогенез. Филогенез (гр. phyle – племя, род, вид, genesis – происхождение) – историческое развитие органического мира, различных систематических групп, отдельных органов и их систем. Различают филогенез групп животных, растений, филогенез органов.
В ходе эволюции наблюдается интеграция организма – установление все более тесных динамических связей между его структурами. Этот принцип отчасти отражается и в ходе эмбриогенеза. Эволюция жизни сопровождается постепенным усилением дифференциации и целостности онтогенеза, увеличением устойчивости онтогенеза в ходе эволюции жизни. Организм в онтогенезе ни на одной из стадий развития не является мозаикой частей, органов или признаков. Морфологическая и функциональная целостность организма в его жизненных проявлениях не вызывает никаких сомнений. Еще Аристотель при сравнении различных организмов установил единство их строения и обосновал учение о морфологическом сходстве,
выражающимся в положении и строении органов у разных животных (современная гомология органов), развил представление о соотношении органов, о взаимозависимостях в их строении. Большое значение в истории вопроса о взаимозависимостях частей организма имели взгляды Ж. Кювье. По его представлениям, как было отмечено ранее, организм является целостной системой, строение которой определяется ее функцией; отдельные части и органы находятся во взаимной связи, их функции согласованы и приспособлены к известным условиям внешней среды (принцип корреляции и принцип условий существования). На приспособления организма к внешней среде и усложнение его строения как наиболее яркую характеристику эволюционного процесса указывал Ч. Дарвин. Он отмечал, что координация частей есть результат исторического процесса приспособления организма к условиям жизни. В дальнейшем многие ученые подчеркивали тот факт, что организм всегда развивается как целое. Имеется очень сложная система связей, объединяющих все части развивающегося организма в одно целое. Благодаря наличию этих связей, выступающих в роли основных, внутренних факторов индивидуального развития, из яйца образуется не случайный хаос органов и тканей, а планомерно построенный организм с согласованно функционирующими частями. Вся целесообразность реакций организма при нормальном контакте одной его развивающейся части с другой есть результат исторического развития данных соотношений, т.е. результат эволюции всего механизма индивидуального развития.
Способы (пути) усовершенствования онтогенеза в процессе эволюции: 1) возникновение новых стадий, вызванное формированием комплексов адаптаций, обеспечивающих выживание организма и достижение половозрелости, приводящих к усложнению онтогенеза; 2) исключение определенных стадий и прекращение идущей на них элиминации, сопровождающееся вторичным упрощением.
Эмбрионизация, автономизация, канализация онтогенеза. Э мбрионизация, автономизация, а также рационализация являются результатами эволюции онтогенеза. Эмбрионизация – это путь развития, когда онтогенез проходит под защитой яйцевых оболочек, дольше изолирован от внешней среды, имеет меньшую сложность организации эмбриональных стадий. Путем эмбрионизации шла эволюция от споровых растений к голосеменным и от них к покрытосеменным. Переход от личиночного развития (у беспозвоночных, рыб, земноводных) к откладке крупных, защищенных плотными оболочками яиц (у рептилий, птиц), к внутриутробному развитию, живорождению (у млекопитающих) – результат эмбрионизации. Эмбрионизация проявляется в заботе о потомстве – насиживание яиц, вынашивание детенышей, строительство гнезд, передача индивидуального опыта потомству, защита семени завязью, плодом. Она проявляется в упрощении циклов развития – это переход от развития с метаморфозом к прямому развитию, к неотении. Автономизация проявляется в возрастании независимости онтогенеза от внешних и внутренних воздействий, этот путь эволюции создает преемственность форм в эволюционном процессе. Автономизация индивидуального развития обусловлена действием стабилизирующего отбора. Рационализация заключается в усовершенствовании процесса с помощью его упрощения.
Одна из тенденций эволюции ведет к канализации онтогенеза (И.И. Шмальгаузен, К. Уоддингтон и др.). Главный действующий агент при этом – естественный отбор, выступающий в виде канализирующего отбора. Он определяет возникновение «стандартного» фенотипа в самых разнообразных, колеблющихся условиях внутренней и внешней среды.
В общем, эволюция онтогенеза имеет некоторые особенности, идет определенными путями, приводит к важным результатам, находится во взаимосвязи с филогенезом, что отражено в биогенетическом законе (будет рассмотрен далее).
Значение корреляций и координаций. В процессе онтогенеза имеет место дифференциация организма (разделение целого на части) и его интеграция (объединение частей в единое целое). Это осуществляется одним и тем же механизмом – взаимодействием развивающихся зачатков. В онтогенезе последовательно накладываются друг на друга три волны коррелятивных зависимостей: корреляции геномные, морфогенетические, эргонтические. Геномные корреляции – корреляции, основанные на взаимодействии генов, выражающемся в явлениях сцепления генов и плейотропии (действие одного гена на формирование разных признаков). Морфогенетические корреляции – взаимодействия развивающихся зачатков, основанные на функционировании генов. Любой дифференцировке развивающихся зачатков предшествует генетическая, выражающаяся в дифференциальной репрессии и дерепрессии генов. Эргонтические корреляции – коррелятивные изменения органов относительно друг друга. Примером может служить усиленное развитие костей, образование на них гребней в местах прикрепления мышц.
Координации означают взаимозависимости в процессах филогенетических преобразований. Исторически они развиваются на базе наследственных изменений частей, связанных системой корреляций, т.е. неизбежным изменением последних, или на другой основе – наследственного изменения частей, непосредственно корреляциями не связанных. Если организм представляет собой согласованное целое, то и в изменениях своего строения в процессе эволюции он должен сохранять значение согласованного целого. Это предполагает координированное изменение частей и органов. Примеров координаций много. Это зависимости в изменениях величины, формы черепной коробки и величины и формы головного мозга – в процессе эволюции выработано очень точное соответствие формы и величины этих органов. Координацией является соотношение между относительной величиной глаз и формой черепа – увеличение размеров глаз связано с увеличением размеров глазниц. К координациям относятся зависимости между степенью развития органов чувств (обоняния, осязания и др.) и степенью развития соответствующих центров и областей головного мозга. Имеются координации между внутренними органами как зависимость между прогрессивным развитием грудной мышцы, сердца и легких у птиц. Очень простая биологическая координация проявляется между длиной передних и задних конечностей у копытных.
3 Биогенетический закон, рекапитуляция, учение о филэмбриогенезах. Впервые взаимосвязь онтогенеза и филогенеза раскрыл К. Бэр в ряде положений, которым Ч. Дарвин дал обобщенное название «Закон зародышевого сходства». В зародыше потомков, писал Ч. Дарвин, мы видим «смутный портрет» предков. Большое сходство разных видов в пределах типа выявляется уже на ранних стадиях эмбриогенеза. Следовательно, по индивидуальному развитию можно проследить историю данного вида. В 1864 г. Ф. Мюллер сформулировал положение о том, что филогенетические преобразования связаны с онтогенетическими изменениями и что эта связь проявляется двумя путями. В первом случае индивидуальное развитие потомков идет аналогично развитию предков лишь до появления в онтогенезе нового признака. Изменение процессов морфогенеза обусловливает повторение в эмбриональном развитии истории предков лишь в общих чертах. Во втором случае потомки повторяют все развитие предков, но к концу эмбриогенеза добавляются новые стадии. Повторение признаков взрослых предков в эмбриогенезе потомков Ф. Мюллер назвал рекапитуляцией. Работы Ф. Мюллера послужили основой для формулировки Э. Геккелем (1866 г.) биогенетического закона, согласно которому «онтогенез есть краткое и быстрое повторение филогенеза». Основа биогенетического закона, как и рекапитуляции, заключается в эмпирической закономерности, отраженной в законе зародышевого сходства К. Бэра. Суть его заключается в следующем: самая ранняя стадия сохраняет значительное сходство с соответствующими стадиями развития родственных форм. Таким образом, процесс онтогенеза представляет собой известное повторение (рекапитуляцию) многих черт строения предковых форм, на ранних стадиях развития – более отдаленных предков, а на более поздних – более родственных форм.
В настоящее время явление рекапитуляции трактуют более широко, как последовательность стадий эмбриогенеза, отражающую историческую последовательность эволюционных преобразований данного вида. Рекапитуляция объясняется сложностью корреляций, в особенности на ранних стадиях развития, и трудностью перестройки системы взаимозависимостей между формообразовательными процессами. Коренные нарушения эмбриогенеза сопровождаются летальными последствиями. Рекапитуляции оказываются наиболее полными у тех организмов и в тех системах органов, в которых морфогенетические зависимости достигают особо большой сложности. Поэтому, лучшие примеры рекапитуляции имеются в онтогенезе высших позвоночных.
Филэмбриогенезы – это изменения, возникающие в разные моменты онтогенеза, ведущие к филогенетическим преобразованиям (филэмбриогенезы – эволюционные преобразования организмов путем изменения хода эмбрионального развития их предков, приводящие к появлению новых признаков у взрослых организмов). Создателем теории филэмбриогенеза является А.Н. Северцов. Согласно его представлениям онтогенез весь перестраивается в процессе эволюции. Новые изменения нередко происходят на последних стадиях формообразования. Усложнения онтогенеза путем прибавления, или надставки стадий, называются анаболией. Надставка добавляет новые черты строения органов, происходит дальнейшее их развитие. В этом случае имеются все предпосылки для повторения в онтогенезе исторических этапов развития данных частей у далеких предков. Поэтому именно при анаболии соблюдается основной биогенетический закон. На поздних стадиях развития происходят обычно изменения в строении скелета позвоночных, возникают изменения в дифференцировке мышц, в распределении кровеносных сосудов. Путем анаболии возникает четырехкамерное сердце у птиц и млекопитающих. Перегородка между желудочками представляет надставку, она формируется на последних стадиях развития сердца. Как анаболии появились рассеченные листья у растений. Онтогенез может, однако изменяться и на средних стадиях развития, отклоняя при этом все позднейшие стадии от прежнего пути. Такой путь изменения онтогенеза называется девиация. Девиация ведет к перестройке органов, существовавших у предков. Примером девиации является формирование роговых чешуй рептилий, которые первоначально формируются, как плакоидные чешуи акуловых рыб. Затем у акуловых начинают интенсивно развиваться соединительнотканные образования в сосочке, а у рептилий – эпидермальная часть. Путем девиации формируются колючки, происходит преобразование побегов в клубень или луковицу. Кроме отмеченных путей (способов) изменения онтогенеза, возможно также изменение самих зачатков органов или их частей – этот путь называется архаллаксис. Хорошим примером его является развитие волоса у млекопитающих. Путем архаллаксисов изменяется число позвонков, число зубов у животных и др. Архаллаксис имел место при удвоении числа тычинок, происхождении однодольности у растений. Рассмотренные эволюционные изменения в онтогенезе отражены на рисунках 4, 5.
Основное значение теории филэмбриогенезов заключается в том, что она объясняет механизм эволюции онтогенеза, механизм эволюционных преобразований органов, возникновение новых признаков в онтогенезе, объясняет факт рекапитуляции. Филэмбриогенез представляет собой результат наследственной перестройки формообразовательных аппаратов, комплекс наследственно обусловленных адаптивных преобразований онтогенеза.
Целостность организма, мультифункциональность. Положение о целостности организма довольно подробно рассмотрено выше. Однако следует отметить, что одновременно с этой особенностью для организма характерна автономность его отдельных органов. Это положение находит подтверждение в явлении мультифункциональности и возможности качественных и количественных изменений функций. Филогенетические преобразования органов и их функций имеют две предпосылки: для каждого органа характерна мультифункциональность, а для функций - способность изменяться количественно. Эти категории и лежат в основе принципов эволюционного изменения органов и их функций. Мультифункциональность органов заключается в том, что каждый орган несет, кроме характерной для него главной функции, еще ряд второстепенных. Так, главная функция листа – фотосинтез, но, кроме того, он выполняет функции отдачи и поглощения воды, запасающего органа, органа размножения и т.д. Пищеварительный тракт у животных – это не только орган пищеварения, но и важнейшее звено в цепи органов внутренней секреции, важное звено в лимфатической и кровеносной системах. Одна и та же функция может проявляться у организмов с большей или меньшей интенсивностью, поэтому любые формы жизнедеятельности имеют не только качественную, но и количественную характеристику. Функция бега,
например, выражена сильнее у одних видов млекопитающих и слабее – у других. По любому из свойств всегда существуют количественные различия между особями вида. Любая из функций организма количественно меняется в процессе индивидуального развития особи.
4 Принципы преобразования органов и функций. Известно более полутора десятков способов эволюции органов и функций, принципов их преобразования. Главнейшими из них являются приведенные ниже.
1) Смена функций: при изменении условий существования главная функция может терять значение, а какая-либо из второстепенных – приобрести значение главной (разделение у птиц желудка на два – железистый и мускульный).
2) Принцип расширения функций: нередко сопровождает прогрессивное развитие (хобот слона, уши африканского слона).
3) Принцип сужения функций (ласты кита).
4) Усиление, или интенсификация функций: связано с прогрессивным развитием органа, большей его концентрацией (прогрессивное развитие головного мозга млекопитающих).
5) Активация функций – превращение пассивных органов в активные (ядовитый зуб у змей).
6) Иммобилизация функций: преобразование активного органа в пассивный (потеря подвижности верхней челюсти в ряду позвоночных).
7) Разделение функций: сопровождается разделением органа (например, мышцы, части скелета) на самостоятельные отделы. Примером может служить разделение непарного плавника рыб на отделы и связанные с этим изменения функций отдельных частей. Передние отделы - спинные и анальные плавники становятся рулями, направляющими движение рыбы, хвостовой отдел – основным двигательным органом.
8) Фиксация фаз: стопоходящие животные при хождении и беге приподымаются на пальцах, через эту фазу устанавливается пальцехождение копытных.
9) Субституция органов: в этом случае какой-либо орган утрачивается и его функцию выполняет другой (замена хорды позвоночником).
10) Симиляция функций: ранее различные по форме и функциям органы становятся подобными друг другу (у змей сходные сегменты тела возникли в результате симиляции их функций).
11) Принципы олигомеризации и полимеризации. При олигомеризации умень- шается количество гомологичных и функционально однотипных органов, что сопровождается принципиальными изменениями коррелятивных связей между органами и системами. Так, тело кольчатых червей состоит из многих повторяющихся сегментов, у насекомых их количество значительно уменьшено, а у высших позвоночных одинаковых сегментов тела нет совсем. Полимеризация сопровождается умножением числа органелл и органов. Она имела большое значение в эволюции простейших. Такой путь развития приводил к появлению колоний, а затем и к появлению многоклеточности. Увеличение числа однородных органов происходило и у многоклеточных животных (как у змей). В ходе эволюции олигомеризация сменялась полимеризацией и наоборот.
Следует отметить, что любой организм – координированное целое, в котором отдельные части находятся в сложном соподчинении и взаимозависимости. Как было отмечено выше, взаимозависимость отдельных структур (корреляция) хорошо изучена в процессе онтогенеза, также как и корреляции, проявляющиеся в процессе филогенеза и обозначаемые как координации. Сложность эволюционных взаимоотношений органов и систем видна при анализе принципов преобразования органов и функций. Эти принципы позволяют глубже представить эволюционные возможности преобразования той или иной организации в разных направлениях, несмотря на ограничения, накладываемые корреляциями.
Скорость эволюции отдельных признаков и структур, а также скорость эволюции форм (видов, родов, семейств, отрядов и т. д.) определяют темпы эволюции в целом.. Последнее необходимо принимать во внимание в практической деятельности человека. Например, применяя химические препараты, следует знать, как быстро у того или другого вида может возникнуть устойчивость к препаратам: лекарственным - у человека, инсектицидам - у насекомых и др. Скорость эволюции отдельных признаков в популяциях, так же как и скорость эволюции целых структур и органов, зависит от многих факторов: числа популяций внутри вида, плотности особей в популяциях, продолжительности жизни поколений. Любые факторы первично окажут свое воздействие на скорость изменения популяции и вида посредством изменения давления элементарных эволюционных факторов.
Антропный принцип и глобальный эволюционизм
Итак, несколько десятилетий назад Б. Картер выдвинул так называемый антропный принцип (АП), декларирующий наличие взаимосвязи между параметрами Вселенной и существованием в ней разума. Формальный толчок началу дискуссии о месте человека во Вселенной дало обсуждение проблемы совпадения больших чисел – странной численной взаимосвязи параметров микромира (постоянной Планка, заряда электрона, размера нуклона) и глобальных характеристик Метагалактики (ее массы, размера, времени существования). Эта проблема поставила вопрос: а на сколько случайны параметры нашего мира, насколько они взаимосвязаны между собой, и что произойдет при их незначительном изменении? Анализ возможного варьирования основных физических параметров показал, что даже незначительное их изменение приводит к невозможности существования нашей Метагалактики в наблюдаемой форме и не совместимо с появлением в ней жизни и, соответственно разума.
Взаимосвязь между параметрами Вселенной и появлением в ней разума была выражена Картером в двух формулировках – сильной и слабой. Слабый АП лишь констатирует, что имеющееся во Вселенной условия не противоречат существованию человека: «Наше положение во Вселенной с необходимостью является привилегированным в том смысле, что оно должно быть совместимо с нашим существованием как наблюдателей» (Картер Б. Совпадение больших чисел и антропологический принцип в космологии // Космология: теория и наблюдения. М., 1978). Сильный АП выдвигает более жесткую взаимосвязь параметров Вселенной с возможностью и необходимостью появления в ней разума: «Вселенная (и, следовательно, фундаментальные параметры, от которых она зависит), должна быть такой, чтобы в ней на некотором этапе эволюции допускалось существование наблюдателей» .
Можно сформулировать два крайних предположения обосновывающих АП:
1) разум в нашей Метагалактике явление абсолютно случайное, которое стало возможным лишь благодаря маловероятному, но реализованному совпадению многих независимых физических параметров;
2) наличие биологической и социальной форм движения закономерное следствие развития Вселенной, а все ее физические характеристики взаимосвязаны и взаимообусловлены таким образом, что с необходимостью вызывают появление разума.
Вероятностная гипотеза объяснения АП, низводит его с позиции глобальной эвристической идеи взаимосвязи появления разумной жизни и строения Вселенной к абсолютно случайному совпадению, чем, по сути, выводит его за рамки научного рассмотрения. Но вероятностную гипотезу можно рассматривать лишь как терминологическую – вместо вопроса о возможности появления наблюдателя во Вселенной ставится вопрос о его возможном появлении в совокупности Вселенных. А исходя из понятия категории «Вселенная» мы должны признать, что оно абсолютно тождественно термину «совокупность Вселенных». Можно лишь различать варианты единично- или множественно-определенной Вселенной, что естественно, не существенно с точки решения проблемы о возможности и необходимости появления разума во Вселенной.
Гипотеза осциллирующей Вселенной, в качестве возможного объяснения АП, не отличается от предположения множественности Вселенных. Численная бесконечность заменяется временной, опять же сводя объяснение АП к абсолютно случайному моменту.
Другая соломинка, предлагаемая теорией осциллирующей Вселенной для формальной поддержки АП – это констатация возможности передачи информации о параметрах Вселенной через момент сингулярности. То есть предполагается, что физические характеристики Метагалактики не случайны и их значения обусловлены предыдущим циклом ее развития (до сингулярности). Вполне логично и возможно. Но опять же эта гипотеза, отодвигая решение проблемы к гипотетическим предыдущим этапам истории Вселенной, не приближает нас к научной интерпретации АП.
Рассмотрим лишь второй, не вероятностный вариант обоснования АП, который непосредственно стыкуется с широко обсуждаемой в настоящее время проблемой глобального эволюционизма. В общих чертах эволюционный подход к проблеме АП можно сформулировать так: Вселенная находится в непрерывном процессе эволюции, и появление жизни и разума закономерный результат этого процесса . При достаточно убедительном построении теории глобального эволюционизма АП должен свестись к ее частному моменту – констатации факта, что разум есть необходимый этап эволюции Вселенной.
Эволюция. На данный момент термин «эволюция» используется либо в узком смысле – для описания процессов формирования и развития биологических систем, либо в самом широком смысле, когда речь идет вообще о появление новых свойств, определений, структурных образований. Естественно, что при использовании далее термина «эволюция» в обсуждении проблемы глобального эволюционизма, мы не будем иметь в виду лишь его частное биологическое значение. И конечно, чтобы избежать терминологических проблем, следует серьезно обратить внимание на расширенную трактовку термина «эволюция», например, при описании астрономических или геологических процессов. Часто мы сталкиваемся с терминологической проблемой использования категории «эволюция», вместо категорий «развитие» и «изменение», хотя различение этих трех категорий вполне вписывается в естественное понимание значения этих слов.
Так, говоря об эволюции, мы имеем в виду появление принципиально новых, уникальных определений (параметров, категорий, систем, и т.д.), не имеющихся ранее. Причем, для того чтобы однозначно различить категорию «эволюция» от категории «развитие», новизна определений должна быть принципиальной не только для рассматриваемой частной системы, но для Мира в целом.
Развитие констатирует появление новых, ранее не присущих некой системе признаков, но не являющихся уникальными для Мира в целом. Так появление разума является эволюционным явлением, но изменения происходящие при рождении и росте отдельного человека, конечно, должно описываться термином «развитие».
Категория изменение описывает процессы, происходящие без появления новых определений. Так процессы, происходящие в современных астрономических объектах корректнее описывать в терминах «изменение» и «развитие», но ни как не «эволюция». Эволюционное формирование наблюдаемых сегодня космологических образований произошло на ранних этапах истории Вселенной, и сейчас мы наблюдаем лишь изменение их параметров. То же можно сказать и о геологических системах – их эволюционное становление происходило на ранних стадиях формирования нашей планеты, а сейчас мы наблюдаем лишь изменения геологических объектов, но ни как не их эволюцию.
Приведенные терминологические уточнения снимают многие проблемы, стоящие на пути распространения теории эволюции на не биологические объекты. И следуют простые выводы.
Первый: различные системы (виды движения) можно рассматривать, как эволюционные лишь на этапе их становления, сопровождающегося появлением принципиально новых качеств и структур, и новых не только для этой системы, а и для Мира в целом. После завершения этого этапа системы продолжают изменяться, но это не носит эволюционный характер. И второй вывод: абсолютно эволюционным объектом является только Мир.
В данном месте рассуждений разумно рассмотреть еще один терминологический вопрос – говорить ли об эволюции Метагалактики или Вселенной? Более уместно было бы использовать термин Вселенная, отодвигающий на второй план проблему множественности Метагалактик. Хотя, наверное, корректнее использовать вообще термин Мир, не носящий космологическую окраску, но по сути совпадающий с категорией Вселенная и более однозначно включающий в себя и биологическую, и социальную формы движения. Что мы и делали выше, но и, не избегая терминов Метагалактика и Вселенная, рассматривая все их как синонимы, опуская имеющиеся различия, как несущественные в контексте данных заметок.
И еще одно сугубо терминологическое замечание – умышленное исключение из лексикона категории «материя» и «дух». Сделано это из-за отсутствия возможности дать их корректное определение в рамках кратких заметок, и дабы не вызывать неизбежных – повторяю чисто терминологических – проблем. Хотя, кому так легче, может вполне термин «вид движения» читать как «форма материи», а под «эволюцией Вселенной» понимать «развитие мирового Духа». Пласт обсуждаемых здесь проблем лежит не на уровне различения этих категорий.
Помимо терминологических есть еще ряд вполне существенных проблем, с которыми связаны сложности развития эволюционного подхода. Здесь выразим их в виде тезисов, которые будут конкретизированы в ходе дальнейших рассуждений. Итак, проблемы:
Структурная – применение термина эволюция к элементам эволюционных систем, хотя изменение свойств этих элементов является вторичным по отношению к действительным эволюционным процессам во всей системе целиком.
Экстраполяционная – попытка перенести принципы эволюционных изменений с одной системы на другие (например, принципы изменчивости и отбора в биологических системах, на эволюцию других видов движения).
Интеграционная – поиск единых закономерностей функционирования эволюционных систем, попытка формального объединения теорий в одну.
Глобальный эволюционизм. Возникновение концепции глобального эволюционизма (ГЭ), во многом связано не только с проблемой обоснования антропного принципа, но и, конечно, с расширением границ эволюционного подхода, принятого в биологической и социальных науках. Сам факт исторического появления и эволюции этих видов движения заставляет усомниться в абсолютной статичности и вечности других видов движения. Загадочность качественных скачков к биологическому и от биологического к социальному миру, наверняка можно постичь только исходя из допущения необходимости подобных переходов между другими видами движения. То есть, исходя из факта наличия эволюции Мира на последних этапах его истории, можно сделать предположение, что Мир в целом является эволюционной системой, то есть и все другие виды движения (помимо биологического и социального) сформировались в результате эволюции . Это высказывание и есть самая общая формулировка парадигмы Глобального эволюционизма.
Заметим, что словосочетание «глобальная эволюция» не обозначает никакого другого понятия отличного от просто «эволюция» (о котором мы говорили в предыдущем параграфе). Эвристичность введения дополнительного термина заключается в распространении эволюционного взгляда на все виды движения, на всю Вселенную, на все этапы ее истории, включая момент ее Начала. При этом отдельные эволюционные процессы (например, биологический или социальный) не являются подчиненными элементами глобальной эволюции, они и есть суть эволюция, ее частные реализации на конкретных этапах истории Вселенной.
Эволюционный подход к существующим видам движения не подразумевает, что все они находятся в постоянном процессе эволюции, а наоборот констатирует необходимость их последовательного формирования на определенных этапах истории Мира. Вообще эволюция, как процесс, относящийся ко всей Вселенной в каждый момент времени реализуется локально только в одном виде движения. То есть всегда существует только одна локальная система (то есть не тождественная всему Миру), которую можно назвать эволюционной, в которой происходит появление принципиально новых, уникальных определений Мира. Чтобы отличить эту систему от других, уже прошедших эволюцию, можно ввести термин «авангард эволюции». Естественно, что авангардом эволюции всегда является последнее по времени появившееся в Мире форма движения (сейчас социальная система). Все предыдущие виды движений, пройдя эволюционный этап и достигнув равновесного состояния (не статичного, а, скорее всего состояния медленного изменения параметров, либо повторяющегося процесса развития отдельных элементов) служат основой для формирования и эволюции нового движения. Возможны появления и новых характеристик у «предыдущих» движений, но они необходимо связаны с эволюцией последнего по времени появления типа движения (системы) – авангарда эволюции.
Итак, в каждый момент истории Вселенной можно выделить только две эволюционные системы – весь Мир и вид движения, являющийся на данный момент авангардом эволюции. Сам эволюционный процесс в них абсолютно совпадает, то есть появление новых характеристик в частной системе есть конкретная реализация эволюции Мира. Глобальный эволюционизм Мира отличается от эволюции отдельных систем лишь своей временной непрерывностью, переносом процесса эволюции с одного вида движения на другой. В то время как эволюционный процесс в отдельной системе необходимо заканчивается при достижении некоего равновесного состояния, а эволюция продолжается в последующем виде движения.
В каждой эволюционной системе (в каждом виде движения в момент, когда он являлся авангардом эволюции) можно выделить эволюционный параметр – параметр, который безусловно, непрерывно изменяется и связан с появлением принципиально новых определений данного типа движения. Так в социальном движении это процесс научно-технического развития – он непрерывен, необратим и определяет изменения других параметров социального движения. Также, говоря об эволюционных процессах в отдельной системе, следует обратить внимание на то, что они относятся ко всей системе целиком и недопустимо механистически переносить их на элементы системы. Так, исследуя социальную эволюцию следует иметь в виду социальную систему в целом, а не развитие и упадок отдельных государств и т.д. (хотя, конечно, все процессы в социальной системе функционально значимы, их совокупность и составляет само социальное движение, но следует четко отделять эволюционную составляющую движения, от частных изменений и развития отдельных элементов).
Один из моментов критики ГЭ связан с абсолютной несхожестью процессов эволюции в биологической и социальной системах и проистечением космологических и геологических процессов. Отмечается их разное направление и результат – одни движутся в сторону усложнения структурной организации, что не скажешь о последних. Наиболее обще на эту критику можно ответить исходя из понимания временности эволюционного развития той или иной системы. Геологическая система была авангардом эволюции на вполне определенном этапе эволюции Мира. Именно в этот период происходили эволюционные процессы образования новых для Мира геологических объектов и структур. А дальнейшие процессы распада геологических объектов не имеют ничего общего с процессом эволюции. Тоже можно сказать и о космологических объектах. Образование астрономических элементов структуры Мира происходило на ранних этапах эволюции Вселенной, а современные процессы видоизменения астрономических объектов не носят эволюционного характера. И только в исторически последнем типе движения (авангарде эволюции) ГЭ реализуется в конкретном эволюционном процессе. (Имея в виду выше приведенные аргументы, можно сделать предположение, что современные научные эксперименты по синтезу живых систем безусловно обречены на неудачу. Возникновение первичных признаков биологических систем было возможно лишь на конкретном этапе эволюции Вселенной, при конкретных физических параметрах, которые естественно не реализуемы в данный момент.)
Перед тем как приступить к тезисному изложению нашего понимания, какой должна быть общая теория глобального эволюционизма, вернемся к проблеме АП, взглянув на нее уже с позиции ГЭ.
Антропный принцип с позиции глобального эволюционизма. Сильный АП с точки зрения ГЭ лишь констатирует, постулирует эволюцию Вселенной. То есть сильный АП, выдвинутый Картером в формулировке: «Вселенная (и, следовательно, фундаментальные параметры, от которых она зависит), должна быть такой, чтобы в ней на некотором этапе эволюции допускалось существование наблюдателей» равносилен утверждению, высказанному с позиции ГЭ: «Вселенная находится в непрерывном процессе эволюции, необходимо приводящем на определенном этапе к возникновению социальной формы движения».
АП высказанный в терминах ГЭ лишен существенного проблемного момента – в нем ничего не говорится о параметрах Вселенной. Во-первых, распространение процесса эволюции Вселенной в прошлое до ее начала снимает вообще вопрос о начальных параметрах – в начальный момент отсутствуют какие-либо определения Мира, а все виды движения и, следовательно, их параметры появляются в процессе эволюции. Во-вторых, ГЭ, констатируя появление социального движения, как необходимого этапа развития Мира, совершенно не регламентирует форму воплощения разума, чем абсолютно снимает проблему соответствия физических параметров Вселенной некой конкретной форме реализации «наблюдателя». То есть при постулировании принципа ГЭ разум во Вселенной возникает необходимо и независимо от конкретных физических параметров «предыдущих» видов движения, при любых их отклонениях (если таковы возможны). И более того с точки зрения обусловленности реализации конкретной формы социальное движение ничем не отличается от других «предыдущих» видов движения (биологического, химического и т.д.).
Следовательно, АП должен быть сформулирован не как принцип соответствия физических и прочих условий возможности появления наблюдателя, а наоборот, как соответствие конкретной формы реализации разума имеющимся условиям. И в этом АП сводится лишь к самой общей форме высказывания принципа глобального эволюционизма.
Для лучшего понимания трактовки АП в рамках ГЭ рассмотрим, к примеру, соответствие окраски некоего вида животного цвету геологических пород в некоторой местности. Можно обсуждать проблему реализации конкретного цвета породы, без которого не возможно было бы существование данного животного – и с какой вероятностью мог бы появиться именно такой цвет, и выдвинуть гипотезу множественности местностей и, что только одна из них совершенно случайно такова, что соответствует окраске животного... Или просто констатировать, что это соответствие есть результат развития вида в условиях данной местности.
Развивая Слабый и Сильный АП, провозглашающие обусловленность существования разума физическими параметрами Вселенной Ф. Типлер дополнительно выдвинул Финалистический АП в следующей формулировке: «Во Вселенной должна возникнуть разумная обработка информации, и, раз возникнув, она никогда не прекратится» (Barrow J.D., Tipler F.J. The anthropic cosmological principle. Oxfotd, 1986). То есть финалистический АП утверждает не только взаимосвязь появления разума с историей Вселенной, но и безусловное влияние его на дальнейшую ее эволюцию.
Как и в случаях слабого и сильного АП, Финалистический АП, рассматриваемый с позиции ГЭ, является лишь констатацией последовательной эволюции отдельных видов движения. Он только обращает внимание не на обусловленность появления конкретной эволюционной системы (в частном случае социальной) предыдущими видами движения, а на необходимое влияние этой системы на последующий процесс эволюции Мира. И с этой позиции финалистический принцип естественно применим ко всем видам движения в равной степени.
Но можно выдвинуть и Сильный Финалистический АП: с появлением разума, с развитием социального движения Мир вступил в новый этап эволюции, характеризуемый активным влиянием одной из форм движения на всю систему (в рамках этих заметок я не буду давать определения этапов эволюции мира). Конец Мира с позиции ГЭ можно понимать как некий акт синкретического единства всех видов движения и он должен осуществиться именно в результате развития разума, как акт абсолютного самопознания Мира. (Это утверждение не несет ни теологического, ни катастрофического оттенка).
И так, подводя итоги, различные АП с позиции ГЭ можно выразить следующим образом:
- Слабый АП: Разум (социальная система) один из видов движения Мира;
- Сильный АП: Разум (социальная система) обязательный этап эволюции Мира, обусловленный всем предыдущими видами движения;
- Финалистический АП: Разумная форма движения Мира неотъемлемый этап развития Мира, определяющий его дальнейшую эволюцию;
- Сильный Финалистический АП: Конец Мира необходимо связан с развитием разума и может трактоваться как момент абсолютного самопознания Миром самого себя.
К общей теории глобального эволюционизма (ОТГЭ). Общая теория глобального эволюционизма – это теория, распространяющая принцип эволюции на весь Мир, на все этапы истории Вселенной. Главное исходное утверждение ОТГЭ – это постулирование эволюционного, последовательного появления и дальнейшего изменения всех существующих видов движения начиная от гравитационного и заканчивая социальным. И водораздел между ОТГЭ и другими теориями описания мира проходит именно в принятии или не принятии самого факта возможности эволюции. Мы акцентирую внимание на этом положении, чтобы четко задать рамки теории и чтобы предостеречь от возражений по частным вопросам при не принятии главного постулата.
Прежде всего, говоря об общих принципах построения ОТГЭ следует исходить из представления, что сама теория, ее сущность, ее структура должны быть адекватны изучаемому, описываемому объекту – Миру, находящемуся в процессе эволюции. Какие выводы можно сделать исходя из этого самого общего взгляда?
Во-первых, ОТГЭ, прежде всего, должна быть сама эволюционной системой. А это значит, что для того, чтобы корректно описывать закономерности эволюционных переходов, она сама должна быть саморазвивающейся системой, продуцирующей категории, понятия и законы непосредственно в процессе своего развития (то есть, предположительно, это должна быть философская система по типу диалектической логики Гегеля.)
Во-вторых, поскольку мы наблюдаем, что все виды движения в процессе эволюции необходимо обуславливая возникновение последующих систем, не сводятся к ним, не растворяются в них, а продолжают функционирование в качестве равновесных систем, то и частные науки описывающие эти движения (физика, химия, биология и др.) не должны быть сводимы друг к другу. То есть, как любой вид движения сохраняет свою специфику, свою самостоятельность в процессе эволюции Мира, так и законы научного описания отдельных видов движения не могут распространяться на предыдущие виды движения (и уж конечно на последующие). И, следовательно, научное описание какого-либо вида движения (в частности, физического взаимодействия) не может рассматриваться как частный случай (частное решение) научной системы, описывающей последующий вид (а именное такой подход, направленный на представление тех или иных научных теорий как частных решений некой обобщенной системы практикуется в современной науке).
Итак, выводы:
ОТГЭ в полном законченном виде должна представлять собой философскую (логическую в гегелевском смысле) систему в которой прослеживается эволюция Мира, в форме последовательного вывода взаимообуславливаемых категорий (определений). Различные части (этапы) этой логической системы должны однозначно соответствовать различным этапам эволюции Мира (отдельным видам движения). Завершаться система должна выводом необходимости Конца Мира, как абсолютного результата эволюции, как возврата к синкретическому единству всех его определений (видов движения), возврата к неопределенному состоянию.
ОТГЭ не включает в себя остальные наука в качестве «частных решений», а лишь определяет их логическую взаимосвязь, указывая, что в рамках каждой из этих научных систем, должен присутствовать механизм развития необходимо приводящий к внутренним противоречиям данной системы, которые разрешаются лишь при переходе к следующему этапу, к следующей системе.
Основное направление эволюции Мира в ОТГЭ понимается не как движение от хаоса к порядку, а наоборот – за исходную точку принимается состояние непосредственного, абсолютно неопределенного (не имеющего определений) порядка, и эволюция начинается с распада этого порядка в результате первого определения Мира самим фактом Начала. Появление первого различенного определения Мира продуцирует новые определения, задающее новый порядок, который распадается в результате соотнесения нового и старых определений. Этот процесс движения Мира, через самоопределение нового порядка и разложение его «под давлением» предыдущих определений и можно назвать эволюцией.
Эту философскую тираду можно изложить и в других терминах. Начало мира можно представить как начало некоего элементарного движения. Изменение мира в ходе этого движения выводит его из текущего равновесного состояния (первым из которых было само неопределенное начало Мира). Непрерывность элементарного движения задает необратимость состояний Мира и, следовательно, новое равновесное состояние может быть достигнуто лишь образованием новых систем, с новыми параметрами и свойствами. Так эволюцию Мира можно представить как поэтапное возникновение новых равновесных состояний на фоне общего движения Мира.
Каждое из этих состояний является динамическим (а не статическим) и изменения параметров предыдущих систем (движений) необходимо приводит к распаду вновь появившегося равновесного состояния и возникновению нового, которое является не только новым этапом развития Мира, но поддерживает стабильность предыдущих систем и продолжает их развитие в качестве своих элементов.
Первыми определениями Мира, непосредственно выводимыми из самого факта Начала являются пространство и время. Пространство, как некое еще неопределенное, непосредственное бытие Мира (Начало «чего») и Время, как некий переход от Небытия к Бытию и т.д. (то есть так далее должна строится, развиваться ОТГЭ).
Гипотеза о последовательном во времени появлении основных видов взаимодействий, а также предположение о принципиальной не сводимости научных теорий, описывающих отдельные виды движения, возможно стимулируют продвижение к созданию единой физической картины Мира. Основной исходной парадигмой для этого должно стать рассмотрение различных физических взаимодействий не как частных реализаций некоего «единого взаимодействия», а как цепочки (последовательности) самодостаточных систем, включающих в себя элемент временного видоизменения, и необходимо порождающих новые системы при возникновении противоречивых решений в предыдущей системе.
Следовательно, при создании единой физической теории следует искать не единую систему уравнений, частными решениями которой описывались бы отдельные типы взаимодействий, а стремиться создать математический аппарат, необходимо содержащий в себе элемент развития. Схема приблизительно может выглядеть так: некая система уравнений описывает определенные процессы, в этой системе имеется параметр при изменении которого система теряет однозначность (появляются бесконечные или прочие противоречивые решения). Для сохранения «равновесности» системы в нее необходимо требуется введение нового параметра, компенсирующего «противоречивые решения». Но в результате получается уже другая система уравнений, не сводящаяся к предыдущей обычными математическими преобразованиями и описывающая уже новый тип процессов. Новая система уравнений должна включать в себя предыдущую как необходимый элемент, но не как частное решение.
Предположение об эволюционном (последовательном) появлении видов движения (физических взаимодействия и др.) снимает одну из существенных проблем космологии. Современная физика без каких либо оговорок предполагает, что законы, описывающие различные виды взаимодействий стационарны и существуют независимо от эволюции Мира. То есть принимается, что хотя в первые моменты после начала Вселенной вследствие особых физических условий (высокая плотность энергии и т.д.) и невозможно было существование элементарных частиц, атомов и т.д., но сами законы (взаимодействия) уже имелись. Возникает несколько вопросов. Во-первых, правомерно ли приписывать некоторые свойства, параметры, законы взаимодействия и т.д. объектам, которые не только не могут еще существовать, но еще и в принципе никогда не существовали? Во-вторых, где хранилась информация об этих законах в момент времени T = 0? Третий вопрос – если считать некоторые законы (и, следовательно, виды движения) Мира вечными и неизменными, то где проходит грань между ними и видами движения явно возникшими лишь на определенном этапе развития Вселенной (биологическое, социальное)? Принимая основные положения ОТГЭ на эти вопросы можно дать однозначные ответы – в момент времени T = 0 физические взаимодействия не существовали, то есть не только не действовали по причине отсутствия объектов, но и категорически не могли быть формулируемы. А дальше... Дальше стоит задача развития как философской системы, логически продуцирующей последовательность категорий, определений Мира, так и создание нового диалектико-эволюционного физико-математического аппарата, способного корректно описать закономерности переходов между типами взаимодействий. (В качестве приложения к этим мыслям можно предложить познакомиться с работой Болдачева А.В «Космология академика В.А. Амбарцумяна и гипотеза о переменности мировых констант»).
Филогенез
В современной биологии значение эволюционной идеи велико, как ни в каком другом разделе естествознания. Причина в том, что материал по разнообразию живых организмов дает больше всего пищи для размышлений о превращениях одних вещей в другие. И вряд ли случайно, что формирование современного глобального эволюционизма началось именно с дарвиновской теории эволюции, объясняющей происхождение биологических видов.
Поскольку биологическое разнообразие есть результат длительного исторического развития, то понять строение и функционирование живых существ невозможно без знания их истории. Это обстоятельство делает исторические реконструкции одной из приоритетных задач в современной биологии.
Познанием путей и закономерности исторического развития организмов занимается особая дисциплина – филогенетика .
Филогенетика зародилась в 60-е годы XIX в. вскоре после выхода в 1859 г. книги Ч.Дарвина «Происхождение видов путем естественного отбора». Термин «филогенез» впервые появился в 1866 г. в фундаментальной работе немецкого биолога-эволюциониста Э.Геккеля «Общая морфология животных». Этим понятием автор обозначил и процесс исторического развития организмов, и структуру родственных (филогенетических) отношений между ними. Введенный приблизительно в эти же годы английским философом Г.Спенсером в научный оборот термин «эволюция» в его современном историческом понимании (ранее им обозначали индивидуальное развитие) также быстро завоевал популярность.
В результате понятия филогенеза и эволюции стали восприниматься как очень близкие по смыслу. Такая классическая трактовка бытует и по сей день: филогенез определяют как пути, закономерности и причины исторического развития организмов. Выдающийся отечественный эволюционист И.И.Шмальгаузен рассматривал филогенез как цепочку сменяющих друг друга онтогенезов.
В настоящее время формируется биотоцентрическая концепция филогенеза, согласно которой биологическая эволюция – это саморазвитие биоты как целостной системы, а филогенез – один из аспектов этого развития. Такое понимание биологической эволюции вообще и филогенеза в частности более всего соответствует современным представлениям об общих законах развития, которые разрабатывает синергетика.
Один из важнейших результатов эволюции – глобальная структура биоты Земли, которая проявляется в многоуровневой иерархии групп, по-разному интегрированных и организованных. В некотором приближении можно считать, что эта структура состоит из двух иерархий. Одна из них связана с разнообразием биоценозов (природных экосистем), объединенных экологическими отношениями. Историческое становление этой иерархии обозначают как филоценогенез . Вторая связана с разнообразием таксонов – филогенетических групп, члены которых находятся в родственных отношениях. Становление именно этой иерархии и есть филогенез .
Филогенез, или Филогеиия (др.-греч. phylon – племя, раса и др.-греч. genetikos – имеющий отношение к рождению) – историческое развитие организмов. В биологии филогенез рассматривает развитие биологического вида во времени. Биологическая классификация основана на филогенезе, но методологически может отличаться от филогенетического представления организмов.
Филогенез рассматривает эволюцию в качестве процесса, в котором генетическая линия – организмы от предка к потомкам – разветвляется во времени, и ее отдельные ветви могут приобретать те или иные изменения или исчезать в результате вымирания.
Имеющиеся на сегодняшний день знания о ветвлении филогенетического древа получены путем построения классификации живых организмов, которая исходно была задумана Карлом Линнеем как отражение «Естественной Системы» всей природы (в том числе и неживой). Впоследствии было установлено, что такой «Естественной Системы» не существует, а то, что К. Линней принимал за проявление этой системы у животных и растений, является филогенией, то есть результатом биологической эволюции. Для более эффективного анализа филогении в настоящее время разрабатываются принципы, в которых метод записи классификации усовершенствован по сравнению с линнеевским, что позволяет более адекватно записать филогению в форме классификации и продолжить ее анализ.
Рис. Филогенез основных групп организмов
В филогенезе выделяется несколько компонент, позволяющих исследовать историческое развитие с разных сторон. В начале XX в. австрийский палеонтолог О.Абель разграничил их следующим образом: ряды предков – «истинные филогенезы»; ряды приспособлений, касающихся одного органа; ряды ступеней совершенствования организации. В настоящее время «истинный филогенез» – развитие (появление и/или изменение состава) филогенетических групп организмов как таковых, безотносительно их свойств, – обозначают как кладогенез, или кладистическая история. Этот термин предложил английский биолог Дж. Гексли в 40-х годах, противопоставив его анагенезу – изменению уровня организации живых существ в процессе эволюции. Исторические изменения отдельных органов и свойств организмов немецкий систематик и эволюционист В.Циммерман в 60-х годах предложил называть семофилогенезом. Более удачен термин семогенез, обозначающий историческое развитие (появление, изменение, исчезновение) отдельных морфологических и иных структур, рассматриваемые вне связи с конкретными группами организмов, которым они присущи.
Группы, порождаемые кладогенезом, называют кладами: например, хордовые, а в их пределах – позвоночные. Группы, порождаемые анагенезом, – грады, ступени эволюционного развития: таковы многоклеточные животные по отношению к одноклеточным, а среди позвоночных – гомойотермные животные (птицы и млекопитающие) по отношению к пойкилотермным (низшие позвоночные). Принципиальное отличие двух этих категорий состоит в том, каким путем приобретаются общие свойства. Клады наследуют их от общего предка, а у град они появляются а результате параллельной или конвергентной эволюции. Современная филогенетика по преимуществу изучает становление иерархии филогенетических групп и специфичных для них свойств, ее основная задача – реконструкция кладогенеза. Из этого видно, что на нынешнем этапе своего развития филогенетика является по преимуществу кладогенетикой.
От естественной системы к эволюционной идее. В классической биологии в XVII-XIX вв. одним из ключевых было представление о естественной системе как всеобщем законе, которому подчинено все сущее. Эта система объединяет естественные группы организмов, существующие в самой природе. Ее воссозданием занималась и занимается систематика, задача которой состоит в том, чтобы распознать эти группы по их «естеству».
Коль скоро конечная цель научного познания – выяснение причин того или иного явления, сама естественная система нуждалась в объяснении: в чем ее изначальная причина. В Средние века таковой считался божественный план творения. Со становлением науки Нового времени причину стали искать в самой Природе: так в естественнонаучное мировоззрение вошла идея исторического развития, давшая наиболее правдоподобное с материалистической точки зрения толкование естественной системы.
Важнейшую роль в формировании систематики и филогенетики сыграла немецкая натурфилософия. С ней связано, с одной стороны, восходящее к Аристотелю представление о «Великой лестнице Природы» (Г.Лейбниц), с другой – уподобление Природы сверхорганизму (Ф.Шеллинг, Л.Окен). Они дали два способа представления естественной системы – линейную и иерархическую. В первом случае имелся в виду непрерывный ряд организмов («природа не делает скачков»), направленный от низших форм жизни к высшим. Во втором случае подразумевалось, что биота, подобно организму, поделена на части разного уровня общности – естественные группировки.
Уподобление биоты сверхорганизму имело особое значение для формирования эволюционного мировоззрения: ведь живое существо немыслимо без развития, направленного в сторону все большего совершенства и дифференциации. На этой основе, вкупе с идеей «лестницы совершенствования» (другое название «лестницы природы»), сформировалась ключевая идея классического эволюционизма, а с ним и классической филогенетики – уподобление исторического развития биоты индивидуальному развитию организма.
Эволюционная интерпретация непрерывного ряда форм породила учение Ж.-Б.Ламарка, согласно которому основная задача эволюционной теории сводилась к объяснению того, каким образом возникают новые свойства организмов, обеспечивающие повышение их уровня организации. Так в филогенетике возникла адаптационистская парадигма.
Иерархической системе более всего соответствует дивергентная концепция эволюции, связываемая с именем Ч.Дарвина. В данном случае история фактически сведена к многократно ветвящимся «цепочкам» превращения одних видов в другие. Такое видение эволюционного развития сформировало в филогенетике генеалогическую парадигму.
Биологи с натурфилософским складом ума воспринимали историческое развитие как макромасштабный процесс. Для них сверхзадачей была (и по сей день остается) реконструкция становления основных групп живых организмов – царств, типов, ступеней развития. Для сторонников набиравшей силу дарвиновской трактовки эволюции ключевыми были события, происходящие на видовом уровне. Соответственно, в реконструкции истории основной акцент делается на «локальные» события – превращения одних видов в другие.
В начале XX в. вся натурфилософская картина мира была признана «ненаучной», поскольку ее утверждения не проверялись опытным путем. В новое мировоззрение как нельзя лучше вписался дарвиновский селекционизм, дополненный мутационной теорией. Постепенно сложилась синтетическая теория эволюции, идеологическую основу которой составило популяционное мышление. В ней терминологически закрепилось разделение исторического развития на микроэволюцию и макроэволюцию (автор терминов – Ю.А.Филипченко); при этом была абсолютизирована первая и фактически отвергнута вторая.
Сведение механизмов эволюции к популяционным процессам позволило рассматривать филогенез как их «механическое» следствие, возникающее просто из-за того, что жизнь существует и развивается очень продолжительное время. Такое развитие эволюционной идеи в первой половине – середине XX в. сделало филогенетику в известной степени «вещью второго сорта», лишило ее прежнего приоритетного статуса. Впрочем, во второй половине того же столетия популяционные идеи сыграли значительную роль в формировании современного этапа развития этой дисциплины.
Классическая филогенетика. Понимание филогенеза как закономерного исторического процесса не мешает по-разному трактовать смысл филогенетического единства, выделяющего естественные группы. Это породило несколько линий развития классической филогенетики, которые соответствуют трем уже упоминавшимся ключевым компонентам филогенеза – кладогенезу, семогенезу и отчасти анагенезу.
Признание филогенетического единства преимущественно как общности происхождения, т.е. монофилии, стимулировало развитие филогенетики в кладогенетическом направлении. Именно для кладогенетики своего рода символом стала метафора ветвящегося филогенетического дерева с единственным корнем.
В геккелевой трактовке эта концепция филогенеза соединяет натурфилософское и дарвиновское понимания эволюции. Историческое развитие рассматривается на «макроуровне», но при этом родоначальником всякой монофилетической группы считается вид. В качестве «наследия» натурфилософии в геккелевой филогенетике присутствует и идея эволюционного прогресса: она отчетливо видна из формы филогенетического дерева, в котором доминирует устремленный ввысь центральный ствол, от которого отходят боковые ветви.
При изучении семогенетической составляющей филогенеза в основную задачу входит выяснение закономерностей и причин преобразования отдельных структур. В данном случае филогенетическое единство организмов определяется главным образом через общность эволюционных тенденций, которая выражена в единстве направлений и этапности исторического развития. На такое понимание филогенеза во многом повлияли представления об эволюции как однонаправленном процессе, развиваемые некоторыми палеонтологами (Э.Коп, В.О.Ковалевский, Г.Осборн). Этому более соответствует метафора не дерева, а куста или даже травяного поля, на котором каждый «стебель» – филетическая линия – произрастает из своего собственного корня (Л.С.Берг).
Методология классической филогенетики базируется на методе тройного параллелизма, т.е. на максимальном согласовании данных сравнительной анатомии, эмбриологии и палеонтологии. Ключевое значение имеет концепция гомологии, исходно разработанная английским анатомом-платоником Р.Оуэном: подразумевается, что для реконструкции филогенеза важны лишь гомологичные структуры, тогда как аналогичные препятствуют ей. Большое место отводится определению вероятных путей эволюции этих структур на основе критериев, вырабатываемых в рамках перфекционистской (от простого к сложному, от менее к более структурированному) и адаптационистской (от менее к более приспособленным организмам) моделей. Частью классической методологии служит анализ предковых форм: с их помощью разрозненные группы «связываются» в монофилетические образования. Особое внимание уделяется ископаемым материалам, среди которых ищутся реальные предки; при их отсутствии предковые признаки реконструируются.
Важная роль отводится взвешиванию признаков. В классической филогенетике оно всегда трактуется как дифференциальное: разным признакам придается разное значение (разный «вес»). Однако в разных школах филогенетики эта общая концепция трактуется специфическим образом в зависимости от понимания сути филогенеза.
Генеалогическая трактовка предполагает деление признаков на «филогенетические», не связанные с частными адаптациями, и на «адаптивные». Подразумевается, что именно первые следует использовать для установления родственных связей между видами, поскольку они унаследованы от предков (ближайших или отдаленных), что свидетельствует о филогенетической общности их потомков.
Адаптационистская трактовка, напротив, придает наибольший вес тем признакам, которые отражают адаптивную специфику данной группы. Фактически эта идея – отголосок прежнего понимания «главной сущности» естественных таксонов. В данном случае существенность морфологических структур связывается с их адаптивной (чаще всего функциональной) значимостью.
В настоящее время традиции классической филогенетики развивает подход, названный филистикой. В ней филогенетическое развитие по-прежнему рассматривается как упорядоченный адаптациогенез, в котором закономерные тенденции преобладают над случайными. Поэтому при характеристике филогенетического единства групп большое значение придается не только единству происхождения, но и единству эволюционных тенденций их представителей как лишнему доказательство монофилии.
Новая филогенетика. Большинство современных реконструкций выполняется в рамках новой филогенетики. Сформировалась она в результате объединения трех подходов, поначалу (в 60-е годы) развивавшихся независимо, – кладистики, нумерической филетики и генофилетики. Ее можно определить как раздел филогенетики, разрабатывающий филогенетические гипотезы на основе кладистической методологии средствами нумерической филетики при использовании молекулярно-генетических данных.
Идеологическое ядро новой филогенетики составляет кладистика, становление которой связывается с именем немецкого энтомолога В.Геннига. Его основополагающая книга была опубликована на немецком в 1950 г., но особую популярность это направление получило после выхода англоязычного издания (Hennig W. Phylogenetic systematics. Urbana, 1966.). Тогда же появился и сам термин «кладистика».
Методологической основой кладистического подхода служит принцип экономии, который по возможности исключает исходные домыслы об исследуемом явлении. Соответственно, исторические реконструкции проводятся при минимальных априорных допущениях о свойствах эволюционного процесса. Общая концепция эволюции как адаптациогенеза, обязывающая детально прописывать эволюционные сценарии, взвешивать признаки согласно их адаптивной значимости и т.п., оказывается избыточной. Этим обосновывается ключевая для новой филогенетики редукция исторического развития к кладогенезу, в котором нет ни параллельных линий, ни ступеней эволюционного развития.

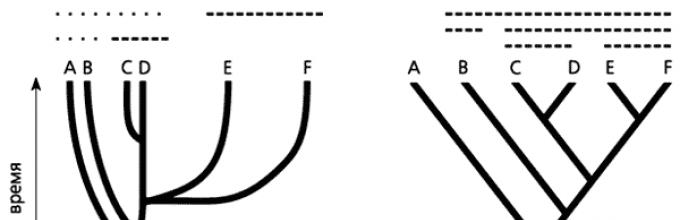
Рис. Стилизованные представления филогенетических схем в современной филогенетике: а – филограмма, б – кладограмма. Сплошными линиями показаны парафилетическе группы, штриховыми – голофилетические группы.
Такое упрощенное понимание филогенеза существенно изменило его графическое представление. Своего рода символом кладистики, а с ней и всей новой филогенетики стала кладограмма – стилизованное филогенетическое дерево (рис.). На нем, в отличие от филограммы, показывающей степень дивергенции и уровни продвинутости групп, присутствуют только точки ветвления, отражающие последовательность становления монофилетических групп. Анагенетическая составляющая эволюционного развития отсутствует; поэтому, глядя на кладограмму, нельзя судить об уровнях дивергенции и продвинутости групп организмов.
Одна из причин успеха кладистики – эффективная методология филогенетических реконструкций, которая сделала принципиально возможной их воспроизводимость. Ее важная часть – более «узкая» трактовка понятий монофилии и монофилетической группы (см. рис.4). В классической филогенетике монофилетической считается группа, включающая любых потомков данной предковой формы. Очевидно, при выделении на таком филогенетическом дереве монофилетических групп возможны весьма широкие разночтения. В кладистике признаются только голофилетические группы, включающие всех потомков данной предковой формы. Группы, в которых лишь часть таких потомков (парафилетические), не рассматриваются.

Рис.Категории сходства, связанные с концепцией синапоморфии. Заглавными буквами обозначены апоморфии, строчными – плезиоморфии; цветом выделены апоморфии, определяющие голофилетические группы.
Частью этой методологии стал парадоксальный с точки зрения классической филогенетики отказ от учета отношений предок-потомок при обсуждении родства и монофилии. Основанием для этого послужило то соображение, что конкретный предковый вид для подавляющего большинства групп, особенно высокого ранга, нельзя установить. В таком случае более корректна отсылка к сестринской группе, возникшей в результате одного кладистического события вместе с исследуемой. Это дает возможность игнорировать геохронологическую составляющую эволюции и «уравнивать» палеонтологические данные с современными. Такая позиция подводит теоретическую базу под генофилетические реконструкции, в которых предковые формы изначально не нужно исследовать.
Основу большинства алгоритмов новой филогенетики составляет принцип синапоморфии, согласно которому монофилию группы можно выявить только через сходство по производному состоянию признака (апоморфии, рис). В отличие от этого симплезиоморфия – сходство по исходному состоянию признака (плезиоморфии) – не позволяет определить монофилию группы. При этом особо выделяется синапоморфное сходство, унаследованное от ближайшего предка данной группы (истинная синапоморфия). Напротив, сходство в результате параллельной эволюции (ложная синапоморфия) считается незначимым: это принципиально отличает кладистику от классической филогенетики.
Последнее составляет один из важнейших критериев – значимость признаков: их «вес» обратно пропорционален вероятности параллелизмов и реверсий в эволюции. Чем выше эта вероятность, тем менее значимы признаки, поскольку они с меньшей надежностью позволяют судить о монофилии. Вместо выбора наиболее существенных признаков вводится количественный критерий суммарной оценки значимости сходства: чем большим числом общих апоморфий обладают два таксона, тем выше вероятность их принадлежности к монофилетической группе. Следовательно, чтобы узнать, монофилетичны ли те или иные группировки таксонов, достаточно просуммировать специфические для каждой из них синапоморфии и сравнить результаты. Это позволяет достаточно просто решать конфликты, возникающие между разными признаками, по которым могут получаться разные филогенетические схемы.
Такой критерий стимулировал активное освоение современной филогенетикой количественных методов, которые разрабатывает нумерическая филетика. Сегодня существует своего рода «индустрия» по производству методов количественной оценки сходства организмов и конструирования филогенетических деревьев и их реализации в виде компьютерных программ.
В настоящее время наиболее популярны два метода – экономии (парсимонии) и наибольшего правдоподобия. Принципиальные различия между ними состоят в разной трактовке начальных условий реконструкций. Методы первой группы, выделяемые со ссылкой на рассмотренный выше принцип экономии (отсюда их название), предполагают полный отказ от априорного определения эволюции признаков, которые трактуются фенетически. Во второй группе признаки вводятся в качестве элементарных гипотез о семогенезах, в которых определены вероятные направления эволюционных преобразований соответствующих структур. В первом случае итоговая гипотеза оптимальна, если в ней минимально количество параллелизмов; во втором случае – если она наиболее правдоподобно суммирует все эволюционные сценарии для признаков.
Несомненное достоинство нумерической филетики в том, что она позволяет манипулировать большими массивами данных. Морфолог чаще всего способен охватить интуицией лишь ограниченное число структур, которые можно описать несколькими десятками формализованных признаков. Однако без применения количественных техник едва ли можно сравнить десятки таксонов по нуклеотидным последовательностям, состоящим из нескольких тысяч пар оснований. Но здесь кроются и свои «подводные камни». Так, суммирование нескольких десятков морфологических и нескольких сотен молекулярных признаков приводит к тому, что первые «поглощаются» вторыми. В результате филогенетическая схема почти целиком определяется молекулярно-генетическими данными.
Активная «нумеризация» новой филогенетики все более переносит акцент с теории и методологии реконструкций на их «технологию». Обсуждаются детали вычисления доверительных интервалов при применении вероятностных критериев, скорость работы компьютерных программ, их ограничения на манипулирование большими массивами данных и т.п. При этом все меньше внимания уделяется биологическому осмыслению результатов, что во многом выхолащивает эволюционный смысл подобных реконструкций.
Особый раздел нумерической филетики составляет компьютерное моделирование филогенетических процессов. Оно позволяет исследовать некоторые свойства исторического развития организмов на основе симуляционных моделей.
Генофилетика (филогеномика) – раздел новой филогенетики, в котором заключения о кладистической истории выводятся на основании анализа исключительно молекулярно-генетических данных. Это новейший этап развития молекулярной филогенетики, связанный с изучением нуклеотидных последовательностей. Теоретические предпосылки для использования таких данных в филогенетике были сформулированы в рамках синтетической теории эволюции (СТЭ), которая свела эволюцию к изменениям, протекающим на генетическом уровне. На этом основании была выдвинута идея: чем «ближе» признаки к генотипу, тем больше их значение для филогенетических реконструкций. Нечего и говорить, что получение технического доступа к первичной структуре информационных макромолекул было расценено как «молекулярная революция» в филогенетике.
В генофилетике при реконструкции родственных отношений особое значение (со ссылкой на Дарвина) придается изменениям в первичной структуре макромолекул, не связанным с выработкой частных адаптаций. Такая трактовка эволюции вполне согласуется с принципами кладистики, что естественно объединяет последнюю и генофилетику в рамках новой филогенетики. Данное обстоятельство следует подчеркнуть особо: если бы в современной филогенетике доминировала адаптационистская концепция, в ней не нашлось бы места генофилетическим реконструкциям, коль скоро они не интерпретируемы с позиций адаптациогенеза.
В генофилетике организм сводится к первичной структуре биополимеров, каждый элемент которых (нуклеотид) рассматривается как отдельный признак, а их может набираться несколько тысяч. Суждения о родстве базируются на суммарной оценке сходства по этим элементарным признакам, причем чем их больше (чем длиннее последовательность), тем более надежными считаются эти суждения. Поэтому единственный приемлемый метод оценки сходства – количественный.
Для перехода от сходства по молекулярным структурам к родству была разработана концепция молекулярных часов. В ее основу легла так называемая модель «нейтральной эволюции», предполагающая стохастическую природу накопления нейтральных мутаций в филетических линиях. Ключевым для концепции стало допущение, что ход этих «часов» приблизительно равномерен и одинаков для разных молекулярных структур и в разных группах организмов. Следовательно, если известна скорость накопления мутаций, то можно определять время разделения групп и на этом основании устанавливать между ними генетическое родство.
Количественная оценка различий по молекулярным структурам (генетическая дистанция) дает собственное время дивергенции, напрямую несопоставимое для разных групп организмов. Перевод его в единое для них абсолютное время (в миллионах лет) осуществляется посредством калибровки «молекулярных часов». Для этого вычисленные дистанции сопоставляются с временем появления в палеонтологической летописи первых представителей монофилетических групп, выявленных молекулярными средствами. Затем проводится перерасчет дистанций в единицах скорости изменения первичной структуры макромолекул и определяется абсолютное время дивергенции.
Отсюда видно, что задача калибровки «молекулярных часов» не решается в рамках самой молекулярной биологии: результат зависит от палеонтологических данных. Здесь очень важны точность определения времени захоронения ископаемых остатков и правильность их отнесения к группам, выделенным по молекулярным маркерам. Кроме того, разные методы оценки генетических дистанций, а также самой калибровки «часов» могут давать разные результаты. Все это служит серьезным основанием для критики исходной упрощенной трактовки концепции «молекулярных часов». Накопленные данные показывают, что скорость изменения первичной структуры различна для разных макромолекул, а для одних и тех же макромолекул – для разных их участков и в разных группах организмов. Значит, универсальных «молекулярных часов» не существует в силу неравномерности их хода.
Возможности генофилетики существенно ограничены тем, что молекулярно-генетические данные, пригодные для филогенетических исследований, накоплены пока лишь для несколько десятков тысяч из почти 2 млн. современных видов, известных науке. А ведь еще есть и многие миллионы вымерших видов, по которым такого рода данные никогда не будут доступны. Все это делает особо актуальной проблему неполноты выборки: чем меньше в исследуемой группе представлено относящихся к ней таксонов, тем менее точна оценка филогенетических отношений между ними.
Приложения новой филогенетики. Многие биологические дисциплины (систематика, биогеография, в последние годы также экология и этология) в той или иной форме используют результаты филогенетических исследований. И в этом смысле их можно считать сферами приложения филогенетики как научной дисциплины.
В настоящее время существует целый класс задач, решение которых так или иначе связано с анализом филогенетического (точнее, кладогенетического) сигнала – информации о происхождении организмов, содержащейся в структуре их сходственных отношений. Оценка этого сигнала равносильна выявлению исторических причин, наряду с другими обусловивших наблюдаемое разнообразие. Очевидно, чтобы выявить названный сигнал, необходимо достаточно полно реконструировать кладистическую историю организмов.
Действительно, всякая развивающаяся система, образно говоря, есть «жертва своей истории». Это значит, что строение или образ жизни организмов могут быть обусловлены особенностями не только их среды обитания, но и особенностями их предыстории. Например, почему у разных наземных животных разное количество конечностей – четыре у позвоночных, шесть у насекомых, восемь у паукообразных? Ответы вряд ли имеет смысл искать в их условиях обитания. Скорее, они кроются в морфологической организации предковых форм каждой из этих групп, от которых потомки унаследовали свои особенности. Понимание этого привело к формированию исследовательской программы - сравнительной филогенетики. Она изучает проявления исторических причин в особенностях образа жизни, морфологии, поведения и т.п. в разных группах организмов.
Чуть ли не с первых шагов своего развития филогенетика тесно связана с исторической биогеографией, предоставляя ей сведения об истории формирования таксонов, составляющих ту или иную локальную биоту. В настоящее время эта связь оформилась в виде кладистической биогеографии, которая целиком опирается на анализ кладистической составляющей филогенеза.
Если изначально филогенетика фактически «выросла» из биологической систематики, то сегодня, напротив, новая филогенетика оказывает существенное влияние на систематику, ее принципы и методы. Классическая филогенетика определяет задачи систематики как отражение результатов филогенеза в системе таксонов, характеризуемых филогенетическим единством. Уточнение этого единства дало начало так называемой кладистической систематике, цель которой сводится к «переводу» кладограммы в таксономическую систему.
Парадокс ситуации в том, что это различие, очень незначительное по форме, весьма сильно повлияло на состояние современной систематики. Очень многие таксоны, признанные поколениями систематиков, оказались парафилетическими и потому подлежащими “упразднению”. Это вызвало волну существенных перестроек естественной системы живых организмов и, соответственно, резкую полемику по поводу их правомочности и обоснованности. Типичным примером таких разногласий может служить ситуация, сложившаяся с системой современных представителей рептилий и птиц (рис). В классической систематике в качестве естественных групп принимаются линнеевские классы Reptilia (черепахи, гаттерии, лепидозавры и крокодилы) и Aves (птицы). Причина достаточно очевидна: эволюция птиц вывела их за пределы рептилийной адаптивной нормы, так что они утратили единство с ними.

Рис. Классическая (слева) и кладистическая (справа) классификации рептилий и птиц.
Но в генеалогической схеме среди перечисленных групп ближайшие родственники крокодилов – птицы. С точки зрения кладистики, классический таксон Reptilia – парафилетический: в его составе должны остаться только гаттерии и лепидозавры (ящерицы и змеи). Черепах нередко объединяют в класс Parareptilia, а птиц и крокодилов следует выделять в класс Archosauria (куда из ископаемых форм относятся также динозавры и птерозавры).
Генофилетический раздел новой филогенетики разрабатывает так называемую геносистематику, в которой таксоны формируются исключительно на основе генетических данных. Ее заслуга в уникальном фактологическом обеспечении старой идеи «Дерева жизни» – глобальной классификации мира живых организмов. В частности, очень детально прорабатывается таксономическая система для прокариот, приведшая к выделению новых царств живых организмов.
Кладистическая систематика не оставила без внимания и таксономическую номенклатуру. В настощее время разрабатывается так называемый «Филокодекс», идеологическую основу которого составляет принцип монофилии в его “узкой” трактовке (Cantino P.D., Queiroz K. The PhyloCode: a phylogenetic code of biological nomenclature. 2003).
Развитие науки в некотором смысле подобно биологической эволюции. Разнообразие идей, подходов, методов столь же присуще динамично развивающейся научной дисциплине, как разнообразие видов – эволюционирующей биоте. Постоянная конкуренция идей, возникновение и отмирание научных школ, развитие новых направлений – все это и следствие, и причина научного прогресса.
Филогенетика в этом отношении не составляет исключения. Ее основной вектор развития во все времена задан очевидной «сверхцелью» – все более полной реконструкцией истории формирования биоты. Как достичь ее – вот основная проблема этой дисциплины. Способы ее решения порождают разные подходы к филогенетическим реконструкциям. Формирование новой филогенетики связано с уточнением этой общей задачи и попытками по-новому решить старые проблемы. Но каждый подход действует в рамках своих возможностей, а его неоправданно оптимистичная трактовка часто уводит в неверном направлении. Поэтому было бы ошибочным считать, что предлагаемые сегодня исторические реконструкции – окончательные.
Очевидно, что на смену новой филогенетике придет какой-то другой подход. Можно полагать, что, подчиняясь тенденциям развития эволюционной теории, он будет свободен от избытка редукционизма, присущего современным исследованиям.
Теперь обратимся к рассмотрению причин и движущих факторов биологической эволюции.
(1) Что такое филогенез
Процесс необратимого исторического развития мира живых организмов, как в целом, так и отдельных таксономических групп, называется филогенезом (от греч. phylon – род, племя и genesis – происхождение, возникновение).
Развитие, как индивидуальное, так и историческое, – важнейшее свойство жизни. Но развитие, самоорганизация присущи материи вообще. В широком смысле процесс развития – это движение, изменение в пространстве и времени. Движение как способ существования материи не возникает и не исчезает, существует вечно – без начала и конца. В то же время движение – векторный, направленный, необратимый процесс, а значит, на отдельных отрезках имеет начало и конец: ... à ... à ... à ...
Это означает, что историческое развитие как движение бесконечно и вместе с тем существует всякий раз как отдельный конечный процесс . Эти кажущиеся противоречия разрешаются при философском взгляде на процесс развития. Историческое развитие любой системы: физической, биологической, социальной – подчиняется общим законам диалектики и синергетики. Развитие совершаетсяпо спирали (это от диалектики) и сопровождается периодическимибифуркациями (согласно законам синергетики) (рис. 6.5).
В самом деле, один виток спирали – некоторый конечный процесс. В схеме развития жизни на Земле такой виток представляет отдельный онтогенез – индивидуальное развитие отдельной особи . Бифуркации, или дивергенции, – расхождение путей развития, образование множества новых онтогенезов, т.е. новых видов. Тогда спираль с ее многочисленными бифуркационными разветвлениями как бесконечный исторический процесс представляет филогенез – необратимую эволюцию организмов, ведущую к увеличению и усложнению биоразнообразия .
Рис. 6.5. Спирально- бифуркационный принцип развития
Каждый виток спирали – отдельный онтогенез; вся разветвленная спираль – филогенез
Таким образом, филогенетические преобразования происходят посредством перестройки онтогенезов особей ; при этом приспособительную ценность могут иметь изменения любой стадии индивидуального развития. Можно сказать, что филогенез представляет собой преемственный ряд (а с учетом дивергенций – древо) онтогенезов последовательных поколений .
(2) От Ламарка и Дарвина к синтетической теории эволюции
Какие же факторы и каким образом обеспечивают филогенетическое развитие? Почему возникают изменения в индивидуальных онтогенезах? Что происходит в точке бифуркации (дивергенции), за которой один биологический вид превращается в два или более видов с новыми признаками? На эти вопросы дает ответы современная синтетическая теория эволюции . Ее формирование имеет свою сложную историю и вряд ли может считаться завершенным.
Первым настоящим эволюционистом был французский биолог Жан Батист Ламарк (1744–1829). В своей известной книге “Философия зоологии” (1809) он предложил первую целостную теорию эволюции живого мира, отвергнув метафизическую идею о постоянстве и неизменности видов. Учение Ламарка основано на большом сравнительно-анатомическом материале и на новом видении системы – от простых организмов к сложным. Оно утверждало прогрессивное развитие жизни на Земле. В то же время, подавляющее большинство теоретиков биологии считают, что Ламарк ошибался в оценке движущих сил (факторов) эволюции. Совершенно справедливо признавая ведущую роль в изменении видов за внешними условиями (климат, почва, пища, свет, теплота и т.д.), он полагал, что самим организмам присуще “внутреннее стремление к совершенствованию”. Ламарк считал, что упражнение органов у отдельной особи ведет к их определенному, целенаправленному изменению, совершенствованию, и эти полезные изменения автоматически закрепляются в потомстве, передаются по наследству (наследование благоприобретенных признаков) . Общеизвестен утрированный пример по этому поводу: жирафы в результате постоянных “упражнений” на срывание листьев с деревьев в ряду многих поколений обрели длинную шею.
Причины и движущие силы филогенетических процессов вскоре после Ламарка были переосмыслены Дарвином. Чарльз Дарвин (1809–1882) – великий английский натуралист, создатель учения о происхождении видов путем естественного отбора. В 1831–1836 годах молодой Дарвин в качестве натуралиста совершил кругосветное путешествие на корабле “Бигль”, в ходе которого собрал большой биологический, географический, геологический и палеонтологический материал, легший в основу его учения. Кроме того, Дарвин досконально изучил историю разведения и селекции домашних животных и растений и пришел к выводу, что подобно искусственному отбору , применяемому в сельскохозяйственной практике, в дикой природе“работает” естественный отбор , который и играет основную творческую роль в формировании наиболее приспособленных видов.
Синтетическая теория эволюции сложилась к середине XX века на основе классической теории эволюции Дарвина, генетики и популяционной экологии . В наше время свою лепту пытается внести иммунология . По сути, синтетическая теория развивает на новом уровне знания идею естественного отбора, обоснованную Дарвином за 100 лет до этого синтеза. Какие же базовые положения были привнесены в синтетическую теорию из предшествующих источников?
(3) Дарвинизм – основа синтетической теории эволюции
Дарвинизм остался фундаментом новой теории. В классическом произведении Дарвина “Происхождение видов путем естественного отбора...” обозначены главные факторы, или движущие силы, эволюционного процесса. Теория Дарвина включает несколько положений.
1. Особи производят больше потомков, чем это необходимо для поддержания их численности, следовательно, организмы способны к неограниченному размножению; при этом им свойственна неопределенная наследуемая изменчивость (ненаправленная, случайная – в отличие от определенной, направленной изменчивости у Ламарка).
2. Из-за ограниченности природных ресурсов (пищи, пространства и др.) между особями происходит борьба за существование , в ходе которой большая часть гибнет и не дает потомства. Заметим, что идея борьбы за существование “витала в воздухе”, Дарвин подхватил и развил ее вслед за Т. Мальтусом, который “борьбу за существование” использовал в своей социологической теории регулирования народонаселения.
3. Выживают и оставляют потомство организмы, наиболее приспособленные к данным условиям среды, в чем и проявляется естественный отбор.
4. В результате отбора особи одного вида накапливают разные приспособительные признаки (творческая роль отбора), что ведет к их дивергенции (лат. divergere – отклоняться, расходиться) и образованию новых видов .
Таким образом, по Дарвину, наследственная изменчивость представляет предпосылку эволюции, борьба за существование и естественный отбор – ее процесс, а видообразование – результат.
(4) Вклад генетики в теорию эволюции
Генетика уже в новом XX веке дала понимание природы наследственности (хромосомы, ДНК) и механизмов изменчивости (мутации и рекомбинации генов) . Открытые Менделем еще в 1865 г. и переоткрытые в 1900 г. законы о независимом наследовании отдельных признаков и их сохранении в скрытом состоянии позволили объяснить то, что Дарвин не понимал и не мог понимать, так как природа наследственности в его время была неизвестна. Генетика и цитология показали, что каждый признак у организма кодируется двумя аллельными генами, расположенными в гомологичных хромосомах – от двух родителей (хромосомный набор диплоидный). Это позволяет организму без риска немедленной гибели “пробовать” в одном из аллелей различные варианты мутационной изменчивости.
Сегодня мы знаем, что мутационная изменчивость особей возникает в результате спонтанных или индуцированных мутаций в половых клетках родителей и несет потенциальные новые признаки. Спонтанные (самопроизвольные) мутации возникают как ошибки подбора нуклеотидов во время репликации ДНК. Индуцированные, или наведенные, мутации вызываются воздействием разнообразных мутагенных факторов. Естественными мутагенами являются: ионизирующая радиация космоса и земных недр, ультрафиолетовый солнечный свет, химические, в том числе пищевые, вещества (особенно свободные активные радикалы кислорода), вирусы и др.
В развитие этих генетических построений российский биолог С.С. Четвериков в 20-х годах XX века разработал важное положение о генофонде вида и популяций , под которым понимается совокупность всех диких и мутантных вариантов генов (у всех особей), присутствующих у данного вида или популяции и участвующих в свободном скрещивании. При половом размножении прежде скрытые признаки выходят в новые комбинации и проявляют себя как новый цвет, новая форма, новая ферментная активность и т.п., а далее проверяются путем естественного отбора на свою полезность или вредность. Таким образом, чем более популяция насыщена скрытыми мутациями, тем чаще возникают новые мутантные комбинации как новые предложения для отбора. Генофонд составляет резерв наследственной изменчивости популяции и ее эволюционный потенциал .
Современная иммуногенетика в поисках механизмов развития и наследования так называемого приобретенного иммунитета приходит к неожиданному альтернативному выводу в духе неоламаркизма . Ряд ученых считает, что мутации на выработку полезных иммуноглобулинов, возникающие в лимфоцитах(а это соматические клетки), могут отбираться и передаваться в половые клетки и таким образом закрепляться в потомстве. Получается, что родительский организм создает в соматических клетках (лимфоцитах) направленную , заведомо полезную иммунную изменчивость и закрепляет ее через половые клетки в потомстве (Стил и др., 2002). Эти новые данные, в случае их широкого признания, могут изменить или существенно дополнить уже сложившиеся представления о механизмах наследственной изменчивости.
(5) Экологические закономерности в теории эволюции
Наконец, популяционная экология дала для синтетической теории эволюции понимание того, что элементарные процессы изменчивости, скрещивания и наследования происходят в рамках целой популяции организмов. Популяция – структурная единица эволюционного процесса. Разработано представление о популяционных волнах – колебаниях численности популяций, регулярно совершаемых при саморегуляции в пищевых цепях, в связи с лунными (месячными), сезонными (годичными), солнечными (11-летними) и другими циклами, в результате природных или техногенных катаклизмов – пожаров, засух, наводнений, оледенений и т.п. Учение о популяционных волнах способствовало пониманию явлений дрейфа генов , т.е. резкого изменения частоты определенных генов в критически малых популяциях. Понятие дрейфа генов, в свою очередь, стало рассматриваться как важный механизм быстрых локальных эволюционных преобразований.
Кроме того, из экологии в эволюционной теории используются знания о причинах и механизмах географических и экологических изоляций . Изоляция – это возникновение разнообразных препятствий к свободному скрещиванию особей внутри популяции, а также между близкими популяциями, которые еще сохраняют генетический обмен на краях своих ареалов. При возникновении изоляции размножение идет в пределах изолятов – обособленных групп, так что между ними прекращается обмен генами. Возникшие ранее изменения генотипа закрепляются в потомстве и усиливаются, что ведет к расхождению признаков особей (дивергенции) в пределах бывшей однородной популяции.
В целом можно заключить, что биологическая эволюция, как и зарождение жизни, – естественный материальный процесс развития Земли, продолжение геологической эволюции. Вполне логично было бы найти в этих глобальных преобразованиях природы общие причины и движущие силы.
Попробуем применить к определению существа биологической эволюции уже испытанный нами тернарный принцип, найти “три кита”, поддерживающих эволюцию живых организмов. Во-первых, это генетика – наследственность видов-основателей, на базе которой идут эволюционные процессы. Данные молекулярной генетики показывают, что у близких видов сходство (гомология) ДНК составляет более 99%. Во-вторых, это эпигенетика – вся совокупность разнообразных факторов среды, направляющих и мутационный процесс (наследственную изменчивость), и естественный отбор. Наконец, в-третьих, синергетика – самоорганизационные процессы. Чтобы найти этот третий компонент в механизмах эволюции, вспомним, что в учении Дарвина важнейшее место занимает понятие дивергенции, означающее расхождение родственных организмов по вновь образуемым признакам. Это непосредственный результат наследственной изменчивости и отбора, который и ведет в итоге к образованию новых видов из вида-предшественника. Дивергенция в популяциях происходит в результате случайных генетических мутаций и рекомбинаций (последние возникают при половом размножении). Совершенно очевидно, что мутации и рекомбинации генов представляют частное биологическое проявление флуктуаций в синергетике, а дарвиновская дивергенция – это типичная бифуркация , за которой следует элементарная “катастрофа” в форме акта самоорганизации качественно новой системы. Как видно, Дарвин предвосхитил современные синергетические понятия механизмов развития, так как процесс видообразования , описанный им, включает типичные элементы самоорганизации, а новый вид и есть качественно новая система.
Подчеркнем, что такой подход не означает сведение биологической эволюции исключительно к процессам самоорганизации, как это пытались делать некоторые авторы. Тем более он не отрицает теорию естественного отбора Дарвина. Напротив, тернарный принцип определения существа эволюции указывает свое место различным движущим факторам – наследственным, самоорганизационным и факторам внешней среды.
Еще раз убеждаемся, что развитие как общая форма движения материи имеет и принципиально общие движущие силы. Триаду “генетика – синергетика – эпигенетика” мы видели в индивидуальном развитии организмов (см. 4.7), в предполагаемых механизмах зарождения жизни на Земле (см. 6.2) и находим в той же смысловой комбинации в процессах глобальной биологической эволюции.
Решение:
Опыт по превращению низкомолекулярных веществ (цианидов, ацетилена, формальдегида и фосфатов) во фрагмент нуклеотида подтверждает гипотезу самопроизвольного синтеза мономеров нуклеиновых кислот из достаточно простых исходных веществ, которые могли быть в условиях ранней Земли.
Опыт, в котором при пропускании электрического разряда через смесь нуклеотидов были получены нуклеиновые кислоты, доказывает возможность синтеза биополимеров из низкомолекулярных соединений в условиях ранней Земли.
Эксперимент, в котором при смешивании в водной среде биополимеров были получены их комплексы, обладающие зачатками свойств современных клеток, подтверждает идею о возможности самопроизвольного образования коацерватов.
6. Установите соответствие между концепцией возникновения жизни и ее содержанием:
2) стационарное состояние
3) креационизм
начало жизни связано с абиогенным образованием органических веществ из неорганических
виды живой материи, как и Земля, никогда не возникали, а существовали вечно
жизнь была создана Творцом в далеком прошлом
жизнь занесена из космоса в виде спор микроорганизмов
Решение:
Согласно концепции биохимической эволюции , начало жизни связано с абиогенным образованием органических веществ из неорганических. Согласно концепции стационарного состояния , виды живой материи, как и Земля, никогда не возникали, а существовали вечно. Сторонники креационизма (от лат. сreatio – сотворение) считают, что жизнь была создана Творцом в далеком прошлом.
7. Установите соответствие между концепцией возникновения жизни и ее содержанием:
1) теория биохимической эволюции
2) стационарное состояние
3) креационизм
возникновение жизни есть результат длительных процессов самоорганизации неживой материи
проблемы зарождения жизни не существует, жизнь была всегда
жизнь есть результат божественного творения
земная жизнь имеет космическое происхождение
Решение:
Согласно концепции биохимической эволюции
, жизнь возникла в результате процессов самоорганизации неживой материи в условиях ранней Земли. Согласно концепции стационарного состояния
, проблемы зарождения жизни не существует, жизнь была всегда. Сторонники креационизма
(от лат. сreatio – сотворение) считают, что жизнь есть результат божественного творения.
Тема 25: Эволюция живых систем
1.Историческая эволюция живых систем (филогенез) является …
самопроизвольной
ненаправленной
обратимой
строго предсказуемой
Решение:
Историческая эволюция живых систем самопроизвольна, она является результатом внутренних возможностей живых систем и действия сил естественного отбора.
2. Синтетическая теория эволюции структурно состоит из теорий микро- и макроэволюций. Теория микроэволюции изучает …
направленные изменения генофондов популяций
основные закономерности развития жизни на Земле в целом
эволюционные преобразования, приводящие к возникновению новых родов
развитие отдельных организмов от рождения до смерти
Решение:
Теория микроэволюции изучает направленные изменения генофондов популяций под воздействием различных факторов. Микроэволюция завершается формированием новых видов организмов, таким образом, она изучает процесс видообразования, но не процесс образования более крупных таксонов.
3. Согласно синтетической теории эволюции, элементарным эволюционным явлением служит изменение …
генофонда популяции
генотипа организма
отдельного гена
хромосомного набора организма
Решение:
Элементарным эволюционным явлением служит изменение генофонда популяции. Отдельная особь претерпевает только онтогенетическое развитие от рождения до смерти и не имеет возможности эволюционировать, поэтому элементарным эволюционным явлением не могут быть изменения отдельных генов, набора генов (генотипов) или набора хромосом отдельного организма.
4. Историческая эволюция живых систем (филогенез) является …
необратимой
ненаправленной
не самопроизвольной
строго предсказуемой
Решение:
Историческая эволюция живых систем является необратимой. Эволюция организмов основывается на вероятностных процессах, в частности – на возникновении случайных мутаций, а потому является необратимой.
5. Эволюционным фактором, благодаря которому эволюция приобретает направленный характер, является(-ются) …
естественный отбор
мутационный процесс
изоляция
популяционные волны
Решение:
Эволюционным фактором, благодаря которому эволюция приобретает направленный характер, является естественный отбор.
Тема 26: История жизни на Земле и методы исследования эволюции (эволюция и развитие живых систем)
1.Морфологические методы исследования эволюции живой природы включают изучение …
недоразвитых и утративших свое основное значение рудиментарных органов, которые могут указать на предковые формы
реликтовых форм, то есть небольших групп организмов с комплексом признаков, характерных для давно вымерших видов
ранних стадий онтогенеза, на которых обнаруживается больше сходства между различными группами организмов
взаимной приспособленности видов друг к другу в природных сообществах
Решение:
Морфологические методы исследования эволюции связаны с изучением особенностей строения органов и организмов сравниваемых форм, а следовательно, изучение недоразвитых и утративших свое основное значение рудиментарных органов, которые могут указать на предковые формы, относится к методам морфологии.
2. Биогеографические методы исследования эволюции живой природы включают …
сопоставление состава фауны и флоры островов с историей их происхождения
изучение рудиментарных органов, указывающих на предковые формы ныне живущих организмов
сравнение ранних стадий онтогенеза организмов разных групп
исследование взаимной приспособленности видов друг к другу в природных сообществах
Решение:
Биогеографические методы исследования эволюции связаны с изучением распространения растений и животных по поверхности нашей планеты, а следовательно, сопоставление состава фауны и флоры островов с историей их происхождения относится к методам биогеографии.
3. Следствием возникновения эукариот в истории жизни на Земле является …
упорядоченность и локализация аппарата наследственности в клетке
возникновение аэробного дыхания
Решение:
Следствием возникновения эукариот в истории жизни на Земле является упорядоченность и локализация аппарата наследственности в клетке. Протоплазма эукариотической клетки сложно дифференцирована, в ней обособлены ядро и другие органоиды. В ядре локализован хромосомный аппарат, в котором сосредоточена основная часть наследственной информации.
4. Экологические методы исследования эволюции живой природы включают изучение …
роли конкретных адаптаций на модельных популяциях
связи между своеобразием флоры, фауны и геологической историей территорий
недоразвитых и утративших свое основное значение рудиментарных органов
процесса онтогенеза организмов данного вида на ранних стадиях
Решение:
Эволюционный процесс является процессом возникновения и развития адаптаций. Экология, изучая условия существования и взаимоотношения между живыми организмами в природных системах или на модельных популяциях, вскрывает значение конкретных адаптаций.
5. Следствием фотосинтеза – важнейшего ароморфоза в истории жизни на Земле – является …
формирование озонового экрана
локализация аппарата наследственности в клетке
дифференциация тканей, органов и их функций
совершенствование анаэробного дыхания
Решение:
Следствием фотосинтеза – важнейшего ароморфоза в истории жизни на Земле – является формирование озонового экрана, который возник по мере накопления кислорода в атмосфере Земли.
6. Расширению арены жизни в истории развития органического мира способствовало …
накопление кислорода в атмосфере
возникновение эукариотов
резкое понижение средней температуры поверхности Земли
затопление наибольшей части континентов водами морей
Решение:
Расширению арены жизни в истории развития органического мира способствовало накопление кислорода в атмосфере с последующим образованием озонового слоя. Озоновый экран защитил от жесткой ультрафиолетовой радиации, в результате чего организмы освоили верхние слои водоемов, более богатые энергией, затем – прибрежные участки, а далее вышли на сушу. В отсутствии озонового экрана жизнь была возможна только под защитой слоя воды толщиной около 10 метров.
7. Ароморфозом, возникшим в ходе эволюции органического мира, является …
возникновение фотосинтеза
появление приспособлений к опылению
изменение окраски цветка
появление защитных игл и колючек
Решение:
Ароморфозы – это такие изменения строения и функций органов, которые имеют общее значение для организма в целом и поднимают уровень его организации. Важнейшим ароморфозом, возникшим в ходе эволюции органического мира, является фотосинтез. Возникновение фотосинтеза привело к целому ряду эволюционных преобразований, как в живых организмах, так и в окружающей среде: появление аэробного дыхания, расширение автотрофного питания, насыщение атмосферы Земли кислородом, появление озонового слоя, заселение организмами суши и воздушной среды.
Тема 27: Генетика и эволюция
1.Установите соответствие между видом изменчивости и ее примером:
1) мутационная изменчивость
пороки развития нервной системы, являющиеся результатом нарушения структуры участка хромосомы
изменение окраски цветка в зависимости от температуры и влажности воздуха
отличающийся от родителей цвет глаз ребенка, являющийся результатом комбинации генов при половом размножении
Решение:
Пороки развития нервной системы, являющиеся результатом нарушения структуры участка хромосомы, – это мутационная изменчивость. Изменение окраски цветка в зависимости от температуры и влажности воздуха представляет модификационную изменчивость.
2. Установите соответствие между генотипами и их проявлением в фенотипе:
два генотипа по одному признаку, одинаково проявляющиеся в фенотипе
два генотипа по одному признаку, по-разному проявляющиеся в фенотипе
два генотипа по двум разным признакам, по-разному проявляющиеся в фенотипе
Решение:
Аллельные гены определяют развитие разных вариантов одного и того же признака, обозначаются одной и той же буквой латинского алфавита – прописной буквой, если ген является доминантным, и строчной, если ген рецессивный. Два генотипа – АА, Аа – одинаково проявляются в фенотипе, поскольку в гетерозиготе Аа проявляется признак доминантного гена. Два генотипа по одному и тому же признаку – АА, аа – по-разному проявляются в фенотипе, поскольку рецессивный ген проявляет себя в гомозиготном состоянии аа.
3. Установите соответствие между свойством генетического материала и проявлением этого свойства:
1) дискретность
2) непрерывность
существует элементарные единицы наследственного материала – гены
жизнь характеризуется продолжительностью существования во времени, которая обеспечивается способностью живых систем к самовоспроизведению
единицы наследственности – гены – расположены в хромосомах в определенной последовательности
Решение:
Дискретность генетического материала проявляется в том, что существуют элементарные единицы наследственного материала – гены. Жизнь как особое явление характеризуется продолжительностью существования во времени, некоторой непрерывностью , которая обеспечивается способностью живых систем к самовоспроизведению – происходит смена поколений клеток, организмов в популяциях, смена видов в системе биоценоза, смена биоценозов, образующих биосферу
4. Установите соответствие между типом признака и его способностью проявляться в поколении:
1) голубой цвет глаз – рецессивный признак
2) карий цвет глаз – доминантный признак
не проявляется в гетерозиготном состоянии
проявляется в гетерозиготном состоянии
не проявляется в гомозиготном состоянии
Решение:
Рецессивный признак проявляются только в гомозиготном состоянии, а в гетерозиготном состоянии рецессивный признак подавляется доминантным и не проявляется. Доминантный признак при полном доминировании проявляется как в гомозиготном, так и в гетерозиготном состоянии.
5. Установите соответствие между свойством генетического материала и проявлением этого свойства:
1) линейность
2) дискретность
гены расположены в хромосомах в определенной последовательности
ген определяет возможность развития отдельного качества данного организма
наследственный материал обладает способностью к самовоспроизведению
Решение:
Линейность генетического материала проявляется в том, что гены расположены в хромосомах в определенной последовательности, а именно в линейном порядке. Ген определяет возможность развития отдельного качества данного организма, что характеризует дискретность его действия.
6. Установите соответствие между понятием и его определением:
1) генотип
2) фенотип
совокупность всех генов диплоидного набора хромосом организма
совокупность всех свойств и признаков конкретного организма
совокупность генов гаплоидного набора хромосом организма
Решение:
Генотип – совокупность всех генов диплоидного набора хромосом организма. Фенотип – совокупность всех свойств и признаков конкретного организма.
7. Установите соответствие между видом изменчивости и ее примером:
1) мутационная изменчивость
2) модификационная изменчивость
изменение структуры хромосом в процессе клеточного деления
изменение окраски цветков при переносе растения из комнатных условий в теплую влажную оранжерею
изменения, связанные с различной комбинацией генов при половом размножении
Решение:
Изменение структуры хромосом в процессе клеточного деления – это мутационная изменчивость. Изменение окраски цветков при переносе растения из комнатных условий в теплую влажную оранжерею представляет модификационную изменчивость.
Тема 28: Экосистемы (многообразие живых организмов - основа организации и устойчивости живых систем)
1.Установите соответствие между функциональной группой организмов экосистемы и примерами организмов:
1) консументы
2) продуценты
3) редуценты
зайцы и волки
зеленые растения и фотосинтезирующие бактерии
гетеротрофные бактерии и грибы
водоросли и почвенные микроорганизмы
Решение:
Консументы – это гетеротрофные организмы, потребляющие органическое вещество продуцентов или других консументов. Консументами являются зайцы и волки. Продуценты – это автотрофные организмы, способные синтезировать органические соединения и строить из них свои тела. К продуцентам относятся зеленые растения, водоросли и фотосинтезирующие бактерии. Редуценты – организмы, живущие за счет мертвого органического вещества, переводя его вновь в неорганические соединения. Редуцентами являются бактерии, грибы.
Главным элементом живого мира, образующим структурную основу всего многообразия органических соединений является Кислород
Гипотеза представляет собой… Теоретическое предположение о сущности непосредственно наблюдаемых явлений.
Главным элементом живого мира,образующим структурную основу всего многообразия органических соединений, является…Углерод.
Главным элементом живого мира,образующим структурную основу всего многообразия органических соединений, является -углерод
Гомеостаз- одно из характерных свойств систем – проявляется в том, что..
СУЩЕСТВУЮТ МОЛЕКУЛЯРНЫЕ МЕХАНИЗМЫ ПОДДЕРЖАНИЯ ОТОЯНСТВА ВНУТРЕННЕЙ СЕДЫ ВОЙ СИСТЕМЫ-
Для измерения галактических и межгалактич. расстояний в качестве единиц длины используют:
световые года
Естественные науки характеризуются… ^ Обязательной опытной проверкой полученных знаний.
Земля отличается от других планет земной группы (Меркурия, Венеры и Марса) ...
большим количеством жидкости на поверхности
Устанавливают сходство организмов на стадии зародышевого развития
Иерархичность природных ситем проявляется в том, что
системы низшего уровнЯ организации входят в состав систем более высокого уровня
Изучая процессы эволюции, в молекулярной биологии Анализируют степень сходства нуклеиновых кислот разных организмов
Историческая эволюция живых систем (филогенез) является… Необратимой
Изучая процессы эволюции, в молекулярной биологии… ^ Анализируют степень сходства нуклеиновых кислот разных организмов.
Интегративность – одно из…Все свойства системы взаимосвязаны и обуславл друг друга
Историческая эволюция живых систем (филогенез) является … самопроизвольной
И в механической и в электромагнитной картинах мира существовали представления о случайности, как отражении неполноты имеющихся знаний; единственно возможном развитии событий при заданном начальном состоянии.
Интегративность – одно из характерных свойств природных систем – проявляется в том, что… У системы появляются новые свойства, отсутствующие у компонентов и возникающие в результате взаимодействия компонентов
^ К деструктивному загрязнению окружающей среды приводит… Строительство автомагистралей.
К явлениям самоорганизации можно отнести:
-переход хим. Реакции в колебательный режим при высоких концентрациях реагентов
К биокосному веществу относится Речная вода
К принципу универсального эволюционизма относится положение о том, что…
Вселенная в целом и все ее подсистемы существуют в постоянном развитии
Кометы, иногда появляющиеся на земном небосводе, … обращаются вокруг Солнца
Космогония изучает происхождение …небесных тел и их систем
К принципам универсального эволюционизма относится положение о том чтоВселенная в целом и все её подсистемы существуют в постоянном развитии
Космология изучает происхождение и развитие ВСЕЛЕННОЙ
К числу статистических научных теорий принадлежит… ^ Квантовая механика.
К числу условий, необходимых для самоорганизации в системе, относится…
СИЛЬНАЯ НЕРАвНОвЕсНОСТЬ СИСТЕМ-
Концепция устойчивого развития человечества означает… Компромисс между стремлением человечества к удовлетворению своих потребностей и необходимостью сохранения биосферы .
Континуальная исследовательская программа Аристотеля была основана на идеях о том, что… ! ^ Пустого пространства не существует, материя заполняет все без пустот ; Материя непрерывна и бесконечно делима ;
Космология изучает происхождение и развитие Вселенной
Какие бы формы жизни ни были обнаружены (или не обнаружены) экспедицией в других мирах, это не может стать опровержением концепции происхождения жизни на Земле, известной как… концепция креационизма ;теория биохимической эволюции
К числу условий, необходимых для самоорганизации в системе, относятся… ^ Сильная неравновесность системы
Линейность генетич материала проявляется в том, что
…
ЕДИНИЦЫ НАСЛЕДСТВЕННОСТИ-ГЕНЫ-РАСПОЛОЖ В ХРОМАСОМАХ В ОПРЕД ПОСЛЕДОВАТ.-; ЕДИНСТВЕННО ВОЗМОЖНОМ РАЗВИТИИ СОБЫТИЙ ПРИ ЗАДАННОМ НАЧАЛЬНОМ СОСТОЯНИИ-
Разные иерархические уровни одной системы-Вселенной
Молекула представляет собой
-структурную еденицу вещества
Мегамир, макромир, микромир – это…РАЗНЫЕ ИЕРАРХИЧЕСКИЕ УРОВНИ ОДНОЙ СИСТЕМЫ – ВСЕЛЕННАЯ
Мегамир, макромир, микромир- это
Молекулярная формула глюкозы (С6Н12О6) и любого другого вещества указывает на…вещества.
КАЧЕСТВЕННЫЙ И КОЛИЧЕСТВЕННЫЙ СОСТАВ
Мегамир, макромир, микромир – это… Разные иерархические уровни одной системы - Вселенной
Между объектами мегамира преобладает _ взаимодействие Гравитационное
Масса обычного вещества, доступного непосредственным наблюдениям в телескопы и сосредоточенного в основном в звездах, составляет_____________ от полной материи вселенной менее 5%
Не относится к числу известных в физике и вообще в естественных науках форм энергии… Положительная и отрицательная психоэнергия.
Наша галактика является… Спиральной
Наше Солнце в настоящее время находится на стадии …преобладания термоядерных реакций синтеза ядер гелия из ядер водорода
Ньютон ввел понятие «Абсолютное время». Время является Абсолютным, поскольку …
Его течение совершенно не зависит от материальных тел и того, что с ним происходит.
Не относится к числу известных в физике и вообще в естественных науках форм энергии… ^ Положительная и отрицательная психоэнергия .
Наша галактика является -Спиральной
Ньютон ввел понЯтие «Абсолютное простраство». Пространство является Абсолютным,поскольку его свойства не зависят от материальных тел и того, что с ними происходит
Ньютон ввел понятие "Абсолютное время". Время является Абсолютным, поскольку...
его течение совершенно не зависит от материальных тел и того, что с ними происходит
Никакие научные данные, полученные в ходе поисков жизни в других мирах, не могут опровергнуть концепцию происхождения жизни, известную как креационизм, поскольку она…неопровержима в принципе; не может быть предметом научного рассмотрения
Научная космология начала развиваться в… Эпоху Возрождения на основе гелиоцентрической системы Коперника
Никакие научные данные, полученные в ходе поисков жизни в других мирах, не могут подтвердить концепцию происхождения жизни на Земле, известную как…
Концепция стационарного состояния
Гипотеза постоянного самозарождения
Ньютон приписывал введённому им понятию «Абсолютного времени» свойство Независимости от материальных тел
Ньютон ввел понятие «Абсолютное время». Время является абсолютным, поскольку…
ЕГО ТЕЧЕНИЕ СОВЕРШЕННО НЕ ЗАВИСИТ ОТ МАТЕРИАЛЬНЫХ ТЕЛ И ТОГО, ЧТО С НИМ ПРОИСХОДИТ-
Наблюдение – получении сведений об агрегатном состоянии, цвете, запахе веществ, выделяющихся в химической реакции.
Ньютон не приписывал введенного им понятию «Абсолютного времени» свойства…зависеть от движения наблюдателя
Научная космология начала развиваться в ^ XX веке на основе общей теории относительности .
Ньютон приписывал введенному им понятию «Абсолютного времени» свойство…
Независимо от материальных тел
Никакие научные данные, полученные в ходе поисков жизни в других мирах, не могут подтвердить концепцию происхождения жизни на Земле. Известную как концепция стационарного состояния, гипотеза постоянного самозарождения.
Ньютон ввел понятие абсолютное пространство. Пространство является Абсолютным, поскольку…
Его свойства не зависят от материальных тел и того, что ними происходи��
Общая теория относительности , в отличие от специальной теории относительности, применима … в очень сильных гравитационных полях
Одним из эмпирических доказательств справедливости общей теории относительности служит -
Одним из индикаторов глобального экологического кризиса современности является
антропогенное изменение состава атмосферы
Относительно симметрий пространства справедливо утверждение, что пространство...
изотропно
массивные материальные тела своим присутствием и движением искривляют геометрию пространства -времени
Аномальное смещение перигелия орбиты Меркурия
Относительно симметрий пространства справедливо утверждение, что пространство… Изотропно
Одним из эмпирических доказательств справедливости общей теории относительности служит … Аномальное смещения перигелия орбиты меркурия
Одним из индикаторов глобального экологического кризиса современности является…
деградация водных, земельных и лесных ресурсов
Одной из причин химической активности органических молекул явл-ся наличие в них функциональных групп, которые являются результатом способности углерода…
образовывать активные связи с атомами кислорода, азота, серы
образовывать четыре связи с другими атомами
Одним из эмпирических доказательств справедливости общей теории относительности служит… Аномальное смещение перигелия орбиты Меркурия
Одним из критериев классификации элементарных частиц является… Время жизни
Одним из следствий принципа эквивалентности выводимым в общей теории относительности, является то, что…Масса и энергия эквивалентны, они связаны соотношением Эйнштейн��
Одним из следствий принципа эквивалентности, выводимым в общей теории относительности, является то, что...в сильных гравитационных полях темп времени замедляется
Основную информацию о составе и динамике глубоких недр Земли приносит...анализ распространения сейсмических волн
Отличие популярного в ХIХ веке понятия мирового эфира от ньютоновского понятия Абсолютного пространства состояло в том, что эфир предполагался...взаимодействующим с материальными телами
Опыт Майкельсона-Морли по измерению скорости света относительно движущегося наблюдателя, проведенный многократно в конце ХIX века, показал … полное соответствие концепции мирового эфира реальности нашего мира
Одним из экологических последствий неолитической революции, являются техногенное загрязнение среды
О расширении вселенной говорит наблюдательный факт …
-КРАСНОГО СМЕЩЕНИЯ В СПЕКТРАХ ВСЕХ ДАЛЕКИХ ГАЛАКТИК-
Одним из экологических последствий неолитической революции(10-8 до н.э.) является
ОПУСТЫНИВАНИЕ ТЕРРИТОРИЙ-
Одним из эмпирических доказательств справедливости общей теории относитетельности служит
-отклонение световых лучей, проходящих вблизи солнца
О присутствии и функционировании живых организмов сигнализирует наличие…Плотной атмосферы с развитой циркуляцие
-Достаточно больших открытых водоемов
Объект,занимающий основную часть поля на данной фотографии не может быть нашей Галактикой(Млечный путь), поскольку мы находимся внутри нашей Галактики, а снимок явно сделан извне
Объект, занимающий основную часть поля зрения, на данной фотографии, не может быть нашей Галактикой (Млечный путь) поскольку Наша Галактика-эллиптическая, а на фото приведена спиральная
Одним из критериев классификации элементарных частиц является спин
Одним из эмпирических доказательств справедливости общей теории относительности служит… Аномальное смещение перигелия орбиты Меркурия
О расширении Вселенной говорит наблюдательный факт… ^ Красного смещения в спектрах всех далеких галактик
^ Одним из следствий принципа эквивалентности, выводимым в общей теории относительности, является то, что…массивные материальные тела своим присутствием и движением искривляют геометрию пространства-времени
Парой рецессивных генов, определяющих развитие одного признака, будет набор… аа
Против концепции происхождения жизни, известной как гипотеза панспермии, могло бы свидетельствовать открытие… того факта, что в космосе жизни нет нигде, кроме Земли; живых организмов земного типа в планетной системе Сириуса и жизни с совершенно иной химической основой на Марсе.
Парой аллельных генов являются гены, определяющие…карий и голубой цвет глаз
Псевдонаука отличается от истинной науке Невосприимчивостью к критике и восхваление своих результатов
Постулатом который был введен в общей теории относительности, является положение о Неизменности законов природы в разных инерциальных системах отсчета
Примером мутационной изменчивости является изменение Структуры хромосомы в процессе клеточного деления
Псевдонаука отличается от истинной науки…Невосприимчивостью к критике и восхваление своих результат��
По принципу дополнительности в его широком смысле связаны друг с другом…методы дедукции и индукции в научном познании
По принципу дополнительности в его широком смысле связаны друг с другом…Квантовая и классическая механик��
Представление о том , что Земля занимает рядовое, ничем не примечательное положение во Вселенной, характерно для .
Представление о том
, что любое изменение в мире можно свести к перемещениям некоторых корпускул, характерно для … атомистических учений Левкиппа и Демокрита
Представление о пространстве-времени как полноправной, активной, сложно устроенной составляющей материального мира характерно для: современной научной картины мира
Проведение многократных экспериментов для установления истинности выдвинутого предположения является примером применения в науке принципа… Верификация
Предсказания специальной теории относительности и классической механики совпадает при условии, что… Скорости движения гораздо меньше скорости света
По своим размерам Земля занимает_______место среди восьми планет Солнечной системы. пятое
Против концепции происхождения жизни, известной как гипотеза панспермии, могло бы свидетельствовать открытие…(1 из 2)
Живых организмов земного типа на Марсе и жизни с совершенно иной химической основой в планетной системе Сириуса
^ Того факта, что в космосе жизни нет нигде, кроме Земли
Понимание пространства и времени как некоторых субстанции, обладающих самостоятельным существованием и не связанных с материальными телами, характерно для… натурфилософской картины мира Аристотеля
Примером модификационной изменчивости является изменение -окраски крыльев бабочек под влиянием условий внешней среды
Примером мутационной изменчивости является изменение… Структуры хромосомы в процессе клеточного деления.
Примером модификационной изменчивости является изменение…
Окраски крыльев бабочек под влиянием условий внешней среды
Представление о пространстве-времени как полноправной, активной, сложно устроенной составляющей материального мира характерно для…
Современной научной картины мира
Предсказания специальной теории относительности и классической механики совпадают при условии,что…^Скорости движения гораздо меньше скорости света.
Постулатом,который был введен в общей теории относительности,является положение о (об)...Эквивалентности ускоренного движения и покоя в гравитационном поле.
Понимание пространства и времени как всего лишь системы отношений между материальными телами характерно для… учений древнегреческих атомистов
Продуцентами в наземной экосистеме являются..растения
Парой рецессивных генов, определяющих развитие одного признака, будет набор...аа
По современным представлениям, примерно через 5 миллиардов лет Солнце исчерпает основные запасы своего термоядерного горючего и … превратится в белый карлик
Представление о пространстве - времени как полноправной, активной, сложно устроенной составляющей материального мира характерно для...современной научной картины мира
Проведение многократных экспериментов для установления истинности выдвинутого предположения является примером применения в науки принципа
Верификации
Парой рецессивных генов, определяющих развитие одного признака, будет набор… аа .
Продуцентами в наземной экосистеме являются… Растения.
Понимание пространства и времени как некоторых субстанций, обладающих самостоятельным существованием и не связанных с материальными телами, характерно для… Механической научной картины мира .
Предположение о наличии у света квантовых (корпускулярных) свойств оказалось необходимым для объяснения экспериментально установленных законов...ФОТОЭФФЕКТА
Распад свободного нейтрона на электрон и антипротон запрещен законом...СОХРАНЕНИЯ ЭЛЕКТРИЧЕСКОГО ЗАРЯДА
Расширение Вселенной происходит… Однородно, без какого-либо особого или выделенного центра расширения.
Расширению арены жизни в истории развития органического мира способствовало … накопление кислорода в атмосфере
Роль для живой природы аномальной плотности льда - твёрдого состояния воды - проявляется в том, что возможно...сохранение жизни в замерзающих водоёмах
Результатом процесса дифференциации научного знания является…Биохимия
Результатом дифференциации научного знания является… Эмбриология
Результатом процесса дифференциации
научного знания является … квантовая механика
С
С точки зрения термодинамики
, предприятия электроэнергетики … превращают неудобные для использования формы энергии частично в электроэнергию, частично в низкокачественные формы энергии
Синергетика изучает… Закономерности самоорганизации в природных и социальных системах.
Синтетическая теория эволюции структурно состоит из теорий микро- и макроэволюций. Теория микроэволюции изучает … направленные изменения генофондов популяций
Согласно синтетической теории эволюции, элементарным эволюционным явлением служит изменение … генофонда популяции
Следствием возникновения эукариот в истории жизни на Земле является.... упорядоченность и локализация аппарата наследственности в клетке
Следствием фотосинтеза – важнейшего ароморфоза в истории жизни на Земле – является … формирование озонового экрана
Суть открытия, сделанного американским астрономом-наблюдателем Э. Хабблом в 20-х годах XX века и ставшего эмпирической основой для становления научной космологии, заключалась в том, что галактики … удаляются друг от друга тем быстрее, чем больше расстояние между ними
Согласно современным космологическим представлениям, расширение Вселенной … ускоряется и будет происходить, чем дальше, тем быстрее
^ Согласно одной из исторических концепций происхождение жизни – гипотезе панспермии, жизнь… Занесена на Землю из космоса.
Согласно представлениям механической картины мира, если бы из Вселенной можно было убрать все материальные тела, то … Свойства пространства и времени остались бы неизменными
Согласно современным концепциям биологической эволюции, первые живые организмы были анаэробными, то есть…
Согласно теории относительности , при переходе от одной системы отсчета к другой может измениться тот факт, что … свет в половине Калифорнии погас в тот момент, когда Мария Ивановна Петрова в Москве щелкнула выключателем в своей квартире
Согласно специальной теории относительности, при переходе от одной системы отчета к другой может измениться… Промежуток времени между двумя событиями
Согласно соотношению неопределенностей В. Гейзенберга…
Невозможно точно и одновременно измерить и координату, и импульс материального объекта
Согласно одной из формулировок второго закона термодинамики, …
Невозможно тепловую энергию полностью перевести в полезную работу
Синергетика изучает …
Структурами макромира является…Молекула воды
Согласно соотношению неопределенности Гейзенберга, точное значение импульса конкретного объекта…Делает неточным знание его координа��
Создание макета газотурбинной установки является примером использования метода...моделирования
Согласно соотношению неопределённостей В. Гейзенберга, точное знание импульса конкретного объекта...делает неточным знание его координат
занесена на Землю из космоса
Способность атомов углерода связываться с атомами азота, фосфора с образованием... существование функциональных групп. обуславливающих...
Солнечная система устроена так, что...ПЛАНЕТЫ-ГИГАНТЫ В СРЕДНЕМ В ДЕСЯТКИ РАЗ ДАЛЬШЕ ОТ СОЛНЦА ЧЕМ ПЛАНЕТЫ ЗЕМНОЙ ГРУППЫ
Способность атомов углерода связываться друг с другом и другими атомами...образование структуры основы...
Способность атома углерода образовывать органические соединения...возможность образования высокомолекулярных(ного)...
Сходство между Большим Взрывом (процессом, в ходе которого образовалась и приобрела свои свойства наша Вселенная) и обычным взрывом артиллерийского снаряда состоит в том, что..расстояния между галактиками с течением времени увеличиваются подобно тому, как разлетаются в разные стороны осколки взорвавшегося снаряда
Среди участников дискуссии о движении мнение о том, что движение продолжается, пока действует вызвавшая его сила, разделили....Аристотель ; Гераклит
Согласно одной из формулировок второго закона термодинамики, ...невозможно тепловую энергию полностью перевести в полезную работу
Согласно представлениям о пространстве и времени, свойственным механической картине мира, если бы из Вселенной исчезли все материальные тела, то пространство и время..продолжали бы существовать и их свойства ничуть не изменились бы
Согласно представлениям о пространстве и времени, свойственным механической картине мира, если бы из Вселенной исчезли все материальные тела, то пространство и время… Продолжали бы существовать и их свойства ничуть не изменились бы
Согласно современным космологическим представлениям, расширение Вселенной… ускоряется и будет происходить чем дальше , тем быстрее
Согласно концепции корпускулярно-волнового дуализма, любой материальный объект… обладает свойствами как волны, так и частицы, но в конкретном эксперименте проявляет либо те, либо другие
Соответствие между методом научного познания и примером его использования:
Моделирование
– написание структурных формул биоорганических соединений;
Эксперимент
– многократное проведение синтеза органических молекул, свойственных живому, из неорганических газов;
Измерения толщины слоя осадочных пород, накопившихся за историю Земли
Согласно современным концепциям биологической эволюции, первые живые организмы были анаэробами, то есть…Были способны жить в отсутствии кислорода
Синтетическая теория эволюции структурно состоит из теории микроэволюции и макроэволюции. Теория макроэволюции изучает…Эволюционные преобразования за длительный исторический период на уровнях выше видового
Состав простого вещества отображает формула… O3
Структурами макромира являются…Бактерии.
Синергетика изучает… Закономерности самоорганизации в природных и социальных системах .
Самая точная оценка возраста Земли получена путем… Измерения концентрации радиоактивных изотопов и продуктов их распада в земных породах и метеоритах.
Согласно современным концепциям биологической эволюции, первые живые организмы были гетеротрофами, то есть… ^ Питались готовыми органическими веществами .
Состав простого вещества отображает формулу… О 3
Создание макета газотурбинной установки является примером использования метода…
Моделирования
Суть открытия, сделанного американским астрономом-наблюдателем Э. Хабблом в 20-х 20 века и ставшего эмпирической основой для становления научной космологии, заключалась в том, что галактики… удаляются друг от друга тем быстрее, чем больше расстояние между ними
Системность научного знания отражается в Четкой структуре знания и методов познания
Составной элементарной частицей является -протон
Роль для живой природы аномальной плотности льда-твердого состояния воды-проявляется в том, что возможно…Сохранение жизни в замерзающих водоемов
Согласно концепции механического детерминизма -любое будущее событие можно точно предсказать, зная начальное состояние и используЯ законы механики
Самой низкой энтропией при равных колличествах обладает(-ют) вода в виде льда
Состояние системы,приближающейся к точке бифуракции,характеризуется неустойчивостью
Самая точная оценка возраста Земли получена путем -измерении концентрации радиоактивных изотопов и продуктов их распада в земных породахи метиоритах
Согласно одной из исторических концепций происхождения жизни-концепции креационизма -жизнь есть результат божественного творения
Синтетическая теория эволюции структурно состоит из теорий микро- и макроэволюций.
Свойство, которое отражает процесс получения научного знания путём логических доказательств и проверки экспериментом, называется...достоверностью
Структурами микромира являются...молекулы воды
Состав простого вещества отображает формула...O 3
Согласно одной из формулировок второго закона термодинамики, в ходе процесса, протекающего в изолированной системе, ...энтропия возрастает
Состояние системы, приближающейся к точке бифуркации, характеризуется...
неустойчивостью
Согласно одной из исторических концепций происхождения жизни - гипотезе панспермии, жизнь...занесена на Землю из космоса
Согласно современ концепц биологич эволюции, первые жив организмы были аноэробами т.е.?
-БЫЛИ СПОСОБНЫ ЖИТЬ В ОТСУТСТВИИ КИСЛОРОДА-
Согласно синтетической теории эволюции и теории Ч.дарвина, главным движущим факторром эволюц является…
ЕСТЕСТВЕННЫЙ ОТБОР-
Согласно современной космологии, возраст вселенной-в несколько раз больше возраста солнца и земли
Системность научного познания отражается в Четкой структуре знания методов познани







